остеокласт
| остеокласт | |
|---|---|
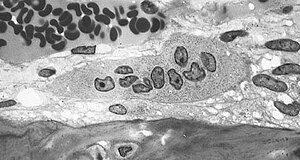 Светлая микрофотография остеокласта с типичными отличительными характеристиками: крупная клетка с множеством ядер и «пенистым» цитозолем. | |
 Иллюстрация, показывающая один остеокласт | |
| Подробности | |
| Предшественник | предшественники остеокластов |
| Расположение | Кость |
| Функция | Разрушение костной ткани |
| Идентификаторы | |
| латинский | остеокласт |
| МеШ | D010010 |
| ТД | Х2.00.03.7.00005 |
| ФМА | 66781 |
| Анатомические термины микроанатомии | |
Остеокласт костной (от древнегреческого ὀστέον (остеон) «кость» и κλαστός (кластос) «сломанный») — это тип клетки , которая разрушает костную ткань . для поддержания, восстановления ремоделирования костей . позвоночного скелета и Эта функция имеет решающее значение Остеокласты разбирают и переваривают смесь гидратированных белков и минералов на молекулярном уровне, секретируя кислоту и коллагеназу ; этот процесс известен как резорбция кости . Этот процесс также помогает регулировать уровень кальция в крови .
Остеокласты обнаруживаются на тех поверхностях кости, которые подвергаются резорбции. На таких поверхностях остеокласты располагаются в неглубоких углублениях, называемых резорбционными заливами (лакуны Хошипа) . Резорбционные заливы образуются в результате эрозивного действия остеокластов на подлежащую кость. Граница нижней части остеокласта имеет пальцеобразные отростки из-за наличия глубоких складок клеточной мембраны ; эта граница называется взъерошенной границей . Волнистая граница соприкасается с поверхностью кости в зоне резорбции. Периферия волнистой границы окружена кольцевидной зоной цитоплазмы , лишенной клеточных органелл , но богатой актиновыми нитями . Эта зона называется чистой зоной или зоной уплотнения . Актиновые нити позволяют клеточной мембране, окружающей зону уплотнения, прочно прикрепляться к костной стенке лакун Хошипа. Таким образом, между взъерошенной границей и резорбируемой костью создается закрытый субостеокластический отсек. Остеокласты секретируют ионы водорода , коллагеназа , катепсин К и гидролитические ферменты попадают в этот отсек. Резорбция костного матрикса остеокластами включает два этапа: (1) растворение неорганических компонентов (минералов) и (2) переваривание органических компонентов костного матрикса. Остеокласты перекачивают ионы водорода в субостеокластический компартмент и, таким образом, создают кислую микросреду, которая увеличивает растворимость костных минералов, что приводит к высвобождению и повторному входу костных минералов в цитоплазму остеокластов для доставки в близлежащие капилляры. После удаления минералов в субостеокластический отсек секретируются коллагеназа и желатиназа. Эти ферменты переваривают и разрушают коллаген и другие органические компоненты декальцинированного костного матрикса. Продукты деградации фагоцитируются остеокластами на взъерошенной границе. Из-за своих фагоцитарных свойств остеокласты считаются компонентом системы мононуклеарных фагоцитов (МПС). Активность остеокластов контролируется гормонами и цитокинами. Кальцитонин, гормон щитовидной железы, подавляет остеокластическую активность. Остеокласты не имеют рецепторов паратиреоидного гормона (ПТГ). Однако ПТГ стимулирует остеобласты секретируют цитокин, называемый фактором, стимулирующим остеокласты, который является мощным стимулятором активности остеокластов. [ 1 ]
Одонтокласт (/odon·to·clast/; o-don´to-klast) — остеокласт, связанный с рассасыванием корней временных зубов . [ 2 ] [ 3 ] [ 4 ]
Структура
[ редактировать ]

Остеокласт – это крупная многоядерная клетка , а остеокласты человека в кости обычно имеют четыре ядра. [ 5 ] и имеют диаметр 150–200 мкм. Когда цитокины, индуцирующие остеокласты, используются для превращения макрофагов в остеокласты, возникают очень большие клетки, которые могут достигать 100 мкм в диаметре. Они могут иметь десятки ядер и обычно экспрессируют основные белки остеокластов, но имеют существенные отличия от клеток живой кости из-за неприродного субстрата. [ 6 ] [ 7 ] Размер многоядерного собранного остеокласта позволяет ему сосредоточить возможности транспорта ионов, секреции белков и везикулярного транспорта многих макрофагов на локализованном участке кости.
Расположение
[ редактировать ]В кости остеокласты обнаруживаются в ямках на поверхности кости, которые называются резорбционными заливами или Хошипа лакунами . Остеокласты характеризуются цитоплазмой однородного, «пенистого» вида. Такой внешний вид обусловлен высокой концентрацией везикул и вакуолей . Эти вакуоли включают лизосомы , заполненные кислой фосфатазой . Это позволяет охарактеризовать остеокласты по их окрашиванию на высокую экспрессию тартрат -резистентной кислой фосфатазы (TRAP) и катепсина К. Шероховатая эндоплазматическая сеть остеокластов редкая, комплекс Гольджи обширен. [ 8 ] [ 9 ] [ 10 ]
В месте активной резорбции кости остеокласт образует специализированную клеточную мембрану — «гофрированную кайму», противостоящую поверхности костной ткани. Эта сильно складчатая или волнистая граница облегчает удаление кости за счет значительного увеличения поверхности клеток для секреции и поглощения содержимого резорбционного отдела и является морфологической характеристикой остеокластов, которые активно резорбируют кость.
Разработка
[ редактировать ]С момента их открытия в 1873 году возникли серьезные споры об их происхождении. Доминировали три теории: с 1949 по 1970 годы была популярна соединительнотканная теория происхождения, утверждавшая, что остеокласты и остеобласты относятся к одной линии, а остеобласты сливаются вместе, образуя остеокласты. После многих лет споров теперь стало ясно, что эти клетки развиваются в результате самослияния макрофагов. [ 11 ] В начале 1980-х годов моноцитарная фагоцитарная система была признана предшественником остеокластов. [ 12 ] Для образования остеокластов необходимо присутствие RANKL (рецептор-активатор лиганда ядерного фактора κβ) и M-CSF (макрофагальный колониестимулирующий фактор) . Эти мембраносвязанные белки продуцируются соседними стромальными клетками и остеобластами , что требует прямого контакта между этими клетками и предшественниками остеокластов .
M-CSF действует через свой рецептор на остеокластах, c-fms ( рецептор колониестимулирующего фактора 1 ), трансмембранный рецептор тирозинкиназы , что приводит к активации вторичного мессенджера тирозинкиназы Src. Обе эти молекулы необходимы для остеокластогенеза и широко участвуют в дифференцировке клеток, происходящих из моноцитов/макрофагов.
RANKL является членом семейства некрозов опухолей ( TNF ) и играет важную роль в остеокластогенезе. У мышей, нокаутных по RANKL, наблюдается фенотип остеопетроза и дефекты прорезывания зубов, а также отсутствие или недостаточность остеокластов. RANKL активирует NF-κβ (ядерный фактор-κβ) и NFATc1 (ядерный фактор активированных Т-клеток, цитоплазматический, кальциневрин-зависимый 1) посредством RANK . Активация NF-κβ стимулируется почти сразу после возникновения взаимодействия RANKL-RANK и не усиливается. Однако стимуляция NFATc1 начинается примерно через 24–48 часов после связывания, и было показано, что его экспрессия зависит от RANKL.
Дифференцировка остеокластов ингибируется остеопротегерином (OPG), который продуцируется остеобластами и связывается с RANKL, тем самым предотвращая взаимодействие с RANK. В то время как остеокласты происходят из гемопоэтической линии, остеобласты происходят из мезенхимальных стволовых клеток. [ 13 ] [ 14 ]
Функция
[ редактировать ]После активации остеокласты перемещаются в области микропереломов кости посредством хемотаксиса . Остеокласты лежат в небольших полостях, называемых лакунами Хошипа, образующихся в результате переваривания подлежащей кости. остеокластов Зона уплотнения представляет собой прикрепление плазматической мембраны к подлежащей кости. Зоны уплотнения ограничены поясами специализированных адгезионных структур, называемых подосомами . Прикреплению к костному матриксу способствуют рецепторы интегрина, такие как αvβ3, через специфический аминокислотный мотив Arg-Gly-Asp в белках костного матрикса, таких как остеопонтин . Остеокласт выделяет ионы водорода под действием карбоангидразы ( H 2 O + CO 2 → HCO 3 − + Ч + ) через волнистую границу в резорбтивную полость, подкисляя и способствуя растворению минерализованного костного матрикса в Ca 2+ , H 3 PO 4 , H 2 CO 3 , вода и другие вещества. Установлено, что дисфункция карбоангидразы вызывает некоторые формы остеопетроза. Ионы водорода перекачиваются против высокого градиента концентрации с помощью протонных насосов , в частности уникальной вакуолярной АТФазы . Этот фермент был нацелен на профилактику остеопороза . Кроме того, несколько гидролитических ферментов , таких как члены групп катепсина для переваривания органических компонентов матрикса высвобождаются и матриксных металлопротеаз (ММП). Эти ферменты высвобождаются в компартмент лизосомами . Из этих гидролитических ферментов наибольшее значение имеет катепсин К.
Катепсин К и другие катепсины
[ редактировать ]Катепсин К представляет собой коллагенолитическую папаинподобную цистеиновую протеазу , которая в основном экспрессируется в остеокластах и секретируется в резорбтивную ямку. Катепсин К — основная протеаза , участвующая в деградации коллагена I типа и других неколлагеновых белков. Мутации в гене катепсина К связаны с пикнодизостозом — наследственным остеопетрозным заболеванием, характеризующимся отсутствием функциональной экспрессии катепсина К. Исследования нокаута катепсина К на мышах приводят к остеопетротическому фенотипу, который частично компенсируется повышенной экспрессией протеаз, отличных от катепсина К, и усилением остеокластогенеза.
Катепсин К обладает оптимальной ферментативной активностью в кислых условиях. Он синтезируется в виде профермента с молекулярной массой 37 кДа и при активации путем автокаталитического расщепления трансформируется в зрелую активную форму с молекулярной массой ~27 кДа.
При поляризации остеокласта над местом резорбции катепсин К секретируется из взъерошенной границы в резорбтивную ямку. Катепсин К трансмигрирует через волнистую границу межклеточными везикулами и затем высвобождается функциональным секреторным доменом . Внутри этих межклеточных везикул катепсин К вместе с активными формами кислорода, генерируемыми TRAP , дополнительно разрушает костный внеклеточный матрикс.
Некоторые другие катепсины экспрессируются в остеокластах, включая B , C , D , E , G и L. катепсины Функция этих цистеиновых и аспарагиновых протеаз в костях обычно неизвестна, и они экспрессируются на гораздо более низких уровнях, чем катепсин К.
Исследования на мышах с нокаутом катепсина L были неоднозначными: сообщалось об уменьшении трабекулярной кости у гомозиготных и гетерозиготных мышей с нокаутом катепсина L по сравнению с мышами дикого типа, а в другом отчете не было обнаружено никаких скелетных аномалий.
Матриксные металлопротеиназы
[ редактировать ]Матриксные металлопротеиназы (ММП) включают семейство, состоящее из более чем 20 цинкзависимых эндопептидаз. Роль матриксных металлопротеиназ (ММП) в биологии остеокластов плохо определена, но в других тканях они связаны с активностью, способствующей развитию опухоли, такой как активация факторов роста , и необходимы для метастазирования опухоли и ангиогенеза.
MMP9 связан с микроокружением кости. Он экспрессируется остеокластами и, как известно, необходим для миграции остеокластов и является мощной желатиназой. У трансгенных мышей, лишенных MMP-9, развиваются дефекты развития костей, внутрикостного ангиогенеза и восстановления переломов.
Считается, что MMP-13 участвует в резорбции костей и дифференцировке остеокластов, поскольку у нокаутных мышей наблюдалось снижение количества остеокластов, остеопетроз и снижение резорбции костей.
ММП, экспрессируемые остеокластами, включают ММП-9, -10, -12 и -14. кроме MMP-9, мало что известно об их отношении к остеокластам, однако высокие уровни MMP-14 обнаруживаются в зоне уплотнения.
Физиология остеокластов
[ редактировать ]В 1980-х и 90-х годах была подробно изучена физиология типичных остеокластов. После выделения взъерошенной границы транспорт ионов через нее был изучен непосредственно в биохимических деталях. Был проверен энергозависимый транспорт кислоты и очищен постулируемый протонный насос. [ 15 ] [ 16 ] Благодаря успешному культивированию остеокластов стало очевидно, что они организованы для поддержки массивного транспорта протонов для подкисления резорбционного отдела и солюбилизации костных минералов. Сюда входит гофрированная кайма Cl. − проницаемость для контроля мембранного потенциала и базолатерального Cl − /HCO 3 − обмен для поддержания цитозольного pH в физиологически приемлемом диапазоне. [ 17 ] [ 18 ] [ 19 ]
Эффективность секреции ионов зависит от остеокластов, образующих эффективное уплотнение вокруг отделения резорбции. Позиционирование этой «зоны запечатывания», по-видимому, опосредовано интегринами, экспрессируемыми на поверхности остеокластов. [ 20 ] Когда зона уплотнения установлена, многоядерный остеокласт реорганизуется. Развитие сильно инвагинированной взъерошенной мембраны, примыкающей к резорбционному отсеку, обеспечивает массивную секреторную активность. Кроме того, он обеспечивает везикулярный трансцитоз минерала и деградированного коллагена от взъерошенной границы к свободной мембране клетки и его высвобождение во внеклеточный отсек. [ 21 ] [ 22 ] Эта деятельность завершает резорбцию кости, и минеральные компоненты и фрагменты коллагена высвобождаются в общий кровоток.
Регулирование
[ редактировать ]Остеокласты регулируются несколькими гормонами , включая паратиреоидный гормон (ПТГ) паращитовидной железы, кальцитонин щитовидной железы и фактор роста интерлейкин 6 (IL-6). Последний гормон, IL-6 , является одним из факторов заболевания остеопорозом . Остеопороз возникает, когда существует дисбаланс между активностью остеокластов по резорбции кости и деятельностью остеобластов по костеобразованию. [ 23 ]
Активность остеокластов также опосредуется взаимодействием двух молекул, продуцируемых остеобластами, а именно остеопротегерина и лиганда RANK . Эти молекулы также регулируют дифференцировку остеокластов. [ 24 ] [ 25 ]
одонтокласт
[ редактировать ]Одонтокласт (/odon·to·clast/; o-don´to-klast) — остеокласт, связанный с рассасыванием корней временных зубов . [ 2 ] [ 3 ] [ 4 ]
Альтернативное использование термина
[ редактировать ]Остеокласт также может быть инструментом, используемым для перелома и восстановления костей (происхождение греческого остеон — кость и кластос — сломанный). Во избежание путаницы клетку первоначально называли озотокластом. Когда хирургический инструмент вышел из употребления, клетка стала называться под своим нынешним названием.
Клиническое значение
[ редактировать ]Гигантские остеокласты могут возникать при некоторых заболеваниях, включая болезнь Педжета и токсичность бисфосфонатов .
У кошек аномальная активность одонтокластов может вызывать одонтокластические резорбтивные поражения , приводящие к необходимости удаления пораженных зубов.
Остеокласты играют важную роль в ортодонтическом перемещении зубов и патологической миграции зубов с нарушениями пародонта.
История
[ редактировать ]Остеокласты были открыты Кёлликером в 1873 году. [ 12 ]
См. также
[ редактировать ]- Список типов клеток человека, полученных из зародышевых листков
- Список различных типов клеток в организме взрослого человека
Ссылки
[ редактировать ]- ^ Медицинская гистология Лайка Хуссейна Сиддики (6-е издание)
- ^ Перейти обратно: а б «Одонтокласт» . Фарлекс, Бесплатный словарь. 2007 . Проверено 6 ноября 2013 г.
- ^ Перейти обратно: а б Ван З, МакКоли Л.К. (март 2011 г.). «Остеокласты и одонтокласты: сигнальные пути развития и болезней» . Заболевания полости рта . 17 (2): 129–42. дои : 10.1111/j.1601-0825.2010.01718.x . ПМИД 20659257 .
- ^ Перейти обратно: а б Чаттерджи К. (1 декабря 2006 г.). Основы оральной гистологии . Издательство Jaypee Brothers. п. 155. ИСБН 978-81-8061-865-9 . [ постоянная мертвая ссылка ]
- ^ Пайпер К., Бойд А., Джонс С.Дж. (1992). «Взаимосвязь между количеством ядер остеокласта и его резорбтивной способностью in vitro». Анатомия и эмбриология . 186 (4): 291–9. дои : 10.1007/BF00185977 . ПМИД 1416078 . S2CID 7570118 .
- ^ Базель МФ, Мазо П., Малкани К., Кретьен М.Ф., Моро М.Ф., Ребел А (1988). «Выделение остеокластов из костной ткани Пейджета: морфометрия и цитохимия на изолированных клетках». Кость . 9 (1): 1–6. дои : 10.1016/8756-3282(88)90020-8 . ПМИД 2837260 .
- ^ Джайн Н., Вайнштейн Р.С. (2009). «Гигантские остеокласты после длительной терапии бисфосфонатами: проблемы диагностики» . Нат Рев Ревматол . 5 (6): 341–6. дои : 10.1038/nrreum.2009.87 . ПМЦ 2860596 . ПМИД 19491914 .
- ^ Стэндринг С., изд. (2005). Анатомия Грея (39-е изд.). Эльзевир.
- ^ Холтроп М.Э., Кинг Дж.Дж. (1977). «Ультраструктура остеокластов и ее функциональные последствия» . Клиническая ортопедия и связанные с ней исследования . 123 (123): 177–96. дои : 10.1097/00003086-197703000-00062 . ПМИД 856515 . S2CID 30010574 . Архивировано (PDF) из оригинала 25 марта 2024 г.
- ^ Вяэнянен Х.К., Чжао Х., Мулари М., Халлин Дж.М. (февраль 2000 г.). «Клеточная биология функции остеокластов» . Журнал клеточной науки . 113 (3): 377–81. дои : 10.1242/jcs.113.3.377 . ПМИД 10639325 .
- ^ С. Л. Тейтельбаум (2000), «Резорбция кости остеокластами», Science , 289 (5484): 1504–1508, Бибкод : 2000Sci...289.1504T , doi : 10.1126/science.289.5484.1504 , PMID 10968780
- ^ Перейти обратно: а б Ниджвейде П.Дж., Бургер Э.Х., Фейен Дж.Х. (октябрь 1986 г.). «Клетки кости: пролиферация, дифференцировка и гормональная регуляция». Физиологические обзоры . 66 (4): 855–86. дои : 10.1152/physrev.1986.66.4.855 . ПМИД 3532144 .
- ^ Мишу, Летиция; Нуман, Мохамед; Дружелюбная, Натали; Браун, Жак П. (2015). «Болезнь Педжета костей: остеоиммунологическое заболевание?» . Дизайн, разработка и терапия лекарств . 9 : 4695–707. дои : 10.2147/DDDT.S88845 . ПМЦ 4544727 . ПМИД 26316708 .
- ^ Нуман, Мохамед; Браун, Жак; Мишу, Летиция (2015). «Влияние загрязнителей воздуха на окислительный стресс при распространенных заболеваниях старения, опосредованных аутофагией» . Международный журнал экологических исследований и общественного здравоохранения . 12 (2): 2289–2305. дои : 10.3390/ijerph120202289 . ПМЦ 4344726 . ПМИД 25690002 .
- ^ Блэр Х.К., Тейтельбаум С.Л., Гизелли Р., Глюк С. (август 1989 г.). «Остеокластическая резорбция кости с помощью поляризованного вакуолярного протонного насоса». Наука . 245 (4920): 855–7. Бибкод : 1989Sci...245..855B . дои : 10.1126/science.2528207 . ПМИД 2528207 .
- ^ Мэттссон Дж.П., Шлезингер П.Х., Килинг DJ, Тейтельбаум С.Л., Стоун Д.К., Се XS (октябрь 1994 г.). «Выделение и восстановление протонного насоса вакуолярного типа мембран остеокластов» . Журнал биологической химии . 269 (40): 24979–82. дои : 10.1016/S0021-9258(17)31485-0 . ПМИД 7929181 .
- ^ Тети А., Блэр Х.К., Тейтельбаум С.Л., Кан А.Дж., Козиол С., Консек Дж., Замбонин-Заллоне А., Шлезингер П.Х. (январь 1989 г.). «Регуляция цитоплазматического pH и обмен хлоридов/бикарбонатов в остеокластах птиц» . Журнал клинических исследований . 83 (1): 227–33. дои : 10.1172/jci113863 . ПМК 303666 . ПМИД 2910910 .
- ^ Блэр Х.К., Тейтельбаум С.Л., Тан Х.Л., Козиол С.М., Шлезингер П.Х. (июнь 1991 г.). «Заряд пассивной проницаемости хлоридов, связанный с H (+)-АТФазой взъерошенной мембраны остеокластов птиц». Американский журнал физиологии . 260 (6, ч. 1): C1315-24. дои : 10.1152/ajpcell.1991.260.6.C1315 . ПМИД 1829326 .
- ^ Шлезингер П.Х., Блэр Х.К., Тейтельбаум С.Л., Эдвардс Дж.К. (июль 1997 г.). «Характеристика хлоридного канала с взъерошенной границей остеокластов и его роль в резорбции кости» . Журнал биологической химии . 272 (30): 18636–43. дои : 10.1074/jbc.272.30.18636 . ПМИД 9228032 .
- ^ Вяэнянен Х.К., Чжао Х., Мулари М., Халлин Дж.М. (февраль 2000 г.). «Клеточная биология функции остеокластов». Журнал клеточной науки . 113 (Часть 3) (3): 377–81. дои : 10.1242/jcs.113.3.377 . ПМИД 10639325 .
- ^ Сало Дж., Лехенкари П., Мулари М., Метсикко К., Вяэнянен Х.К. (апрель 1997 г.). «Удаление продуктов резорбции кости остеокластов путем трансцитоза». Наука . 276 (5310): 270–3. дои : 10.1126/science.276.5310.270 . ПМИД 9092479 .
- ^ Несбитт С.А., Хортон Массачусетс (апрель 1997 г.). «Транспорт матриксных коллагенов через остеокласты, резорбирующие кость». Наука . 276 (5310): 266–9. дои : 10.1126/science.276.5310.266 . ПМИД 9092478 .
- ^ Танец, Эмбер (23 февраля 2022 г.). «Забавные факты о костях: больше, чем просто строительные леса» . Знающий журнал . дои : 10.1146/knowable-022222-1 . Проверено 8 марта 2022 г.
- ^ Шоппет М., Прейсснер К.Т., Хофбауэр Л.С. (апрель 2002 г.). «Лиганд RANK и остеопротегерин: паракринные регуляторы костного метаболизма и сосудистой функции» . Атеросклероз, тромбоз и сосудистая биология . 22 (4): 549–53. дои : 10.1161/01.ATV.0000012303.37971.DA . ПМИД 11950689 .
- ^ Ширази, Саджад; Равиндран, Шрирам; Купер, Линдон Ф. (2022). «Топографически-опосредованная иммуномодуляция при остеоинтеграции; союзник или враг» . Биоматериалы . 291 : 121903. doi : 10.1016/j.bimaterials.2022.121903 . ISSN 1878-5905 . ПМЦ 10148651 . ПМИД 36410109 .
Внешние ссылки
[ редактировать ]- MedicineNet. Архивировано 25 мая 2013 г. в Wayback Machine.
- Остеокласты Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- Жизнь остеокласта
- Random42 : Анимация от Random42 Scientific Communication о роли остеокластов в ремоделировании кости.