Диметродон
| Диметродон | |
|---|---|

| |
| Скелет Д. Лимбатуса , Государственный музей естественной истории Карлсруэе | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Synapsida |
| Семья: | † sphenacodontidae |
| Подсемейство: | † sphenacodontinae |
| Род: | † Диметродон Коуп , 1878 |
| Тип видов | |
| † Dimetrodon Limbatus Коуп, 1877
| |
| Разновидность | |
|
См . ниже | |
| Синонимы | |
|
Род синонимия Виды синонимия | |
Dimetrodon ( / D aɪ ˈ m iː t r ə ˌ d ɒ n / [ 1 ] или не D , это он t ½ ¯ du ½ ɒ n ; он [ 2 ] зажженная « Две меры зубов » ) является вымершим родом , не являющейся млекопитающими, синапсид принадлежащей к семейству Sphenacodontidae , которая жила в цисуральный век раннего пермского периода , около 295–272 миллионов лет назад. [ 3 ] [ 4 ] [ 5 ] С большинством видов длиной 1,7–4,6 м (5,6–15,1 фута) и весом 28–250 кг (62–551 фунт), наиболее заметной особенностью диметродона является большой нейрора плавание позвонки . Это был обязательный четвероночный (он мог ходить только на четырех ногах) и имел высокий изогнутый череп с большими зубами разных размеров вдоль челюстей. Большинство окаменелостей были обнаружены на юго -западе Соединенных Штатов , большинство из них поступают из геологического месторождения под названием « Красные слои Техаса и Оклахомы» . Совсем недавно его окаменелости также были обнаружены в Германии , и более дюжины видов были названы с тех пор, как род был впервые возведен в 1878 году.
Диметродон часто принимают за динозавр или как современник динозавров в популярной культуре, но он вымер за 40 миллионов лет до появления динозавров. [ 6 ] [ 7 ] похожа на рептилий по внешнему виду и физиологии, Несмотря на то, что Dimetrodon он гораздо более тесно связан с млекопитающими, чем с рептилиями, хотя он не является прямым предком млекопитающих. [ 4 ] Dimetrodon назначается «не млекопитающим синапсидам», группе традиционно-но неправильно-называемой «рептилиями, похожими на млекопитающие», [ 4 ] Но теперь известные как млекопитающие. Эти группы Dimetrodon вместе с млекопитающими в клады Synapsida, в то время как рептилии помещаются в отдельную кладу, Sauropsida . Одиночные отверстия в черепе за каждым глазом, известные как височные фенестра , и другие особенности черепа отличают Диметродона и настоящих млекопитающих от большинства самых ранних сауропсидов .
Диметродон, вероятно, был одним из хищников из цисульских экосистем, питаясь рыбой и тетраподами , включая рептилии и амфибии . Меньшие виды диметродонов могли иметь разные экологические роли . Парус Диметродона мог быть использован для стабилизации позвоночника или для нагрева и охлаждения его тела как формы терморегуляции . [ 8 ] Некоторые недавние исследования утверждают, что парус был бы неэффективным при удалении тепла из организма, из -за того, что крупные виды обнаружили с небольшими парусами и небольшими видами, обнаруженными с большими парусами, по существу, исключающему тепловой регуляции в качестве основной цели. Парус был, скорее всего, использовался на дисплее ухаживания , в том числе угрожая соперникам или демонстрацию потенциальным товарищам. [ 9 ] [ 10 ]
Описание
[ редактировать ]
Диметродон был четвероночной , поддерживаемой парусом синапсидией, которая, скорее всего, имела полупрокативную позу между млекопитающим и ящерицей, а также мог ходить в более верной позиции с его телом и большинством или весь его хвост от земли. [ 11 ] Большинство видов диметродона длины от 1,7 до 4,6 м (от 6 до 15 футов) и, по оценкам, весом от 28 до 250 кг (60 и 550 фунтов). [ 12 ] Самые маленькие известные виды D. teutonis были длиной около 60 см (24 дюйма) и весили 14 килограммов (31 фунт). [ 12 ] [ 13 ] Более крупные виды диметродона были одними из самых больших хищников раннего пермца, хотя тесно связанный Таппенозавр , известный по скелетным фрагментам в немного более молодых породах, мог быть еще больше на уровне 5,5 метров (18 футов) длиной. [ 14 ] [ 15 ] Хотя некоторые виды Dimetrodon могут расти очень большими, многие юношеские образцы известны. [ 16 ]
Череп
[ редактировать ]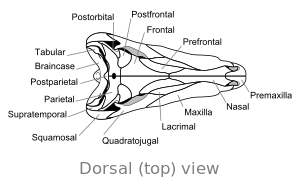 |
 |
 |
 |
Одно большое отверстие по обе стороны от задней части черепа связывает диметродона с млекопитающими и отличает его от большинства самых ранних сауропсидов, в которых либо отсутствует отверстия , либо имеют два отверстия . Такие особенности, как хребты на внутренней стороне носовой полости и гребень на задней части нижней челюсти, являются частью эволюционного прогрессирования от ранних четырех лиммированных позвоночных до млекопитающих .
Череп Диметродона высокий и сжимается в боковом направлении или от стороны в сторону. Глазки расположены высоко и далеко назад в черепе. За каждой глазной гнездой находится единственная дыра, называемая инфрат -временной фенестрой . Дополнительная дыра в черепе, супратромочная фенестра , можно увидеть при просмотре сверху. Задняя часть черепа ( затылака ) ориентирована под небольшим углом вверх, функция, которую он разделяет со всеми другими ранними синапсидами . [ 17 ] Верхний запас черепа наклоняется вниз в выпуклой дуге к кончику морды. Кончик верхней челюсти, образованный костью предчезгиллы , поднимается над частью челюсти, образованной верхней челюстной костью, чтобы сформировать верхнюю часть «шага». Внутри этого шага находится диастема , разрыв в ряду зубов. Его череп был более сильно построен, чем череп динозавра.
Зубы
[ редактировать ]Размер зубов сильно варьируется по длине челюстей, придавая диметродон его название, что означает «два меры зуба» в отношении наборов мелких и больших зубов. [ 18 ] Одна или две пары канинии (большие, заостренные, похожие на собаки ) простираются от верхней челюсти. Большие зубы резца также присутствуют на кончиках верхних и нижних челюстей, укоренившихся в преднамеренных и зубных костях . Маленькие зубы присутствуют вокруг верхнечелюстного «шага» и за канихообразной, становясь меньше назад в челюсти. [ 19 ]

Многие зубы самые широкие в их средних и узких ближе к челюстям, давая им вид слез. Зубы в форме слез уникальны для Dimetrodon и других тесно связанных спгенакодонтидов , которые помогают отличить их от других ранних синапсидов. [ 13 ] Как и во многих других ранних синапсидах , зубы большинства видов диметродонов зазубрены по их краям. [ 13 ] Ссудеры зубов Диметродона были настолько хороши, что они напоминали крошечные трещины. [ 20 ] У динозавра у Альбертозавра были аналогичные трещины, но у основания каждого зубца была круглая пустота , которая бы функционировала для распределения силы по более широкой площади поверхности и предотвращения напряжений кормления, вызвавшего распределение трещины через зуб Полем В отличие от Albertosaurus , зубья -диметродона не имели адаптации, которые остановили бы трещины от формирования их зубцов. [ 20 ] У зубов D. teutonis не хватает зубцов, но все еще имеют острые края. [ 13 ]
Исследование 2014 года показывает, что Dimetrodon находился в гонке вооружений против ее добычи. Меньшие виды, D. milleri , не имели зубных зубцов, потому что он ел небольшую добычу. По мере того, как добыча стала больше, несколько видов Dimetrodon начали развивать зубцы на зубах и увеличиваться в размере. Например, у D. limbatus были эмалевые зубцы, которые помогли ему прорезать плоть (которые были похожи на зубцы, которые можно найти на Secodontosaurus ). У второго по величине вида, D. grandis , есть зубчатые зубцы, похожие на раздувание акул и динозавров теропода , что делает его зубы еще более специализированными для прорезки плоти. По мере того, как добыча Dimetrodon выросла, различные виды реагировали на то, чтобы расти до больших размеров и развивающиеся вечно тщательные зубы. [ 21 ] Толщина и масса зубов Диметродона также могли быть адаптацией для увеличения долговечности зубов. [ 22 ]
Носовая полость
[ редактировать ]На внутренней поверхности носового участка черепа находятся хребты, называемые назотурбиналами , которые могли поддерживать хрящ, который увеличивал площадь обонятельного эпителия , слой ткани, который обнаруживает запахи. Эти хребты намного меньше, чем у последующих синапсидов от позднего пермского и триаса, чьи большие носотурбиналы рассматриваются в качестве доказательства теплокровной, потому что они могли поддерживать слизистые мембраны, которые согревали и увлажняли входящий воздух. Таким образом, носовая полость диметродона является переходной между перенесенными земельными позвоночными и млекопитающими. [ 23 ]
Челюстное соединение и ухо
[ редактировать ]Другая переходная особенность Dimetrodon - гребень в задней части челюсти, называемой отраженной пластинки, которая находится на суставной кости, которая соединяется с квадратной костью черепа, образуя челюстный сустав. У более поздних предков млекопитающих суставной и квадрат отделялись от челюстного сустава, в то время как суставкура превратилась в кость Malleus среднего уха . Отраженная пластинка стала частью кольца, называемого барабанным кольцом, которое поддерживает ушной барабан у всех живых млекопитающих. [ 24 ]
Хвост
[ редактировать ]
Хвост Диметродона составляет большую часть общей длины тела и включает в себя около 50 каудальных позвонков . Хвосты отсутствовали или неполными в первых описанных скелетах Диметродона . Единственными известными каудальными позвонками были 11 ближайших к бедру. С тех пор, как эти первые несколько каудальных позвонков узко узко устроены, когда они прогрессируют дальше от бедра, многие палеонтологи в конце 19 -го и начале 20 -го веков считали, что у Диметродона был очень короткий хвост. В основном полный хвост Диметродона не был описан до 1927 года. [ 25 ]
Плыть
[ редактировать ]
Парус Диметродона образуется удлиненными нейронными шипами, проецирующими от позвонков. Каждый позвоночник варьируется по форме поперечного сечения от его основания до его кончика в так называемой дифференцировке «диматродонта». [ 26 ] Рядом с корпусом позвонка поперечное сечение позвоночника складывается в прямоугольную форму, и, ближе к кончику, он принимает форму восьмой фигуры, когда канавка проходит по обе стороны от позвоночника. Считается, что форма-фигура восьмерка усиливает позвоночник, предотвращая изгибы и переломы. [ 27 ] Поперечное сечение позвоночника одного образца диметродонских гиганхомогенов имеет прямоугольную форму, но сохраняет кольца с фигурной формой вблизи его центра, что указывает на то, что форма шипов может меняться по возрасту. [ 28 ] Микроскопическая анатомия каждого позвоночника варьируется от основания к наконечникам, что указывает на то, где она была встроена в мышцы спины и где он был обнаружен как часть паруса. Нижняя или проксимальная часть позвоночника имеет шероховатую поверхность, которая послужила бы точкой привязки для эпаксиальных мышц спины, а также имеет сеть соединительных тканей, называемых волокнами Шарпи , которые указывают на то, что он был встроен в организм. Выше на дистальной (внешней) части позвоночника поверхность кости более гладкая. Периостеум . , слой ткани, окружающую кость, покрыт небольшими бороздками, которые, по -видимому, поддерживали кровеносные сосуды, которые сосудились на парус [ 29 ]
Большая канавка, которая проходит по длине позвоночника, когда -то считалась каналом для кровеносных сосудов, но, поскольку кость не содержит сосудистых каналов, с считается, что парус не был столь же очень васкуляризирован, как когда -то думали. Некоторые образцы диметродона сохраняют деформированные области нервных шипов, которые, по-видимому, заживают переломы. Корковая кость , которая выросла над этими разрывами, сильно сосудится, что позволяет предположить, что мягкие ткани должны присутствовать на парусе, чтобы снабжать место кровеносными сосудами . [ 27 ] Слоистая пластинчатая кость составляет большую часть поперечного сечения нейронного позвоночника и содержит линии арестованного роста, которые можно использовать для определения возраста каждого человека при смерти. [ 30 ] Во многих образцах D. gigashomogenes дистальные части шипов резко изгибаются, что указывает на то, что парус имел бы нерегулярный профиль в жизни. Их изящество предполагает, что мягкие ткани, возможно, не распространились до кончиков шипов, а это означает, что лямочные ленты паруса, возможно, не были такими обширными, как это обычно представляют. [ 26 ]
Кожа
[ редактировать ]
ископаемых доказательств кожи Диметродона Пока не обнаружено . Впечатления кожи родственного животного, Estemmenosuchus , указывают на то, что оно было бы гладким и хорошо предоставленным железами, но эта форма кожи, возможно, не применима к Dimetrodon , поскольку его происхождение довольно отдаленная. [ 31 ] Диметродон также мог иметь большие закуски на нижней стороне его хвоста и живота, как у других синапсидов. [ 32 ] [ 33 ] Свидетельство от варанопидного Ascendonanus предполагает, что некоторые ранние синапсиды, возможно, имели шкалы, похожие на сквамат . [ 34 ] Тем не менее, некоторые недавние исследования поставили варанопиды таксономически ближе к диапсидов рептилиям . [ 35 ] [ 36 ]
История классификации
[ редактировать ]Самые ранние открытия
[ редактировать ]
Самое раннее открытие окаменелостей Диметродона была из версии, восстановленной в 1845 году человеком по имени Дональд Маклеод, живущий в британской колонии острова принца Эдуарда . [ 37 ] Эти окаменелости были приобретены Джоном Уильямом Джонсоном, канадским геологом, а затем описанным Джозефом Лейди в 1854 году как нижняя челюсть батинатуса бореалиса , большого плотоядного животного , связанного с TheCodontosaurus , [ 38 ] Хотя в 2015 году он был позже реклассифицирован как вид диметродона , как Dimetrodon Borealis . [ 39 ]
Первые описания, справится
[ редактировать ]Окаменелости теперь приписывают Dimetrodon , впервые изученные американским палеонтологом Эдвардом Дим -ным Коупом в 1870 -х годах. Коуп получил окаменелости вместе с таковыми у многих других пермских тетрапод от нескольких коллекционеров, которые изучали группу камней в Техасе, называемые красными кроватями . Среди этих коллекционеров были швейцарский натуралист Джейкоб Болл , Техасский геолог WF Cummins и любительский палеонтолог Чарльз Хазелиус Стернберг . [ 40 ] Большинство экземпляров COPE отправились в Американский музей естественной истории или в Музей Уолкера Университета Чикагского университета (большая часть коллекции ископаемых ходьбы теперь находится в Полевом музее естественной истории ).
Стернберг послал некоторые из своих собственных образцов немецкому палеонтологу Фердинанду Бройли в Мюнхенском университете , хотя Broili не был таким плодовитым, как в описании образцов. Соперник Коупа Отниэль Чарльз Марш также собрал несколько костей Диметродона , которые он отправил в музей Уокера. [ 41 ] Первое использование названия Dimetrodon появилось в 1878 году, когда Коуп назвал вида Dimetrodon incisivus , Dimetrodon Rectiformis и Dimetrodon Gigas в Служении научного журнала Американского философского общества . [ 42 ]
Первое описание ископаемого Dimetrodon появилось годом ранее, когда Коуп назвал виды Clepsydrops Limbatus из красных ложе Техаса. [ 43 ] (Название Clepsydrops было впервые придуманным COPE в 1875 году для спенакодонтида из округа Вермилион, штат Иллинойс , и позже использовался для многих образцов спгенаконтидов из Техаса; многие новые виды спенакодонтидов из Техаса были назначены клепсидроп любое на останков В начале 20 веков.) C. Limbatus был реклассифицирован как вид диметродона в 1940 году, что означает, что статья Cope 1877 года была первой записью Dimetrodon .
Коуп был первым, кто описывал синапсид, поддерживаемую парусом с именом C. natalis в своей статье 1878 года, хотя он назвал парус плавником и сравнил ее с гребнями современной лизарда из базилиска ( Basilicus ). Парусы не были сохранены в образцах D. incisivus и D. gigas , которые Cope описали в его статье 1878 года, но удлиненные шипы присутствовали в образовании D. ertiformis , который он описал. [ 42 ] Коуп прокомментировал цель паруса в 1886 году, написав: «Утилита трудно представить. Если у животного не было водных привычек, и плавал на спине, гребень или плавник, должно быть, были на пути активных движений ... Конечности не достаточно долго, и когти достаточно острые, чтобы продемонстрировать древесные привычки, как в существующем роде Basilicus , где существует подобный гребень ». [ 19 ]
Описания начала 20 -го века
[ редактировать ]
В первые несколько десятилетий 20 -го века американский палеонтологский случай ЕС создал множество исследований на Dimetrodon и описал несколько новых видов. Он получил финансирование от института Карнеги для изучения многих образцов Dimetrodon в коллекциях Американского музея естественной истории и нескольких других музеев. [ 41 ] Многие из этих окаменелостей были собраны CAPE, но не были тщательно описаны, так как COPE был известен тем, что возводил новые виды на основе лишь нескольких фрагментов кости.
Начиная с конца 1920 -х годов, палеонтолог Альфред Ромер воздержал много образцов диметродона и назвал несколько новых видов. В 1940 году Ромер стал соавтором крупного исследования с ценой Llewellyn Ivor , называемой «Обзор Пеликозаурии», в котором были переоценены виды Диметродона, названного COPE и CASE. [ 44 ] Большинство названий видов, считающихся действительными, и цена, все еще используются сегодня. [ 29 ]
Новые образцы
[ редактировать ]В течение десятилетий после монографии Ромера и Прайса многие образцы Диметродона были описаны из населенных пунктов за пределами Техаса и Оклахомы . Первый был описан из региона четырех угла в Юте в 1966 году [ 45 ] а другой был описан из Аризоны в 1969 году. [ 46 ] В 1975 году Олсон сообщил о Диметродона материале из Вашингтонского образования Огайо, которому было дано предварительное назначение D. cf. Лимбатус . [ 47 ] [ 48 ] [ 49 ] Новый вид диметродона под названием D. occidentalis (то есть «Западный Диметродон ») был назван в 1977 году из Нью -Мексико. [ 50 ] Образцы, найденные в Юте и Аризоне, вероятно, также принадлежат D. occidentalis . [ 51 ]
Перед этими открытиями существовала теория о том, что средняя конндентальная морская дорога отделяла то, что сейчас является Техасом и Оклахомой от больших западных земель во время раннего пермского, изолируя диметродон в небольшой регион Северной Америки, в то время как меньший сфенакодонтид, называемый Сфинакодоном, доминировал в западном районе. В то время как этот морской путь, вероятно, существовал, обнаружение окаменелостей за пределами Техаса и Оклахомы показывает, что его протяженность была ограничена и что это не было эффективным барьером для распределения Диметродона . [ 50 ] [ 52 ]
В 2001 году был описан новый вид диметродона, называемый D. teutonis из местности нижнего пермского брамакера в Тюринговом лесу Германии, расширяя географический диапазон диметродона за пределами Северной Америки. впервые [ 12 ]
Разновидность
[ редактировать ]
Двадцать видов диметродона был впервые описан в 1878 году . были названы с тех пор, как род Многие из них были синонимизированы с старшими названными видами, а некоторые теперь принадлежат к различным родам.
Краткое содержание
[ редактировать ]| Разновидность | Власть | Расположение | Статус | Синонимы | Изображения |
|---|---|---|---|---|---|
| Диметродон Анджеленс | Олсон, 1962 |
|
Действительный | 
| |
| Dimetrodon Borealis | Лейди, 1854 |
|
Действительный | Ранее известный как Dinosaur Bathygnathus borealis | 
|
| Dimetrodon Booneorum | Ромер, 1937 |
|
Действительный | ||
| Dimetrodon olovianus | Дело, 1907 |
|
Действительный | Embolophorus oulovianus cope, | |
| Dimetrodon Gigahomogenes | Дело, 1907 |
|
Действительный | 
| |
| Диметродон Грандис | Ромер и Прайс, 1940 |
|
Действительный | Clepsydrops Gigas Cope, 1878 Dimetrodon Gigas Cope, 1878 Дело Theropleura Grandis , 1907 Bathygly Theodore Case, 1911 Dimetrodon Maximus Romer 1936 |

|
| Dimetrodon Kempae | Ромер, 1937 |
|
Возможное имя без сомнения | ||
| Dimetrodon Limbatus | Ромер и Прайс, 1940 |
|
Действительный | Clepsydrops Limbatus Cope, 1877 Dimetrodon incisivus Cope, 1878 Dimetrodon Rectiformis Cope, 1878 Dimetrodon Semiradicatus Cope, 1881 |
|
| ДИМЕТРОДОНСКИЕ ТООТЫ | Ромер, 1937 |
|
Действительный | 
| |
| Dimetrodon macrospondylus | Дело, 1907 |
|
Действительный | Clepsydrops macrospondylus cope, 1884 Demetrodon Platycentrus Case, 1907 |
|
| Dimetrodon Milleri | Ромер, 1937 |
|
Действительный | 
| |
| Диметродон Наталис | Ромер, 1936 |
|
Действительный | Clepsydrops Natalis Cope, 1878 | 
|
| Dimetrodon occidentalis | Берман, 1977 |
|
Действительный | ||
| Dimetrodon Teutonis | Berman et al. , 2001 |
|
Действительный |
Dimetrodon Limbatus
[ редактировать ]
Dimetrodon Limbatus был впервые описан Эдвардом Дрибцом Коуп в 1877 году как Clepsydrops Limbatus . [ 43 ] (Имя Clepsydrops было впервые придуманным COPE в 1875 году для останков спенакодонтида из округа Вермилион, штат Иллинойс , и позже использовался для многих образцов спенаконтидов из Техаса; многие новые виды спенакодонтидов из Техаса были назначены на любой клепсиду или Dimetrodon в конце NineTectyd и Texas и Dimetrodon в покойном Nineletent и Texas и Dimetrodon. В начале двадцатых веков.) Основываясь на образце из красных слоев Техаса , это была первая известная синапсид, поддерживаемая парусным плаванием. В 1940 году палеонтологи Альфред Ромер и Ллевеллин Айвор Прайс переназначил C. limbatus род Dimetrodon , что сделало D. limbatus типовым видом диметродона на . [ 44 ] Останки осторожно назначены для этого вида, также известны из округа Вашингтон, штат Огайо , что соответствует относительно большому человеку. Эти останки немного старше, чем другие, назначенные D. limbatus с запада, хотя потенциальный D. limbatus остается из Нью -Мексико может быть одновременно. [ 49 ]
Dimetrodon incisivus
[ редактировать ]Первое использование названия Dimetrodon произошло в 1878 году, когда Cope назвал вида Dimetrodon incisivus вместе с Dimetrodon Rectiformis и Dimetrodon Gigas . [ 42 ]
Dimetrodon Rectiformis
[ редактировать ]Dimetrodon Rectiformis был назван рядом с Dimetrodon Incisivus в статье Коупа 1878 года и был единственным из трех названных видов, которые сохранили удлиненные нейронные шипины. [ 42 ] В 1907 году палеонтологский случай ЕС переместил D. Rectiformis в вид D. incisivus . [ 41 ] D. Incisivus был впоследствии синонимом типового вида Dimetrodon Limbatus , что делает D. rectiformis синонимом D. limbatus . [ 29 ]
Dimetrodon Semiradicatus
[ редактировать ]Описанный в 1881 году на основе верхних костей челюсти, Dimetrodon Semiradicatus был последним видом, названным COPE. [ 53 ] В 1907 году синонимизировал случай ЕС D. semiradicatus с D. incisivus, основанный на сходстве в форме зубов и костей черепа. [ 41 ] D. incisivus и D. semiradicatus в настоящее время считаются синонимами D. limbatus . [ 29 ]
Dimetrodon olovianus
[ редактировать ]Dimetrodon Allovianus был впервые описан Эдвардом Дрибцом Коуп в 1888 году как Embolophorus olovianus . В 1903 году дело о ЕС опубликовало длинное описание E. Dollovianus , которое он позже сослался на Dimetrodon . [ 54 ]
Диметродон Грандис
[ редактировать ]
Палеонтолог EC Case назвал новый вид Synapsid Synapsid, Theropleura Grandis , в 1907 году. [ 41 ] В 1940 году Альфред Ромер и Ллевеллин Айвор Прайс переназначил Theropleura Grandis на Dimetrodon , возведу виды D. grandis . [ 44 ]
Диметродон Гигас
[ редактировать ]В своей статье 1878 года о окаменелостях из Техаса Коуп назвал Clepsydrops Gigas вместе с первыми названными видами Dimetrodon , D. Limbatus , D. incisivus и D. ertiformis . [ 42 ] Случай реклассифицирован C. gigas как новый вид диметродона в 1907 году. [ 41 ] Кейс также описал очень хорошо сохранившийся череп диметродона в 1904 году, приписывая его видам диметродон Гигас . [ 55 ] В 1919 году Чарльз У. Гилмор приписал почти полный образец диметродона D. Gigas . [ 56 ] Dimetrodon Gigas теперь признан синонимом D. grandis . [ 57 ]
Dimetrodon Giganhomogenes
[ редактировать ]
Dimetrodon Giganhomogenes был назван случаем EC в 1907 году и до сих пор считается действительным видом Dimetrodon . [ 41 ] [ 29 ]
Dimetrodon macrospondylus
[ редактировать ]Dimetrodon Macrospondylus был впервые описан COPE в 1884 году как Clepsydrops Macrospondylus . В 1907 году случай реклассифицировал его как Dimetrodon Macrospondylus . [ 41 ]
Dimetrodon PlatyCentrus
[ редактировать ]Dimetrodon PlatyCentrus был впервые описан по случаю в его монографии 1907 года. В настоящее время он считается синонимом Dimetrodon Macrospondylus . [ 29 ]
Диметродон Наталис
[ редактировать ]
Палеонтолог Альфред Ромер построил вида Dimetrodon Natalis в 1936 году, ранее описанный как Clepsydrops Natalis . Д. Наталис был самым маленьким известным видом диметродона в то время и был найден вместе с остатками более крупного тела D. limbatus . [ 58 ]
Dimetrodon Booneorum
[ редактировать ]Диметродон Бунеорум был впервые описан Альфредом Ромером в 1937 году на основе останков из Техаса. [ 58 ]
"Деметродон" Комбо
[ редактировать ]Dimetrodon Kempae был назван Ромером в 1937 году, в той же статье, что и D. Booneorum , D. Loomisi и D. Milleri . [ 58 ] Dimetrodon Kempae был назван на основе единой плечевой кости и нескольких позвонков, и поэтому может быть номенем дубия , который нельзя выделить как уникальный вид диметродона . [ 12 ] В 1940 году Ромер и Прайс подняли возможность того, что D. Kempae не может попасть в род Dimetrodon , предпочитая классифицировать его как Sphenacodontidae Incertae sedis . [ 44 ]
ДИМЕТРОДОНСКИЕ ТООТЫ
[ редактировать ]
Диметродон Лумиси был впервые описан Альфредом Ромером в 1937 году вместе с Д. Бунеорумом , Д. Кемпэ и Д. Миллери . [ 58 ] Останки были найдены в Техасе и Оклахоме.
Dimetrodon Milleri
[ редактировать ]
Dimetrodon Milleri был описан Ромером в 1937 году. [ 58 ] Это один из самых маленьких видов диметродона в Северной Америке и может быть тесно связан с D. occidentalis , другим видом с мелким телом. [ 51 ] D. Milleri известен по двум скелетам, один почти завершен (MCZ 1365) и еще один менее полный, но больше (MCZ 1367). D. Milleri - самый старый известный вид диметродона .
Помимо своего небольшого размера, D. milleri отличается от других видов диметродона тем, что его нервные колючки имеют круговые, а не фигурирующие в форме поперечного сечения. Его позвонки также более короче по высоте по сравнению с остальной частью скелета, чем у других видов диметродона . Череп высокий, а морда короткая по сравнению с височной областью. Короткие позвонки и высокий череп также видны у вида D. booneorum , D. limbatus и D. grandis , предполагая, что D. milleri может быть первым из эволюционного прогрессирования между этими видами.
Диметродон Анджеленс
[ редактировать ]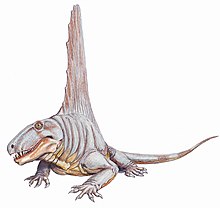
Диметродон Анджеленс был назван палеонтологом Эвереттом С. Олсоном в 1962 году. [ 59 ] Образцы вида были зарегистрированы из формирования Сан -Анджело в Техасе. [ 60 ] Это также самый большой вид диметродона.
Dimetrodon occidentalis
[ редактировать ]Dimetrodon occidentalis был назван в 1977 году из Нью -Мексико. [ 50 ] Его название означает «Западный Диметродон », потому что это единственный североамериканский вид диметродона, известный к западу от Техаса и Оклахомы. Он был назван на основе одного скелета, принадлежащего относительно маленькому человеку. Небольшой размер D. occidentalis похож на размер D. milleri , что предполагает тесную связь. Образцы Диметродона , найденные в Юте и Аризоне, вероятно, также принадлежат D. occidentalis . [ 51 ]
Dimetrodon Teutonis
[ редактировать ]Dimetrodon Teutonis был назван в 2001 году из леса Тюрингян Германии и был первым видом Диметродона, который был описан за пределами Северной Америки. Это также самый маленький вид диметродона . [ 12 ]
Виды, назначенные различным родам
[ редактировать ]Диметродон Круцигер
[ редактировать ]В 1878 году Коуп опубликовал статью под названием «Тераторфузная рептилия», в которой он описал Dimetrodon Cruciger . [ 61 ] Д. Круцигер отличался небольшими проекциями, которые простирались с обеих сторон каждого нейронного позвоночника, как ветви дерева. [ 62 ] В 1886 году Коуп переместил D. cruciger к роду Naosaurus, потому что он считал, что его шипы настолько отличаются от других видов диметродона , что вид заслужил свой собственный род. [ 63 ] Назаурус позже будет синонимизироваться с Edaphosaurus , родом, который справляется с названием в 1882 году на основе черепов, которые, очевидно, принадлежали травоядным животным, учитывая их тупые зубы. [ 64 ]
Диметродон Лонграмус
[ редактировать ]Случай EC назвал вида Dimetrodon Longiramus в 1907 году на основе лопатки и удлиненной нижней челюсти из формирования Belle Plains в Техасе. [ 41 ] В 1940 году Ромер и Прайс признали, что материал D. longiramus принадлежал тому же таксону, что и еще один образец, описанный палеонтологом Сэмюэлем Венделлом Уиллистоном в 1916 году, который включал аналогичную удлиненную нижнюю челюсть и длинную верхнюю верхнюю верхнюю часть. [ 44 ] Уиллистон не считал, что его образец принадлежит Диметродону , но вместо этого классифицировал его как Ophiacodontid . [ 65 ] Ромер и цена назначил дело и образцы Уиллистона новорожденному роду и видам Secodontosaurus longiramus , которые были тесно связаны с Dimetrodon . [ 44 ] [ 66 ]
Филогенетическая классификация
[ редактировать ]Dimetrodon является ранним членом группы, называемой Synapsids , в которую входят млекопитающие и многие из их вымерших родственников, хотя он не является предком какого -либо млекопитающего (которые появились миллионы лет спустя. [ 67 ] ) Он часто принимается за динозавр в популярной культуре, несмотря на то, что он вымер около 40 миллионов лет (MA) до первого появления динозавров в триасовый период. Как синапсид, Dimetrodon более тесно связан с млекопитающими, чем с динозаврами или любой живой рептилий. К началу 1900 -х годов большинство палеонтологов назвали Диметродон рептилия в соответствии с таксономией Линни , которая оценила рептилия как класс и диметродон как род в этом классе. Млекопитающие были назначены в отдельный класс, а Dimetrodon был описан как «рептилия, похожая на млекопитающие». Палеонтологи теоретизировали, что млекопитающие эволюционировали из этой группы в (что они называли) переход рептилий к млекопитающему.
Филогенетическая таксономия синапсиды
[ редактировать ]
При филогенетической систематике потомки последнего общего предка Диметродона . и всех живых рептилий будут включать всех млекопитающих, потому что Диметродон более тесно связан с млекопитающими, чем с любой живой рептилий Таким образом, если желательно избежать клады, которая содержит как млекопитающих, так и живых рептилий, то Dimetrodon не должен быть включен в эту кладу, а не какую-либо другую «млекопитающую рептилия». Потомки последнего общего предка млекопитающих и рептилий (которые появились около 310 млн. Лет в позднем каменноугольнике ), поэтому разделены на две клады: Synapsida, который включает в себя диметродон и млекопитающие, а также Sauropsida , которые включают живые рептилии и все вымершие рептилии более тесно связаны им, чем млекопитающих. [ 4 ]
В рамках Clade Synapsida Dimetrodon является частью Clade Sphenacodontia , которая была впервые предложена в качестве ранней синапсид в 1940 году палеонтологами Альфредом Ромером и Llewellyn Ivor Price, наряду с группами Ophiacodontia и Edaphosauria . [ 44 ] Все три группы известны из позднего каменноугольного и раннего пермца. Ромер и цена отличали их в основном посткраниальными особенностями, такими как формы конечностей и позвонки. Ophiacodontia считалась самой примитивной группой, потому что ее члены казались самыми рептилиями, а спенакодонтия была самой продвинутой, потому что ее члены казались наиболее похожими на группу под названием Therapsida , которая включала ближайших родственников к млекопитающим. Ромер и Прайс разместили еще одну группу ранних синапсидов, называемых варанопидами в сфенакодонтии, считая, что они более примитивны, чем другие спенакодонты, такие как Dimetrodon . [ 68 ] Они думали, что варанопиды и диметродон, похожие на спенакодонты, были тесно связаны, потому что обе группы были плотоядными, хотя варанопиды намного меньше и более похожи на ящерицу, отсутствуют паруса.
Современный взгляд на синапсид был предложен палеонтологом Робертом Р. Рейсом в 1986 году, чье исследование включало особенности, в основном обнаруженные в черепе, а не в посткраниальном скелете. [ 69 ] Диметродон по -прежнему считается сфенакодонтом в этой филогении , но варанодонтиды теперь считаются более базальными синапсидами, падая за пределы клады спенакодонтии. Внутри спенакодонтии находится группа Sphenacodontoidea , которая, в свою очередь, содержит Sphenacodontidae и Therapsida . Sphenacodontidae-это группа, содержащая Dimetrodon и несколько других поддерживаемых парусными синапсидами, таких как Sphenacodon и Secodontosaurus , в то время как Therapsida включает млекопитающих и их в основном перми и триасовые родственники.
Ниже приведена кладограмма Clade Synapsida, которая следует за этой филогении Synapsida, как модифицировано из анализа Benson (2012). [ 68 ]
| Амниота |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Приведенная ниже кладограмма показывает отношения нескольких видов диметродона , от Brink et al. , (2015). [ 70 ]
| Sphenacodontidae |
| ||||||||||||||||||||||||||||||
Палеобиология
[ редактировать ]Функция нейронных шипов
[ редактировать ]
Палеонтологи предложили много способов, которыми парус мог бы функционировать в жизни. Некоторые из первых, кто подумал о ее цели, предположили, что парус, возможно, служил камуфляжем среди тростников, в то время как Диметродон ждал добычи или в качестве настоящего лодочного паруса, чтобы поймать ветер, пока животное находилось в воде. [ 71 ] Другое заключается в том, что длинные нейронные шипы могли бы стабилизировать ствол, ограничивая движение вверх и вниз, что позволило бы более эффективному движению от стороны в сторону во время ходьбы. [ 27 ]
Терморегуляция
[ редактировать ]В 1940 году Альфред Ромер и Ллевеллин Айвор Прайс предложили, чтобы парус выполнял терморегуляторную функцию, позволяя людям согревать свои тела с солнцем. В последующие годы было создано многие модели для оценки эффективности терморегуляции в Dimetrodon . Например, в статье 1973 года в журнале Nature палеонтологи С. Д. Брэмвелл и П. Б. Феллгетт подсчитали, что потребовалось индивидуальное значение на 200 килограммов (440 фунтов) около полутора часов, чтобы температура его тела поднялась с 26 до 32 ° C (79 до 90 ° F). [ 72 ] В 1986 году Стивен С. Хаак пришел к выводу, что потепление было медленнее, чем думал ранее, и что процесс, вероятно, занял четыре часа. Используя модель, основанную на различных факторах окружающей среды и гипотетических физиологических аспектах Dimetrodon , Haack обнаружил, что парус позволил Dimetrodon быстрее утро и достигать немного более высокой температуры тела в течение дня, но он был неэффективным в высвобождении избыточного тепла и не позволил Диметродону сохранить более высокую температуру тела ночью. [ 73 ] В 1999 году группа инженеров -механиков создала компьютерную модель для анализа способности паруса регулировать температуру тела в течение разных сезонов, и пришла к выводу, что парус был полезен для захвата и выпуска тепла в любое время в год. [ 74 ]

Большинство из этих исследований дают две терморегуляторные роли для паруса Диметродона : одно как средство быстрого нагревания утром, а другой - как способ остыть, когда температура тела становится высокой. Предполагается, что Dimetrodon и все другие ранние пермские земельные позвоночные были хладнокровными или пукилотермическими , полагаясь на солнце, чтобы поддерживать высокую температуру тела. Из-за своего большого размера Dimetrodon имел высокую тепловую инерцию , что означает, что изменения в температуре тела происходили медленнее в нем медленнее, чем у животных меньшего тела. По мере того, как температура выросла по утрам, добыча с маленьким телом диметродона могла согреть свои тела намного быстрее, чем что-то вроде размера Dimetrodon . Многие палеонтологи, включая Хаака, предположили, что парус Диметродона , возможно, позволил ему быстро нагреться утром, чтобы идти в ногу со своей добычей. [ 73 ] Большая площадь поверхности паруса также означала, что тепло может быстро распаться в окружающую среду, полезное, если животному нужно было высвобождать избыточное тепло, полученное в результате метаболизма, или поглощать от солнца. Dimetrodon , возможно, уклонился от его паруса от солнца, чтобы остыть или ограниченный кровоток к парусу, чтобы поддерживать тепло ночью. [ 71 ]
В 1986 году Дж. Скотт Тернер и К. Ричард Трейси предложили, чтобы эволюция паруса в Диметродоне была связана с эволюцией теплокровной, предков млекопитающих. Они думали, что парус Диметродона позволил ему быть гомеотермическим , сохраняя постоянную, хотя и низкую температуру тела. Млекопитающие также являются гомеотермическими, хотя они отличаются от диметродона от эндотермических , контролируя температуру их тела внутри, посредством повышенного метаболизма. Тернер и Трейси отметили, что ранние терапиды, более продвинутая группа синапсидов, тесно связанных с млекопитающими, имели длинные конечности, которые могут высвобождать тепло таким образом, сходным с планой диметродона . Годотермия, которая развивалась у животных, таких как Диметродон, могла перенести в терапсиды посредством модификации формы тела, которая в конечном итоге превратится в теплокровную млекопитающие. [ 75 ]

Недавние исследования Паруса Диметродона и других спгенакодонтидов подтверждают утверждение Хаака в 1986 году о том, что парус был плохо адаптирован к выпуску тепла и поддержанию стабильной температуры тела. Присутствие парусов у маленьких видов диметродона, таких как D. milleri и D. teutonis, не соответствует идее, что целью паруса была терморегуляция, потому что меньшие паруса менее способны переносить тепло и потому что мелкие тела могут легко поглощать и высвобождать тепло самостоятельно. Более того, близкие родственники Диметродона, такие как Спенакодон, имеют очень низкие гребни, которые были бы бесполезны в качестве терморегуляторных устройств. [ 29 ] Считается, что большой парус Диметродона постепенно развивался из этих меньших гребней, что означает, что в течение большей части эволюционной истории паруса терморегуляция не могла выполнить важную функцию. [ 76 ]
Хотя функция его паруса остается неопределенной, Dimetrodon и других спгенакодонтидов, вероятно, были эндотермами всего тела, характеризующимися высокой энергетической метаболизмом ( тахиметаболизм ) и, вероятно, способностью поддерживать высокую и стабильную температуру тела. Этот вывод был частью общепринятого исследования, которое обнаружило, что тахиметаболическая эндотермия была широко распространена во всем, и, вероятно, плезиоморфной как синапсидам , так и для сауропсидов . Для Dimetrodon доказательством было эндотерми-индикативное размер отверстия, посредством которого кровь доставлялась к их длинным костям, и высокое кровяное давление, которое было бы необходимо для обеспечения крови для вершин хорошо сосудированных шипов, поддерживающих парус. [ 77 ]
Более крупные образцы диметродона имеют более крупные парусы по сравнению с их размером, пример положительной аллометрии . Положительная аллометрия может принести пользу терморегуляции, потому что это означает, что по мере того, как отдельные люди становятся все больше, площадь поверхности увеличивается быстрее, чем масса. Животные с более крупным телом генерируют большую тепло в результате метаболизма, и количество тепла, которое должно быть рассеивается с поверхности тела, значительно больше, чем то, что должно быть рассеивается животными меньшими телом. Эффективное рассеяние тепла может быть предсказано на многих различных животных с единственной взаимосвязью между массой и площадью поверхности. Тем не менее, исследование аллометрии в 2010 году в Dimetrodon обнаружило другую связь между его парусом и массой тела: фактический показатель масштабирования паруса был намного больше, чем показатель, ожидаемый у животного, адаптированного к рассеянию тепла. Исследователи пришли к выводу, что парус Диметродона рос гораздо быстрее, чем необходимо для терморегуляции, и предположил, что половой отбор был основной причиной его эволюции. [ 76 ]
Сексуальный отбор
[ редактировать ]Аллометрический показатель для высоты паруса по величине аналогичен масштабированию межвидовой длины рога до высоты плеча в шэках . Кроме того, как наблюдал Баккер (1970) в контексте Диметродона , многие виды ящериц поднимают дорсальный гребень кожи во время угроз и ухаживания, а положительно аллометрические, сексуально диморфные изделия и росы присутствуют в существующих ящерицах (Echelle et al. 1978; Christian et al. Существуют также доказательства сексуального диморфизма как в надежности скелета, так и на относительной высоте шипов D. limbatus (Romer and Price 1940). [ 76 ]
Сексуальный диморфизм
[ редактировать ]Диметродон , возможно, был сексуально диморфным , что означает, что у мужчин и женщин были немного разные размеры тела. Некоторые образцы Dimetrodon были предполагаются как мужчины, потому что у них более толстые кости, более крупные парусы, более длинные черепа и более выраженные верхнечелюстные «шаги», чем другие. Основываясь на этих различиях, монтированные скелеты в Американском музее естественной истории (AMNH 4636) и Полевой музей естественной истории могут быть мужчины и скелеты в музее природы и науки в Денвере ( MCZ 1347) и Музей Мичиганского университета естественной истории может быть женщинами. [ 44 ]


Палеоэкология
[ редактировать ]
Окаменелости Диметродона известны из Соединенных Штатов (Техас, Оклахома, Нью -Мексико, Аризона, Юта и Огайо), Канады ( остров Принца Эдуарда ) и Германии, районы, которые были частью суперконтинента Еврамерики во время раннего Пермья. В Соединенных Штатах почти все материалы, приписываемые Диметродону, поступили из трех геологических групп в северо-центральной части Техаса и юго-центральной Оклахомы: Группа Clear Fork , группа Вичито и Группа реки Пиз . [ 78 ] [ 79 ] Большинство ископаемых находок являются частью низменных экосистем, которые во время перми были бы обширными водно -болотными угодьями. В частности, красные ложе Техаса представляют собой область большого разнообразия ископаемых тетрапод или четырехногленных позвоночных. В дополнение к Диметродону , наиболее распространенными тетраподами в красных слоях и во всех ранних пермских месторождениях на юго -западе Соединенных Штатов являются амфибии Аржерия , Диплокаус , Эропс и Тримерхахис , рептилиоморф Сеймур , рептилийский каптор и синапсоиды и , рептилий и эдафосавр синапсои Полем Эти тетраподы составляли группу животных, которые палеонтолог Эверетт С. Олсон назвал «пермо-углевороночной хронофуной», фауной , которая доминировала в континентальной еврамериканской экосистеме в течение нескольких миллионов лет. [ 80 ] Основываясь на геологии месторождений, таких как красные кровати, фауна, как полагают, населяла хорошо вегетативную низменную дельтовую экосистему . [ 81 ]
Продовольственная сеть
[ редактировать ]
Олсон сделал много выводов о палеоэкологии Техасских красных слоев и роли Диметродона в его экосистеме. Он предложил несколько основных типов экосистем, в которых жили самые ранние тетраподы. Диметродон принадлежал к наиболее примитивной экосистеме, которая разработала из водных пищевых сетей. В нем водные растения были основными производителями и в значительной степени питались рыбами и водными беспозвоночными. Большинство земельных позвоночных питались этими водными первичными потребителями. Диметродон , вероятно, был лучшим хищником экосистемы красных слоев, питаясь различными организмами, такими как акула Xenacanthus [ Цитация необходима ] , водные амфибии Trimerorhachis и Diplocaulus , а также наземные тетраподы Seymouria и Trematops . Насекомые известны из ранних пермских красных слоев и, вероятно, были вовлечены в некоторой степени в той же пищевой паутине, что и Dimetrodon , кормив небольшие рептилии, такие как Captorhinus . Сборка красных кроватей также включали некоторые из первых крупных землевладельческих травоядных животных, таких как эдафозавра и диадекты . Кормируя в первую очередь на наземных растениях, эти травоядные не получали свою энергию из водных пищевых сетей. По словам Олсона, лучшим современным аналогом для населенной диметродона экосистемы является Эверглейдс . [ 81 ] Точный образ жизни диметродона (амфибия на наземный) долгое время был противоречивым, но костная микроанатомия поддерживает наземный образ жизни, [ 82 ] что подразумевает, что он питался в основном на суше, на берегах или в очень мелкой воде. Существуют также доказательства того, что Dimetrodon, охотящаяся за эфивацией Diplocaulus во времена засухи, с тремя частично съеденными юношескими диплокаусом в норе из восьми нормовых знаков зубов от диметродона , которые раскопали и убили их. [ 83 ]
Единственным видом диметродона, найденным за пределами юго -запада Соединенных Штатов, является D. Teutonis из Германии. Его останки были обнаружены в формации Тамбаха в ископаемом участке, называемом местностью Bromacker. Собрание Bromacker ранних пермских тетрапод необычна в том смысле, что существует мало синапсидов с крупными телами, служащих роли лучших хищников. D. Teutonis , по оценкам, составлял всего 1,7 метра (5,6 фута) в длину, слишком мал, чтобы добыть большие травоядные диадектиды , которые обильны в сборке Bromacker. Это, скорее всего, съела мелких позвоночных и насекомых. Только три окаменелости могут быть связаны с большими хищниками, и, как полагают, они были либо большими варанопидами , либо небольшими сфенакодонтами , которые могли бы потенциально добывать D. teutonis . В отличие от низменных дельтовых красных слоев Техаса, считается, что отложения Bromacker представляют собой нагорную среду без водных видов. Возможно, что плотоядные животные с крупным телом не были частью сборки Bromacker, потому что они зависели от большого водного водного амфибии для еды. [ 12 ]
Смотрите также
[ редактировать ]- Secodontosaurus - вымерший род синапсидов
- Динозавры в парке Юрского периода
- Edaphosaurus - вымерший род синапсидов
Ссылки
[ редактировать ]- ^ "Диметродон" . Lexico UK English Dictionary . Издательство Оксфордского университета . Архивировано из оригинала 2020-03-22.
- ^ "Диметродон" . Dictionary.com unabridged (онлайн). н.д. Получено 2018-02-12 .
- ^ "Диметродон" . База данных палеобиологии . Архивировано с оригинала 2 октября 2013 года . Получено 23 августа 2012 года .
- ^ Подпрыгнуть до: а беременный в дюймовый Angielczyk, KD (2009). «Диметродон не динозавр: использование деревьев, чтобы понять древних родственников млекопитающих и их эволюцию» . Эволюция: образование и охват . 2 (2): 257–271. doi : 10.1007/s12052-009-0117-4 .
- ^ Huttenlocker, AK; Rega, E. (2012). «Палеобиология и костная микроструктура синапсидов Pelycosauriangrade». В чинэми, А. (ред.). Пероруннеры млекопитающих: радиация, гистология, биология . Издательство Университета Индианы. С. 90–119. ISBN 978-0-253-35697-0 .
- ^ «Известные доисторические животные, которые на самом деле не были динозаврами» . 17 февраля 2021 года. Архивировано из оригинала 2021-04-07.
- ^ Черный, Райли. «Диметродон в вашем семейном древе» . Смитсоновский журнал . Получено 2021-12-05 .
- ^ Cloudsley-Thompson, JL (2005-01-19). Экология и поведение мезозойских рептилий . Springer Science & Business Media. ISBN 978-3-540-22421-1 .
- ^ Фиеста, Энрике; Дэвидсон, Джон (2015-01-10). Dimetrodon - Permian Predator . Мендон Коттедж книги. ISBN 978-1-310-19617-1 .
- ^ Зачос, Фрэнк; Ашер, Роберт (2018-10-22). Эволюция млекопитающих, разнообразие и систематика . Walter de Gruyter Gmbh & Co Kg. ISBN 978-3-11-034155-3 .
- ^ Гонет, Джордан; Бардин, Жреми; Джирондот, Марк; Хатчинсон, Джон Р.; Лаурин, Мишель (2023). «Разрушение постурального разнообразия млекопитающих: вклад перекрестных сечений в палеобиологических выводах» . Журнал эволюции млекопитающих . 30 (2): 321–337. doi : 10.1007/s10914-023-09652-w . S2CID 256788973 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон Берман, DS; Рейш, RR; Martens, T.; Henrici, AC (2001). «Новый вид диметродона (Synapsida: Sphenacodontidae) из нижнего Пермского Германии записей, первого появления рода за пределами Северной Америки» (PDF) . Канадский журнал наук о Земле . 38 (5): 803–812. Bibcode : 2001cajes..38..803b . doi : 10.1139/cjes-38-5-803 .
- ^ Подпрыгнуть до: а беременный в дюймовый Fröbisch, J.; Schoch, RR; Müller, J.; Schindler, T.; Schweiss, D. (2011). «Новая базальная синапсида сфенакодонтид из позднего каменноугольного бассейна саара-Нахе, Германия» (PDF) . Acta Palaeontologica Polonica . 56 (1): 113–120. doi : 10.4202/app.2010.0039 . S2CID 45410472 .
- ^ Олсон, ЕС; Beerbower, Jr (1953). «Формирование Сан -Анджело, Перми из Техаса и его позвоночные». Журнал геологии . 61 (5): 389–423. Bibcode : 1953jg ..... 61..389o . doi : 10.1086/626109 . S2CID 128681671 .
- ^ Олсон, ЕС (1955). «Параллелизм в эволюции пермских рептилийских фаун старых и новых миров» . ФИЛИАНА . 37 (13): 385–401.
- ^ Стернберг, CW (1942). «Скелет незрелого пеликозавра, диметродон ср. Грандис , от Пермия из Техаса». Журнал палеонтологии . 16 (4): 485–486. JSTOR 1298848 .
- ^ Лаурин, М.; Reisz, RR (2012). «Синапсида: млекопитающие и их вымершие родственники» . Веб -проект Tree of Life . Архивировано из оригинала 7 декабря 2012 года . Получено 24 августа 2012 года .
- ^ «Выставленные образцы: Диметродон» . Американский музей естественной истории . Архивировано из оригинала 4 июля 2012 года . Получено 2 июля 2012 года .
- ^ Подпрыгнуть до: а беременный Baur, G.; Дело, ЕС (1899). «История Пеликозаурии, с описанием рода Dimetrodon , Cope». Труды Американского философского общества . 20 (1): 5–62. doi : 10.2307/1005488 . HDL : 2027/UC1.32106020416696 . JSTOR 1005488 .
- ^ Подпрыгнуть до: а беременный Abler, WL 2001. Керф-и бурная модель зубных зубцов тиранозавра. п. 84-89. В: Жизнь мезозойских позвоночных . Ed.S Tanke, DH, Carpenter, K., Skrepnick, MW Indiana University Press.
- ^ "Великая энергетическая вызов" . Среда .
- ^ Махо, чай; Махо, Сиги; Скотт, Дайан; Рейс, Роберт Р. (19 августа 2022 г.). «Пермский гиперкарновор предполагает сложности стоматологии среди ранних амниотов» . Природная связь . 13 (1): 4882. BIBCODE : 2022NATCO..13.4882M . doi : 10.1038/s41467-022-32621-5 . PMC 9391490 . PMID 35986022 .
- ^ Кемп, Т.С. (2006). «Происхождение и раннее излучение рептилий, подобных млекопитающим,: палеобиологическая гипотеза» . Журнал эволюционной биологии . 19 (4): 1231–1247. doi : 10.1111/j.1420-9101.2005.01076.x . PMID 16780524 . S2CID 3184629 .
- ^ Лаурин, М.; Reisz, RR (1997). «Аутипоморфии основных клад синапсидов» . Веб -проект Tree of Life . Получено 24 августа 2012 года .
- ^ Ромер, как (1927). «Примечания о пермо-карбоновой рептилийской диметродоне ». Журнал геологии . 35 (8): 673–689. Bibcode : 1927jg ..... 35..673r . doi : 10.1086/623462 . JSTOR 30060393 . S2CID 140679339 .
- ^ Подпрыгнуть до: а беременный Рега, EA; Noriega, K.; Sumida, SS; Huttenlocker, A.; Ли, А.; Кеннеди, Б. (2012). «Зажившие переломы в нейронных шипах связанного скелета диметродона: последствия для морфологии и функции дорсального паруса». Fieldiana Life and Earth Sciences . 5 : 104–111. doi : 10.3158/2158-5520-5.1.104 . S2CID 108887164 .
- ^ Подпрыгнуть до: а беременный в Rega, E.; Sumida, S.; Noriega, K.; Пелл, C.; Ли А. (2005). «Палеопатология, основанная на фактических данных: онтогенетические и функциональные последствия дорсальных парусов в Диметродоне ». Журнал палеонтологии позвоночных . 25 (S3): 103а. doi : 10.1080/02724634.2005.10009942 . S2CID 220413556 .
- ^ Sumida, S.; Rega, E.; Noriega, K. (2005). «Палеопатология, основанная на фактических данных: влияние на филогенетический анализ рода Dimetrodon ». Журнал палеонтологии позвоночных . 25 (S3): 120a. doi : 10.1080/02724634.2005.10009942 . S2CID 220413556 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин Huttenlocker, AK; Rega, E.; Sumida, SS (2010). «Сравнительная анатомия и остеогистология гиперлонгевых нервных шипов в сфенакодонтидах Sphenacodon и Dimetrodon (Amniota: Synapsida)». Журнал морфологии . 271 (12): 1407–1421. doi : 10.1002/jmor.10876 . PMID 20886514 . S2CID 40899700 .
- ^ Рега, EA; Noriega, K.; Sumida, S.; Ли, А. (2004). «Гистологический анализ травматического повреждения множества нервных шипов сопутствующего скелета диметродона : последствия для заживления ответа, морфологии дорсального паруса и возраста в смерти в более низкой пермской синапсиде». Интегративная и сравнительная биология . 44 : 628.
- ^ Konstantinovich Cchudinov, Petr (1965). «Новые факты о фауне верхнего пермца СССР». Журнал геологии . 73 (1): 117–30. Bibcode : 1965jg ..... 73..117c . doi : 10.1086/627048 . S2CID 129428120 .
- ^ Botha-Brink, J.; Modesto, SP (2007). «Агрегация« Пеликозавр »смешанного возраста из Южной Африки: самые ранние доказательства родительской помощи в амниотах?» Полем Труды Королевского общества б . 274 (1627): 2829–2834. doi : 10.1098/rspb.2007.0803 . PMC 2288685 . PMID 17848370 .
- ^ Niedźwiedzki, G.; Bojanowski, M. (2012). «Предполагаемое впечатление от тела EupelyCosaur от раннего пермца внутрисудетического бассейна, Польша». Ichnos . 19 (3): 150–155. Bibcode : 2012ichno..19..150n . doi : 10.1080/10420940.2012.702549 . S2CID 129567176 .
- ^ Spindler, Frederik; Вернебург, Ральф; Schneider, Joerg W.; Лутардт, Людвиг; Аннакер, Волкер; Rößler, Ronny (2018). «Первое древесное« Пеликозавр »(Synapsida: Varanopidae) из раннего пермского ископаемого химического отложения, SE Germany, с обзором филогения варанопидов». Палц 92 (2): 315–364. Bibcode : Palatinate 2018 ... 92..315s . Doi : 10.1007/s12542-018-0405-9 . S2CID 133846070 .
- ^ Модесто, Шон П. (январь 2020 г.). «Укоренение об отношениях рептилий» . Природа экология и эволюция . 4 (1): 10–11. doi : 10.1038/s41559-019-1074-0 . ISSN 2397-334X . PMID 31900449 . S2CID 209672518 .
- ^ Форд, Дэвид П.; Бенсон, Роджер Б.Дж. (2019). «Передописание маярума Оровентора (Sauropsida, Diapsida) с использованием μCT с высоким разрешением, и последствия для ранней филогения амниот» . Документы по палеонтологии . 5 (2): 197–239. Bibcode : 2019ppal .... 5..197f . doi : 10.1002/spp2.1236 . ISSN 2056-2802 . S2CID 92485505 .
- ^ Spalding, Dae (1995). « Батинатус , первый в Канаде« динозавр ». В Сарджанте был (ред.). Окаменелости позвоночных и эволюция научных концепций . Тейлор и Фрэнсис США. С. 245–254. ISBN 2881249965 .
- ^ Spalding, Dae (1995). « Батинатус , первый в Канаде« динозавр ». В Сарджанте был (ред.). Окаменелости позвоночных и эволюция научных концепций . Тейлор и Фрэнсис США. С. 245–254. ISBN 2881249965 .
- ^ «Канакозавра! Первым канадским« динозавром »становится Dimetrodon Borealis» . Phys.org .
- ^ Мэтью, WD (1908). «Обзор" пересмотра Пеликозаурии Северной Америки " . Наука . 27 (699): 816–818. Bibcode : 1908sci .... 27..816m . doi : 10.1126/science.27.699.816 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час я Дело, ЕС (1907). Пересмотр Пеликозаурии Северной Америки . Вашингтон, округ Колумбия: Карнеги Институт Вашингтона. С. 1 –176.
- ^ Подпрыгнуть до: а беременный в дюймовый и Коуп, Эд (1878). «Описания вымершей Батрахии и рептилии от пермской формирования Техаса». Труды Американского философского общества . 17 (101): 505–530. JSTOR 982652 .
- ^ Подпрыгнуть до: а беременный Коуп, Эд (1877). «Описания вымерших позвонков из пермских и триасовых образов Соединенных Штатов». Труды Американского философского общества . 17 (100): 182–193. JSTOR 982295 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час Ромер, как; Прайс, Ли (1940). «Обзор Пеликозаурии». Геологическое общество Америки Специальная статья . Геологическое общество Америки Специальные документы. 28 : 1–538. doi : 10.1130/spe28-p1 .
- ^ Вон, стр. (1966). «Сравнение ранних пермских фаун позвоночных в регионе четырех углов и северо-центрального Техаса» (PDF) . Вклад в науку. 105 Музей естественной истории округа Лос -Анджелес: 1–13. Архивировано из оригинала (PDF) 2017-01-10 . Получено 2016-04-21 .
{{cite journal}}: CITE Journal требует|journal=( помощь ) - ^ Вон, стр. (1969). «Ранние пермские позвоночные из южного Нью -Мексико и их палеозоогеографическое значение» (PDF) . Вклад в науку. 166 Музей естественной истории округа Лос -Анджелес: 1–22. Архивировано из оригинала (PDF) 2017-01-10 . Получено 2016-04-21 .
{{cite journal}}: CITE Journal требует|journal=( помощь ) - ^ Олсон, Эверетт С; Олсон, Эверетт С. (1970-02-05). «Trematops Stonei sp. Nov. (Temnospondyli Amphibia) из Вашингтонской формирования, Dunkard Group, штат Огайо» . Kirtlandia . 8 : 1–12.
- ^ Олсон, ЕС (1975). «Пемо-карбоновая палеоэкология и морфотипические серии» . Американский зоолог . 15 (2): 371–389. doi : 10.1093/icb/15.2.371 .
- ^ Подпрыгнуть до: а беременный в Лукас, Спенсер Г. (2013-11-01). «Биостратиграфия позвоночных и биохронология Верхней палеозойской группы Данкарда, Пенсильвания - Западная Вирджиния - Охио, США» . Международный журнал геологии угля . SI: Dunkard Group Geology. 119 : 79–87. doi : 10.1016/j.coal.2013.04.007 . ISSN 0166-5162 .
- ^ Подпрыгнуть до: а беременный в Берман, Д.С. (1977). «Новый вид диметродона (Reptilia, Pelycosauria) из не-дельтаических фаций в нижнем пермине северо-центральной части Нью-Мексико». Журнал палеонтологии . 51 (1): 108–115. JSTOR 1303466 .
- ^ Подпрыгнуть до: а беременный в Мадалена, К.; Успех, с.; Zeigler, K.; Rega, E. (2007). Lator Nower Cittery of Jemez Publo, северо-центральный Нью-Мексико » А 27 (3, приложение): 110а. doi : 10.1080/ 02724634.1007.1010458
- ^ Вон, стр. (1973). Позвоночные из группы Катлера долины и окрестности (PDF) . Руководство по геологическому обществу Нью -Мексико. Тол. 24. Геологическое общество Нью -Мексико. С. 99–105.
- ^ Коуп, изд (1881). «На какой -то новой Batrachia и рептилиях от пермских красных кроватей Техаса». Бюллетень Геологической службы США . 6 : 79–82.
- ^ Дело, ЕС (1903). «Остеология эмболофора Долловануса , Коуп, с попыткой восстановления» . Журнал геологии . 11 (1): 1–28. Bibcode : 1903jg ..... 11 .... 1c . doi : 10.1086/621055 . JSTOR 30056735 .
- ^ Дело, ЕС (1904). «Остеология черепа пеликозаврского рода, Dimetrodon » . Журнал геологии . 12 (4): 304–311. Bibcode : 1904jg ..... 12..304c . doi : 10.1086/621157 . JSTOR 30055825 .
- ^ Гилмор, CW (1919). «Конная скелет диметродонских гигас в Национальном музее Соединенных Штатов с заметками об анатомии скелета» (PDF) . Материалы Национального музея США . 56 (2300): 525–539. doi : 10.5479/si.00963801.56-2300.525 .
- ^ Henrici, AC; Берман, DS; Лукас, SG; Heckert, AB; Rinehart, LF; Zeigler, KE (2005). «ЗАДРЫ И ТАРСУС РАННОЙ ПЕММИАНСКИЙ СИНАПСИДИСИСИСИСИСИСИКИ СПГЕНАКОДОН ФЕРОЙК (EUPELYCOSARAIA: SPHENACODONTIDAE)» (PDF) . В Лукасе, SG; Zeigler, Ke (eds.). Не мрачный перми . Музей естественной истории и науки Нью -Мексико. Тол. 30. Альбукерке: музей естественной истории и науки Нью -Мексико. С. 106–110.
- ^ Подпрыгнуть до: а беременный в дюймовый и Ромер, как (1937). «Новые роды и виды пеликозавровских рептилий» (PDF) . Труды Зоологического клуба Новой Англии . 16 : 89–97.
- ^ Олсон, ЕС (1962). «Поздние пермские наземные позвоночные, США и СССР». Труды Американского философского общества . 52 (2): 1–224. doi : 10.2307/1005904 . JSTOR 1005904 .
- ^ Баттейл, Б. (2000). «Сравнение последних Пермских Гондванан и Лаурасиан Амниот Фауны». Журнал африканских наук о Земле . 31 (1): 165–174. Bibcode : 2000jafes..31..165b . doi : 10.1016/s0899-5362 (00) 00081-6 .
- ^ Коуп, Эд (1878). «Теломорфная рептилия» . Американский натуралист . 12 (12): 829–830. doi : 10.1086/272251 .
- ^ Коуп, Эд (1880). «Второй вклад в историю позвонков пермского образования Техаса». Труды Американского философского общества . 19 (107): 38–58. JSTOR 982605 .
- ^ Коуп, изд (1886). «Длинная выявленная тераморфа пермской эпохи» . Американский натуралист . 20 (6): 544–545. doi : 10.1086/274275 .
- ^ Коуп, изд (1882). «Третий вклад в историю позвонков пермского образования Техаса». Труды Американского философского общества . 20 (112): 447–461. JSTOR 982692 .
- ^ Уиллистон, SW (1916). «Остеология некоторых американских пермских позвоночных, II». Вклад музея Уокера . 1 : 165–192.
- ^ Рейш, RR; Берман, DS; Скотт Д. (1992). «Анатомия черепа и отношения Secodontosaurus, необычная рептилия, похожая на млекопитающие (Synapsida: Sphenacodontidae) от раннего пермица Техаса». Зоологический журнал Линневого общества . 104 (2): 127–184. doi : 10.1111/j.1096-3642.1992.tb00920.x . S2CID 56425294 .
- ^ Млекопитающие появляются
- ^ Подпрыгнуть до: а беременный Бенсон, RJ (2012). «Взаимосвязи базальных синапсидов: черепные и посткраниальные морфологические разделы предполагают различные топологии». Журнал систематической палеонтологии . 10 (2): 601–624. Bibcode : 2012jspal..10..601b . doi : 10.1080/14772019.2011.631042 . S2CID 84706899 .
- ^ RR, RR (1986). "Пеликозаврия" В Sues, H.-D. (пшеница.). Ручная масса Тол. 17а Густав Фишер Версия. стр. 1–102. ISBN 978-3-89937-032-4 Полем OL 12985656M .
- ^ Бринк, Кирстин С.; Мэддин, Хиллари С.; Эванс, Дэвид С.; Рейс, Роберт Р.; Sues, Hans-Dieter (2015). «Переоценка исторического канадского ископаемого батинхатуса Borealis от раннего пермца острова принца Эдуарда» . Канадский журнал наук о Земле . 52 (12): 1109–1120. Bibcode : 2015cajes..52.1109b . doi : 10.1139/cjes-2015-0100 .
- ^ Подпрыгнуть до: а беременный Флорид, Джорджия; Kalogirou, SA; Тассу, SA; Wrobel, L. (2001). «Природная среда и тепловое поведение диметродона Limbatus ». Журнал термической биологии . 26 (1): 15–20. Bibcode : 2001jtbio..26 ... 15f . doi : 10.1016/s0306-4565 (00) 00019-x . PMID 11070340 .
- ^ Брэмвелл, CD; Fellgett, PB (1973). «Тепловое регулирование в парусных ящерицах». Природа . 242 (5394): 203–205. Bibcode : 1973natur.242..203b . doi : 10.1038/242203a0 . S2CID 4159825 .
- ^ Подпрыгнуть до: а беременный Haack, SC (1986). «Тепловая модель плавания пеликозавра». Палеобиология . 12 (4): 450–458. Bibcode : 1986pbio ... 12..450H . doi : 10.1017/s009483730000316x . S2CID 124339088 .
- ^ Флорид, Джорджия; Wrobel, LC; Kalogirou, SA; Tassou, SA (1999). «Тепловая модель для рептилий и пеликозавров». Журнал термической биологии . 24 (1): 1–13. Bibcode : 1999jtbio..24 .... 1f . doi : 10.1016/s0306-4565 (98) 00032-1 .
- ^ Тернер, JS; Трейси, Кр (1986). «Размер тела, гомеотермия и контроль теплообмена у млекопитающих рептилий» (PDF) . В Хоттоне, Н. III.; Маклин, PD; Рот, JJ; Рот, Е.С. (ред.). Экология и биология рептилий, подобных млекопитающим . Вашингтон, округ Колумбия: Смитсоновская институциональная пресса. С. 185–194. Архивировано из оригинала (PDF) 2016-04-12 . Получено 2012-07-26 .
- ^ Подпрыгнуть до: а беременный в Томкинс, JL; Лебас, NR; Виттон, депутат; Martill, DM; Humphries, S. (2010). «Положительная аллометрия и предыстория сексуального отбора» (PDF) . Американский натуралист . 176 (2): 141–148. doi : 10.1086/653001 . PMID 20565262 . S2CID 36207 . Архивировано из оригинала (PDF) 2016-04-12.
- ^ Григг, Гордон; Nowack, Джулия; Бикудо, Хосе Эдуардо Перейра Уилкен; Бал, Нареш Чандра; Вудворд, Холли Н.; Сеймур, Роджер С. (2022). «Эндотермия всего тела: древняя, гомологичная и широко распространенная среди предков млекопитающих, птиц и крокодийцев» . Биологические обзоры . 97 (2): 766–801. doi : 10.1111/brv.12822 . HDL : 2440/134060 . ISSN 1464-7931 . PMC 9300183 . PMID 34894040 . S2CID 245021195 .
- ^ Лукас, SG; Spielmann, Ja; Rinehart, LF; Мартенс Т. (2009). Dimetrodon (Amniota: Synapsida: Sphenacodontidae) из нижнего пермского формации ABO, округ Сокорро, Нью -Мексико (PDF) . Руководство по геологическому обществу Нью -Мексико. Тол. 60. Нью -Мексико Геологическое общество. С. 281–284.
- ^ Нельсон, Джон У., Роберт В. Хук и Дэн С. Чейни (2013). Литостратиграфия нижнего пермского (Леонардиан) Формирование вилки северо-центрального Техаса из перехода на каменноугольную пермию: Бюллетень 60, изд. Спенсер Г. Лукас и соавт. Музей естественной истории и науки Нью -Мексико, стр. 286-311. Получено 28 декабря 2017 года.
- ^ Салливан, C.; Рейш, RR; Мэй, WJ (2000). «Большие дискорофоидные скелетные элементы из нижнего пермского Ричардса стимулируют трещины, штат Оклахома и их палеоэкологические последствия». Журнал палеонтологии позвоночных . 20 (3): 456–461. doi : 10.1671/0272-4634 (2000) 020 [0456: ldseft] 2.0.co; 2 . JSTOR 4524117 . S2CID 140709673 .
- ^ Подпрыгнуть до: а беременный Олсон, ЕС (1966). «Эволюция сообщества и происхождение млекопитающих». Экология . 47 (2): 291–302. Bibcode : 1966col ... 47..291o . doi : 10.2307/1933776 . JSTOR 1933776 .
- ^ Kriloff, A.; Жермен, Д.; Canoville, A.; Винсент, П.; Sache, M.; Лаурин М. (2008). «Эволюция костной микроанатомии тетраподной голени и ее использование в палеобиологическом выводе» . Журнал эволюционной биологии . 21 (3): 807–826. doi : 10.1111/j.1420-9101.2008.01512.x . PMID 18312321 . S2CID 6102313 .
- ^ «Панкрированный монстр отбил головы от древних амфибий» . Живая наука . 31 октября 2013 года.
Внешние ссылки
[ редактировать ] Определения из Wiktionary
Определения из Wiktionary  СМИ из Commons
СМИ из Commons  Таксоны от викиспецис
Таксоны от викиспецис  Данные из Wikidata
Данные из Wikidata
- Dimetrodon Palaeos Page на Dimetrodon
- Введение в веб -страницу музея палеонтологии Университета Калифорнийского университета Пеликозавров на ранних синапсидах, включая Dimetrodon
- «Диметродон: наш самый маловероятный предок» . PBS Eons . 21 августа 2017 - через YouTube .






- Пермская Германия
- Sphenacodontidae
- Цисульские синапсиды Европы
- Цисульские синапсиды Северной Америки
- Доисторические синапсидные роды
- Переходные окаменелости
- Таксоны, названные Эдвардом Дрибцом, Коуп
- Ископаемые таксоны, описанные в 1878 году
- Сначала цисульскую семью выступлений
- Цизуральскую семейную вымирание
- Вершина хищников