Сонное тело
| Сонное тело | |
|---|---|
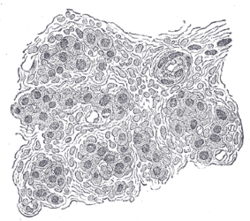 Раздел части тела сонной артерии человека. Сильно увеличено. На разрезе среди клеток видны многочисленные кровеносные сосуды. | |
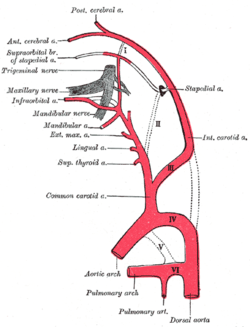 Схема, показывающая начало основных ветвей сонных артерий. | |
| Подробности | |
| нерв | Ветвь языкоглоточного нерва к каротидному синусу |
| Идентификаторы | |
| латинский | каротический клубок |
| МеШ | D002344 |
| ТА98 | А12.2.04.007 |
| ТА2 | 3886 |
| ФМА | 50095 |
| Анатомическая терминология | |
Каротидное тело представляет собой небольшое скопление хеморецепторных клеток и поддерживающих поддерживающих клеток, расположенных в местах разветвления каждой общей сонной артерии в ее наружной оболочке . [1] [2]
Каротидное тельце обнаруживает изменения в составе артериальной крови протекающей через него , главным образом парциального давления артериального кислорода , но также и углекислого газа . Он также чувствителен к изменениям pH крови и температуры .
Структура
[ редактировать ]Тело сонной артерии расположено на задней поверхности бифуркации общей сонной артерии. [3]
Каротидное тело состоит из двух типов клеток, называемых гломусными клетками : гломусные клетки I типа представляют собой периферические хеморецепторы , а гломусные клетки II типа представляют собой поддерживающие поддерживающие клетки.
- Гломусные клетки I типа происходят из нервного гребня . [4] Они высвобождают различные нейротрансмиттеры , включая ацетилхолин , АТФ и дофамин , которые запускают ВПСП в синапсированных нейронах, ведущих к дыхательному центру . Они иннервируются аксонами языкоглоточного нерва, которые вместе называются нервом сонного синуса.
- Клетки гломусного типа II напоминают глиальные клетки , экспрессируют глиальный маркер S100 и действуют как поддерживающие клетки.
Функция
[ редактировать ]Этот раздел нуждается в более надежных медицинских справках для проверки или слишком сильно полагается на первоисточники . ( октябрь 2019 г. ) |  |
Каротидное тельце функционирует как сенсор: оно реагирует на стимул, в первую очередь парциальное давление О 2 , которое обнаруживается клетками I типа (гломусными), и запускает потенциал действия через афферентные волокна языкоглоточного нерва , который передает информацию на центральную нервную систему.
Стимул
[ редактировать ]каротидного тела Периферические хеморецепторы в первую очередь чувствительны к снижению парциального давления кислорода (PO 2 ) . этом отличие от центральных хеморецепторов продолговатого мозга к изменениям рН и РСО2 ( В снижение рН и повышение РСО2 ) , которые преимущественно чувствительны . Хеморецепторы каротидных тел также чувствительны к pH и PCO2 , но лишь вторично. Более конкретно, чувствительность хеморецепторов каротидного тела к уменьшению P O 2 выше, когда pH снижается, а P CO 2 увеличивается.
Частота импульсации каротидных тел особенно чувствительна к изменению артериального РО2 в диапазоне от 60 до 30 мм рт. ст., в диапазоне, в котором насыщение гемоглобина кислородом быстро снижается. [5]
Выход каротидных телец низкий при парциальном давлении кислорода выше примерно 100 мм рт. ст. (13,3 кПа ) (при нормальном физиологическом рН), но ниже 60 мм рт. ст. активность клеток I типа (гломусных) быстро возрастает за счет снижения насыщение гемоглобина кислородом ниже 90%.
Обнаружение
[ редактировать ]Механизм обнаружения снижения P O 2 еще не идентифицирован; механизмов может быть несколько, и они могут различаться у разных видов. [6] гипоксии Было показано, что обнаружение зависит от повышенного образования сероводорода , вырабатываемого цистатионин-гамма-лиазой, поскольку обнаружение гипоксии снижается у мышей, у которых этот фермент нокаутирован или фармакологически ингибирован. Процесс обнаружения включает взаимодействие цистатионин-гамма-лиазы с гемооксигеназой-2 и образование окиси углерода . [7] Тем не менее, некоторые исследования показывают, что физиологическая концентрация сероводорода может быть недостаточно сильной, чтобы вызвать такие реакции.
Другие теории предполагают, что в этом могут участвовать митохондриальные сенсоры кислорода и гемсодержащие цитохромы, которые подвергаются обратимому одноэлектронному восстановлению во время окислительного фосфорилирования. Гем обратимо связывает O 2 со сродством, подобным сродству каротидного тельца, что указывает на то, что гем-содержащие белки могут играть роль в O 2 ; потенциально это может быть один из комплексов, участвующих в окислительно-фосфорилировании. Это приводит к увеличению количества активных форм кислорода и повышению внутриклеточного кальция. 2+ . Однако неизвестно, приводит ли гипоксия к увеличению или уменьшению количества активных форм кислорода. Роль активных форм кислорода в чувствительности к гипоксии также находится под вопросом. [8]
Кислородзависимый фермент гемоксидаза также был предложен в качестве сенсора гипоксии. При нормоксии гемоксигеназа генерирует монооксид углерода (CO), CO активирует калиевый канал с большой проводимостью, активируемый кальцием, BK. Падение CO, возникающее вследствие гипоксии, приведет к закрытию этого калиевого канала, что приведет к деполяризации мембраны и, как следствие, к активации каротидного тельца. [9] Роль «энергетического сенсора» AMP-активируемой протеинкиназы (AMPK) также была предложена в распознавании гипоксии. Этот фермент активируется во время использования чистой энергии и метаболического стресса, включая гипоксию. AMPK имеет ряд мишеней, и, по-видимому, в каротидном теле, когда AMPK активируется гипоксией, это приводит к закрытию расположенных ниже калиевых каналов как O 2 -чувствительных TASK-подобных, так и BK-каналов. [10]
Увеличение P CO 2 обнаруживается потому, что CO 2 диффундирует в клетку, где увеличивает концентрацию угольной кислоты и, следовательно, протонов . Точный механизм восприятия CO 2 неизвестен, однако было продемонстрировано, что CO 2 и низкий уровень pH ингибируют TASK-подобную калиевую проводимость, уменьшая калиевый ток. Это приводит к деполяризации клеточной мембраны, что приводит к выделению кальция. 2+ вход, возбуждение гломусных клеток и последующее высвобождение нейромедиатора. [11]
Артериальный ацидоз ( метаболический или вследствие изменения PCO Na 2 ) ингибирует переносчики кислотных оснований (например, + -ЧАС + ), которые повышают внутриклеточный pH и активируют транспортеры (например, Cl − -ОХС 3 − ), которые уменьшают его. Изменения концентрации протонов, вызванные ацидозом (или противоположностью алкалоза ) внутри клетки, стимулируют те же пути, которые участвуют в P CO 2 чувствительности .
Другой механизм – через чувствительные к кислороду калиевые каналы. Падение растворенного кислорода приводит к закрытию этих каналов, что приводит к деполяризации. Это приводит к высвобождению нейромедиатора дофамина в языкоглоточном и блуждающем нервах, идущих к вазомоторной области.
Потенциал действия
[ редактировать ]Клетки типа I (гломусные) в сонных артериях (и аортальных телах) происходят из нейроэктодермы и, таким образом, являются электрически возбудимыми. Снижение парциального давления кислорода, увеличение парциального давления углекислого газа и снижение артериального pH могут вызвать деполяризацию клеточной мембраны , и они влияют на это, блокируя калия токи . Это снижение мембранного потенциала открывает потенциалзависимые кальциевые каналы, что вызывает повышение внутриклеточной концентрации кальция. Это вызывает экзоцитоз везикул , содержащих различные нейротрансмиттеры , включая ацетилхолин , норадреналин , дофамин , аденозин , АТФ , вещество Р и метэнкефалин . Они действуют на рецепторы афферентных нервных волокон, прилегающих к гломусной клетке, вызывая потенциал действия.
Реле
[ редактировать ]Обратная связь от каротидного тела передается в кардиореспираторные центры продолговатого мозга через афферентные ветви языкоглоточного нерва . (Эфферентные волокна хеморецепторов тела аорты передаются через блуждающий нерв .) Эти центры, в свою очередь, регулируют дыхание и артериальное давление, при этом гипоксия вызывает усиление вентиляции.
Клиническое значение
[ редактировать ]
Параганглиома
[ редактировать ]Параганглиома – это опухоль, которая может поражать тело сонной артерии и обычно является доброкачественной . В редких случаях злокачественная нейробластома может исходить из тела сонной артерии.
См. также
[ редактировать ]Список различных типов клеток в организме взрослого человека
Ссылки
[ редактировать ]- ^ «Сонное тело и каротидный синус – общие сведения» . Протоколы Айовы для головы и шеи. Medicine.uiowa.edu . Проверено 23 октября 2019 г.
- ^ Холл, Джон Эдвард (20 мая 2015 г.). Учебник Гайтона и Холла по медицинской физиологии (13-е изд.). Филадельфия, Пенсильвания. ISBN 978-1-4557-7005-2 . OCLC 900869748 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Кадасне, ДК (2009). Учебник анатомии Кадасне (1-е изд.). Нью-Дели: Медицинские издательства Jaypee Brothers. п. 916. ИСБН 978-81-8448-455-7 . OCLC 682534511 .
- ^ Гонсалес С., Альмарас Л., Обесо А., Ригуаль Р. (1994). «Хеморецепторы каротидного тела: от естественных раздражителей к сенсорным разрядам». Физиол. Преподобный . 74 (4): 829–98. дои : 10.1152/physrev.1994.74.4.829 . ПМИД 7938227 .
- ^ Холл, Джон Эдвард (20 мая 2015 г.). Учебник Гайтона и Холла по медицинской физиологии (13-е изд.). Филадельфия, Пенсильвания. ISBN 978-1-4557-7005-2 . OCLC 900869748 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Уорд Дж.П. (2008). «Датчики кислорода в контексте» . Биохим Биофиз Акта . 1777 (1): 1–14. дои : 10.1016/j.bbabio.2007.10.010 . ПМИД 18036551 .
- ^ Пэн Ю.Дж., Нандури Дж., Рагураман Г., Суваннакитти Д., Гадалла М.М., Кумар Г.К., Снайдер Ш.Х., Прабхакар Н.Р. (2010). H2S опосредует чувствительность O2 в каротидном теле PNAS 107 (23) 10719-10724. дои : 10.1073/pnas.1005866107
- ^ Гонсалес К., Санс-Альфайате Г., Агапито М.Т., Гомес-Ниньо А., Роше А., Обесо А. (2002). «Датчики кислорода в контексте» . Дыхание Физиол Нейробиол . 132 (1): 17–41. дои : 10.1016/S1569-9048(02)00047-2 . ПМИД 12126693 . S2CID 25674998 .
- ^ Уильямс С.Э., Вуттон П., Мейсон Х.С., Боулд Дж., Айлс Д.Е., Риккарди Д., Пирс С., Кемп П.Дж. (2004). «Гемоксигеназа-2 — сенсор кислорода для кальций-чувствительного калиевого канала» . Наука . 306 (5704): 2093–7. Бибкод : 2004Sci...306.2093W . дои : 10.1126/science.1105010 . ПМИД 15528406 . S2CID 41811182 .
- ^ Вятт К.Н., Мастард К.Дж., Пирсон С.А., Даллас М.Л., Аткинсон Л., Кумар П., Пирс С., Харди Д.Г., Эванс А.М. (2007). «АМФ-активируемая протеинкиназа опосредует возбуждение сонных артерий вследствие гипоксии» . J Биол Хим . 282 (11): 8092–8. дои : 10.1074/jbc.M608742200 . ПМЦ 1832262 . ПМИД 17179156 .
- ^ Баклер К.Дж., Уильямс Б.А., Оноре Э. (2000). «TASK-подобный фоновый калиевый канал, чувствительный к кислороду, кислоте и анестезии, в артериальных хеморецепторных клетках крысы» . Дж. Физиол . 525 (1): 135–142. дои : 10.1111/j.1469-7793.2000.00135.x . ПМК 2269923 . ПМИД 10811732 .