Трихома
Эта статья нуждается в дополнительных цитатах для проверки . ( октябрь 2022 г. ) |


Трихомы ( / ˈt r aɪ k oʊ m t z , ˈ r ɪ k oʊ m ») — z / ; от древнегреческого τρίχωμα ( tríkhōma ) « волосы тонкие выросты или придатки на растениях , водорослях , лишайниках и некоторых протистах . Они имеют разнообразную структуру и функции. Примерами являются волосы, железистые волоски, чешуйки и сосочки. Покрытие любого вида волосков на растении — волосок , а поверхность, несущая их, называется опушенной .
Водорослевые трихомы
[ редактировать ], обычно нитчатых, У некоторых водорослей концевая клетка образует удлиненную волосоподобную структуру, называемую трихомой. [ нужен пример ] Тот же термин применяется к таким структурам у некоторых цианобактерий , таких как Spirulina и Oscillatoria . Трихомы цианобактерий могут быть непокрытыми, как у Oscillatoria , или покрытыми оболочкой, как у Calothrix . [1] Эти структуры играют важную роль в предотвращении эрозии почвы , особенно в холодном пустынном климате . [ нужна ссылка ] Нитчатые оболочки образуют стойкую липкую сеть, которая помогает поддерживать структуру почвы.
Растительные трихомы
[ редактировать ]




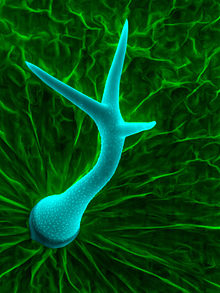


Трихомы растений имеют множество различных особенностей, которые различаются как у разных видов растений, так и у органов отдельного растения. Эти особенности влияют на подкатегории, в которые помещаются трихомы. Некоторые определяющие особенности включают следующее:
- Одноклеточные или многоклеточные
- Прямой (вертикальный, практически без разветвлений), спиральный (в форме штопора) или крючковатый (изогнутая вершина). [2]
- Наличие цитоплазмы
- Железистый (секреторный) против эгландулярного
- Извилистые, простые (неразветвленные и одноклеточные), пельтатные (чешуйчатые), звездчатые (звездчатые). [3]
- Адаксиальный или абаксиальный , относящийся к тому, присутствуют ли трихомы соответственно на верхней поверхности (адаксиальный) или нижней поверхности (абаксиальный) листа или другого бокового органа.
В модельном организме Cistus salviifolius на этом растении присутствует больше адаксиальных трихом, поскольку эта поверхность страдает от большего стресса от ультрафиолетового (УФ) и солнечного излучения , чем абаксиальная поверхность. [4]
Трихомы могут защитить растение от широкого спектра вредных воздействий, таких как ультрафиолетовое излучение, насекомые, транспирация и непереносимость заморозков. [5]
Воздушные поверхностные волоски
[ редактировать ]Трихомы на растениях представляют собой эпидермальные разного рода выросты. Термины «появление» или «колючки» относятся к выростам, которые затрагивают не только эпидермис. Это различие не всегда легко применить (см. « Дерево погоди-минуты» ). Также существуют нетрихоматозные эпидермальные клетки, выступающие из поверхности, например, корневые волоски .
Распространенный тип трихомы — волос . Волосы растений могут быть одноклеточными и многоклеточными , разветвленными и неразветвленными. Многоклеточные волоски могут иметь один или несколько слоев клеток. Разветвленные волоски могут быть дендритными (древовидными), как у лапы кенгуру ( Anigozanthos ), пучковыми или звездчатыми (звездообразными), как у Arabidopsis thaliana .
Другим распространенным типом трихом являются чешуйчатые или пельтатные волосы , которые имеют пластинчатую или щитовидную группу клеток, прикрепленных непосредственно к поверхности или расположенных на каком-либо стебле. Распространенными примерами являются листовые чешуи бромелиевых, таких как ананас , рододендрон и облепиха ( Hippophae rhamnoides ).
Любой из различных типов волос может быть железистые , вырабатывающие какой-то секрет, например, эфирные масла, вырабатываемые мятой и многими другими представителями семейства яснотковых .
, используется множество терминов, Для описания внешнего вида органов растения, таких как стебли и листья относящихся к наличию, форме и внешнему виду трихом. Примеры включают в себя:
- голые , голые – без волосков и трихом; поверхность гладкая
- волосатый – грубо волосатый
- hispid – имеющий щетинистые волосы
- членистые – простые многоклеточно-однорядные волоски
- пуховый – имеющий почти шерстяной покров из длинных волосков.
- волосатый - опушенный длинными, прямыми, мягкими, раскидистыми или стоячими волосками.
- половозрелый – ежеминутно опушенный; наличие тонких, коротких, обычно стоячих волос.
- половозрелый – слегка покрыт мелкими мягкими и торчащими волосками.
- опушенный - несущий волоски или трихомы любого типа.
- стригиллоза - стригиллоза минутная
- полосатый - прямые волосы направлены более или менее в том же направлении, что и по краю или средней жилке.
- tomentellous – мелко войлочный
- войлочный – покрыт густыми, спутанными, пушистыми волосками.
- ворсинчатый – мелковорсинчатый
- ворсинчатый – с длинными мягкими волосками, часто изогнутыми, но не спутанными.
Размер, форма, густота и расположение волосков на растениях чрезвычайно изменчивы по их наличию у разных видов и даже внутри вида на разных органах растения. Можно перечислить несколько основных функций или преимуществ наличия волос на поверхности. что во многих случаях волосы мешают питанию по крайней мере некоторых мелких травоядных , а также, в зависимости от жесткости и раздражительности нёба Вполне вероятно , , и крупных травоядных. Волосы на растениях, растущих в районах, подверженных морозу, защищают живые поверхностные клетки от мороза. В ветреных местах волоски прерывают поток воздуха по поверхности растения, уменьшая транспирацию . Плотный покров волосков отражает солнечный свет , защищая более нежные ткани под ним в жарких, сухих и открытых местах обитания. Кроме того, в местах, где большая часть доступной влаги поступает из капель тумана , волосы, по-видимому, усиливают этот процесс, увеличивая площадь поверхности, на которой могут накапливаться капли воды. [ нужна ссылка ]
Железистые трихомы
[ редактировать ]Железистые трихомы широко изучены, хотя они встречаются только примерно у 30% растений. Их функция — выделять метаболиты для растения. Некоторые из этих метаболитов включают:
- терпеноиды , которые имеют множество функций, связанных с защитой, ростом и развитием. [6]
- фенилпропаноиды , которые играют роль во многих путях развития растений, таких как вторичные метаболиты, реакция на стресс, и действуют как медиаторы взаимодействий растений в окружающей среде. [7]
- флавоноиды
- метилкетоны
- ацилсахара [8]
Нежелезистые трихомы
[ редактировать ]Нежелезистые трихомы служат структурной защитой от различных абиотических стрессоров, включая потерю воды, экстремальные температуры и УФ-излучение, а также биотические угрозы, такие как нападение патогенов или травоядных животных. [9]
Например, модельное растение C. salviifolius встречается в районах с сильным световым стрессом и плохими почвенными условиями вдоль побережий Средиземного моря. Он содержит нежелезистые, звездчатые и дендритные трихомы, которые обладают способностью синтезировать и хранить полифенолы, которые влияют как на поглощение радиации, так и на высыхание растений. Эти трихомы также содержат ацетилированные флавоноиды, которые могут поглощать УФ-В, и неацетилированные флавоноиды, которые поглощают более длинные волны УФ-А. В нежелезистых трихомах единственная известная роль флавоноидов — блокировать самые короткие волны для защиты растения; это отличается от их роли в железистых трихомах. [4]
У родов Salix и gossypium модифицированные трихомы образуют ватные волокна, которые обеспечивают анемохию или распространение с помощью ветра. Эти семенные трихомы являются одними из самых длинных растительных клеток. [10]
Полифенолы
[ редактировать ]Было обнаружено, что нежелезистые трихомы рода Cistus содержат эллагитаннины, гликозиды и производные кемпферола . Основная цель эллаготаннинов — помочь адаптироваться во время стресса, ограничивающего питательные вещества. [4]
Развитие трихома и корневых волосков
[ редактировать ]И трихомы, и корневые волоски , ризоиды многих сосудистых растений , представляют собой боковые выросты одной клетки эпидермального слоя. Корневые волоски образуются из трихобластов — волосообразующих клеток эпидермиса корня растения . Корневые волоски имеют диаметр от 5 до 17 микрометров и длину от 80 до 1500 микрометров (Диттмар, цит. по Исаву, 1965). Корневые волоски могут сохраняться в течение двух-трех недель, а затем отмирать. В то же время на верхушке корня постоянно образуются новые корневые волоски. Таким образом, покрытие корней волос останется прежним. Поэтому понятно, что пересадку следует производить с осторожностью, поскольку по большей части корневые волоски удаляются. Вот почему высадка может привести к увяданию растений.
Генетический контроль формирования рисунка трихом и корневых волос имеет схожие механизмы контроля. Оба процесса включают ядро родственных транскрипционных факторов , которые контролируют инициацию и развитие эпидермального разрастания. Активация генов, которые кодируют специфические белковые факторы транскрипции (названные GLABRA1 (GL1), GLABRA3 (GL3) и TRANSPARENT TESTA GLABRA1 (TTG1)) являются основными регуляторами судьбы клеток для производства трихом или корневых волосков. [11] Когда эти гены активируются в эпидермальной клетке листа, внутри этой клетки инициируется образование трихрома. ГЛ1, ГЛ3. и TTG1 также активируют негативные регуляторы, которые ингибируют образование трихромов в соседних клетках. Эта система контролирует расстояние между трихомами на поверхности листа. Как только трихомы разовьются, они могут делиться или разветвляться. [12] Напротив, корневые волоски ветвятся лишь изредка. При образовании трихом и корневых волосков регулируется деятельность многих ферментов. Например, непосредственно перед развитием корневых волосков существует точка повышенной активности фосфорилазы . [13]
Многое из того, что ученые знают о развитии трихом, происходит от модельного организма Arabidopsis thaliana , поскольку его трихомы простые, одноклеточные и нежелезистые. Путь развития регулируется тремя факторами транскрипции: R2R3 MYB , основной спираль-петля-спираль и повтором WD40 . Три группы ТФ образуют тримерный комплекс (MBW) и активируют экспрессию продуктов ниже по ходу процесса, что активирует образование трихом. Однако только MYB действуют как ингибитор, образуя негативный комплекс. [14]
Фитогормоны
[ редактировать ]Фитогормоны растений оказывают влияние на рост и реакцию растений на раздражители окружающей среды. Некоторые из этих фитогормонов участвуют в образовании трихом, к ним относятся гибберелловая кислота (ГК), цитокинины (КК) и жасмоновые кислоты (ЖК). ГК стимулирует рост трихом путем стимуляции GLABROUS1 (GL1), однако белки SPINDLY и DELLA подавляют действие GA, поэтому меньшее количество этих белков создает больше трихом. [14]
Некоторые другие фитогормоны, способствующие росту трихом, включают брассиностероиды, этилен и салициловую кислоту. Это удалось понять, проведя эксперименты с мутантами, у которых практически нет каждого из этих веществ. В каждом случае на обеих поверхностях растений наблюдалось меньшее образование трихом, а также неправильное формирование присутствующих трихом. [14]
Значение для таксономии
[ редактировать ]Тип, наличие, отсутствие и расположение трихом являются важными диагностическими признаками при идентификации растений и таксономии растений. [15] При судебно-медицинской экспертизе такие растения, как Cannabis sativa, можно идентифицировать путем микроскопического исследования трихом. [16] [17] Хотя трихомы редко встречаются сохранившимися в окаменелостях , основания трихом встречаются регулярно, и в некоторых случаях их клеточная структура важна для идентификации.
Arabidopsis thaliana Классификация трихом
[ редактировать ]Трихомы Arabidopsis thaliana классифицируются как воздушные, эпидермальные, одноклеточные и трубчатые структуры. [18]
Значение для молекулярной биологии растений
[ редактировать ]У модельного растения Arabidopsis thaliana образование трихом инициируется белком GLABROUS1. Нокауты соответствующего гена приводят к голым растениям. Этот фенотип уже использовался в экспериментах по редактированию генома и может представлять интерес в качестве визуального маркера для исследований растений с целью улучшения методов редактирования генов, таких как CRISPR/Cas9 . [19] [20] Трихомы также служат моделями клеточной дифференцировки и формирования узоров у растений. [21]
Использование
[ редактировать ]Листья фасоли исторически использовались для отлова клопов в домах Восточной Европы . Трихомы на листьях фасоли захватывают насекомых, пронзая их лапки ( лапки ). Тогда листья будут уничтожены. [22]
Трихомы являются важной частью строительства гнезда европейской пчелы-чесальщика шерсти ( Anthidium manicatum ). Этот вид пчел включает трихомы в свои гнезда, соскребая их с растений и используя в качестве подкладки для полостей гнезд. [23]
Оборона
[ редактировать ]Растения могут использовать трихомы для отражения атак травоядных с помощью физических и/или химических средств, например, в специализированных жалящих волосках видов Urtica (Крапива), которые доставляют воспалительные химические вещества, такие как гистамин . Исследования трихом были направлены на защиту сельскохозяйственных культур, что является результатом отпугивания травоядных животных (Brookes et al. 2016). [24] Однако некоторые организмы выработали механизмы сопротивления воздействию трихом. Личинки Heliconius charithonia , например, способны физически освобождаться от трихом, способны откусывать трихомы, способны образовывать шелковые покрывала, чтобы лучше перемещаться по листьям. [25]
Жалящие трихомы
[ редактировать ]Жалящие трихомы различаются по своей морфологии и распространению между видами, однако сходное воздействие на крупных травоядных предполагает, что они выполняют схожие функции. В районах, восприимчивых к травоядным животным, наблюдалась более высокая плотность жалящих трихом. У Urtica жалящие трихомы вызывают болезненные ощущения, продолжающиеся в течение нескольких часов при контакте с человеком. Это ощущение считается защитным механизмом против крупных животных и мелких беспозвоночных и играет роль в усилении защиты посредством секреции метаболитов. Исследования показывают, что это ощущение связано с быстрым выбросом токсина (например, гистамина ) при контакте и проникновении через шаровидные кончики указанных трихом. [26]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Определите эту водоросль» . Проверено 20 сентября 2013 г.
- ^ Пейн, Уиллард В. (1978). «Словарь терминологии растительных волос». Бриттония . 30 (2): 239–255. Бибкод : 1978Britt..30..239P . дои : 10.2307/2806659 . ISSN 0007-196X . JSTOR 2806659 . S2CID 42417527 .
- ^ Левин, Дональд А. (1973). «Роль трихом в защите растений». Ежеквартальный обзор биологии . 48 (1): 3–15. дои : 10.1086/407484 . ISSN 0033-5770 . JSTOR 2822621 . S2CID 83358711 .
- ^ Перейти обратно: а б с Таттини, М.; Маттейни, П.; Сарачини, Э.; Траверси, МЛ; Джордано, К.; Агати, Г. (2007). «Морфология и биохимия нежелезистых трихом листьев Cistus salvifolius L., произрастающих в экстремальных местообитаниях Средиземноморского бассейна». Биология растений . 9 (3): 411–419. Бибкод : 2007PlBio...9..411T . дои : 10.1055/s-2006-924662 . ISSN 1438-8677 . ПМИД 17143807 .
- ^ Хюльскамп, Мартин (июнь 2004 г.). «Растительные трихомы: модель дифференцировки клеток». Молекулярно-клеточная биология . 5 (6): 471–480. дои : 10.1038/nrm1404 . ПМИД 15173826 . S2CID 26541461 – через EBSCO.
- ^ Толл, Доротея (2015), «Биосинтез и биологические функции терпеноидов в растениях», Шрейдер, Йенс; Больманн, Йорг (ред.), Биотехнология изопреноидов , Достижения в области биохимической инженерии/биотехнологии, том. 148, Springer International Publishing, стр. 63–106, номер документа : 10.1007/10_2014_295 , ISBN. 9783319201078 , PMID 25583224
- ^ Ясинский, Михал; Бяла, Ванда (2018). «Дело о фенилпропаноиде: важен транспорт» . Границы в науке о растениях . 9 : 1610. дои : 10.3389/fpls.2018.01610 . ISSN 1664-462X . ПМК 6221964 . ПМИД 30443262 .
- ^ Хухельманн, Александр; Бутри, Марк; Хачес, Чарльз (2017). «Железистые трихомы растений: фабрики натуральных клеток, представляющие высокий биотехнологический интерес1 [ОТКРЫТЬ]» . Физиология растений . 175 (1): 6–22. дои : 10.1104/стр.17.00727 . ISSN 0032-0889 . ПМЦ 5580781 . ПМИД 28724619 .
- ^ Карабурниотис, Джордж; Лиакопулос, Георгиос; Николопулос, Димосфенис; Бреста, Панайота (01 февраля 2020 г.). «Защитная и защитная роль нежелезистых трихом против множественных стрессов: координация структуры и функции» . Журнал лесных исследований . 31 (1): 1–12. дои : 10.1007/s11676-019-01034-4 . ISSN 1993-0607 .
- ^ Шуй Ван, Цзя-Вэй Ван, Нань Юй, Чун-Хун Ли, Бинь Ло, Цзинь-Ин Гоу, Лин-Цзянь Ван и Сяо-Я Чен (сентябрь 2004 г.). «Контроль развития трихом растений с помощью хлопкового волокна MYB GeneW⃞» . Растительная клетка . 16 (9): 2323–2334. дои : 10.1105/tpc.104.024844 . ПМК 520936 . ПМИД 15316114 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Шеллманн, С.; Шнитгер, А.; Кирик, В.; Вада, Т.; Окада, К.; Берманн, А.; Тумфарт, Дж.; Юргенс, Г.; Хюльскамп, М. (01 октября 2002 г.). «TRIPTYCHON и CAPRICE опосредуют латеральное торможение во время формирования рисунка трихом и корневых волосков у Arabidopsis» . Журнал ЭМБО . 21 (19): 5036–5046. дои : 10.1093/emboj/cdf524 . ISSN 0261-4189 . ПМК 129046 . ПМИД 12356720 .
- ^ Шеллманн, Свен; Халскамп, Мартин (1 сентября 2004 г.). «Эпидермальная дифференциация: трихомы Arabidopsis как модельная система» . Международный журнал биологии развития . 49 (5–6): 579–584. дои : 10.1387/ijdb.051983ss . ISSN 0214-6282 . ПМИД 16096966 .
- ^ Дозье, Ларри В.; Риопель, Дж.Л. (1977). «Дифференциальная активность ферментов во время дифференцировки трихобластов у Elodea canadensis». Американский журнал ботаники . 64 (9): 1049–1056. дои : 10.2307/2442160 . JSTOR 2442160 .
- ^ Перейти обратно: а б с Юань, Линг; Сингх, Санджай Кумар; Патра, Барунава; Паттанаик, Ситаканта (2014). «Обзор генной регуляторной сети, контролирующей развитие трихом у модельного растения Arabidopsis» . Границы в науке о растениях . 5 : 259. дои : 10.3389/fpls.2014.00259 . ISSN 1664-462X . ПМК 4071814 . ПМИД 25018756 .
- ^ Дэвис, штат Пенсильвания ; Хейвуд, В.Х. (1963). Принципы систематики покрытосеменных . Принстон, Нью-Джерси: Ван Ностранпейдж. п. 154.
- ^ Бхатия, RYP; Рагхаван, С.; Рао, КВС; Прасад, В.Н. (1973). «Судебно-медицинская экспертиза листьев и фрагментов листьев в свежем и сушеном состоянии». Журнал Общества судебно-медицинской экспертизы . 13 (3): 183–190. дои : 10.1016/S0015-7368(73)70794-5 . ПМИД 4774149 .
- ^ Управление ООН по наркотикам и преступности (2009 г.). Рекомендуемые методы идентификации и анализа каннабиса и продуктов каннабиса (пересмотренные и обновленные) . Нью-Йорк: Организация Объединенных Наций. стр. 30–32. ISBN 9789211482423 .
- ^ Майтур, Джайдип; Чуа, Нам-Хай (апрель 2000 г.). «Стабилизация микротрубочек приводит к переориентации роста трихом арабидопсиса» (PDF) . Растительная клетка . 12 (4): 465–477. дои : 10.1105/tpc.12.4.465 . ПМК 139846 . ПМИД 10760237 .
- ^ Хан, Флориан; Мантегацца, Отон; Грейнер, Андре; Хегеманн, Питер; Эйзенхут, Мэрион; Вебер, Андреас П.М. (2017). «Эффективный визуальный контроль активности CRISPR/Cas9 у Arabidopsis thaliana» . Границы в науке о растениях . 8:39 . дои : 10.3389/fpls.2017.00039 . ISSN 1664-462X . ПМК 5258748 . ПМИД 28174584 .
- ^ Хан, Флориан; Эйзенхут, Мэрион; Мантегацца, Отон; Вебер, Андреас П.М. (5 апреля 2018 г.). «Гомологическое восстановление дефектного голого гена у арабидопсиса с помощью нацеливания на ген на основе Cas9» . Границы в науке о растениях . 9 : 424. doi : 10.3389/fpls.2018.00424 . ПМЦ 5895730 . ПМИД 29675030 .
- ^ Хюльскамп, М.; Шнитгер, А.; Фолкерс, У. (1999). «Формирование паттерна и дифференциация клеток: трихомы Arabidopsis как генетическая модельная система». Международный обзор цитологии . 186 : 147–178. дои : 10.1016/S0074-7696(08)61053-0 . ISBN 9780123645906 . ISSN 0074-7696 . ПМИД 9770299 .
- ^ Шиндлер, М.В.; Хейнс, К.Ф.; Поттер, МФ; Кукуруза, РМ; Лаудон, К. (2013). «Поимка постельных клопов трихомами листьев вдохновляет на создание биомиметических поверхностей» . Журнал интерфейса Королевского общества . 10 (83): 20130174. doi : 10.1098/rsif.2013.0174 . ISSN 1742-5662 . ПМЦ 3645427 . ПМИД 23576783 .
- ^ Эльц, Томас; Кюттнер, Дженнифер; Лунау, Клаус; Толлриан, Ральф (6 января 2015 г.). «Секреты растений предотвращают паразитизм ос в гнездах пчел-чесальщиков шерсти, что имеет значение для разнообразия материалов для гнезд у Megachilidae» . Границы экологии и эволюции . 2 . дои : 10.3389/fevo.2014.00086 .
- ^ Карли, А (2016). «Использование свойств физической защиты для защиты сельскохозяйственных культур: трихомы листьев Rubus idaeus оказывают сдерживающее воздействие на паутинного клеща, но не на тлю». Анналы прикладной биологии . 168 (2): 159–172. doi : 10.1111/aab.12252 – через Academic Search Premier.
- ^ Кардосо, Марсио З. (2008). «Экология, поведение и биномика: обращение травоядных с трихомами растений: случай Heliconius Charithonia (L.) (Lepidoptera: Nymphalidae) и Passiflora Lobata (Kilip) Hutch. (Passifloraceae)» . Неотропическая энтомология . 37 (3): 247–52. дои : 10.1590/s1519-566x2008000300002 . ПМИД 18641894 .
- ^ Фу, HY; Чен, С.Дж.; Куо-Хуанг, LL (2003). «Сравнительное исследование жалящих трихом и некоторых связанных с ними эпидермальных структур в листьях Dendrocnide meyeniana, Girardinia diversifolia и Urtica thunbergiana». Тайваньия . 48 (4): 213–223.
Библиография
[ редактировать ]- Исав, К. 1965. Анатомия растений , 2-е издание. Джон Уайли и сыновья. 767 стр.
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| растений Группы | |||||||||||
| Анатомия растений |
| ||||||||||
| Физиология растений Материалы | |||||||||||
| Рост растений и привычка | |||||||||||
| Воспроизведение | |||||||||||
| Таксономия растений | |||||||||||
| Упражняться | |||||||||||
| |||||||||||