Криптомонада
| Криптомонады | |
|---|---|
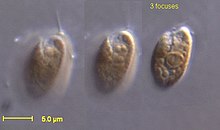 | |
| Родомонас Салина | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | потогонные средства |
| Подтип: | Ролломонадия |
| Суперкласс: | Криптомонада Кавальер-Смит , стат 2004 г. ноябрь 2015 год [1] |
| Занятия и заказы [1] | |
Криптомонады криптофиты или ( ) [2] это группа водорослей , [3] большинство из которых имеют пластиды . они традиционно считаются водорослей под . В среде психологов названием Cryptophyta подразделением [4] Они распространены в пресной воде, а также встречаются в морских и солоноватых средах обитания. Каждая клетка имеет размер около 10–50 мкм , уплощенную форму, с передней бороздкой или карманом. По краю кармана обычно имеются два слегка неравных жгутика . Некоторые могут проявлять миксотрофию . [5] Их относят к кладе Cryptomonada , которая делится на два класса: гетеротрофные Goniomonadea и фототрофные Cryptophyceae . Эти две группы объединены тремя общими морфологическими характеристиками: наличием перипласта , эжектомом со вторичной закруткой и митохондриальных крист с плоскими канальцами. [6] Генетические исследования, проведенные еще в 1994 году, также подтвердили гипотезу о том, что Goniomonas была сестрой Cryptophyceae. [7] Исследование, проведенное в 2018 году, выявило убедительные доказательства того, что общий предок Cryptomonada был автотрофным протистом. [8]
Характеристики
[ редактировать ]
Криптомонады отличаются наличием характерных экструсом, называемых эжектосомами , которые состоят из двух соединенных спиральных лент, удерживаемых под натяжением. [9] Если клетки раздражаются механическим, химическим или световым стрессом, они разряжаются, перемещая клетку зигзагообразным курсом в сторону от возмущения. С карманом связаны крупные эектосомы, видимые под световым микроскопом; более мелкие встречаются под перипластом , окружением клеток, специфичных для криптофитов. [10] [11]
За исключением класса Goniomonadea , у которого полностью отсутствуют пластиды. [12] и Cryptomonas paramecium (ранее называвшаяся Chilomonas paramecium ), имеющая лейкопласты , криптомонады имеют один или два хлоропласта. Они содержат хлорофиллы а и с вместе с фикобилипротеинами и другими пигментами и различаются по цвету (от коричневого, красного до синевато-зеленого). Каждая окружена четырьмя мембранами, а находится редуцированное клеточное ядро, называемое нуклеоморфом между двумя средними . Это указывает на то, что пластида произошла от эукариотического симбионта, который, как показали генетические исследования, представлял собой красную водоросль . [13] Однако пластиды сильно отличаются от пластид красных водорослей: фикобилипротеины присутствуют, но только в просвете тилакоида и присутствуют только в виде фикоэритрина или фикоцианина . В случае Rhodomonas кристаллическая структура была определена до 1,63 Å; [14] и было показано, что альфа-субъединица не имеет никакого отношения ни к одному другому известному фикобилипротеину.
Некоторые криптомонады, такие как Cryptomonas , могут образовывать пальмеллоидные стадии, но легко покидают окружающую слизь и снова становятся свободноживущими жгутиконосцами. Некоторые виды Cryptomonas также могут образовывать неподвижные микробные цисты — стадии покоя с жесткими клеточными стенками, позволяющие выжить в неблагоприятных условиях. Жгутики криптомонад вставлены параллельно друг другу и покрыты двудольными волосками, называемыми мастигонемами , которые образуются внутри эндоплазматической сети и транспортируются на поверхность клетки. Небольшие чешуйки также могут присутствовать на жгутиках и теле клетки. Митохондрии имеют плоские кристы , митоз открытый; половом размножении Сообщалось также о .
Классификация
[ редактировать ]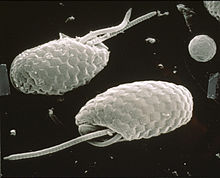

Первое упоминание о криптомонадах, по-видимому, было сделано Кристианом Готфридом Эренбергом в 1831 году. [15] во время изучения Инфузории . Позднее ботаники отнесли их к отдельной группе водорослей , классу Cryptophyceae или отделу Cryptophyta, а зоологи отнесли к жгутиковым простейшим отряда Cryptomonadina. В некоторых классификациях криптомонады считались близкими родственниками динофлагеллят из -за их (казалось бы) схожей пигментации и группировались как Pyrrophyta . Хлоропласты криптомонад тесно связаны с хлоропластами гетероконтов и гаптофитов , и Кавальер-Смит объединил эти три группы как Chromista . Однако тот факт, что сами организмы тесно связаны, противоречит основным различиям в клеточной организации ( ультраструктурная идентичность ), что позволяет предположить, что три основные линии, отнесенные к хромистам, приобрели пластиды независимо и что хромисты полифилетичны. Представление о том, что криптомонады примитивно гетеротрофны и имеют вторично приобретенные хлоропласты, подтверждается молекулярными данными. [16] Парфри и др. и Бурки и др. поместил Cryptophyceae в родственную кладу зеленых водорослей . [17] или зеленые водоросли плюс глаукофиты . [18] Сестринской группой криптомонад, вероятно, являются катаблефариды (также называемые катаблефаридами), группа жгутиконосцев, которые также имеют эжектомосомы. [19]
Одна из предложенных группировок выглядит следующим образом: (1) Cryptomonas , (2) Chroomonas / Komma и Hemiselmis , (3) Rhodomonas / Rhinomonas / Storeatula , (4) Guillardia / Hanusia , (5) Geminigera / Plagioselmis / Teleaulax , (6) Proteomonas sulcata , (7) Falcomonas daucoides . [20]
Ссылки
[ редактировать ]- ^ Jump up to: а б Кавальер-Смит, Томас (2018). «Королевство Chromista и его восемь типов: новый синтез, подчеркивающий нацеливание на перипластидные белки, эволюцию цитоскелета и перипластид, а также древние расхождения» . Протоплазма . 255 : 297–357. дои : 10.1007/S00709-017-1147-3 . ПМЦ 5756292 . ПМИД 28875267 .
- ^ Барнс, Ричард Стивен Кент (2001). Беспозвоночные: синтез . Уайли-Блэквелл. п. 41. ISBN 978-0-632-04761-1 .
- ^ Хан Х, Арчибальд Дж. М. (май 2008 г.). «Латеральный перенос интронов в пластидном геноме криптофитов» . Нуклеиновые кислоты Рез . 36 (9): 3043–53. дои : 10.1093/нар/gkn095 . ПМК 2396441 . ПМИД 18397952 .
- ^ Окамото, Н.; Иноуе, И. (2005). «Катаблефариды являются дальней сестринской группой Cryptophyta: предложение для Katablepharidophyta Divisio Nova/Kathablepharida Phylum Novum на основе рДНК SSU и филогении бета-тубулина». Протист . 156 (2): 163–179. дои : 10.1016/j.protis.2004.12.003 . ПМИД 16171184 .
- ^ «Криптофиты – криптомонады» . Архивировано из оригинала 10 июня 2011 г. Проверено 2 июня 2009 г.
- ^ Кавальер-Смит, Томас (2004). Органеллы, геномы и филогения эукариот . стр. 87–88.
- ^ Макфадден, Гилсон и Хилл (1994), «Гониомонады: последовательности рРНК указывают на то, что этот фаготрофный жгутиконосец является близким родственником компонента-хозяина криптомонад», European Journal of Phycology , 29 (1): 29–32, doi : 10.1080/ 09670269400650451
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ченчи (2018), «Последовательность ядерного генома криптомонады без пластид Goniomonas avonlea дает представление об эволюции вторичных пластид», BMC Biology , 16 (1): 137, doi : 10.1186/s12915-018-0593-5 , PMC 6260743 , PMID 30482201
- ^ Грэм, Ле; Грэм, Дж. М.; Уилкокс, Л.В. (2009). Водоросли (2-е изд.). Сан-Франциско, Калифорния: Бенджамин Каммингс (Пирсон). ISBN 9780321559654 .
- ^ Морролл, С.; Гринвуд, AD (1980). «Сравнение периодических подструктур трихоцист Cryptophyceae и Prasinophyceae». БиоСистемы . 12 (1–2): 71–83. дои : 10.1016/0303-2647(80)90039-8 . ПМИД 6155157 .
- ^ Грим, Дж. Н.; Стэхелин, Луизиана (1984). «Эжектомомы жгутиковых Chilomonas paramecium - визуализация с помощью методов замораживания-перелома и выделения». Журнал протозоологии . 31 (2): 259–267. дои : 10.1111/j.1550-7408.1984.tb02957.x . ПМИД 6470985 .
- ^ Последовательность ядерного генома криптомонады без пластид Goniomonas avonlea дает представление об эволюции вторичных пластид.
- ^ Дуглас, С.; и др. (2002). «Сильно редуцированный геном порабощенного ядра водоросли» . Природа . 410 (6832): 1091–1096. Бибкод : 2001Natur.410.1091D . дои : 10.1038/35074092 . ПМИД 11323671 .
- ^ Уилк, К.; и др. (1999). «Эволюция светособирающего белка путем добавления новых субъединиц и перестановки консервативных элементов: кристаллическая структура криптофитного фикоэритрина с разрешением 1,63 Å» . ПНАС . 96 (16): 8901–8906. дои : 10.1073/pnas.96.16.8901 . ПМК 17705 . ПМИД 10430868 .
- ^ Новарино, Г. (2012). «Таксономия криптомонад в 21 веке: первые 200 лет» . Психологические отчеты: современные достижения в таксономии водорослей и ее применениях: филогенетическая, экологическая и прикладная перспектива : 19–52 . Проверено 16 октября 2018 г.
- ^ Ченчи, У.; Сиббальд, С.Дж.; Кертис, бакалавр; Камикава, Р.; Эме, Л.; Муг, Д.; Генриссат, Б.; Марешаль, Э.; Чаби, М.; Джемиэль, К.; Роджер, Эй Джей; Ким, Э.; Арчибальд, Дж. М. (2018). «Последовательность ядерного генома криптомонады Goniomonas avonlea, лишенной пластид, дает представление об эволюции вторичных пластид» . БМК Биология . 16 (1): 137. дои : 10.1186/s12915-018-0593-5 . ПМК 6260743 . ПМИД 30482201 .
- ^ Парфри, Лаура Вегенер ; Лар, Дэниел Дж.Г.; Нолл, Эндрю Х.; Кац, Лаура А. (16 августа 2011 г.). «Оценка времени ранней диверсификации эукариот с помощью мультигенных молекулярных часов» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (33): 13624–13629. Бибкод : 2011PNAS..10813624P . дои : 10.1073/pnas.1110633108 . ПМК 3158185 . ПМИД 21810989 .
- ^ Бурки, Фабьен; Каплан, Майя; Тихоненков Денис Владимирович; Златогурский, Василий; Минь, Буй Куанг; Радайкина Людмила Владимировна; Смирнов, Алексей; Мыльников Александр П.; Килинг, Патрик Дж. (27 января 2016 г.). «Распутывание ранней диверсификации эукариот: филогеномное исследование эволюционного происхождения Centrohelida, Haptophyta и Cryptista» . Учеб. Р. Сок. Б. 283 (1823): 20152802. doi : 10.1098/rspb.2015.2802 . ISSN 0962-8452 . ПМК 4795036 . ПМИД 26817772 .
- ^ Нисимура, Юки; Куме, Кейтаро; Сонехара, Кейто; Танифудзи, Горо; Ширатори, Такаши; Исида, Кен-Ичиро; Хашимото, Тецуо; Инагаки, Юдзи; Окума, Мория (2020). «Митохондриальные геномы Hemiarma marina и Leucocryptos marina пересмотрели эволюцию созревания цитохрома c у Cryptista» . Границы экологии и эволюции . 8 . дои : 10.3389/fevo.2020.00140 .
- ^ «Криптомонады» . Проверено 24 июня 2009 г.