Протеаза
Протеаза ферментом (также называемая пептидазой , протеиназой или протеолитическим ) [ 1 ] представляет собой фермент , который катализирует протеолиз , расщепляя белки на более мелкие полипептиды или отдельные аминокислоты и стимулируя образование новых белковых продуктов. [ 2 ] Они делают это, расщепляя пептидные связи внутри белков путем гидролиза – реакции, при которой вода разрывает связи . Протеазы участвуют во многих биологических путях, включая переваривание потребляемых белков, катаболизм белков (расщепление старых белков), [ 3 ] [ 4 ] и клеточная сигнализация .
В отсутствие функциональных ускорителей протеолиз будет очень медленным и займет сотни лет . [ 5 ] Протеазы можно найти во всех формах жизни и вирусах . независимо Они неоднократно эволюционировали , и разные классы протеаз могут выполнять одну и ту же реакцию с помощью совершенно разных каталитических механизмов .
Классификация
[ редактировать ]На основе каталитического остатка
[ редактировать ]Протеазы можно разделить на семь широких групп: [ 6 ]
- Сериновые протеазы - с использованием серинового спирта
- Цистеиновые протеазы - с использованием цистеина. тиола
- Треониновые протеазы - с использованием треонина . вторичного спирта
- Аспарагиновые протеазы - с использованием аспартаткарбоновой кислоты.
- Глутаминовые протеазы - с использованием глутаматкарбоновой кислоты.
- Металлопротеазы – с использованием металла , обычно цинка. [ 3 ] [ 4 ]
- Аспарагиновые пептидлиазы - использование аспарагина для проведения реакции элиминирования (не требующей воды)
Впервые протеазы были сгруппированы в 84 семейства в соответствии с их эволюционным родством в 1993 году и классифицированы по четырем каталитическим типам: сериновые , цистеиновые , аспарагиновые и металлопротеазы . [ 7 ] Треониновые протеазы не были описаны до 1995 и 2004 и глутаминовые годов соответственно. Механизм, используемый для расщепления пептидной связи, включает в себя превращение аминокислотного остатка, содержащего цистеин и треонин (протеазы) или молекулу воды (аспарагиновая, глутаминовая и металлопротеазы), в нуклеофильный характер, чтобы он мог атаковать карбонильную группу пептида. Одним из способов создания нуклеофила является каталитическая триада , где остаток гистидина используется для активации серина , цистеина или треонина в качестве нуклеофила. Однако это не эволюционная группировка, поскольку типы нуклеофилов эволюционировали конвергентно в разных суперсемействах , а некоторые суперсемейства демонстрируют дивергентную эволюцию с образованием множества разных нуклеофилов. Металлопротеазы, аспарагиновые и глутаминовые протеазы используют остатки своего активного центра для активации молекулы воды, которая затем атакует разрезаемую связь. [ 8 ]
Пептидные лиазы
[ редактировать ]Седьмой каталитический тип протеолитических ферментов, аспарагин-пептидлиаза , был описан в 2011 году. Его протеолитический механизм необычен, поскольку вместо гидролиза он осуществляет реакцию элиминирования . [ 9 ] Во время этой реакции каталитический аспарагин образует циклическую химическую структуру, которая при правильных условиях расщепляется по остаткам аспарагина в белках. Учитывая его принципиально иной механизм, его включение в качестве пептидазы может быть дискуссионным. [ 9 ]
На основе эволюционной филогении
[ редактировать ]Современная классификация эволюционных суперсемейств протеаз содержится в базе данных MEROPS. [ 10 ] В этой базе данных протеазы классифицируются в первую очередь по «кланам» ( суперсемействам ) на основе структуры, механизма и порядка каталитических остатков (например, клан PA , где P указывает на смесь семейств нуклеофилов). Внутри каждого «клана» протеазы классифицируются на семейства на основе сходства последовательностей (например, семейства S1 и C3 в клане PA). Каждое семейство может содержать многие сотни родственных протеаз (например , трипсин , эластаза , тромбин и стрептогризин внутри семейства S1).
В настоящее время известно более 50 кланов, каждый из которых указывает на независимое эволюционное происхождение протеолиза. [ 10 ]
На основе оптимального pH
[ редактировать ]Альтернативно, протеазы можно классифицировать по оптимальному pH , при котором они активны:
- Кислые протеазы
- Нейтральные протеазы, участвующие в гиперчувствительности 1 типа . Здесь он высвобождается тучными клетками и вызывает активацию комплемента и кининов . [ 11 ] В эту группу входят кальпаины .
- Основные протеазы (или щелочные протеазы )
Ферментативная функция и механизм
[ редактировать ]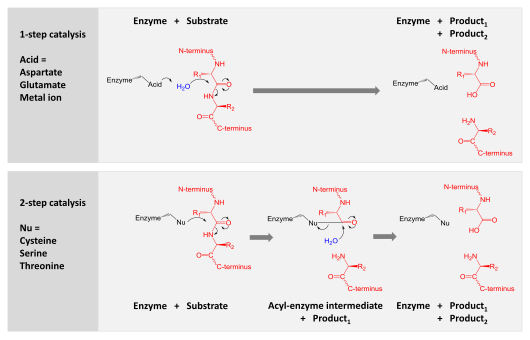
Протеазы участвуют в расщеплении длинных белковых цепей на более короткие фрагменты путем расщепления пептидных связей , связывающих аминокислотные остатки. Некоторые отсоединяют концевые аминокислоты от белковой цепи ( экзопептидазы , такие как аминопептидазы , карбоксипептидаза А ); другие атакуют внутренние пептидные связи белка ( эндопептидазы , такие как трипсин , химотрипсин , пепсин , папаин , эластаза ).
Катализ
[ редактировать ]Катализ осуществляется по одному из двух механизмов:
- Аспарагиновая, глутаминовая и металлопротеазы активируют молекулу воды, которая осуществляет нуклеофильную атаку на пептидную связь, гидролизуя ее.
- Сериновые, треониновые и цистеиновые протеазы используют нуклеофильный остаток (обычно в каталитической триаде ). Этот остаток осуществляет нуклеофильную атаку, ковалентно связывая протеазу с белком-субстратом, высвобождая первую половину продукта. Этот ковалентный промежуточный продукт ацил-фермента затем гидролизуется активированной водой для завершения катализа путем высвобождения второй половины продукта и регенерации свободного фермента.
Специфика
[ редактировать ]Протеолиз может быть очень беспорядочным , так что гидролизуется широкий спектр белковых субстратов. Это относится к пищеварительным ферментам, таким как трипсин , которые должны быть способны расщеплять массив потребляемых белков на более мелкие пептидные фрагменты. Неразборчивые протеазы обычно связываются с одной аминокислотой на субстрате и поэтому обладают специфичностью только к этому остатку. Например, трипсин специфичен для последовательностей ...K\... или ...R\... ('\'=сайт расщепления). [ 12 ]
И наоборот, некоторые протеазы высокоспецифичны и расщепляют субстраты только с определенной последовательностью. Свертывание крови (например, тромбин ) и обработка вирусных полипротеинов (например, протеаза TEV ) требуют такого уровня специфичности для достижения точных событий расщепления. Это достигается за счет протеаз, имеющих длинную связывающую щель или туннель с несколькими карманами, которые связываются с указанными остатками. Например, протеаза TEV специфична для последовательности ...ENLYFQ\S... ('\'=сайт расщепления). [ 13 ]
Деградация и аутолиз
[ редактировать ]Протеазы, будучи сами белками, расщепляются другими молекулами протеаз, иногда того же типа. Это действует как метод регуляции активности протеазы. Некоторые протеазы менее активны после аутолиза (например, протеаза TEV ), тогда как другие более активны (например, трипсиноген ).
Биоразнообразие протеаз
[ редактировать ]Протеазы встречаются во всех организмах, от прокариот до эукариот и вирусов . Эти ферменты участвуют во множестве физиологических реакций, от простого переваривания пищевых белков до высокорегулируемых каскадов (например, процессов свертывания крови) . каскад , система комплемента , пути апоптоза и каскад, активирующий профенолоксидазу беспозвоночных). Протеазы могут либо разрывать специфические пептидные связи ( ограниченный протеолиз ), в зависимости от аминокислотной последовательности белка, либо полностью расщеплять пептид до аминокислот ( неограниченный протеолиз ). Активность может представлять собой деструктивное изменение (отмена функции белка или расщепление его на основные компоненты), активация функции или сигнал в сигнальном пути.
Растения
[ редактировать ]Геномы растений кодируют сотни протеаз, функции которых в основном неизвестны. Те, функция которых известна, в значительной степени участвуют в регуляции развития . [ 14 ] Растительные протеазы также играют роль в регуляции фотосинтеза . [ 15 ]
Животные
[ редактировать ]Протеазы используются в организме для различных метаболических процессов. Кислые протеазы, секретируемые в желудке (например, пепсин ), и сериновые протеазы, присутствующие в двенадцатиперстной кишке ( трипсин и химотрипсин ), позволяют нам переваривать белок пищи. Присутствующие в сыворотке крови протеазы ( тромбин , плазмин , фактор Хагемана и др.) играют важную роль в свертывании крови, а также лизисе тромбов, правильном действии иммунной системы. Другие протеазы присутствуют в лейкоцитах ( эластаза , катепсин G ) и играют несколько различных ролей в метаболическом контроле. Некоторые змеиные яды также являются протеазами, например, гадюки гемотоксин , и мешают каскаду свертывания крови жертвы. Протеазы определяют время жизни других белков, играющих важную физиологическую роль, таких как гормоны, антитела или другие ферменты. Это один из самых быстрых «включающихся» и «выключающихся» регуляторных механизмов в физиологии организма.
Благодаря сложному совместному действию протеазы могут катализировать каскадные реакции, которые приводят к быстрому и эффективному усилению ответа организма на физиологический сигнал.
Бактерии
[ редактировать ]Бактерии секретируют протеазы для гидролиза пептидных связей в белках и, следовательно, расщепления белков на составляющие их аминокислоты . Бактериальные и грибковые протеазы особенно важны для глобальных циклов углерода и азота при переработке белков, и такая активность, как правило, регулируется сигналами питания в этих организмах. [ 16 ] Чистое влияние пищевой регуляции активности протеаз среди тысяч видов, присутствующих в почве, можно наблюдать на уровне общего микробного сообщества, поскольку белки расщепляются в ответ на ограничение углерода, азота или серы. [ 17 ]
ААА+ Бактерии содержат протеазы, ответственные за общий контроль качества белков (например, протеасома ), разрушая развернутые или неправильно свернутые белки .
Секретируемая бактериальная протеаза также может действовать как экзотоксин и быть примером фактора вирулентности в бактериальном патогенезе (например, эксфолиативный токсин ). Бактериальные экзотоксические протеазы разрушают внеклеточные структуры.
Вирусы
[ редактировать ]Геномы некоторых вирусов кодируют один массивный полипротеин , для расщепления которого на функциональные единицы требуется протеаза (например, вирус гепатита С и пикорнавирусы ). [ 18 ] Эти протеазы (например, протеаза TEV ) обладают высокой специфичностью и расщепляют лишь очень ограниченный набор последовательностей субстрата. Поэтому они являются общей мишенью для ингибиторов протеазы . [ 19 ] [ 20 ]
Архея
[ редактировать ]Археи используют протеазы для регулирования различных клеточных процессов, включая передачу сигналов клетками , метаболизм , секрецию и контроль качества белка. [ 21 ] [ 22 ] У архей обнаружены только две АТФ-зависимые протеазы: мембраносвязанная протеаза LonB и растворимый 20S протеосомный комплекс. [ 21 ]
Использование
[ редактировать ]Область исследований протеаз огромна. около 8000 статей в этой области. С 2004 года ежегодно публикуется [ 23 ] Протеазы используются в промышленности, медицине и в качестве основного инструмента биологических исследований. [ 24 ] [ 25 ]
Пищеварительные протеазы входят в состав многих стиральных порошков , а также широко используются в хлебной промышленности в качестве улучшителя хлеба . В медицине используются различные протеазы как для их естественной функции (например, контроль свертывания крови), так и для полностью искусственных функций ( например , для целенаправленной деградации патогенных белков). Высокоспецифичные протеазы, такие как протеаза TEV и тромбин, обычно используются для расщепления слитых белков и аффинных меток контролируемого . Содержащие протеазу растительные растворы, называемые вегетарианским сычужным ферментом, уже сотни лет используются в Европе и на Ближнем Востоке для изготовления кошерных и халяльных сыров . Вегетарианский сычужный фермент Withania coagulans использовался на протяжении тысячелетий в качестве аюрведического средства для улучшения пищеварения и лечения диабета на Индийском субконтиненте. Его также используют для приготовления панира .
Ингибиторы
[ редактировать ]Активность протеаз ингибируется ингибиторами протеаз . [ 26 ] Одним из примеров ингибиторов протеаз является суперсемейство серпинов . В его состав входят альфа-1-антитрипсин (защищающий организм от чрезмерного воздействия собственных воспалительных протеаз), альфа-1-антихимотрипсин (который действует аналогично), С1-ингибитор (защищающий организм от чрезмерной активации собственной системы комплемента , вызванной протеазами). ), антитромбин (который защищает организм от чрезмерной коагуляции ), ингибитор активатора плазминогена-1 (который защищает организм от неадекватной коагуляции путем блокирования фибринолиза, запускаемого протеазой ) и нейросерпина . [ 27 ]
Природные ингибиторы протеаз включают семейство белков липокалинов , которые играют роль в регуляции и дифференцировке клеток. липофильные Было обнаружено, что лиганды, прикрепленные к белкам липокалина, обладают свойствами ингибирования опухолевых протеаз. Природные ингибиторы протеазы не следует путать с ингибиторами протеазы, используемыми в антиретровирусной терапии. Некоторые вирусы , в том числе ВИЧ/СПИД , в своем репродуктивном цикле зависят от протеаз. Таким образом, ингибиторы протеаз разрабатываются как противовирусные терапевтические средства.
Другие природные ингибиторы протеаз используются в качестве защитных механизмов. Распространенными примерами являются ингибиторы трипсина, обнаруженные в семенах некоторых растений, наиболее примечательными для людей являются соевые бобы, основная продовольственная культура, где они действуют, отпугивая хищников. Сырые соевые бобы токсичны для многих животных, включая человека, пока содержащиеся в них ингибиторы протеазы не будут денатурированы.
См. также
[ редактировать ]- Лигаза
- Протеаза
- Клан ПА
- Конвергентная эволюция
- Протеолиз
- Каталитическая триада
- Карта протеолиза
- Протеазы в ангиогенезе
- Внутримембранные протеазы
- Ингибитор протеазы (фармакология)
- Ингибитор протеазы (биология)
- TopFIND - база данных специфичности протеаз, субстратов, продуктов и ингибиторов
- MEROPS - База данных эволюционных групп протеаз
Ссылки
[ редактировать ]- ^ «Протеолитический фермент | Описание, типы и функции | Британника» .
- ^ Лопес-Отин С., Bond JS (ноябрь 2008 г.). «Протеазы: многофункциональные ферменты в жизни и болезни» . Журнал биологической химии . 283 (45): 30433–30437. дои : 10.1074/jbc.R800035200 . ПМЦ 2576539 . ПМИД 18650443 .
- ^ Jump up to: а б Кинг СП, Лян В.Г., Шерпельц К.П., Шиллинг А.Б., Мередит С.К., Тан В.Дж. (июль 2014 г.). «Молекулярные основы распознавания и деградации субстратов протеазой препоследовательности человека» . Структура . 22 (7): 996–1007. doi : 10.1016/j.str.2014.05.003 . ПМК 4128088 . ПМИД 24931469 .
- ^ Jump up to: а б Шен Ю, Иоахимиак А, Рознер М.Р., Тан В.Дж. (октябрь 2006 г.). «Структуры человеческого фермента, расщепляющего инсулин, открывают новый механизм распознавания субстрата» . Природа . 443 (7113): 870–874. Бибкод : 2006Natur.443..870S . дои : 10.1038/nature05143 . ПМЦ 3366509 . ПМИД 17051221 .
- ^ Радзичка А., Вольфенден Р. (июль 1996 г.). «Скорость некаталитического гидролиза пептидных связей в нейтральном растворе и сродство протеаз к переходному состоянию». Журнал Американского химического общества . 118 (26): 6105–6109. дои : 10.1021/ja954077c .
Чтобы оценить относительную эффективность ферментов, катализирующих гидролиз внутренних и С-концевых пептидных связей [...]
- ^ Ода К. (январь 2012 г.). «Новые семейства карбоксилпептидаз: серин-карбоксилпептидазы и глутаминовые пептидазы» . Журнал биохимии . 151 (1): 13–25. дои : 10.1093/jb/mvr129 . ПМИД 22016395 .
- ^ Роулингс Н.Д., Барретт А.Дж. (февраль 1993 г.). «Эволюционные семейства пептидаз» . Биохимический журнал . 290 (Часть 1): 205–218. дои : 10.1042/bj2900205 . ПМЦ 1132403 . ПМИД 8439290 .
- ^ Санман, Лаура Э. (июнь 2014 г.). «Профилирование протеаз на основе активности» . Ежегодный обзор биохимии . 83 : 249–273. doi : 10.1146/annurev-biochem-060713-035352 . ПМИД 24905783 .
- ^ Jump up to: а б Роулингс Н.Д., Барретт А.Дж., Бейтман А. (ноябрь 2011 г.). «Аспарагиновые пептидлиазы: седьмой каталитический тип протеолитических ферментов» . Журнал биологической химии . 286 (44): 38321–38328. дои : 10.1074/jbc.M111.260026 . ПМК 3207474 . ПМИД 21832066 .
- ^ Jump up to: а б Роулингс Н.Д., Барретт А.Дж., Бейтман А. (январь 2010 г.). «МЕРОПС: база данных пептидаз» . Исследования нуклеиновых кислот . 38 (Проблема с базой данных): D227–D233. дои : 10.1093/nar/gkp971 . ПМК 2808883 . ПМИД 19892822 .
- ^ Митчелл Р.С., Кумар В., Аббас А.К., Фаусто Н. (2007). Основная патология Роббинса (8-е изд.). Филадельфия: Сондерс. п. 122. ИСБН 978-1-4160-2973-1 .
- ^ Родригес Дж., Гупта Н., Смит Р.Д., Певзнер П.А. (январь 2008 г.). «Разрезает ли трипсин раньше, чем пролин?». Журнал исследований протеома . 7 (1): 300–305. дои : 10.1021/pr0705035 . ПМИД 18067249 .
- ^ Ренике С., Спадаччини Р., Таксис С. (24 июня 2013 г.). «Протеаза вируса травления табака с повышенной толерантностью к субстрату в положении P1'» . ПЛОС ОДИН . 8 (6): e67915. Бибкод : 2013PLoSO...867915R . дои : 10.1371/journal.pone.0067915 . ПМК 3691164 . ПМИД 23826349 .
- ^ ван дер Хорн Р.А. (2008). «Растительные протеазы: от фенотипов к молекулярным механизмам». Ежегодный обзор биологии растений . 59 : 191–223. doi : 10.1146/annurev.arplant.59.032607.092835 . hdl : 11858/00-001M-0000-0012-37C7-9 . ПМИД 18257708 .
- ^ Зелиско А., Джековский Г. (октябрь 2004 г.). «Зависимая от старения деградация Lhcb3 опосредуется тилакоидной мембраносвязанной протеазой». Журнал физиологии растений . 161 (10): 1157–1170. дои : 10.1016/j.jplph.2004.01.006 . ПМИД 15535125 .
- ^ Симс ГК (2006). «Азотное голодание способствует биодеградации N-гетероциклических соединений в почве» . Биология и биохимия почвы . 38 (8): 2478–2480. doi : 10.1016/j.soilbio.2006.01.006 . Архивировано из оригинала 28 апреля 2021 г. Проверено 29 декабря 2018 г.
- ^ Симс ГК, Вандер ММ (2002). «Протеолитическая активность при ограничении азота или серы». Прил. Почвенный Экол . 568 (3): 1–5. Бибкод : 2002AppSE..19..217S . дои : 10.1016/S0929-1393(01)00192-5 .
- ^ Тонг Л. (декабрь 2002 г.). «Вирусные протеазы». Химические обзоры . 102 (12): 4609–4626. дои : 10.1021/cr010184f . ПМИД 12475203 .
- ^ Скореньский М, Сеньчик М (2013). «Вирусные протеазы как мишени для разработки лекарств». Текущий фармацевтический дизайн . 19 (6): 1126–1153. дои : 10.2174/13816128130613 . ПМИД 23016690 .
- ^ Курт Йилмаз Н., Суонстром Р., Шиффер, Калифорния (июль 2016 г.). «Улучшение ингибиторов вирусных протеаз для борьбы с лекарственной устойчивостью» . Тенденции в микробиологии . 24 (7): 547–557. дои : 10.1016/j.tim.2016.03.010 . ПМЦ 4912444 . ПМИД 27090931 .
- ^ Jump up to: а б Хименес М.И., Черлетти М., Де Кастро Р.Э. (2015). «Архейные мембраносвязанные протеазы: сведения о Haloferax volcanii и других галоархей» . Границы микробиологии . 6:39 . дои : 10.3389/fmicb.2015.00039 . ПМЦ 4343526 . ПМИД 25774151 .
- ^ Мопен-Ферлоу, Дж. А. (декабрь 2018 г.). Робинсон Н.П. (ред.). «Протеолитические системы архей: нарезка, нарезка кубиками и измельчение в крайнем случае» . Новые темы в науках о жизни . 2 (4): 561–580. дои : 10.1042/ETLS20180025 . ПМЦ 7497159 . ПМИД 32953999 .
- ^ Барретт А.Дж., Роулингс Н.Д., Весснерд Дж.Ф. (2004). Справочник по протеолитическим ферментам (2-е изд.). Лондон, Великобритания: Elsevier Academic Press. ISBN 978-0-12-079610-6 .
- ^ Хупер Н.М., изд. (2002). Протеазы в биологии и медицине . Лондон: Портленд Пресс. ISBN 978-1-85578-147-4 .
- ^ Фейджу-Сиота Л., Вилла ТГ (28 сентября 2010 г.). «Нативные и биотехнологически модифицированные растительные протеазы с промышленным применением». Пищевые и биотехнологические технологии . 4 (6): 1066–1088. дои : 10.1007/s11947-010-0431-4 . S2CID 84748291 .
- ^ Саутэн С. (июль 2001 г.). «Геномный взгляд на человеческие протеазы как мишени для лекарств». Открытие наркотиков сегодня . 6 (13): 681–688. дои : 10.1016/s1359-6446(01)01793-7 . ПМИД 11427378 .
- ^ Пуэнте XS, Лопес-Отин С (апрель 2004 г.). «Геномный анализ крысиных протеаз и ингибиторов протеаз» . Геномные исследования . 14 (4): 609–622. дои : 10.1101/гр.1946304 . ПМЦ 383305 . ПМИД 15060002 .
Внешние ссылки
[ редактировать ]- Международное общество протеолиза
- MEROPS - база данных пептидаз. Архивировано 14 ноября 2006 г. в Wayback Machine.
- Список ингибиторов протеазы
- Предиктор разрезания протеазы
- Список протеаз и их особенностей (см. также [1] ). Архивировано 30 апреля 2011 г. на Wayback Machine .
- MAP протеолиза от Центра протеолитических путей
- База данных сайта протеолиза Cut Site — тщательно подобранные экспертные аннотации пользователей.
- Графический интерфейс сайтов разреза протеазы
- База данных протеаз TopFIND, охватывающая сайты разрезов, субстраты и концы белков.
- Протеазы Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)