Глюкозамин-фосфат N-ацетилтрансфераза
| Глюкозамин 6-фосфат N-ацетилтрансфераза | |||
|---|---|---|---|
 Глюкозамин 6-фосфат N-ацетилтрансфераза 1, гомодимер, человек | |||
| Идентификаторы | |||
| ЕС №. | 2.3.1.4 | ||
| CAS №. | 9031-91-8 | ||
| Базы данных | |||
| Intenz | Intenz View | ||
| Бренда | Бренда вход | ||
| Расширение | Вид Nicezyme | ||
| Кегг | Кегг вход | ||
| Метатический | Метаболический путь | ||
| Напрямую | профиль | ||
| PDB Структуры | RCSB PDB PDBE PDBSUM | ||
| Джин Онтология | Друг / Quickgo | ||
| |||
При фермете , глюкозамин-фосфат N-ацетилтрансфераза (GNA) ( 2.3.1.4 ) представляет собой фермент который катализирует перенос ацетильной группы из -CoA первично ацетил EC к -ацетил-D-глюкозамин-6-фосфат . [ 1 ]
Этот фермент принадлежит к семейству трансфераз , группе ферментов, которая переносит очень специфическую функциональную группу, в данном случае ацетил, от донора в рецептор. В частности, этот фермент можно охарактеризовать как часть семейства ацилтрансфераз , поскольку он включает перенос общей ацильной группы с метилом в качестве заместителя.
Номенклатура
[ редактировать ]Систематическим названием этого класса ферментов является ацетил-КоА: D-глюкозамин-6-фосфат N-ацетилтрансфераза. Другие названия в общем использовании включают фосфоглюкозамин-трансацетилазу, фосфоглукозамин ацетилазу, глюкозамин-6-фосфат ацетилазу, D-глюкозамин-6-P N-ацетилтрансферазу, ацинодезоксисфосфосфат-ацетилтрансфера, гликоза ацетилазо-ацетилазосфона -ацетилглюкозамин -6-фосфатная синтаза, фосфоглукозамин N-ацетилаза, глюкозамин-фосфат N-ацетилтрансфераза и глюкозамин-6-фосфат N-ацетилтрансфераза.
Функция
[ редактировать ]Этот фермент является частью пути биосинтеза гексозамина [ 2 ] (HBP), который является одним из путей обработки глюкозы в общем метаболизме. Этот путь разделяет два начальных шага с гликолизом и расходится лишь небольшой части потока глюкозы от этого более традиционного гликолитического пути. Следовательно, он предпочитается, когда существует отрицательная регуляция обратной связи по гликолизу, как в случае большого количества свободных жирных кислот . Конечным продуктом этого пути является UDP-N-ацетилглюкозамин , который участвует в модификации сложных молекул, таких как гликолипиды , протеогликаны [ 3 ] и гликопротеины . Этот конечный продукт действует как носитель N-ацетилглюкозамина , который является мономерной единицей хитина, [ 4 ] раковины ракообразных и насекомых , а также клеточную стенку грибов Структурный полимер, который сочетает в себе . Кроме того, N-ацетилглюкозамин также является единицей пептидогликанового полимера, который сочетает в себе бактерий клеточную стенку [ 5 ] наряду с N-ацетилмурамным дисахаридом.

Более конкретно, фермент GNA катализирует четвертую стадию пути HBP у эукариот , способствуя переносу углерода из ацетил-КоА к другому субстрату, D-глюкозамин-6-фосфату , который, наконец, даст UDP-N-ацетилглукозамин . Это небольшая, но важная химическая стадия, которая имеет решающее значение для свойств подпродуктов этого метаболического пути. Ацетилирование ацетилглюкозамином проводится до самого конечного продукта гексаминового пути, и очень характерно для полимеров, образованных с N- . Например, он является одним из основных различий в молекулярной структуре хитина и целлюлозы , [ 7 ] и объясняет многие физические и химические свойства этих полимеров. Например, в случае хитина вычислительные исследования показали, что ацилирование способствует образованию водородных связей , которые стабилизируют кристаллическую структуру этого полимера, обеспечивая большую устойчивость к разрушению. [ 8 ]
Тем не менее, в прокариотическом метаболизме путь биосинтеза гексозамина следует за другой стадией реакции, на котором другой фермент действует на одни и те же характерные субстраты [ 6 ] (Рисунок 1). В прокариотах перенос фосфата от 6-углерода к 1-углероду происходит перед ацилированием, так что субстратом реакции углерода является глюкозамин-1-фосфатом, а не D-глюкозамин-6-фосфатом. На этот раз фермент, ответственный за ацетилирование, является бифункциональный белок Glmu (N-ацетилглюкозамин-1-фосфат уридилтрансфераза), [ 9 ] Это также катализирует добавление UDP в фосфатную группу на N-ацетил-D-глюкозамин-1-фосфат.
У людей глюкозамин-фосфат N-ацетилтрансфераза представляет собой димер с двумя идентичными субъединицами , [ 10 ] и кодируется в геном gnpnat [ 11 ] (Символ HGNC). Более конкретно, фермент сильно экспрессируется в тканях печени , желудка и желудочно -кишечного тракта, а внутри клетки он расположен в эндосомах и в аппарате Гольджи (путем ручной аннотации). [ 11 ]
Механизм
[ редактировать ]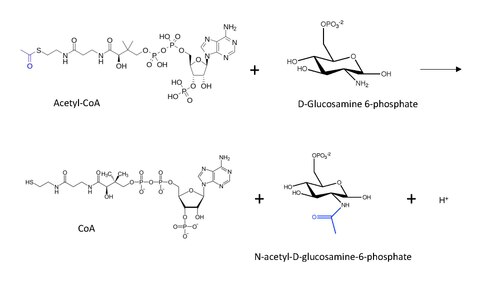
Молекулярная структура реакции, катализируемая GNA, показана ниже, с переносимой ацетильной группой синим.
Общий механизм реакции, постулируемый для ацетилирования белка N-энда (вдохновленного механизмом ацетилирования лизина) с помощью ацетил-КоА включает нуклеофильную атаку аминогрузки (в данном случае от D-глюкозамина-6-фосфата) на терминальный карбонил в углероде Передача, что приводит к образованию углеродного тетраэдрического промежуточного соединения. [ 13 ] Реакция продолжается с восстановлением карбонила , удаляя COA в качестве уходной группы, так что теперь ацетильная группа подключена к аминогруппе в другом субстрате.
В частности, для этих катализаторов N-ацетилтрансферазы, исследования с ферментом S. cerevisiae GNA показали, что некоторые специфические аминокислоты способствуют связыванию субстрата, повышенной нуклеофильности аминогрузки и, наконец, катализом, который подтверждает постулируемый механизм, описанный выше. [ 14 ] Glu98, ASP99 и ILE100 поляризуют карбонильную связь в ацетил-КоА , увеличивая углеродную электрофильность , а также стабилизируя тетраэдрическое промежуточное здание углерода. Tyr143 отвечает за стабилизацию тиолатного аниона, в пользу S-COA в качестве уходной группы из тетраэдрического углерода. Наконец, ASP134 повышает нуклеофильность аминогрузки в D -глюкозамид-6-фосфате за счет пожертвований электронной плотности на атом азота . В другом организме, C. albicans , было обнаружено, что аналогичный набор аминокислот необходим для каталитической активности, [ 15 ] соответственно система GLU88-ASP-89-ILE90, ASP125 и TYR133.
Структура
[ редактировать ]
В конце 2019 года 13 структур для этого класса ферментов были решены к PDB , с кодами доступа 1I12 ( Saccharomyces cerevisiae ), 1i1d ( Saccharomyces cerevisiae 1i21 ( Saccharomyces cerevisiae ) 2huz ( sapien , ) , homo Sapiens ), 4AG7 ( Caenorhabditis elegans ), среди других.
На рисунке 3 показана предложенная кристаллическая структура GNA у людей , [ 17 ] с каждой каталитической субъединицей в другом цвете. Ацетил -КоА, ограниченный ферментом, показан в светло-розовом, а продукт, все еще связанный с каталитическим сайтом, показан в фиолетовом виде. Передаваемая ацетильная группа в N-ацетил-D-глюкозамин-6-фосфатном произведении в фиолетовом виде показана желтым. Эта предложенная трехмерная структура белка показывает , что специфические части субстратов, участвующих в этой реакции - терминальный конец линейной части ацетил -КоА и азота , прикрепленной к глюкозаминовому кольцу - находятся в большой близости.
Ссылки
[ редактировать ]- ^ Kato N, Mueller CR, Wessely V, Lan Q, Christensen BM (июнь 2005 г.). «Москит глюкозамин-6-фосфат N-ацетилтрансфераза: кДНК, структура генов и кинетика фермента». Биохимия насекомых и молекулярная биология . 35 (6): 637–46. doi : 10.1016/j.ibmb.2005.02.005 . PMID 15857769 .
- ^ Schwarzer M, Doenst T (2016). Руководство ученых по метаболизму сердца . Академическая пресса. С. 39–55. ISBN 9780128023945 .
- ^ Ким Й.Х., Накаяма Т., Наяк Дж. (Январь 2018 г.). «Гликолиз и биосинтетический путь гексозамина в качестве новых целей для воспаления верхних и нижних дыхательных путей» . Исследования аллергии, астмы и иммунологии . 10 (1): 6–11. doi : 10.4168/aair.2018.10.1.6 . PMC 5705485 . PMID 29178672 .
- ^ Коэн (октябрь 2001 г.). «Синтез и ингибирование хитина: повторное посещение». Наука по борьбе с вредителями . 57 (10): 946–50. doi : 10.1002/ps.363 . PMID 11695188 .
- ^ Meroueh SO, Bencze KZ, Hesek D, Lee M, Fisher JF, Stemmler TL, Mobashery S (март 2006 г.). «Трехмерная структура пептидогликана бактериальной клеточной стенки» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (12): 4404–9. Bibcode : 2006pnas..103.4404M . doi : 10.1073/pnas.0510182103 . PMC 1450184 . PMID 16537437 .
- ^ Jump up to: а беременный Риглер Х., Хертер Т., Гришковская И., Луде А., Рингаджлло М., Болгер М.Е., Эссигман Б., Усадель Б (апрель 2012 г.). «Кристаллическая структура и функциональная характеристика глюкозамин-6-фосфатной N-ацетилтрансферазы из Arabidopsis thaliana». Биохимический журнал . 443 (2): 427–37. doi : 10.1042/bj20112071 . PMID 22329777 .
- ^ «Рис. 1 Химическая структура целлюлозы и хитина» . Researchgate . Получено 2019-03-15 .
- ^ Cui J, Yu Z, Lau D (январь 2016 г.). «Влияние ацетильной группы на механические свойства нанокристалла хитина/хитозана: исследование молекулярной динамики» . Международный журнал молекулярных наук . 17 (1): 61. doi : 10.3390/ijms17010061 . PMC 4730306 . PMID 26742033 .
- ^ Vithani N, Bais V, Prakash B (июнь 2014 г.). «Glmu (N-ацетилглюкозамин-1-фосфат уридилтрансфераза) связана с тремя ионами магния и АТФ в активном сайте» . Acta Crystallographica Раздел f . 70 (Pt 6): 703–8. doi : 10.1107/s2053230x14008279 . PMC 4051520 . PMID 24915076 .
- ^ Wang J, Liu X, Liang YH, Li LF, Su XD (сентябрь 2008 г.). «Связывание акцепторного субстрата, выявленное кристаллической структурой глюкозамин-6-фосфата N-ацетилтрансферазы 1». Письма Febs . 582 (20): 2973–8. doi : 10.1016/j.febslet.2008.07.040 . PMID 18675810 . S2CID 5131540 .
- ^ Jump up to: а беременный «GNPNAT1 - глюкозамин 6 -фосфат N -ацетилтрансфераза - homo sapiens (человек) - ген и белок Gnpnat1» . www.uniprot.org . Получено 2019-03-15 .
- ^ «Flybase-глюкозамин 6-фосфатная N-ацетилтрансфераза» .
- ^ Lim S, Smith KR, Lim St, Tian R, Lu J, Tan M (2016-04-14). «Регуляция митохондриальных функций с помощью фосфорилирования и дефосфорилирования белка» . Клеточная и биологическая наука . 6 : 25. DOI : 10.1186/S13578-016-0089-3 . PMC 4832502 . PMID 27087918 .
- ^ Мио Т, Ямада-Окабе Т., Арисава М., Ямада-Окабе Х (январь 1999 г.). «Saccharomyces cerevisiae gna1, необходимый ген, кодирующий новую ацетилтрансферазу, участвующую в синтезе UDP-N-ацетилглюкозамина» . Журнал биологической химии . 274 (1): 424–9. doi : 10.1074/jbc.274.1.424 . PMID 9867860 .
- ^ Милевский С., Габриэль I, Олчови Дж (январь 2006 г.). «Ферменты биосинтеза UDP-GLCNAC у дрожжей» . Дрожжи . 23 (1): 1–14. doi : 10.1002/да.1337 . PMID 16408321 . S2CID 39940329 .
- ^ PDB : 2O28 ; Плотников А.Н., Бохкарев А., Эдвардс А.М., Эрроусмит Ч., Санстрем М., Вейгельт Дж. И др. «Кристаллическая структура глюкозамин-фосфата N-ацетилтрансферазы 1» . Всемирный банк данных белка . doi : 10.2210/pdb2o28/pdb .
- ^ Peneff C, Mengin-Lecreulx D, Bourne Y (май 2001). «Кристаллические структуры апо и комплексных Saccharomyces cerevisiae GNA1 проливают свет на каталитический механизм амино-сахарной N-ацетилтрансферазы» . Журнал биологической химии . 276 (19): 16328–34. doi : 10.1074/jbc.m009988200 . PMID 11278591 .