Мышечная клетка
| Мышечная клетка | |
|---|---|
 | |
| Подробности | |
| Расположение | Мышцы |
| Идентификаторы | |
| латинский | миоцит |
| МеШ | Д032342 |
| ТД | Х2.00.05.0.00002 |
| ФМА | 67328 |
| Анатомические термины микроанатомии | |
, Мышечная клетка также известная как миоцит , представляет собой зрелую сократительную клетку в мышце животного. [1] У человека и других позвоночных животных выделяют три типа: скелетные , гладкие и сердечные (кардиомиоциты). [2] Клетка скелетных мышц длинная, нитевидная, со множеством ядер и называется мышечным волокном . [3] Мышечные клетки развиваются из эмбриональных клеток-предшественников, называемых миобластами . [1]
Клетки скелетных мышц образуются путем слияния миобластов с образованием многоядерных клеток ( синцитий ) в процессе, известном как миогенез . [4] [5] Клетки скелетных мышц и клетки сердечной мышцы содержат миофибриллы и саркомеры и образуют поперечно-полосатую мышечную ткань . [6]
Клетки сердечной мышцы образуют сердечную мышцу в стенках камер сердца и имеют одно центральное ядро . [7] Клетки сердечной мышцы соединены с соседними клетками с помощью вставочных дисков , и когда они объединены в видимую единицу, они называются волокнами сердечной мышцы . [8]
Гладкомышечные клетки контролируют непроизвольные движения, такие как перистальтические сокращения пищевода и желудка . Гладкая мышца не имеет миофибрилл и саркомеров и поэтому не исчерчена. Гладкомышечные клетки имеют одно ядро.
Структура
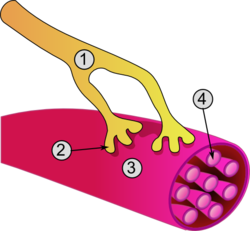
[ редактировать ]Необычная микроскопическая анатомия мышечной клетки породила ее терминологию. Цитоплазма ; называется саркоплазмой мышечной клетки гладкая эндоплазматическая сеть мышечной клетки называется саркоплазматической сетью ; а клеточная мембрана в мышечной клетке называется сарколеммой . [9] Сарколемма воспринимает и проводит раздражения.
Клетки скелетных мышц
[ редактировать ]
Клетки скелетных мышц представляют собой отдельные сократительные клетки внутри мышцы и чаще известны как мышечные волокна из-за их более длинного нитевидного вида. [10] В целом существует два типа мышечных волокон, участвующих в сокращении мышц : медленные ( тип I ) или быстрые ( тип II ).
Одна мышца, такая как двуглавая мышца плеча, у молодого взрослого мужчины содержит около 253 000 мышечных волокон. [11] Скелетные мышечные волокна — единственные мышечные клетки, которые имеют многоядерные ядра , обычно называемые миоядрами . Это происходит во время миогенеза при слиянии миобластов , каждый из которых вносит ядро во вновь образованную мышечную клетку или мышечную трубку . [12] Слияние зависит от специфичных для мышц белков, известных как фузогены, называемые миомакер и миомергер . [13]
Поперечно-полосатое мышечное волокно содержит миофибриллы, состоящие из длинных белковых цепей миофиламентов . Существует три типа миофиламентов: тонкие, толстые и эластичные, которые вместе вызывают сокращение мышц . [14] Тонкие миофиламенты состоят в основном из актина , а толстые — из миозина , и они скользят друг по другу, укорачивая длину волокон при мышечном сокращении. Третий тип миофиламентов представляет собой эластичную нить, состоящую из тайтина , очень крупного белка.
В исчерченности мышечных полос миозин образует темные нити, составляющие А. полосу Тонкие нити актина — это легкие нити, составляющие I-диапазон . Наименьшая сократительная единица волокна называется саркомером и представляет собой повторяющуюся единицу внутри двух Z-полос . Саркоплазма также содержит гликоген , который обеспечивает клетки энергией во время интенсивных физических упражнений, и миоглобин , красный пигмент, который сохраняет кислород до тех пор, пока он не понадобится для мышечной деятельности. [14]
Саркоплазматический ретикулум , специализированный тип гладкой эндоплазматической сети , образует сеть вокруг каждой миофибриллы мышечного волокна. Эта сеть состоит из групп двух расширенных концевых мешочков, называемых терминальными цистернами, и одного Т-трубочки (поперечного канальца), который проходит через клетку и выходит на другую сторону; вместе эти три компонента образуют триады , существующие в сети саркоплазматической сети, в которой каждая Т-трубочка имеет две терминальные цистерны с каждой стороны. Саркоплазматический ретикулум служит резервуаром для ионов кальция, поэтому, когда потенциал действия распространяется по Т-трубочкам, он сигнализирует саркоплазматическому ретикулуму выпустить ионы кальция из закрытых мембранных каналов для стимуляции мышечного сокращения. [14] [15]
В скелетных мышцах на конце каждого мышечного волокна наружный слой сарколеммы соединяется с сухожильными волокнами в области миосухожильного соединения . [16] [17] Внутри мышечного волокна, прижатого к сарколемме, располагаются многократно уплощенные ядра ; Эмбриологически это многоядерное состояние возникает в результате слияния нескольких миобластов с образованием каждого мышечного волокна, где каждый миобласт вносит свой вклад в одно ядро. [14]
Клетки сердечной мышцы
[ редактировать ]Клеточная мембрана клетки сердечной мышцы имеет несколько специализированных областей, которые могут включать вставочный диск и поперечные канальцы . Клеточная мембрана покрыта пластинчатой оболочкой шириной около 50 нм. Ламинарное покрытие можно разделить на два слоя; и плотная пластинка светлая пластинка . Между этими двумя слоями может находиться несколько разных типов ионов, включая кальций . [18]
Сердечная мышца, как и скелетная мышца, также поперечно-полосатая, а клетки содержат миофибриллы, миофиламенты и саркомеры, как и клетки скелетных мышц. клетки Клеточная мембрана прикреплена к цитоскелету с помощью якорных волокон шириной примерно 10 нм. Обычно они располагаются на линиях Z, образуя бороздки и отходящие поперечные канальцы. В кардиомиоцитах это образует фестончатую поверхность. [18]
Цитоскелет — это то, из чего строится остальная часть клетки, и он имеет две основные цели; первая — стабилизировать топографию внутриклеточных компонентов, а вторая — помочь контролировать размер и форму клетки. Хотя первая функция важна для биохимических процессов, последняя имеет решающее значение для определения соотношения поверхности и объема клетки. Это сильно влияет на потенциальные электрические свойства возбудимых клеток . Кроме того, отклонение от стандартной формы и размера клетки может иметь негативное прогностическое влияние. [18]
Гладкие мышечные клетки
[ редактировать ]Гладкомышечные клетки называются так потому, что они не имеют ни миофибрилл, ни саркомеров и, следовательно, не имеют исчерченности . Они обнаружены в стенках полых органов , в том числе желудка , кишечника , мочевого пузыря и матки , в стенках кровеносных сосудов , в путях дыхательной , мочевыделительной и репродуктивной систем . В глазах расширяют цилиарные мышцы и сужают радужную оболочку и изменяют форму хрусталика . В коже гладкомышечные клетки, такие как клетки, поднимающие волосы, заставляют волосы вставать дыбом в ответ на холод или страх . [19]
Гладкостмышечные клетки имеют веретенообразную форму с широкой серединой и суженными концами. Они имеют одно ядро и имеют от 30 до 200 микрометров длину . Это в тысячи раз короче волокон скелетных мышц. Диаметр их клеток также намного меньше, что устраняет необходимость в Т-трубочках, присутствующих в поперечнополосатых мышечных клетках. Хотя гладкомышечные клетки лишены саркомеров и миофибрилл, они содержат большое количество сократительных белков актина и миозина. Актиновые нити прикреплены плотными телами (похожими на Z-диски в саркомерах) к сарколемме. [19]
Разработка
[ редактировать ]Миобласт клетка - — это эмбриональная предшественник , которая дифференцируется , образуя различные типы мышечных клеток. [20] Дифференцировка регулируется миогенными регуляторными факторами , включая MyoD , Myf5 , миогенин и MRF4 . [21] GATA4 и GATA6 также играют роль в дифференцировке миоцитов. [22]
Скелетные мышечные волокна образуются при слиянии миобластов; Таким образом, мышечные волокна представляют собой клетки с несколькими ядрами , известные как миоядра , причем каждое ядро клетки происходит из одного миобласта. Слияние миобластов специфично для скелетных мышц, а не для сердечной мышцы или гладких мышц .
Миобласты в скелетных мышцах, не образующие мышечных волокон, дедифференцируются обратно в миосателлитные клетки . Эти сателлитные клетки остаются прилегающими к скелетным мышечным волокнам, расположенным между сарколеммой и базальной мембраной. [23] эндомизий . (соединительнотканная оболочка, разделяющая мышечные пучки на отдельные волокна) Чтобы повторно активировать миогенез, необходимо стимулировать сателлитные клетки к дифференцировке в новые волокна.
Миобласты и их производные, включая сателлитные клетки, теперь можно создавать in vitro посредством направленной дифференцировки плюрипотентных стволовых клеток . [24]
Киндлин-2 играет роль в удлинении развития во время миогенеза. [25]
Функция
[ редактировать ]Сокращение мышц поперечно-полосатой мышцы
[ редактировать ]
Сокращение скелетных мышц
[ редактировать ]При сокращении тонкие и толстые нити скользят друг относительно друга с помощью аденозинтрифосфата . Это сближает Z-диски вместе в процессе, называемом механизмом скользящей нити. Сокращение всех саркомеров приводит к сокращению всего мышечного волокна. Это сокращение миоцита запускается потенциалом действия над клеточной мембраной миоцита. Потенциал действия использует поперечные канальцы , чтобы проникнуть от поверхности к внутренней части миоцита, которая непрерывна внутри клеточной мембраны.Саркоплазматическая сеть представляет собой мембранные мешочки, которые соприкасаются поперечными канальцами, но остаются отдельными. Они окружают каждый саркомер и заполнены Ca. 2+ . [26]
Возбуждение миоцита вызывает деполяризацию его синапсов, нервно-мышечных соединений , что запускает потенциал действия. Благодаря единственному нервно-мышечному соединению каждое мышечное волокно получает сигнал только от одного соматического эфферентного нейрона. Потенциал действия в соматическом эфферентном нейроне вызывает высвобождение нейромедиатора ацетилхолина . [27]
Когда ацетилхолин высвобождается, он диффундирует через синапс и связывается с рецептором на сарколемме (термин, уникальный для мышечных клеток, который относится к клеточной мембране). Это вызывает импульс, который проходит через сарколемму. [28]
Когда потенциал действия достигает саркоплазматического ретикулума, он запускает высвобождение Са. 2+ из Ка 2+ каналы. Калифорния 2+ течет из саркоплазматической сети в саркомер обеими своими нитями. Это приводит к тому, что нити начинают скользить, а саркомеры становятся короче. Для этого требуется большое количество АТФ, поскольку он используется как для прикрепления, так и для высвобождения каждой головки миозина . Очень быстро, Ка 2+ активно транспортируется обратно в саркоплазматический ретикулум, что блокирует взаимодействие тонкой и толстой нити. Это, в свою очередь, заставляет мышечные клетки расслабляться. [28]
Различают четыре основных типа мышечного сокращения : изометрическое, изотоническое, эксцентрическое и концентрическое. [29] Изометрические сокращения – это сокращения скелетных мышц, которые не вызывают движения мышц. а изотонические сокращения — это сокращения скелетных мышц, которые действительно вызывают движение. Эксцентрическое сокращение – это когда мышца движется под нагрузкой. Концентрическое сокращение – это когда мышца укорачивается и генерирует силу.
Сокращение сердечной мышцы
[ редактировать ]Специализированные кардиомиоциты в синоатриальном узле генерируют электрические импульсы, которые контролируют частоту сердечных сокращений. Эти электрические импульсы координируют сокращение оставшейся сердечной мышцы через систему электропроводности сердца . Активность синоатриального узла модулируется, в свою очередь, нервными волокнами как симпатической , так и парасимпатической нервной системы. Эти системы действуют, увеличивая и уменьшая соответственно скорость выработки электрических импульсов синоатриальным узлом.
Эволюция
[ редактировать ]Эволюционное . происхождение мышечных клеток у животных широко обсуждается: одна точка зрения состоит в том, что мышечные клетки эволюционировали однажды, и, следовательно, все мышечные клетки имеют одного общего предка Другая точка зрения состоит в том, что мышечные клетки развивались более одного раза, и любое морфологическое или структурное сходство обусловлено конвергентной эволюцией и развитием общих генов, которые предшествуют эволюции мышц – даже мезодермы ( мезодерма – это зародышевый листок , дающий начало мышечные клетки у позвоночных).
Шмид и Зайпель (2005) [30] утверждают, что происхождение мышечных клеток является монофилетическим признаком, который произошел одновременно с развитием пищеварительной и нервной систем всех животных, и что это происхождение можно проследить до единственного предка многоклеточных животных, у которого присутствуют мышечные клетки. Они утверждают, что молекулярное и морфологическое сходство между мышечными клетками Cnidaria и Ctenophora настолько схоже со сходством билатерий , что у многоклеточных животных должен быть один предок, от которого произошли мышечные клетки. В этом случае Шмид и Зайпель утверждают, что последним общим предком Bilateria, Ctenophora и Cnidaria был триплобласт ( организм, имеющий три зародышевых листка), и что диплобластия , то есть организм с двумя зародышевыми листками, развилась вторично из-за их наблюдение отсутствия мезодермы или мышц, обнаруженное у большинства книдарий и гребневиков. Сравнивая морфологию книдарий и гребневиков с билатериями, Шмид и Зайпель смогли заключить, что . в щупальцах и кишечнике некоторых видов книдарий и щупальцах гребневиков имеются миобластоподобные структуры Поскольку эта структура уникальна для мышечных клеток, эти ученые на основе данных, собранных их коллегами, определили, что это маркер поперечно-полосатые мышцы, подобные тем, которые наблюдаются у билатерий. Авторы также отмечают, что мышечные клетки, обнаруженные у книдарий и гребневиков, часто оспариваются из-за того, что эти мышечные клетки происходят из эктодермы , а не из мезодермы или мезендодермы.
Другие авторы утверждают, что источником настоящих мышечных клеток является энтодермальная часть мезодермы и энтодерма. Однако Шмид и Зайпель (2005) [30] противодействовать скептицизму - относительно того, являются ли мышечные клетки, обнаруженные у гребневиков и книдарий, «настоящими» мышечными клетками, - учитывая, что книдарии развиваются через стадию медузы и стадию полипа. Они отмечают, что на стадии медузы гидрозойных существует слой клеток, отделяющийся от дистальной стороны эктодермы, который образует поперечно-полосатые мышечные клетки, аналогично мезодерме; они называют этот третий отдельный слой клеток эктокодоном . Шмид и Зайпель утверждают, что даже у билатерий не все мышечные клетки происходят из мезодермы: их ключевым примером является то, что как в глазных мышцах позвоночных, так и в мышцах спиралей эти клетки происходят из эктодермальной мезодермы, а не из энтодермальной. мезодерма. Более того, они утверждают, что, поскольку миогенез действительно происходит у книдарий с помощью тех же молекулярных регуляторных элементов, которые обнаружены в характеристиках мышечных клеток у билатерий, существуют доказательства единого происхождения поперечно-полосатых мышц. [30]
В отличие от этого аргумента в пользу единого происхождения мышечных клеток, Штейнмец, Краус и др . (2012) [31] утверждают, что молекулярные маркеры, такие как белок миозин II , используемые для определения этого единственного происхождения поперечно-полосатых мышц, предшествуют образованию мышечных клеток. Они используют пример сократительных элементов, присутствующих у Porifera или губок, у которых действительно нет поперечно-полосатой мышцы, содержащей этот белок. Кроме того, Штейнмец, Краус и др . представили доказательства полифилетического происхождения развития поперечнополосатых мышечных клеток посредством анализа морфологических и молекулярных маркеров, которые присутствуют у билатерий и отсутствуют у книдарий, гребневиков и билатерий. Штейнмец, Краус и др . показали, что традиционные морфологические и регуляторные маркеры, такие как актин , способность связывать фосфорилирование боковых цепей миозина с более высокими концентрациями положительных концентраций кальция и другие элементы MyHC , присутствуют у всех многоклеточных животных, а не только у организмов, у которых было показано наличие мышц. клетки. Таким образом, использование любого из этих структурных или регуляторных элементов для определения того, достаточно ли мышечные клетки книдарий и гребневиков схожи с мышечными клетками билатерий, чтобы подтвердить единственную линию, сомнительно, по мнению Штейнмеца, Крауса, и др . Кроме того, они объясняют, что ортологи генов Myc, которые использовались для выдвижения гипотезы о происхождении поперечно-полосатых мышц, возникли в результате события дупликации генов, которое предшествует появлению первых настоящих мышечных клеток (имеется в виду поперечнополосатая мышца), и они показывают, что гены Myc присутствуют. у губок есть сократительные элементы, но нет настоящих мышечных клеток. Штейнмец, Краус и др . также показали, что локализация этого дуплицированного набора генов, которые выполняют как функцию облегчения образования генов поперечнополосатых мышц, так и генов клеточной регуляции и движения, уже была разделена на многополосатый и немышечный MHC. Это разделение дуплицированного набора генов проявляется в локализации поперечно-полосатой части в сократительной вакуоли у губок, в то время как немышечная часть экспрессируется более диффузно во время формы и изменения клеток в процессе развития. Штейнмец, Краус и др . обнаружили аналогичную картину локализации у книдарий, за исключением книдарий N. vectensis, у которых этот маркер поперечно-полосатых мышц присутствует в гладких мышцах пищеварительного тракта. Таким образом, они утверждают, что плеизиоморфный признак отдельных ортологов Much не может быть использован для определения монофилогении мышц, а также утверждают, что присутствие поперечнополосатого мышечного маркера в гладких мышцах этого книдария указывает на принципиально иной механизм формирования мышечных клеток. Развитие и строение книдарий. [31]
Штейнмец, Краус и др . (2012) [31] далее доказывают множественное происхождение поперечнополосатых мышц у многоклеточных животных, объясняя, что ключевой набор генов, используемый для формирования тропонинового комплекса для регуляции и формирования мышц у билатерий, отсутствует у книдарий и гребневиков, а также наблюдалось 47 структурных и регуляторных белков, Штейнмец, Краус и др . не удалось обнаружить даже уникальный белок поперечно-полосатых мышечных клеток, который экспрессировался как у книдарий, так и у билатерий. Более того, Z-диск, по-видимому, развивался по-разному даже у билатерий, и даже между этой кладой существует большое разнообразие белков, что указывает на большую степень радиации мышечных клеток. Благодаря этому расхождению Z-диска Штейнмец, Краус и др . утверждают, что существует только четыре общих белковых компонента, которые присутствовали у всех мышечных предков билатерий, и что из них в качестве необходимых компонентов Z-диска только белок актин, который, как они уже утверждали, является неинформативным маркером из-за его плеизиоморфного состояния, присутствует у книдарий. В результате дальнейшего тестирования молекулярных маркеров Steinmetz et al. отмечают, что у небилатерий отсутствуют многие регуляторные и структурные компоненты, необходимые для формирования мышц билатерий, и не обнаружено какого-либо уникального набора белков как для билатерий, так и для книдарий, а также для гребневиков, которого нет у более ранних, более примитивных животных, таких как губки и гребневики. амебозои . Благодаря этому анализу авторы пришли к выводу, что из-за отсутствия элементов, от которых зависят структура и использование билатеральных мышц, небилерианные мышцы должны иметь другое происхождение и иметь другой набор регуляторных и структурных белков. [31]
Другой взгляд на аргумент: Андрику и Арноне (2015). [32] используйте недавно доступные данные о сетях регуляции генов, чтобы посмотреть, как иерархия генов и морфогенов, а также другой механизм спецификации тканей расходятся и похожи среди ранних вторичноротых и протостом. Понимая не только то, какие гены присутствуют у всех билатерий, но также время и место размещения этих генов, Андрику и Арноне обсуждают более глубокое понимание эволюции миогенеза. [32]
В своей статье Андрику и Арноне (2015) [32] утверждают, что для истинного понимания эволюции мышечных клеток функцию регуляторов транскрипции необходимо понимать в контексте других внешних и внутренних взаимодействий. В ходе своего анализа Андрику и Арноне обнаружили, что как у беспозвоночных билатерий, так и у книдарий существуют консервативные ортологи регуляторной сети генов. Они утверждают, что наличие этой общей схемы регулирования позволило в значительной степени отклониться от единой хорошо функционирующей сети. Андрику и Арноне обнаружили, что ортологи генов, обнаруженных у позвоночных, были изменены в результате различных типов структурных мутаций у вторичноротых и протостом беспозвоночных, и они утверждают, что эти структурные изменения в генах привели к значительному расхождению мышечных функций и формирования мышц у беспозвоночных. эти виды. Андрику и Арноне смогли распознать не только любые различия, обусловленные мутациями в генах, обнаруженных у позвоночных и беспозвоночных, но также интеграцию видоспецифичных генов, которые также могли вызвать отклонение от исходной функции регуляторной сети генов. Таким образом, хотя была определена общая система формирования мышечного паттерна, они утверждают, что это может быть связано с тем, что более предковая регуляторная сеть генов несколько раз использовалась в разных линиях с дополнительными генами и мутациями, вызывающими очень дивергентное развитие мышц. Таким образом, кажется, что структура миогенного паттерна может быть наследственной чертой. Однако Андрику и Арноне объясняют, что базовую структуру мышечного паттерна также следует учитывать в сочетании с цис-регуляторные элементы присутствуют на разных этапах развития. В отличие от высокого уровня структуры аппаратов семейства генов, Андрику и Арноне обнаружили, что цис-регуляторные элементы не были хорошо консервативны как во времени, так и в месте в сети, что могло указывать на большую степень дивергенции в формировании мышечных клеток. Благодаря этому анализу кажется, что миогенный GRN является предковым GRN с фактическими изменениями в миогенной функции и структуре, возможно, связанным с более поздними кооптами генов в разное время и в разных местах. [32]
Эволюционно специализированные формы скелетных и сердечных мышц предшествовали расхождению эволюционной линии позвоночных и членистоногих . [33] Это указывает на то, что эти типы мышц развились у общего предка где-то до 700 миллионов лет назад (млн лет назад) . Было обнаружено, что гладкие мышцы позвоночных развились независимо от типов скелетных и сердечных мышц.
Типы мышечных клеток беспозвоночных
[ редактировать ]Свойства, по которым различают быстрые, промежуточные и медленные мышечные волокна, могут быть разными для летательных и прыжковых мышц беспозвоночных. [34] Еще больше усложняет эту схему классификации то, что содержание митохондрий и другие морфологические свойства мышечных волокон могут меняться у мухи цеце в зависимости от физических упражнений и возраста. [35]
См. также
[ редактировать ]- Список типов клеток человека, полученных из зародышевых листков
- Список различных типов клеток в организме взрослого человека
Ссылки
[ редактировать ]- ^ Jump up to: а б Миоциты Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- ^ Брюне, Тибо; и др. (2016). «Эволюционное происхождение двухлатеральных гладких и поперечно-полосатых миоцитов» . электронная жизнь . 5 : 1. дои : 10.7554/elife.19607 . ISSN 2050-084X . ПМК 5167519 .
- ^ Саладин, Кеннет С. (2011). Анатомия человека (3-е изд.). Нью-Йорк: МакГроу-Хилл. стр. 72–73. ISBN 9780071222075 .
- ^ Скотт, В; Стивенс, Дж; Биндер-Маклеод, SA (2001). «Классификация типов волокон скелетных мышц человека» . Физиотерапия . 81 (11): 1810–1816. дои : 10.1093/ptj/81.11.1810 . ПМИД 11694174 . Архивировано из оригинала 13 февраля 2015 года.
- ^ «Кто-нибудь знает, почему волокна скелетных мышц имеют периферические ядра, а кардиомиоциты — нет? Каковы их функциональные преимущества?» . Архивировано из оригинала 19 сентября 2017 года.
- ^ Беттс, Дж. Гордон; Янг, Келли А.; Уайз, Джеймс А.; Джонсон, Эдди; По, Брэндон; Круз, Дин Х.; Король, Оксана; Джонсон, Джоди Э.; Уомбл, Марк; Дезе, Питер (6 марта 2013 г.). «Сердечная мышечная ткань» . Проверено 3 мая 2021 г.
- ^ «Мышечные ткани» . Архивировано из оригинала 13 октября 2015 года . Проверено 29 сентября 2015 г.
- ^ «Структура предсердий, волокна и проводимость» (PDF) . Проверено 5 июня 2021 г.
- ^ Саладин, Кеннет С. (2011). Анатомия человека (3-е изд.). Нью-Йорк: МакГроу-Хилл. стр. 244–246. ISBN 9780071222075 .
- ^ «Структура скелетных мышц | Обучение SEER» . Training.seer.cancer.gov .
- ^ Кляйн, CS; Марш, Джорджия; Петрелла, Р.Дж.; Райс, CL (июль 2003 г.). «Количество мышечных волокон в двуглавой мышце плеча у мужчин молодого и старого возраста». Мышцы и нервы . 28 (1): 62–8. дои : 10.1002/mus.10386 . ПМИД 12811774 . S2CID 20508198 .
- ^ Чо, Швейцария; Ли, К.Дж.; Ли, Э.Г. (август 2018 г.). «С величайшей осторожностью белки молекул стромального взаимодействия (STIM) проверяют, что делают скелетные мышцы» . Отчеты БМБ . 51 (8): 378–387. дои : 10.5483/bmbrep.2018.51.8.128 . ПМК 6130827 . ПМИД 29898810 .
- ^ Прасад, В.; Миллей, ДП (8 мая 2021 г.). «Скелетные мышечные волокна для роста рассчитывают на количество ядер» . Семинары по клеточной биологии и биологии развития . 119 : 3–10. дои : 10.1016/j.semcdb.2021.04.015 . ПМК 9070318 . ПМИД 33972174 . S2CID 234362466 .
- ^ Jump up to: а б с д Саладин, К. (2012). Анатомия и физиология: единство формы и функции (6-е изд.). Нью-Йорк: МакГроу-Хилл. стр. 403–405. ISBN 978-0-07-337825-1 .
- ^ Суги, Харуо; Абэ, Т; Кобаяши, Т; Чаен, С; Онуки, Ю; Саэки, Ю; Сугиура, С; Герреро-Эрнандес, Агустин (2013). «Усиление силы, генерируемой отдельными головками миозина в волокнах поясничной мышцы кролика со шкурой при низкой ионной силе» . ПЛОС ОДИН . 8 (5): е63658. Бибкод : 2013PLoSO...863658S . дои : 10.1371/journal.pone.0063658 . ПМЦ 3655179 . ПМИД 23691080 .
- ^ Шарве, Б; Руджеро, Ф; Ле Гельек, D (апрель 2012 г.). «Развитие миосухожильного соединения. Обзор» . Журнал «Мышцы, связки и сухожилия» . 2 (2): 53–63. ПМЦ 3666507 . ПМИД 23738275 .
- ^ Бенцингер, CF; Ван, YX; Рудницкий, Массачусетс (1 февраля 2012 г.). «Наращивание мышечной массы: молекулярная регуляция миогенеза» . Перспективы Колд-Спринг-Харбор в биологии . 4 (2): а008342. doi : 10.1101/cshperspect.a008342 . ПМК 3281568 . ПМИД 22300977 .
- ^ Jump up to: а б с Феррари, Роберто. «Здоровые и больные миоциты: метаболизм, структура и функции» (PDF) . oxfordjournals.org/en . Издательство Оксфордского университета. Архивировано из оригинала (PDF) 19 февраля 2015 года . Проверено 12 февраля 2015 г.
- ^ Jump up to: а б Беттс, Дж. Гордон; Янг, Келли А.; Уайз, Джеймс А.; Джонсон, Эдди; По, Брэндон; Круз, Дин Х.; Король, Оксана; Джонсон, Джоди Э.; Уомбл, Марк; Дезе, Питер (6 марта 2013 г.). «Гладкая мускулатура» . Проверено 10 июня 2021 г.
- ^ страница 395, Биология, пятое издание, Кэмпбелл, 1999 г.
- ^ Перри Р., Рудник М. (2000). «Молекулярные механизмы, регулирующие миогенную детерминацию и дифференцировку» . Фронт Биосци . 5 : Д750–67. doi : 10.2741/Перри . ПМИД 10966875 .
- ^ Чжао Р., Ватт А.Дж., Батл М.А., Ли Дж., Бандоу Б.Дж., Дункан С.А. (май 2008 г.). «Потеря GATA4 и GATA6 блокирует дифференцировку сердечных миоцитов и приводит к акардии у мышей» . Дев. Биол . 317 (2): 614–9. дои : 10.1016/j.ydbio.2008.03.013 . ПМЦ 2423416 . ПМИД 18400219 .
- ^ Заммит, PS; Партридж, штат Техас; Яблонка-Реувени, З (ноябрь 2006 г.). «Сателлитная клетка скелетных мышц: стволовая клетка, пришедшая с холода» . Журнал гистохимии и цитохимии . 54 (11): 1177–91. дои : 10.1369/jhc.6r6995.2006 . ПМИД 16899758 .
- ^ Чал Дж, Огинума М, Аль Танури З, Гобер Б, Сумара О, Хик А, Буссон Ф, Зидуни Ю, Мурш С, Монкуке П, Тасси О, Винсент С, Миядзаки А, Бера А, Гарнье Дж. М., Гевара Дж., Хестон М., Кеннеди Л., Хаяши С., Дрейтон Б., Черрье Т., Гайро-Морель Б., Гуссони Е., Реле Ф., Таджбахш С., Пуркье О. (август 2015 г.). «Дифференцировка плюрипотентных стволовых клеток в мышечные волокна для моделирования мышечной дистрофии Дюшенна» . Природная биотехнология . 33 (9): 962–9. дои : 10.1038/nbt.3297 . ПМИД 26237517 . S2CID 21241434 .

- ^ Доулинг Дж. Дж., Врид А. П., Ким С., Голден Дж., Фельдман Э. Л. (2008). «Киндлин-2 необходим для удлинения миоцитов и необходим для миогенеза» . BMC Клеточная Биол . 9:36 . дои : 10.1186/1471-2121-9-36 . ПМЦ 2478659 . ПМИД 18611274 .
- ^ «Строение и функции скелетных мышц» . курсы.washington.edu . Архивировано из оригинала 15 февраля 2015 года . Проверено 13 февраля 2015 г.
- ^ «Возбуждение мышечных волокон» . курсы.washington.edu . Университет Вашингтона. Архивировано из оригинала 27 февраля 2015 года . Проверено 11 февраля 2015 г.
- ^ Jump up to: а б Зисер, Стивен. «Анатомия и функции мышечных клеток» (PDF) . www.austincc.edu . Архивировано (PDF) из оригинала 23 сентября 2015 года . Проверено 12 февраля 2015 г. [ мертвая ссылка ]
- ^ Гаш, Мэтью С.; Кэндл, Патрисия Ф.; Мюррей, Ян В.; Варакалло, Мэтью (2024). «Физиология, мышечные сокращения» . СтатПерлз . Издательство StatPearls.
- ^ Jump up to: а б с Зейпель, Катя; Шмид, Волкер (1 июня 2005 г.). «Эволюция поперечно-полосатых мышц: медузы и происхождение триплобластии» . Биология развития . 282 (1): 14–26. дои : 10.1016/j.ydbio.2005.03.032 . ПМИД 15936326 .
- ^ Jump up to: а б с д Стейнмец, Патрик Р.Х.; Краус, Йоханна EM; Ларру, Клэр; Хаммель, Йорг У.; Амон-Хассензал, Аннетт; Хоулистон, Эвелин; и др. (2012). «Независимая эволюция поперечно-полосатых мышц у книдарий и билатерий» . Природа . 487 (7406): 231–234. Бибкод : 2012Natur.487..231S . дои : 10.1038/nature11180 . ПМЦ 3398149 . ПМИД 22763458 .
- ^ Jump up to: а б с д Андрику, Кармен; Арноне, Мария Ина (1 мая 2015 г.). «Слишком много способов построить мышцу: эволюция GRN, управляющих миогенезом». Зоологический обзор . Специальный выпуск: Материалы 3-го Международного конгресса по морфологии беспозвоночных. 256 : 2–13. дои : 10.1016/j.jcz.2015.03.005 .
- ^ ООта, С.; Сайто, Н. (1999). «Филогенетические взаимоотношения мышечных тканей, выведенные на основе наложения генных деревьев» . Молекулярная биология и эволюция . 16 (6): 856–867. doi : 10.1093/oxfordjournals.molbev.a026170 . ISSN 0737-4038 . ПМИД 10368962 .
- ^ Хойл, Грэм (1983). «8. Разнообразие мышечных клеток» . Мышцы и их нервный контроль . Нью-Йорк, штат Нью-Йорк: Джон Уайли и сыновья. стр. 293–299 . ISBN 9780471877097 .
- ^ Андерсон, М.; Финлейсон, Л.Х. (1976). «Влияние физических упражнений на рост митохондрий и миофибрилл летательных мышц мухи цеце, Glossina morsitans». Дж. Морфол . 150 (2): 321–326. дои : 10.1002/jmor.1051500205 . S2CID 85719905 .
Внешние ссылки
[ редактировать ] СМИ, связанные с миоцитами, на Викискладе?
СМИ, связанные с миоцитами, на Викискладе? - Структура мышечной клетки