лаковый оперон
Лактозный у оперон ( lac -оперон) — это оперон, E. coli и необходимый для транспорта и метаболизма лактозы многих других кишечных бактерий . Хотя глюкоза является предпочтительным источником углерода для большинства кишечных бактерий, lac- оперон позволяет эффективно переваривать лактозу, когда глюкоза недоступна, за счет активности β-галактозидазы . [1] Генная регуляция оперона lac- была первым механизмом генетической регуляции, который был четко понят, поэтому он стал ярким примером прокариот регуляции генов . По этой причине его часто обсуждают на вводных занятиях по молекулярной и клеточной биологии . Эту систему метаболизма лактозы использовали Франсуа Жакоб и Жак Моно, чтобы определить, как биологическая клетка узнает, какой фермент синтезировать. Работа над лаковым опероном принесла им Нобелевскую премию по физиологии в 1965 году. [1]
У большинства бактериальных клеток, включая E. coli отсутствуют интроны , в геноме . У них также отсутствует ядерная мембрана . Следовательно, регуляция генов с помощью lac-оперона происходит на уровне транскрипции, предотвращая превращение ДНК в мРНК .
Бактериальные опероны представляют собой полицистронные транскрипты , которые способны продуцировать несколько белков из одного транскрипта мРНК . В этом случае, когда лактоза требуется в качестве источника сахара для бактерии, три гена lac- могут экспрессироваться оперона и транслироваться их последующие белки: lacZ , lacY и lacA . Генным продуктом lacZ является β-галактозидаза , которая расщепляет лактозу, дисахарид , на глюкозу и галактозу . lacY кодирует β-галактозидпермеазу , мембранный белок , который внедряется в плазматическую мембрану , обеспечивая клеточный транспорт лактозы в клетку. Наконец, lacA кодирует β-галактозидтрансацетилазу .


1 : РНК-полимераза, 2 : репрессор, 3 : промотор, 4 : оператор, 5 : лактоза, 6 : lacZ , 7 : lacY , 8 : lacA .
Обратите внимание, что количество пар оснований на диаграмме, приведенной выше, указано не для масштаба. На самом деле в lac-опероне более 5300 пар оснований. [2]
Было бы расточительно производить ферменты при отсутствии лактозы или при наличии предпочтительного источника энергии, такого как глюкоза. Лак - оперон использует двухчастный механизм управления, гарантирующий, что клетка расходует энергию на производство ферментов, кодируемых lac- опероном, только при необходимости. [3]
В отсутствие лактозы lac -репрессор , кодируемый lacI, останавливает выработку ферментов и транспортных белков, кодируемых lac- опероном. [4] Это происходит путем блокирования ДНК-зависимой РНК-полимеразы . Эта блокировка/остановка не идеальна, и минимальное количество экспрессии генов происходит постоянно. Белок-репрессор всегда экспрессируется, но lac-оперон (ферменты и транспортные белки) репрессируется. (Но не полностью остановлено)
Когда доступна лактоза, но нет глюкозы, некоторое количество лактозы поступает в клетку с использованием уже существующего транспортного белка, кодируемого lacY. Эта лактоза затем соединяется с репрессором и инактивирует его, тем самым позволяя экспрессировать lac-оперон. больше β-галактозидпермеазы, Затем синтезируется позволяя проникнуть еще большему количеству лактозы, и ферменты, кодируемые lacZ и lacA, могут ее переварить.
Однако в присутствии глюкозы, независимо от присутствия лактозы, оперон будет репрессирован. Это связано с тем, что белок-активатор катаболитов (CAP), необходимый для производства ферментов, остается неактивным, и EIIA Глк закрывает пермеазу лактозы, чтобы предотвратить транспорт лактозы в клетку. Этот механизм двойного контроля вызывает последовательное использование глюкозы и лактозы в двух различных фазах роста, известных как диокси .
Структура
[ редактировать ]
- Лак - оперон состоит из трех структурных генов , а также промотора , терминатора , регулятора и оператора . Три структурных гена: lacZ , lacY и lacA .
- lacZ кодирует β-галактозидазу (LacZ), внутриклеточный фермент , расщепляющий дисахарид лактозу на глюкозу и галактозу .
- lacY кодирует β-галактозидпермеазу (LacY), трансмембранный симпортер , который перекачивает β-галактозиды, включая лактозу, в клетку, используя протонный градиент в том же направлении. Пермеаза увеличивает проницаемость клетки для β-галактозидов .
- lacA кодирует β-галактозидтрансацетилазу (LacA), фермент, который переносит ацетильную группу от ацетил-КоА к тиогалактозиду.
Только lacZ и lacY , по-видимому, необходимы для катаболического пути лактозы .
По цифрам: lacI имеет 1100 п.н., lacZ — 3000 п.н., lacY — 800 п.н., lacA — 800 п.н., причем 3 п.н. соответствуют 1 аминокислоте. [5]
Генетическая номенклатура
[ редактировать ]Трехбуквенные сокращения используются для описания фенотипов бактерий, включая E. coli .
Примеры включают в себя:
- Lac (способность использовать лактозу),
- Его (способность синтезировать аминокислоту гистидин)
- Мот (плавательная моторика)
- см Р (резистентность к антибиотику стрептомицину )
В случае Lac клетки дикого типа представляют собой Lac. + и способны использовать лактозу в качестве источника углерода и энергии, тогда как Lac − мутантные производные не могут использовать лактозу. Те же три буквы обычно используются (строчные, курсивом) для обозначения генов, участвующих в определенном фенотипе, причем каждый отдельный ген дополнительно обозначается дополнительной буквой. Гены lac , кодирующие ферменты, — это lacZ , lacY и lacA . Четвертый lac ген — lacI , кодирующий репрессор лактозы — «I» означает индуцируемость .
Можно различать структурные гены, кодирующие ферменты, и регуляторные гены, кодирующие белки, влияющие на экспрессию генов. Текущее использование расширяет фенотипическую номенклатуру и применяется к белкам: таким образом, LacZ представляет собой белковый продукт гена lacZ , β-галактозидазу. Различные короткие последовательности, которые не являются генами, также влияют на экспрессию генов, включая промотор lac , lac p и оператор lac , laco . Хотя это не является строго стандартным использованием, мутации, влияющие на lac o, называются lac o. с , по историческим причинам.
Регулирование
[ редактировать ]Специфический контроль lac -генов зависит от доступности субстрата лактозы для бактерии. Белки не производятся бактериями, когда лактоза недоступна в качестве источника углерода.Гены lac организованы в оперон ; то есть они ориентированы в одном направлении, непосредственно прилегая к хромосоме, и совместно транскрибируются в одну полицистронную молекулу мРНК. Транскрипция всех генов начинается со связывания фермента РНК-полимеразы (РНКП), ДНК-связывающего белка , который связывается со специфическим участком связывания ДНК, промотором , расположенным непосредственно перед генами. Связыванию РНК-полимеразы с промотором способствует цАМФ связанный с белок-активатор катаболита, (CAP, также известный как белок-рецептор цАМФ). [6] Однако ген lacI (регуляторный ген lac -оперона) продуцирует белок, который блокирует связывание РНКП с оператором оперона. Этот белок можно удалить только тогда, когда аллолактоза связывается с ним и инактивирует его. Белок, образуемый геном lacI, известен как lac-репрессор. Тип регуляции, которому подвергается lac- оперон, называется негативным индуцибельным, что означает, что ген выключается регуляторным фактором ( лак -репрессором), если не добавляется какая-либо молекула (лактоза). После удаления репрессора РНКП приступает к транскрипции всех трех генов ( lacZYA ) в мРНК. Каждый из трех генов на цепи мРНК имеет свою собственную последовательность Шайна-Дальгарно , поэтому гены транслируются независимо. [7] Последовательность ДНК E. coli lac- оперона , lacZYA мРНК и генов lacI доступны в GenBank (вид) .
Первым механизмом контроля является регуляторная реакция на лактозу, которая использует внутриклеточный регуляторный белок, называемый репрессором лактозы, для препятствования выработке β-галактозидазы в отсутствие лактозы. Ген lacI , кодирующий репрессор, лежит рядом с опероном lac и всегда экспрессируется ( конститутивен ). Если лактоза отсутствует в ростовой среде, репрессор очень прочно связывается с короткой последовательностью ДНК сразу за промотором, рядом с началом lacZ, называемой оператором lac . Связывание репрессора с оператором препятствует связыванию РНКП с промотором, и поэтому мРНК, кодирующая LacZ и LacY, образуется только на очень низких уровнях. Однако когда клетки выращиваются в присутствии лактозы, метаболит лактозы, называемый аллолактозой, образующийся из лактозы продуктом гена lacZ , связывается с репрессором, вызывая аллостерический сдвиг. Измененный таким образом репрессор не может связываться с оператором, позволяя РНКП транскрибировать lac -гены и тем самым приводя к более высоким уровням кодируемых белков.
Второй механизм контроля — это реакция на глюкозу, которая использует гомодимер белка-активатора катаболита (CAP) для значительного увеличения производства β-галактозидазы в отсутствие глюкозы. Циклический аденозинмонофосфат (цАМФ) представляет собой сигнальную молекулу, распространенность которой обратно пропорциональна распространенности глюкозы. Он связывается с CAP, что, в свою очередь, позволяет CAP связываться с сайтом связывания CAP (последовательность ДНК длиной 16 п.н. выше промотора слева на диаграмме ниже, примерно на 60 п.н. выше сайта начала транскрипции). [8] который помогает РНКП связываться с ДНК. В отсутствие глюкозы концентрация цАМФ высока, и связывание CAP-цАМФ с ДНК значительно увеличивает выработку β-галактозидазы, позволяя клетке гидролизовать лактозу и высвобождать галактозу и глюкозу.
Совсем недавно было показано, что исключение индуктора блокирует экспрессию lac- оперона в присутствии глюкозы. Глюкоза транспортируется в клетку с помощью PEP-зависимой фосфотрансферазной системы . Фосфатная группа фосфоенолпирувата переносится посредством каскада фосфорилирования, состоящего из общих белков PTS (фосфотрансферазной системы) HPr и EIA и глюкозоспецифичных белков PTS EIIA. Глк и EIIB Глк , цитоплазматический домен транспортера глюкозы EII. Транспорт глюкозы сопровождается ее фосфорилированием с помощью EIIB. Глк , истощая фосфатную группу из других белков PTS, включая EIIA Глк . Нефосфорилированная форма EIIA Глк связывается с лактозой- пермеазой и предотвращает ее попадание лактозы в клетку. Следовательно, если присутствуют и глюкоза, и лактоза, транспорт глюкозы блокирует транспорт индуктора lac - оперона. [9]
Репрессорная структура
[ редактировать ]
Лак -репрессор представляет собой белок, состоящий из четырех частей, тетрамер, с идентичными субъединицами. Каждая субъединица содержит мотив спираль-поворот-спираль (HTH), способный связываться с ДНК. Операторный сайт, с которым связывается репрессор, представляет собой последовательность ДНК с инвертированной симметрией повторов. Два полусайта ДНК оператора вместе связываются с двумя субъединицами репрессора. Хотя две другие субъединицы репрессора в этой модели ничего не делают, это свойство не было понято в течение многих лет.
В конце концов было обнаружено, что в регулировании лака участвуют еще два оператора . [10] Один (O 3 ) находится примерно на -90 п.н. выше O 1 в конце гена lacI , а другой (O 2 ) находится примерно на +410 п.н. ниже O 1 в ранней части lacZ . Эти два сайта не были обнаружены в ранних работах, поскольку они имеют избыточные функции, а отдельные мутации не сильно влияют на репрессию. Одиночные мутации O 2 или O 3 имеют только 2-3-кратный эффект. Однако их важность подтверждается тем фактом, что двойной мутант, дефектный как по O 2 , так и по O 3, резко дерепрессируется (примерно в 70 раз).
В текущей модели lac -репрессор связан одновременно как с основным оператором O 1 , так и с O 2 или O 3 . Промежуточная ДНК выходит из комплекса. Избыточный характер двух второстепенных операторов предполагает, что важен не конкретный зацикленный комплекс. Одна из идей заключается в том, что система работает через привязку; если связанный репрессор на мгновение высвобождается из O 1 , связывание с второстепенным оператором удерживает его поблизости, так что он может быстро повторно связаться. Это увеличит сродство репрессора к O 1 .
Механизм индукции
[ редактировать ]
Репрессор является аллостерическим белком , т.е. он может принимать одну из двух слегка различающихся форм, находящихся в равновесии друг с другом. В одной форме репрессор будет связываться с операторной ДНК с высокой специфичностью, а в другой форме он утратил свою специфичность. Согласно классической модели индукции, связывание индуктора, аллолактозы или ИПТГ, с репрессором влияет на распределение репрессора между двумя формами. Таким образом, репрессор, связанный с индуктором, стабилизируется в несвязывающей с ДНК конформации. Однако эта простая модель не может дать полную картину, поскольку репрессор довольно стабильно связан с ДНК, но быстро высвобождается при добавлении индуктора. Следовательно, кажется очевидным, что индуктор также может связываться с репрессором, когда репрессор уже связан с ДНК. До сих пор не совсем известно, каков точный механизм связывания. [11]
Роль неспецифического связывания
[ редактировать ]Неспецифическое связывание репрессора с ДНК играет решающую роль в репрессии и индукции Lac-оперона. Сайтом специфического связывания белка Lac-репрессора является оператор. Неспецифическое взаимодействие опосредовано главным образом взаимодействиями заряд-заряд, тогда как связывание с оператором усиливается гидрофобными взаимодействиями. Кроме того, существует множество неспецифических последовательностей ДНК, с которыми может связываться репрессор. По сути, любая последовательность, не являющаяся оператором, считается неспецифической. Исследования показали, что без наличия неспецифического связывания индукция (или нерепрессия) Lac-оперона не могла бы произойти даже при насыщенных уровнях индуктора. Было продемонстрировано, что без неспецифического связывания базальный уровень индукции в десять тысяч раз меньше, чем обычно наблюдается. Это связано с тем, что неспецифическая ДНК действует как своего рода «приемник» для белков-репрессоров, отвлекая их от оператора. Неспецифические последовательности уменьшают количество доступного репрессора в клетке. Это, в свою очередь, уменьшает количество индуктора, необходимого для снятия давления в системе. [12]
Аналоги лактозы
[ редактировать ]
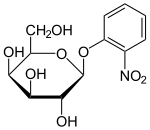

Описан ряд производных или аналогов лактозы, полезных для работы с lac- опероном. Эти соединения в основном представляют собой замещенные галактозиды, в которых глюкозный фрагмент лактозы заменен другой химической группой.
- Изопропил-β-D-тиогалактопиранозид (ИПТГ) часто используется в качестве индуктора lac- оперона для физиологической работы. [1] ИПТГ связывается с репрессором и инактивирует его, но не является субстратом для β-галактозидазы. Одним из преимуществ IPTG для исследований in vivo является то, что он не метаболизируется E. coli . Его концентрация остается постоянной, а скорость экспрессии генов, контролируемых lac p/o, не является переменной в эксперименте. Потребление ИПТГ зависит от действия пермеазы лактозы в P. fluorescens , но не в E. coli . [13]
- Фенил-β-D-галактоза (фенил-Гал) является субстратом для β-галактозидазы, но не инактивирует репрессор и поэтому не является индуктором. Поскольку клетки дикого типа производят очень мало β-галактозидазы, они не могут расти на фенил-гале в качестве источника углерода и энергии. Мутанты, лишенные репрессора, способны расти на фенил-гале. Таким образом, минимальная среда, содержащая только фенил-Гал в качестве источника углерода и энергии, является селективной для мутантов-репрессоров и мутантов-операторов. Если 10 8 клетки штамма дикого типа высеивают на чашки с агаром, содержащим фенил-Гал, редкие колонии, которые растут, представляют собой в основном спонтанные мутанты, влияющие на репрессор. На относительное распределение мутантов-репрессоров и операторов влияет размер цели. Поскольку ген lacI, кодирующий репрессор, примерно в 50 раз крупнее оператора, в отборе преобладают мутанты репрессора.
- Тиометилгалактозид [TMG] – еще один аналог лактозы. Они ингибируют репрессор lacI. При низких концентрациях индуктора как ТМГ, так и ИПТГ могут проникать в клетку через пермеазу лактозы. Однако при высоких концентрациях индуктора оба аналога могут проникать в клетку независимо. ТМГ может снижать скорость роста при высоких внеклеточных концентрациях. [14]
- Другие соединения служат красочными индикаторами активности β-галактозидазы.
- ONPG расщепляется с образованием интенсивно желтого соединения, ортонитрофенола и галактозы, и обычно используется в качестве субстрата для анализа β-галактозидазы in vitro . [15]
- Колонии, продуцирующие β-галактозидазу, окрашиваются в синий цвет под действием X-гал (5-бром-4-хлор-3-индолил-β-D-галактозид), который является искусственным субстратом для B-галактозидазы, расщепление которой приводит к образованию галактозы и 4-Cl. ,3-Br индиго, что дает глубокий синий цвет. [16]
- Аллолактоза является изомером лактозы и является индуктором lac- оперона. Лактоза представляет собой галактозо-β(1→4)-глюкозу, тогда как аллолактоза представляет собой галактозо-β(1→6)-глюкозу. Лактоза превращается в аллолактозу под действием β-галактозидазы в реакции, альтернативной гидролитической. Физиологический эксперимент, демонстрирующий роль LacZ в продукции «истинного» индуктора в клетках E. coli , представляет собой наблюдение, что нулевой мутант lacZ все еще может продуцировать пермеазу LacY при выращивании с IPTG, негидролизуемым аналогом аллолактозы, но не при выращивании с лактозой. Объяснение состоит в том, что для производства индуктора внутри клетки необходим процессинг лактозы в аллолактозу (катализируемый β-галактозидазой).
Развитие классической модели
[ редактировать ]Экспериментальным микроорганизмом, который использовали Франсуа Жакоб и Жак Моно, была обычная лабораторная бактерия E. coli , но многие из основных концепций регуляции, открытых Жакобом и Моно, имеют фундаментальное значение для клеточной регуляции во всех организмах. [17] Основная идея заключается в том, что белки не синтезируются, когда в них нет необходимости: E. coli сохраняет клеточные ресурсы и энергию, не производя три белка Lac, когда нет необходимости метаболизировать лактозу, например, когда другие сахара, такие как глюкоза доступны . В следующем разделе обсуждается, как E. coli контролирует определенные гены в ответ на метаболические потребности.
Во время Второй мировой войны Моно проверял влияние сочетаний сахаров в качестве источников питательных веществ на E. coli и B. subtilis . Моно продолжил аналогичные исследования, проведенные другими учеными с бактериями и дрожжами. Он обнаружил, что бактерии, выращенные с использованием двух разных сахаров, часто демонстрируют две фазы роста. Например, если были предоставлены одновременно глюкоза и лактоза, сначала метаболизировалась глюкоза (фаза роста I, см. Рисунок 2), а затем лактоза (фаза роста II). Лактоза не метаболизировалась в течение первой части диауктической кривой роста, поскольку β-галактозидаза не вырабатывалась, когда в среде присутствовали и глюкоза, и лактоза. Моно назвал это явление диакси . [18]

Затем Моно сосредоточил свое внимание на индукции образования β-галактозидазы, которая происходила, когда лактоза была единственным сахаром в культуральной среде. [19]
Классификация регуляторных мутантов
[ редактировать ]Концептуальный прорыв Джейкоба и Моно [20] заключалась в признании различия между регуляторными веществами и местами, в которых они действуют, изменяя экспрессию генов. Бывший солдат, Джейкоб использовал аналогию с бомбардировщиком, который сбрасывал свой смертоносный груз при получении специальной радиопередачи или сигнала. Для работы системы требуется как наземный передатчик, так и приемник в самолете. Теперь предположим, что обычный передатчик сломался. Эту систему можно заставить работать путем введения второго функционального передатчика. Для сравнения, сказал он, рассмотрим бомбардировщик с неисправной ствольной коробкой. Поведение этого бомбардировщика невозможно изменить введением второго функционального самолета.
Для анализа регуляторных мутантов lac -оперона Джейкоб разработал систему, с помощью которой вторая копия генов lac ( lacI с промотором и lacZYA с промотором и оператором) может быть введена в одну клетку. Культуру таких бактерий, которые являются диплоидными по lac -генам, но в остальном нормальными, затем тестируют на регуляторный фенотип. В частности, определяют, образуются ли LacZ и LacY даже в отсутствие IPTG (поскольку репрессор лактозы, продуцируемый мутантным геном, нефункционален). Этот эксперимент, в котором гены или кластеры генов тестируются попарно, называется тестом комплементации .

Этот тест проиллюстрирован на рисунке ( lacA опущен для простоты). Во-первых, показаны определенные гаплоидные состояния (т.е. клетка несет только одну копию lac- генов). На панели (a) показана репрессия, (b) показана индукция IPTG, а (c) и (d) показан эффект мутации гена lacI или оператора соответственно. На панели (e) показан тест комплементации репрессора. Если одна копия генов lac несет мутацию в lacI , а вторая копия является диким типом для lacI , результирующий фенотип является нормальным, но lacZ экспрессируется при воздействии индуктора IPTG. Говорят, что мутации, влияющие на репрессор, рецессивны по отношению к дикому типу (и этот дикий тип является доминантным ), и это объясняется тем фактом, что репрессор представляет собой небольшой белок, который может диффундировать в клетке. Копия оперона lac , примыкающая к дефектному гену lacI, эффективно отключается белком, продуцируемым из второй копии lacI .
Если тот же эксперимент проводится с использованием мутации оператора, получается другой результат (панель (f)). Фенотип клетки, несущей один мутантный и один операторный сайт дикого типа, заключается в том, что LacZ и LacY продуцируются даже в отсутствие индуктора IPTG; поскольку поврежденный операторный сайт не позволяет связыванию репрессора ингибировать транскрипцию структурных генов. Мутация оператора является доминантной. Когда операторный сайт, с которым должен связываться репрессор, повреждается мутацией, наличие второго функционального сайта в той же клетке не влияет на экспрессию генов, контролируемых мутантным сайтом.
В более сложной версии этого эксперимента используются маркированные опероны, чтобы различить две копии lac -генов и показать, что нерегулируемый структурный ген(ы) находится(ются) рядом с мутантным оператором (панель (g). Например, предположим, что одна копия отмечена мутацией, инактивирующей lacZ , так что она может производить только белок LacY, в то время как вторая копия несет мутацию, влияющую на lacY , и может производить только LacZ. В этой версии только копия оперона . lac соседний с мутантным оператором выражается без IPTG. Мы говорим, что мутация оператора является цис-доминантной , она доминантна по отношению к дикому типу, но влияет только на ту копию оперона, которая непосредственно соседствует с ним.
Это объяснение вводит в заблуждение в важном смысле, поскольку оно исходит из описания эксперимента, а затем объясняет результаты в терминах модели. Но на самом деле часто бывает так, что модель стоит на первом месте, и эксперимент проводится специально для проверки модели. Джейкоб и Моно сначала предположили, что в ДНК должен быть участок со свойствами оператора, а затем разработали свои тесты комплементации, чтобы показать это.
Доминирование операторных мутантов также предполагает процедуру их специфического отбора. Если регуляторные мутанты выбираются из культуры дикого типа с использованием фенил-Гал, как описано выше, операторные мутации редки по сравнению с мутантами-репрессорами, поскольку размер мишени настолько мал. Но если вместо этого мы начнем со штамма, который несет две копии всей lac- области (то есть диплоидной для lac ), репрессорные мутации (которые все еще происходят) не восстанавливаются, поскольку комплементация вторым геном lacI дикого типа придает дикий тип. фенотип. Напротив, мутация одной копии оператора придает мутантный фенотип, поскольку он доминирует над второй копией дикого типа.
Регуляция с помощью циклического АМФ [21]
[ редактировать ]Объяснение диаукси зависело от характеристики дополнительных мутаций, влияющих на lac -гены, отличных от тех, которые объясняются классической моделью. Впоследствии были идентифицированы два других гена, cya и crp , которые картированы далеко от lac и которые при мутировании приводят к снижению уровня экспрессии в присутствии IPTG и даже в штаммах бактерий, лишенных репрессора или оператора. Открытие цАМФ в E. coli привело к демонстрации того, что мутанты, дефектные по гену cya , но не по гену crp , могут быть восстановлены до полной активности путем добавления цАМФ в среду.
Ген cya кодирует аденилатциклазу, которая производит цАМФ. У мутанта cya отсутствие цАМФ делает экспрессию генов lacZYA более чем в десять раз ниже нормальной. Добавление цАМФ корректирует низкую экспрессию Lac, характерную для мутантов cya . Второй ген, crp , кодирует белок, называемый белком-активатором катаболита (CAP) или белком рецептора цАМФ (CRP). [22]
Однако ферменты метаболизма лактозы производятся в небольших количествах в присутствии как глюкозы, так и лактозы (иногда это называется «протекающей экспрессией») из-за того, что РНКП все еще может иногда связываться и инициировать транскрипцию даже в отсутствие CAP. Утечка экспрессии необходима для того, чтобы обеспечить метаболизм некоторого количества лактозы после того, как источник глюкозы израсходован, но до lac полной активации экспрессии .
В итоге:
- Когда лактоза отсутствует, фермент Lac вырабатывается очень мало (у оператора связан с ним репрессор Lac).
- Когда присутствует лактоза, но также присутствует предпочтительный источник углерода (например, глюкоза), тогда вырабатывается небольшое количество фермента (Lac-репрессор не связан с оператором).
- Когда глюкоза отсутствует, CAP-цАМФ связывается со специфическим участком ДНК выше промотора и осуществляет прямое белок-белковое взаимодействие с РНКП, что облегчает связывание РНКП с промотором.
Задержка между фазами роста отражает время, необходимое для производства достаточного количества ферментов, метаболизирующих лактозу. Во-первых, регуляторный белок CAP должен собраться на lac -промоторе, что приводит к увеличению продукции lac -мРНК . Большее количество доступных копий мРНК lac приводит к производству (см. перевод ) значительно большего количества копий LacZ (β-галактозидаза для метаболизма лактозы) и LacY (пермеаза лактозы для транспортировки лактозы в клетку). После задержки, необходимой для повышения уровня ферментов, метаболизирующих лактозу, бактерии вступают в новую быструю фазу роста клеток .

Две загадки катаболитной репрессии связаны с тем, как уровни цАМФ связаны с присутствием глюкозы, и, во-вторых, почему клетки вообще должны беспокоиться. После расщепления лактозы фактически образуются глюкоза и галактоза (легко превращающиеся в глюкозу). С метаболической точки зрения лактоза является таким же хорошим источником углерода и энергии, как и глюкоза. Уровень цАМФ связан не с внутриклеточной концентрацией глюкозы, а со скоростью транспорта глюкозы, которая влияет на активность аденилатциклазы. (Кроме того, транспорт глюкозы также приводит к прямому ингибированию пермеазы лактозы.) О том, почему E. coli действует таким образом, можно только предполагать. Все кишечные бактерии ферментируют глюкозу, что позволяет предположить, что они часто с ней сталкиваются. Возможно, что небольшая разница в эффективности транспорта или метаболизма глюкозы по сравнению с лактозой дает клеткам возможность регулировать lac- оперон таким способом. [23]
Использование в молекулярной биологии
[ редактировать ]Ген lac и его производные можно использовать в качестве репортерного гена в ряде методов селекции на основе бактерий, таких как двухгибридный анализ, в котором необходимо определить успешное связывание активатора транскрипции со специфической последовательностью промотора. [16] В чашках LB , содержащих X-gal , изменение цвета колоний от белых до синего соответствует примерно 20–100 единицам β-галактозидазы, в то время как тетразолий- лактозная среда и лактозная среда МакКонки имеют диапазон 100–1000 единиц и являются наиболее чувствительными к соответственно верхняя и нижняя части этого диапазона. [16] Поскольку лактозная среда МакКонки и тетразолий-лактозная среда основаны на продуктах распада лактозы, они требуют присутствия как lacZ , так и lacY генов . Таким образом, многие lac методы слияния , которые включают только ген lacZ, подходят для пластин X-gal. [16] или ОНПГ . жидкие бульоны [24]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Гриффитс, Энтони Дж. Ф.; Весслер, Сьюзен Р.; Кэрролл, Шон Б.; Добли, Джон (2015). Введение в генетический анализ (11-е изд.). Фриман, WH & Company. стр. 400–412. ISBN 9781464109485 .
- ^ Санганерия, Таниша; Бордони, Бруно (2024), «Генетика, индуцируемый оперон» , StatPearls , Остров сокровищ (Флорида): StatPearls Publishing, PMID 33232031 , получено 20 июня 2024 г.
- ^ МакКлин, Филипп (1997). «Экспрессия прокариотических генов» . ndsu.edu . Проверено 19 мая 2017 г.
- ^ «Экспрессия прокариотических генов» . ndsu.edu . Проверено 19 мая 2017 г.
- ^ «2.6: Лак-Оперон; CAP-сайт; ДНК-следы» . Свободные тексты по биологии . 3 января 2019 года . Проверено 20 июня 2024 г.
- ^ Басби С., Эбрайт Р.Х. (2001). «Активация транскрипции белком-активатором катаболита (CAP)». Дж. Мол. Биол . 293 (2): 199–213. дои : 10.1006/jmbi.1999.3161 . ПМИД 10550204 .
- ^ Кеннелл, Дэвид; Ризман, Ховард (июль 1977 г.). оперона Escherichia coli «Частоты инициации транскрипции и трансляции лакового ». Журнал молекулярной биологии . 114 (1): 1–21. дои : 10.1016/0022-2836(77)90279-0 . ПМИД 409848 .
- ^ Малан, Т. Филип; Колб, Энни; Бюк, Анри; МакКлюр, Уильям (декабрь 1984 г.). «Механизм активации CRP-цАМФ инициации транскрипции лак- оперона, активации промотора P1». Дж. Мол. Биол . 180 (4): 881–909. дои : 10.1016/0022-2836(84)90262-6 . ПМИД 6098691 .
- ^ Гёрке Б., Стюльке Дж (август 2008 г.). «Подавление углеродных катаболитов у бактерий: множество способов максимально эффективно использовать питательные вещества». Обзоры природы. Микробиология . 6 (8): 613–24. дои : 10.1038/nrmicro1932 . ПМИД 18628769 . S2CID 8782171 .
- ^ Олер, С.; Эйсманн, ER; Кремер, Х.; Мюллер-Хилл, Б. (1990). «Три оператора lac- оперона сотрудничают в репрессиях» . Журнал ЭМБО . 9 (4): 973–979. дои : 10.1002/j.1460-2075.1990.tb08199.x . ПМК 551766 . ПМИД 2182324 .
- ^ Гриффитс, Энтони Дж. Ф.; Гелбарт, Уильям М.; Миллер, Джеффри Х.; Левонтин, Ричард К. (1999). «Регуляция лактозной системы» . Современный генетический анализ . Нью-Йорк: WH Freeman. ISBN 0-7167-3118-5 .
- ^ фон Хиппель, PH; Ревзин А.; Гросс, Калифорния; Ван, AC (декабрь 1974 г.). «Неспецифическое связывание ДНК белков, регулирующих геном, как механизм биологического контроля: I. lac -оперон: аспекты равновесия» . ПНАС . 71 (12): 4808–12. дои : 10.1073/pnas.71.12.4808 . ПМК 433986 . ПМИД 4612528 .
- ^ Хансен Л.Х., Кнудсен С., Соренсен С.Дж. (июнь 1998 г.). «Влияние гена lacY на индукцию индуцируемых IPTG промоторов, изученных на Escherichia coli и Pseudomonas fluorescens » . Курс. Микробиол . 36 (6): 341–7. дои : 10.1007/s002849900320 . ПМИД 9608745 . S2CID 22257399 . Архивировано из оригинала 18 октября 2000 года.
- ^ Марбах А., Беттенброк К. (январь 2012 г.). « Индукция лак- оперона в Escherichia coli: систематическое сравнение индукции IPTG и TMG и влияния трансацетилазы LacA». Журнал биотехнологии . 157 (1): 82–88. дои : 10.1016/j.jbiotec.2011.10.009 . ПМИД 22079752 .
- ^ «Тест ОНПГ (β-галактозидаза)» . Сентябрь 2000 г. Архивировано из оригинала 3 ноября 2007 г. Проверено 25 октября 2007 г.
- ^ Jump up to: а б с д Йонг Дж., Рамм Э., Пабо С. (2000). «Бактериальная двухгибридная система селекции для изучения белок-ДНК и белок-белковых взаимодействий» . Proc Natl Acad Sci США . 97 (13): 7382–7. Бибкод : 2000PNAS...97.7382J . дои : 10.1073/pnas.110149297 . ПМК 16554 . ПМИД 10852947 .
- ^ «Веха 2 – Дальновидная пара: Природные вехи в экспрессии генов» . www.nature.com . Проверено 27 декабря 2015 г.
- ^ Мюллер-Хилл, Бенно (1996). Лак парадигмы Оперон, Краткая история генетической . Берлин: Вальтер де Грюйтер. стр. 7–10. ISBN 3-11-014830-7 .
- ^ Макнайт, Стивен Л. (1992). Транскрипционная регуляция . Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор. стр. 3–24 . ISBN 0-87969-410-6 .
- ^ Джейкоб Ф .; Моно Дж. (июнь 1961 г.). «Генетические регуляторные механизмы синтеза белков». Дж Мол Биол . 3 (3): 318–56. дои : 10.1016/S0022-2836(61)80072-7 . ПМИД 13718526 .
- ^ Монтмини, М. (1997). «Регуляция транскрипции с помощью циклического АМФ». Ежегодный обзор биохимии . 66 : 807–822. doi : 10.1146/annurev.biochem.66.1.807 . ISSN 0066-4154 . ПМИД 9242925 .
- ^ Ботсфорд, JL; Харман, Дж. Г. (март 1992 г.). «Циклический АМФ у прокариот» . Микробиологические обзоры . 56 (1): 100–122. дои : 10.1128/MMBR.56.1.100-122.1992 . ISSN 0146-0749 . ПМК 372856 . ПМИД 1315922 .
- ^ Васкес А., Бег К.К., Деменезес М.А. и др. (2008). «Влияние ограничения растворяющей способности на метаболизм E. coli» . BMC Сист Биол . 2 :7. дои : 10.1186/1752-0509-2-7 . ПМК 2270259 . ПМИД 18215292 .
- ^ «Индукция лакового оперона в E. coli» (PDF) . САПС. Архивировано из оригинала (PDF) 20 января 2022 года . Проверено 29 июня 2016 г.
Внешние ссылки
[ редактировать ]- Lac + Operan в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- лак- оперон на книжной полке NCBI [2]
- Представляем коллекцию анимации Virtual Cell Animation : Lac Operon
- Лак - Оперон: наука Бозмана
- Окрашивание цельных эмбрионов мыши на активность β-галактозидазы (lacZ)