Пептидная связь

В органической химии пептидная связь представляет собой амидного типа ковалентную химическую связь , соединяющую две последовательные альфа-аминокислоты от C1 ( углерод номер один) одной альфа-аминокислоты и N2 ( азот номер два) другой вдоль пептида или белка . цепь. [1]
Ее также можно назвать эупептидной связью. [1] чтобы отличить ее от изопептидной связи , которая представляет собой другой тип амидной связи между двумя аминокислотами.
Синтез
[ редактировать ]
Когда две аминокислоты образуют дипептид через пептидную связь , [1] это разновидность реакции конденсации . [2] При таком типе конденсации две аминокислоты сближаются друг с другом, при этом небоковая ( C1) карбоновой кислоты часть одной приближается к небоковой аминогруппе (N2) другой . Один теряет водород и кислород из своей карбоксильной группы (СООН), а другой теряет водород из своей аминогруппы (NH 2 ). В результате этой реакции образуется молекула воды (H 2 O) и две аминокислоты, соединенные пептидной связью (-CO-NH-). Две соединенные аминокислоты называются дипептидом.
Амидная связь синтезируется, когда карбоксильная группа одной молекулы аминокислоты реагирует с аминогруппой другой молекулы аминокислоты, вызывая высвобождение молекулы воды (H 2 O), следовательно, процесс представляет собой реакцию синтеза дегидратации .
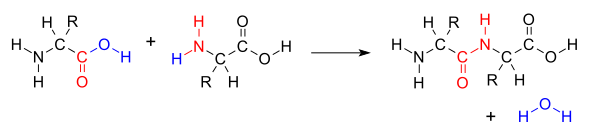
На образование пептидной связи расходуется энергия, которая в организмах получается из АТФ . [3] Пептиды и белки представляют собой цепочки аминокислот , скрепленные пептидными связями (а иногда и несколькими изопептидными связями ). Организмы используют ферменты для производства нерибосомальных пептидов . [4] и рибосомы для производства белков посредством реакций, которые в деталях отличаются от синтеза дегидратации. [5]
Некоторые пептиды, такие как альфа-аманитин , называются рибосомальными пептидами, поскольку они производятся рибосомами. [6] но многие из них являются нерибосомальными пептидами , поскольку они синтезируются специализированными ферментами, а не рибосомами. Например, трипептид глутатион синтезируется в два этапа из свободных аминокислот двумя ферментами : глутамат-цистеиновой лигазой (образует изопептидную связь , которая не является пептидной связью) и глутатионсинтетазой (образует пептидную связь). [7] [8]
Деградация
[ редактировать ]Пептидная связь может быть разорвана гидролизом ( присоединением воды). При гидролизе пептидных связей в воде выделяется 8–16 кДж / моль (2–4 ккал / моль ) энергии Гиббса . [9] Этот процесс чрезвычайно медленный: период полураспада при 25 ° C составляет от 350 до 600 лет на одну связь. [10]
В живых организмах этот процесс обычно катализируется ферментами , известными как пептидазы или протеазы , хотя есть сообщения о гидролизе пептидных связей, вызванном конформационным напряжением, когда пептид/белок сворачивается в нативную структуру. [11] Таким образом, этот неферментативный процесс ускоряется не стабилизацией переходного состояния, а скорее дестабилизацией основного состояния.
Спектры
[ редактировать ]Длина волны поглощения пептидной связи составляет 190–230 нм, [12] что делает его особенно восприимчивым к ультрафиолетовому излучению.
Цис/транс-изомеры пептидной группы
[ редактировать ]Значительная делокализация неподеленной пары электронов на атоме азота придает группе характер частичной двойной связи . Частичная двойная связь делает амидную группу плоской и встречается либо в цис-, либо в транс-изомерах . В развернутом состоянии белков пептидные группы могут изомеризоваться и принимать оба изомера; однако в свернутом состоянии в каждом положении принимается только один изомер (за редкими исключениями). Транс-форма является преимущественно предпочтительной для большинства пептидных связей (соотношение примерно 1000:1 в популяциях транс:цис). Однако пептидные группы X-Pro имеют тенденцию иметь соотношение примерно 30:1, предположительно из-за симметрии между C а и С д Атомы пролина делают цис- и транс-изомеры почти равными по энергии, как показано на рисунке ниже.
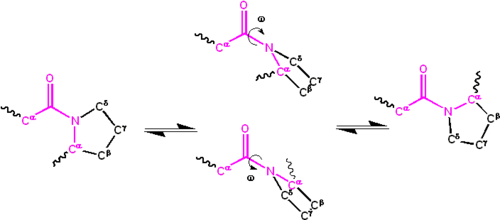
Двугранный угол, связанный с пептидной группой (определяемый четырьмя атомами C а –C'–N–C а ) обозначается ; для цис-изомера ( синперипланарная конформация) и для транс-изомера ( антиперипланарная конформация). Амидные группы могут изомеризоваться по связи C'–N между цис- и транс-формами, хотя и медленно ( секунд при комнатной температуре). Переходные состояния требует частичного разрыва двойной связи, чтобы энергия активации составляла примерно 80 кДж/моль (20 ккал/моль). Однако энергия активации изомеризация может быть снижена (и катализирована ) за счет изменений, которые благоприятствуют форме с одинарной связью, таких как помещение пептидной группы в гидрофобное окружение или передача водородной связи атому азота пептидной группы X-Pro. . Оба эти механизма снижения энергии активации наблюдались в пептидилпролилизомеразах (PPIases), которые представляют собой встречающиеся в природе ферменты, катализирующие цис-транс-изомеризацию пептидных связей X-Pro.





Конформационное сворачивание белка обычно происходит намного быстрее (обычно 10–100 мс), чем цис-транс-изомеризация (10–100 с). Ненативный изомер некоторых пептидных групп может значительно нарушить конформационное сворачивание, либо замедляя его, либо предотвращая его даже до тех пор, пока не будет достигнут нативный изомер. Однако не все пептидные группы оказывают одинаковое влияние на сворачивание; ненативные изомеры других пептидных групп могут вообще не влиять на сворачивание.
Химические реакции
[ редактировать ]Благодаря своей резонансной стабилизации пептидная связь относительно нереактивна в физиологических условиях, даже в меньшей степени, чем аналогичные соединения, такие как сложные эфиры . Тем не менее, пептидные связи могут вступать в химические реакции, обычно посредством атаки электроотрицательного атома на карбонильный углерод , разрывая двойную карбонильную связь и образуя тетраэдрический промежуточный продукт. Это путь, которым следуют при протеолизе и, в более общем смысле, в реакциях обмена N–O-ацилов, таких как реакции интеинов . Когда функциональная группа, атакующая пептидную связь, представляет собой тиол , гидроксил или амин , полученную молекулу можно назвать циклолом или, более конкретно, тиациклолом, оксациклолом или азациклолом соответственно.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с «Номенклатура и символика аминокислот и пептидов. Рекомендации 1983 г.» . Европейский журнал биохимии . 138 (1): 9–37. 1984. doi : 10.1111/j.1432-1033.1984.tb07877.x . ISSN 0014-2956 . ПМИД 6692818 .
- ^ Мюллер, П. (1 января 1994 г.). «Словарь терминов, используемых в физической органической химии (Рекомендации ИЮПАК, 1994 г.)» . Чистая и прикладная химия . 66 (5): 1077–1184. дои : 10.1351/pac199466051077 . ISSN 1365-3075 . S2CID 195819485 .
- ^ Уотсон, Джеймс; Хопкинс, Нэнси; Робертс, Джеффри; Агецингер Штайц, Джоан; Вайнер, Алан (1987) [1965]. Молекулярная биология гена (твердый переплет) (Четвертое изд.). Менло-Парк, Калифорния: The Benjamin/Cummings Publishing Company, Inc. 168 . ISBN 978-0-8053-9614-0 .
- ^ Миллер БР; Гулик А.М. (2016). «Структурная биология нерибосомальных пептидсинтетаз». Биосинтез нерибосомальных пептидов и поликетидов . Методы молекулярной биологии. Том. 1401. стр. 3–29. дои : 10.1007/978-1-4939-3375-4_1 . ISBN 978-1-4939-3373-0 . ПМК 4760355 . ПМИД 26831698 .
- ^ Гриффитс Эй Джей; Миллер Дж. Х.; Сузуки ДТ; Левонтин РЦ; Гелбарт В.М. (2000). Синтез белка (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Уолтон Джей Ди; Халлен-Адамс HE; Ло Х. (2010). «Рибосомальный биосинтез циклических пептидных токсинов грибов мухоморов» . Биополимеры . 94 (5): 659–664. дои : 10.1002/bip.21416 . ПМЦ 4001729 . ПМИД 20564017 .
- ^ У Г.; Клык YZ; Ян С.; Луптон-младший; Тернер Н.Д. (март 2004 г.). «Метаболизм глутатиона и его значение для здоровья» . Журнал питания . 134 (3): 489–492. дои : 10.1093/jn/134.3.489 . ПМИД 14988435 .
- ^ Мейстер А. (ноябрь 1988 г.). «Метаболизм глутатиона и его селективная модификация» . Журнал биологической химии . 263 (33): 17205–17208. дои : 10.1016/S0021-9258(19)77815-6 . ПМИД 3053703 .
- ^ Мартин Р.Б. (декабрь 1998 г.). «Свободные энергии и равновесия гидролиза и образования пептидных связей». Биополимеры . 45 (5): 351–353. doi : 10.1002/(SICI)1097-0282(19980415)45:5<351::AID-BIP3>3.0.CO;2-K .
- ^ Радзичка, Анна; Вулфенден, Ричард (1 января 1996 г.). «Скорость некаталитического гидролиза пептидных связей в нейтральном растворе и сродство протеаз к переходному состоянию». Журнал Американского химического общества . 118 (26): 6105–6109. дои : 10.1021/ja954077c . ISSN 0002-7863 .
- ^ Сандберг А.; Йоханссон Д.Г.; Макао Б.; Херд Т. (апрель 2008 г.). «Аутопротеолиз домена SEA, ускоренный конформационным напряжением: энергетические аспекты». Журнал молекулярной биологии . 377 (4): 1117–1129. дои : 10.1016/j.jmb.2008.01.051 . ПМИД 18308334 .
- ^ Гольдфарб А.Р.; Сайдел Л.Дж.; Мосович Э. (ноябрь 1951 г.). «УФ-спектры поглощения белков» . Журнал биологической химии . 193 (1): 397–404. дои : 10.1016/S0021-9258(19)52465-6 . ПМИД 14907727 .


| Базы данных органов управления : Национальные |
|---|