Диметродон
| Диметродон | |
|---|---|

| |
| Скелет D.limbatus , Государственный музей естественной истории Карлсруэ. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Сторона синапса |
| Семья: | † Сфенакодонтиды |
| Подсемейство: | † Сфенакодонтины |
| Род: | † Диметродон Коуп , 1878 г. |
| Типовой вид | |
| † Диметродон лимбатус Коуп, 1877 г.
| |
| Разновидность | |
|
См . ниже | |
| Синонимы | |
|
Синонимия рода Синонимия видов | |
Диметродон ( / d aɪ ˈ m iː t r ə ˌ d ɒ n / [ 1 ] или / - это aɪ ˈ m ɛ t ə r ˌ и ɒ n / ; [ 2 ] горит. ' две меры зубов ' ) — вымерший род не млекопитающих синапсид , принадлежащий к семейству Sphenacodontidae , живший в приуральскую эпоху ранней перми . , около 295–272 миллионов лет назад [ 3 ] [ 4 ] [ 5 ] Поскольку длина большинства видов составляет 1,7–4,6 м (5,6–15,1 фута) и вес 28–250 кг (62–551 фунт), наиболее заметной особенностью диметродона является большой парус нервного отростка на его спине, образованный удлиненными шипами, идущими от позвонки . Это было облигатное четвероногое животное (оно могло ходить только на четырех ногах) и имело высокий изогнутый череп с крупными зубами разной величины, расположенными вдоль челюстей. Большинство окаменелостей было найдено на юго-западе Соединенных Штатов , большинство из них происходит из геологических месторождений, называемых Красными пластами Техаса и Оклахомы . Совсем недавно его окаменелости были также найдены в Германии было названо более дюжины видов , и с тех пор, как этот род был впервые установлен в 1878 году, .
Диметродона часто принимают за динозавра или за современника динозавров в массовой культуре, но он вымер примерно за 40 миллионов лет до появления динозавров. [ 6 ] [ 7 ] по внешнему виду и физиологии похож на рептилий, Хотя диметродон он гораздо более тесно связан с млекопитающими, чем с рептилиями, хотя и не является прямым предком млекопитающих. [ 4 ] Диметродон отнесен к «синапсидам немлекопитающих», группе, которую традиционно, но ошибочно, называют «рептилиями, подобными млекопитающим». [ 4 ] но теперь известны как стволовые млекопитающие. Это группирует диметродонов вместе с млекопитающими в кладе Synapsida, тогда как рептилии помещаются в отдельную кладу Sauropsida . Одиночные отверстия в черепе за каждым глазом, известные как височные окна , и другие особенности черепа отличают диметродона и настоящих млекопитающих от большинства самых ранних зауропсидов .
Диметродон, вероятно, был одним из высших хищников экосистем Приуралья, питаясь рыбой и четвероногими , включая рептилий и земноводных . Меньшие виды Dimetrodon, возможно, играли разные экологические роли . Парус Диметродона, возможно, использовался для стабилизации его позвоночника или для обогрева и охлаждения его тела в качестве формы терморегуляции . [ 8 ] Некоторые недавние исследования утверждают, что парус был бы неэффективен для отвода тепла от тела из-за того, что крупные виды были обнаружены с маленькими парусами, а мелкие виды были обнаружены с большими парусами, что по существу исключает регулирование тепла как его основную цель. Парус, скорее всего, использовался для демонстрации ухаживания , в том числе для отпугивания соперников или хвастовства перед потенциальными партнерами. [ 9 ] [ 10 ]
Описание
[ редактировать ]
Диметродон был четвероногим синапсидом с парусной спинкой, который, скорее всего, имел полураспростертую позу между млекопитающим и ящерицей, а также мог ходить в более вертикальной стойке, отрывая тело и большую часть или весь хвост от земли. [ 11 ] Длина большинства видов диметродонов варьировалась от 1,7 до 4,6 м (от 6 до 15 футов) и, по оценкам, весила от 28 до 250 кг (от 60 до 550 фунтов). [ 12 ] Самый маленький из известных видов D. teutonis имел длину около 60 см (24 дюйма) и весил 14 кг (31 фунт). [ 12 ] [ 13 ] Более крупные виды Dimetrodon были одними из крупнейших хищников ранней перми, хотя близкородственный Tappenosaurus , известный по фрагментам скелета в несколько более молодых породах, возможно, был еще крупнее и имел длину около 5,5 метров (18 футов). [ 14 ] [ 15 ] Хотя некоторые виды диметродонов могут достигать очень больших размеров, известно множество молодых экземпляров. [ 16 ]
Череп
[ редактировать ]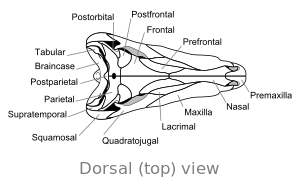 |
 |
 |
 |
Единственное большое отверстие по обе стороны задней части черепа связывает диметродона с млекопитающими и отличает его от большинства самых ранних зауропсидов, у которых либо отсутствуют отверстия , либо есть два отверстия . Такие особенности, как гребни на внутренней стороне носовой полости и гребень в задней части нижней челюсти, считаются частью эволюционного прогресса от ранних наземных позвоночных с четырьмя конечностями до млекопитающих .
Череп диметродона высокий и сжатый с боков или из стороны в сторону. Глазницы расположены высоко и далеко назад в черепе. Позади каждой глазницы есть единственное отверстие, называемое подвисочным окном . Дополнительное отверстие в черепе, надвисочное окно , можно увидеть, если смотреть сверху. Задняя часть черепа ( затылок ) ориентирована под небольшим углом вверх, что характерно для всех других ранних синапсидов . [ 17 ] Верхний край черепа спускается вниз по выпуклой дуге к кончику морды. Кончик верхней челюсти, образованный предчелюстной костью, приподнят над частью челюсти, образованной верхнечелюстной костью, образуя верхнечелюстную «ступеньку». На этом этапе образуется диастема – разрыв в зубном ряду. Его череп был более массивным, чем череп динозавра.
Зубы
[ редактировать ]Размер зубов сильно варьируется по длине челюстей, что и дало диметродону его название, что означает «два размера зуба» в отношении набора маленьких и больших зубов. [ 18 ] Одна или две пары клыкообразных (больших, заостренных, клыкообразных зубов) отходят от верхней челюсти. Крупные резцы имеются также на кончиках верхней и нижней челюстей, укорененные в предчелюстных и зубных костях . Мелкие зубы присутствуют вокруг верхнечелюстной «ступени» и позади клыкообразных, а дальше в челюсти они становятся меньше. [ 19 ]

Многие зубы наиболее широки в средней части и сужаются ближе к челюстям, что придает им вид капли. Зубы каплевидной формы уникальны для диметродона и других близкородственных сфенакодонтид , что помогает отличить их от других ранних синапсидов. [ 13 ] Как и у многих других ранних синапсидов , зубы большинства видов Dimetrodon имеют зазубрины по краям. [ 13 ] Зазубрины зубов диметродона были настолько тонкими, что напоминали крошечные трещинки. [ 20 ] У динозавра Альбертозавра были такие же трещинообразные зубцы, но в основании каждого зубца была круглая полость , которая должна была распределять силу по большей площади поверхности и предотвращать распространение трещины по зубу из-за напряжений, вызванных питанием. . В отличие от альбертозавра , зубы диметродона не имели приспособлений, которые предотвращали бы образование трещин на их зазубринах. [ 20 ] На зубах D. teutonis нет зазубрин, но они все же имеют острые края. [ 13 ]
Исследование 2014 года показывает, что диметродон участвовал в гонке вооружений против своей добычи. У более мелкого вида, D. milleri , не было зубцов на зубах, поскольку он питался мелкой добычей. По мере того как добыча становилась больше, у некоторых видов диметродонов на зубах появлялись зазубрины, и они увеличивались в размерах. Например, у D.limbatus были зубцы на эмали, которые помогали ему прорезать плоть (которые были похожи на зубцы, которые можно найти у секодонтозавра ). Второй по величине вид, D. grandis , имеет зубцы, похожие на зубцы акул и тероподовых динозавров, что делает его зубы еще более специализированными для разрезания плоти. По мере того, как добыча диметродона становилась больше, различные виды в ответ увеличивались в размерах и развивали все более острые зубы. [ 21 ] Толщина и масса зубов диметродона также могли быть приспособлением для увеличения продолжительности жизни зубов. [ 22 ]
Носовая полость
[ редактировать ]На внутренней поверхности носовой части черепа имеются гребни, называемые носотурбиналами , которые, возможно, поддерживали хрящи, увеличивающие площадь обонятельного эпителия — слоя ткани, улавливающей запахи. Эти гребни намного меньше, чем у более поздних синапсидов поздней перми и триаса, большие носотрубки которых считаются свидетельством теплокровности, поскольку они, возможно, поддерживали слизистые оболочки, согревавшие и увлажнявшие поступающий воздух. Таким образом, носовая полость диметродона является переходной между таковыми ранних наземных позвоночных и млекопитающих. [ 23 ]
Челюстной сустав и ухо
[ редактировать ]Еще одной переходной особенностью диметродона является гребень в задней части челюсти, называемый отраженной пластинкой, который находится на суставной кости, которая соединяется с квадратной костью черепа, образуя челюстной сустав. У более поздних предков млекопитающих суставная и квадратная кости отделились от челюстного сустава, а суставная часть развилась в молоточек среднего уха . Отраженная пластинка стала частью кольца, называемого барабанным кольцом, которое поддерживает барабанную перепонку у всех современных млекопитающих. [ 24 ]
Хвост
[ редактировать ]
Хвост диметродона составляет большую часть общей длины тела и включает около 50 хвостовых позвонков . У первых описанных скелетов диметродона хвосты отсутствовали или были неполными . Единственными известными хвостовыми позвонками были 11, расположенные ближе всего к бедру. Поскольку эти первые несколько хвостовых позвонков быстро сужаются по мере продвижения от бедра, многие палеонтологи конца 19 - начала 20 веков считали, что у диметродона очень короткий хвост. Практически полный хвост диметродона не был описан до 1927 года. [ 25 ]
Плыть
[ редактировать ]
Парус диметродона образован удлиненными нервными отростками, выступающими из позвонков. Каждый шип различается по форме поперечного сечения от основания до кончика, что известно как «диметродонтная» дифференциация. [ 26 ] Около тела позвонка поперечное сечение позвоночника сжато с боков до прямоугольной формы, а ближе к вершине оно принимает форму восьмерки, поскольку по обеим сторонам позвоночника проходит бороздка. Считается, что форма восьмерки укрепляет позвоночник, предотвращая его изгибы и переломы. [ 27 ] Поперечное сечение позвоночника одного экземпляра Dimetrodon giganhomogenes имеет прямоугольную форму, но вблизи его центра сохраняются кольца в форме восьмерки, что указывает на то, что форма шипов может меняться с возрастом особи. [ 28 ] Микроскопическая анатомия каждого шипа варьируется от основания до кончика, указывая, где он был встроен в мышцы спины, а где он был обнажен как часть паруса. Нижняя или проксимальная часть позвоночника имеет шероховатую поверхность, которая могла бы служить точкой крепления для эпаксиальных мышц спины, а также имеет сеть соединительных тканей, называемых волокнами Шарпи, что указывает на то, что она встроена в тело. Выше, в дистальной (внешней) части позвоночника, поверхность кости более гладкая. Надкостница . , слой ткани, окружающей кость, покрыта небольшими бороздками, которые, предположительно, поддерживали кровеносные сосуды, кровоснабжающие парус [ 29 ]
Когда-то считалось, что большая борозда, проходящая по всей длине позвоночника, является каналом для кровеносных сосудов, но, поскольку кость не содержит сосудистых каналов, считается, что парус не так сильно васкуляризирован, как считалось раньше. У некоторых экземпляров диметродона сохраняются деформированные участки нервных отростков, которые кажутся зажившими переломами. Кортикальная кость , выросшая над этими разрывами, сильно васкуляризирована, что позволяет предположить, что на парусе должны были присутствовать мягкие ткани, снабжающие это место кровеносными сосудами . [ 27 ] Многослойная пластинчатая кость занимает большую часть поперечного сечения нервного отдела позвоночника и содержит линии остановки роста, которые можно использовать для определения возраста каждого человека на момент смерти. [ 30 ] У многих экземпляров D. gigashomogenes дистальные части шипов резко изгибаются, что указывает на то, что при жизни парус имел неправильный профиль. Их искривленность предполагает, что мягкие ткани, возможно, не доходили до кончиков шипов, а это означает, что перепонка паруса могла быть не такой обширной, как обычно думают. [ 26 ]
Кожа
[ редактировать ]
Никаких окаменелостей, свидетельствующих о наличии кожи диметродона , пока не обнаружено. Отпечатки кожи родственного животного, Эстемменозуха , указывают на то, что оно должно было быть гладким и хорошо снабженным железами, но эта форма кожи, возможно, не применима к Диметродону , поскольку его происхождение довольно отдаленное. [ 31 ] Диметродон также мог иметь большие щитки на нижней стороне хвоста и живота, как и у других синапсидов. [ 32 ] [ 33 ] Данные, полученные от варанопида Ascendonanus, позволяют предположить, что некоторые ранние синапсиды могли иметь чешуйки, похожие на чешуйки . [ 34 ] Однако некоторые недавние исследования показали, что варанопиды таксономически ближе к диапсидным рептилиям . [ 35 ] [ 36 ]
История классификации
[ редактировать ]Самые ранние открытия
[ редактировать ]
Самым ранним открытием окаменелостей диметродона была верхняя челюсть, обнаруженная в 1845 году человеком по имени Дональд МакЛеод, жившим в британской колонии на острове Принца Эдуарда . [ 37 ] Эти окаменелости были приобретены Джоном Уильямом Джонсоном, канадским геологом, а затем описаны Джозефом Лейди в 1854 году как нижняя челюсть Bathygnathus Borealis , крупного хищника, родственного Thecodontosaurus . [ 38 ] хотя позже в 2015 году он был реклассифицирован как вид Dimetrodon , как Dimetrodon Borealis . [ 39 ]
Первые описания Коупа
[ редактировать ]Окаменелости, теперь приписываемые диметродону, были впервые изучены американским палеонтологом Эдвардом Дринкером Коупом в 1870-х годах. Коуп получил окаменелости вместе с окаменелостями многих других пермских четвероногих от нескольких коллекционеров, которые исследовали группу скал в Техасе, называемую Красными пластами . Среди этих коллекционеров были швейцарский натуралист Джейкоб Болл , техасский геолог У. Ф. Камминс и палеонтолог-любитель Чарльз Хазелиус Штернберг . [ 40 ] Большинство образцов Коупа было отправлено в Американский музей естественной истории или в Музей Уокера Чикагского университета (большая часть коллекции окаменелостей Уокера сейчас находится в Филдовском музее естественной истории ).
Штернберг отправил несколько своих образцов немецкому палеонтологу Фердинанду Бройли в Мюнхенский университет , хотя Бройли не был так плодовит, как Коуп, в описании образцов. Соперник Коупа Отниэль Чарльз Марш также собрал несколько костей диметродона , которые он отправил в музей Уокера. [ 41 ] Первое использование названия Dimetrodon произошло в 1878 году, когда Коуп назвал виды Dimetrodon incisivus , Dimetrodon Rectiformis и Dimetrodon gigas в научном журнале Proceedings of the American Philosophical Society . [ 42 ]
Однако первое описание ископаемого диметродона появилось годом ранее, когда Коуп назвал вид Clepsydropslimbatus из Красных пластов Техаса. [ 43 ] (Название Clepsydrops было впервые предложено Коупом в 1875 году для сфенакодонтид останков из округа Вермилион, штат Иллинойс , а позже использовалось для многих образцов сфенакодонтид из Техаса; многие новые виды сфенакодонтид из Техаса были отнесены либо к Clepsydrops , либо к Dimetrodon в конце 19-го и 1870-х гг. начало 20-го века.) C.limbatus был реклассифицирован как вид Dimetrodon в 1940 году, а это означает, что статья Коупа 1877 года была первой записью Dimetrodon .
Коуп был первым, кто в своей статье 1878 года описал синапсид с парусной спинкой, назвав его C. natalis , хотя он назвал парус плавником и сравнил его с гребнями современной ящерицы-василиска ( Basilicus ). Паруса не сохранились у экземпляров D. incisivus и D. gigas , описанных Коупом в его статье 1878 года, но удлиненные шипы присутствовали у описанного им экземпляра D. Rectiformis . [ 42 ] Коуп прокомментировал назначение паруса в 1886 году, написав: «Полезность паруса трудно себе представить. Если только животное не имело водных привычек и не плавало на спине, гребень или плавник, должно быть, мешали активным движениям… Конечности недостаточно длинные, а когти недостаточно острые, чтобы демонстрировать древесные привычки, как у существующего рода Basilicus , у которого существует подобный гребень». [ 19 ]
Описания начала 20 века
[ редактировать ]
В первые несколько десятилетий 20-го века американский палеонтолог Э. К. Кейс провел множество исследований диметродона и описал несколько новых видов. Он получил финансирование от Института Карнеги для изучения многих экземпляров диметродона в коллекциях Американского музея естественной истории и нескольких других музеев. [ 41 ] Многие из этих окаменелостей были собраны Коупом, но не были подробно описаны, поскольку Коуп был известен тем, что вывел новые виды на основе всего лишь нескольких фрагментов костей.
Начиная с конца 1920-х годов палеонтолог Альфред Ромер повторно изучил многие экземпляры диметродона и назвал несколько новых видов. стал соавтором большого исследования В 1940 году Ромер вместе с Ллевеллином Айвором Прайсом под названием «Обзор пеликозаврии», в котором были повторно оценены виды диметродона, названные Коупом и Кейсом. [ 44 ] Большинство названий видов, которые Ромер и Прайс считали действительными, используются до сих пор. [ 29 ]
Новые образцы
[ редактировать ]В течение десятилетий после выхода монографии Ромера и Прайса многие экземпляры диметродона были описаны из мест за пределами Техаса и Оклахомы . Первый был описан из региона Четырех Углов штата Юта в 1966 году. [ 45 ] еще один был описан из Аризоны в 1969 году. [ 46 ] В 1975 году Олсон сообщил о материале диметродона из Огайо. [ 47 ] Новый вид диметродона под названием D. occidentalis (что означает «западный диметродон ») был назван в 1977 году из Нью-Мексико. [ 48 ] Экземпляры, найденные в Юте и Аризоне, вероятно, также принадлежат D. occidentalis . [ 49 ]
До этих открытий существовала теория, согласно которой в ранней перми среднеконтинентальный морской путь отделял то, что сейчас является Техасом и Оклахомой, от более западных земель, изолируя диметродона от небольшого региона Северной Америки, в то время как в западной части доминировал меньший сфенакодонтид, называемый сфенакодоном . Хотя этот морской путь, вероятно, действительно существовал, обнаружение окаменелостей за пределами Техаса и Оклахомы показывает, что его протяженность была ограничена и что он не был эффективным барьером для распространения диметродона . [ 48 ] [ 50 ]
В 2001 году новый вид диметродона под названием D. teutonis был описан из нижнепермского местонахождения Бромакер в Тюрингенском лесу в Германии, что расширило географический ареал диметродона за пределы Северной Америки. впервые [ 12 ]
Разновидность
[ редактировать ]
Двадцать видов диметродона был впервые описан в 1878 году . были названы с тех пор, как этот род Многие из них были синонимами более старых названных видов, а некоторые теперь принадлежат к разным родам.
Краткое содержание
[ редактировать ]| Разновидность | Власть | Расположение | Статус | Синонимы | Изображения |
|---|---|---|---|---|---|
| Диметродон ангеленсис | Олсон, 1962 год. | Техас | Действительный | 
| |
| Диметродон северный | Лейди, 1854 г. | Остров Принца Эдуарда | Действительный | Ранее известный как динозавр Bathygnathus Borealis. | 
|
| Диметродон боонеорум | Ромер, 1937 год. | Техас | Действительный | ||
| Диметродон кукольный | Кейс, 1907 год. | Техас | Действительный | Embolophorus dolovianus Cope, 1888 г. | |
| Диметродон гигагомогенес | Кейс, 1907 год. | Техас | Действительный | 
| |
| Диметродон Грандис | Ромер и Прайс, 1940 год. | Оклахома Техас |
Действительный | Клепсидропс гигас Коуп, 1878 г. Диметродон гигас Коуп, 1878 г. Дело Theropleura grandis , 1907 г. Батигглипт Теодора Кейса, 1911 г. Диметродон Максимус Ромер 1936 г. |

|
| Диметродон кемпае | Ромер, 1937 год. | Техас | Возможное имя сомнения | ||
| Диметродон лимбатус | Ромер и Прайс, 1940 год. | Оклахома Техас |
Действительный | Клепсидропс лимбатус Коуп, 1877 г. Диметродон инцизивус Коуп, 1878 г. Диметродон прямоугольный Коуп, 1878 г. Диметродон полурадикатус Коуп, 1881 г. |
|
| Диметродонские существа | Ромер, 1937 год. | Техас Оклахома |
Действительный | 
| |
| Диметродон макроспондил | Кейс, 1907 год. | Техас | Действительный | Клепсидропы макроспондилус Коуп, 1884 г. Кейс Dimetrodon platycentrus , 1907 г. |
|
| Диметродон миллери | Ромер, 1937 год. | Техас | Действительный | 
| |
| Диметродон наталис | Ромер, 1936 год. | Техас | Действительный | Клепсидропс наталис Коуп, 1878 г. | 
|
| Диметродон западный | Берман, 1977 г. | Аризона Нью-Мексико Юта |
Действительный | ||
| Диметродон тевтонис | Берман и др. , 2001 г. | Германия | Действительный |
Диметродон лимбатус
[ редактировать ]
Dimetrodonlimbatus был впервые описан Эдвардом Дринкером Коупом в 1877 году как Clepsydropslimbatus . [ 43 ] (Название Clepsydrops было впервые предложено Коупом в 1875 году для останков сфенакодонтид из округа Вермилион, штат Иллинойс , а позже использовалось для многих экземпляров сфенакодонтид из Техаса; многие новые виды сфенакодонтид из Техаса были отнесены либо к Clepsydrops , либо к Dimetrodon в конце XIX и начале двадцатого века.) Основанный на образце из Красных пластов Техаса , это был первый известный синапсид с парусной спинкой. В 1940 году палеонтологи Альфред Ромер и Айвор Прайс отнесли C.limbatus к роду Dimetrodon , сделав D.limbatus типовым видом Dimetrodon Ллевелин . [ 44 ]
Диметродон острый
[ редактировать ]Первое использование названия Dimetrodon произошло в 1878 году, когда Коуп назвал вид Dimetrodon incisivus наряду с Dimetrodon Rectiformis и Dimetrodon gigas . [ 42 ]
Диметродон прямоугольный
[ редактировать ]Dimetrodon rectiformis был назван наряду с Dimetrodon incisivus в статье Коупа 1878 года и был единственным из трех названных видов, сохранившим удлиненные нервные отростки. [ 42 ] В 1907 году палеонтолог Э. К. Кейс отнес D. rectiformis к виду D. incisivus . [ 41 ] Позже D. incisivus стал синонимом типового вида Dimetrodonlimbatus , что сделало D.rectiformis синонимом D.limbatus . [ 29 ]
Диметродон полурадикатус
[ редактировать ]Описанный в 1881 году на основе костей верхней челюсти, Dimetrodon semiradicatus был последним видом, названным Коупом. [ 51 ] В 1907 году ЕС Кейс синонимизировал D. semiradicatus с D. incisivus на основании сходства формы зубов и костей черепа. [ 41 ] D. incisivus и D. semiradicatus теперь считаются синонимами D.limbatus . [ 29 ]
Диметродон кукольный
[ редактировать ]Dimetrodon Dollovianus был впервые описан Эдвардом Дринкером Коупом в 1888 году как Embolophorus Dollovianus . В 1903 году Э. Кейс опубликовал пространное описание E. dolovianus , которого он позже отнес к Dimetrodon . [ 52 ]
Диметродон Грандис
[ редактировать ]
В 1907 году палеонтолог Э.К. Кейс назвал новый вид парусных синапсидов — Theropleura grandis . [ 41 ] В 1940 году Альфред Ромер и Ллевеллин Айвор Прайс переназначили Theropleura grandis на Dimetrodon , выведя вид D. grandis . [ 44 ]
Диметродон гигас
[ редактировать ]В своей статье 1878 года об окаменелостях из Техаса Коуп назвал Clepsydrops gigas наряду с первыми названными видами Dimetrodon , D.limbatus , D. incisivus и D.rectiformis . [ 42 ] В 1907 году Кейс реклассифицировал C. gigas как новый вид диметродона . [ 41 ] Кейс также описал очень хорошо сохранившийся череп Dimetrodon в 1904 году, отнеся его к виду Dimetrodon gigas . [ 53 ] В 1919 году Чарльз В. Гилмор приписал почти полный экземпляр диметродона D. gigas . [ 54 ] Dimetrodon gigas теперь признан синонимом D. grandis . [ 55 ]
Диметродон гигантомогенес
[ редактировать ]
Dimetrodon giganhomogenes был назван EC Case в 1907 году и до сих пор считается действительным видом Dimetrodon . [ 41 ] [ 29 ]
Диметродон макроспондил
[ редактировать ]Dimetrodon macrospondylus был впервые описан Коупом в 1884 году как Clepsydrops macrospondylus . В 1907 году Кейс реклассифицировал его как Dimetrodon macrospondylus . [ 41 ]
Диметродон платицентрус
[ редактировать ]Dimetrodon platycentrus был впервые описан Кейсом в его монографии 1907 года. Сейчас он считается синонимом Dimetrodon macrospondylus . [ 29 ]
Диметродон наталис
[ редактировать ]
Палеонтолог Альфред Ромер в 1936 году вывел вид Dimetrodon natalis , ранее описанный как Clepsydrops natalis . D. natalis был самым маленьким известным видом Dimetrodon в то время и был обнаружен рядом с остатками более крупного D.limbatus . [ 56 ]
Диметродон боонеорум
[ редактировать ]Dimetrodon booneorum был впервые описан Альфредом Ромером в 1937 году на основе останков из Техаса. [ 56 ]
Кампания «Диметродон»
[ редактировать ]Dimetrodon kempae был назван Ромером в 1937 году в той же статье, что и D. booneorum , D. loomisi и D. milleri . [ 56 ] Dimetrodon kempae был назван на основе одной плечевой кости и нескольких позвонков и, следовательно, может быть nomen dubium , который нельзя отличить как уникальный вид Dimetrodon . [ 12 ] В 1940 году Ромер и Прайс предположили, что D. kempae может не относиться к роду Dimetrodon , предпочитая классифицировать его как Sphenacodontidae incertae sedis . [ 44 ]
Диметродонские существа
[ редактировать ]
Dimetrodon loomisi был впервые описан Альфредом Ромером в 1937 году вместе с D. booneorum , D. kempae и D. milleri . [ 56 ] Останки были найдены в Техасе и Оклахоме.
Диметродон миллери
[ редактировать ]
Dimetrodon milleri был описан Ромером в 1937 году. [ 56 ] Это один из самых маленьких видов Dimetrodon в Северной Америке, который может быть тесно связан с D. occidentalis , другим видом с небольшим телом. [ 49 ] D. milleri известен по двум скелетам: одному почти полному (MCZ 1365), а другому менее полному, но более крупному (MCZ 1367). D. milleri — старейший известный вид диметродона .
Помимо небольшого размера, D. milleri отличается от других видов Dimetrodon тем, что его нервные отростки имеют круглую, а не восьмерочную форму в поперечном сечении. Его позвонки также короче по высоте по сравнению с остальной частью скелета, чем у других видов Dimetrodon . Череп высокий, морда короткая относительно височной области. Короткие позвонки и высокий череп также наблюдаются у видов D. booneorum , D.limbatus и D. grandis , что позволяет предположить, что D. milleri может быть первым представителем эволюционного прогресса между этими видами.
Диметродон ангеленсис
[ редактировать ]
Dimetrodon angelensis был назван палеонтологом Эвереттом К. Олсоном в 1962 году. [ 57 ] Сообщалось, что экземпляры этого вида были обнаружены в формации Сан-Анджело в Техасе. [ 58 ] Это также самый крупный вид диметродона.
Диметродон западный
[ редактировать ]Dimetrodon occidentalis был назван в 1977 году из Нью-Мексико. [ 48 ] Его название означает «западный диметродон », потому что это единственный североамериканский вид диметродона, известный к западу от Техаса и Оклахомы. Свое название он получил по единственному скелету, принадлежавшему относительно небольшой особи. Небольшой размер D. occidentalis аналогичен размеру D. milleri , что предполагает близкое родство. Экземпляры диметродона , найденные в Юте и Аризоне, вероятно, также принадлежат к D. occidentalis . [ 49 ]
Диметродон тевтонис
[ редактировать ]Dimetrodon teutonis был назван в 2001 году из Тюрингенского леса Германии и стал первым видом Dimetrodon, описанным за пределами Северной Америки. Это также самый маленький вид диметродона . [ 12 ]
Виды, отнесенные к разным родам
[ редактировать ]Диметродон крестообразный
[ редактировать ]В 1878 году Коуп опубликовал статью под названием «Тероморфные рептилии», в которой описал Dimetrodon Crusger . [ 59 ] D. Cruger отличался небольшими выступами, которые простирались с обеих сторон каждого нервного отростка, как ветви дерева. [ 60 ] В 1886 году Коуп отнес D. Crusger к роду Naosaurus, поскольку считал, что его шипы настолько отличаются от шипов других видов Dimetrodon , что этот вид заслуживает отдельного рода. [ 61 ] Позднее наозавр был синонимом Эдафозавра , рода, который Коуп назвал в 1882 году на основании черепов, которые, очевидно, принадлежали травоядным животным, учитывая их тупые дробящие зубы. [ 62 ]
Диметродон лонгирам
[ редактировать ]ЕС Кейс назвал этот вид Dimetrodon longiramus в 1907 году на основе лопатки и удлиненной нижней челюсти из формации Бель-Плейнс в Техасе. [ 41 ] В 1940 году Ромер и Прайс признали, что материал D. longiramus принадлежал к тому же таксону, что и другой экземпляр, описанный палеонтологом Сэмюэлем Венделлом Уиллистоном в 1916 году, который включал такую же удлиненную нижнюю челюсть и длинную верхнюю челюсть. [ 44 ] Уиллистон не считал свой экземпляр принадлежащим диметродону , а вместо этого классифицировал его как офиакодонтида . [ 63 ] Ромер и Прайс отнесли образцы Кейса и Уиллистона к недавно созданному роду и виду Secodontosaurus longiramus , который был тесно связан с Dimetrodon . [ 44 ] [ 64 ]
Филогенетическая классификация
[ редактировать ]Диметродон — ранний член группы, называемой синапсидами , в которую входят млекопитающие и многие их вымершие родственники, хотя он не является предком какого-либо млекопитающего (появившегося миллионы лет спустя). [ 65 ] ). В популярной культуре его часто принимают за динозавра, несмотря на то, что он вымер примерно за 40 миллионов лет (млн лет назад) до первого появления динозавров в триасовый период. Как синапсид, диметродон более тесно связан с млекопитающими, чем с динозаврами или любой другой живой рептилией. К началу 1900-х годов большинство палеонтологов называли Диметродона рептилией в соответствии с таксономией Линнея , которая относила Reptilia к классу , а Dimetrodon — к роду внутри этого класса. Млекопитающие были выделены в отдельный класс, а Диметродон описывался как «млекопитающая рептилия». Палеонтологи предположили, что млекопитающие произошли от этой группы в результате (так называемого) перехода от рептилий к млекопитающим.
Филогенетическая систематика Synapsida
[ редактировать ]
Согласно филогенетической систематике , потомки последнего общего предка диметродона . и всех ныне живущих рептилий должны включать всех млекопитающих, поскольку диметродон более тесно связан с млекопитающими, чем с любой ныне живущей рептилией Таким образом, если желательно избежать клада, состоящего как из млекопитающих, так и из ныне живущих рептилий, то диметродон не должен быть включен в эту кладу, как и любая другая «рептилия, похожая на млекопитающих». Потомки последнего общего предка млекопитающих и рептилий (появившегося около 310 млн лет назад в позднем карбоне ) поэтому разделены на две клады: Synapsida, которая включает Dimetrodon и млекопитающих, и Sauropsida , которая включает ныне живущих рептилий и всех вымерших рептилий, более тесно связанных между собой. для них, чем для млекопитающих. [ 4 ]
В пределах клады Synapsida Dimetrodon является частью клады Sphenacodontia , которая была впервые предложена в качестве ранней группы синапсид в 1940 году палеонтологами Альфредом Ромером и Ллевеллином Айвором Прайсом, наряду с группами Ophiacodontia и Edaphosauria . [ 44 ] Все три группы известны из позднего карбона и ранней перми. Ромер и Прайс различали их прежде всего по посткраниальным особенностям, таким как форма конечностей и позвонков. Ophiacodontia считалась самой примитивной группой, потому что ее члены выглядели наиболее рептильными, а Sphenacodontia была самой развитой, потому что ее члены больше всего напоминали группу под названием Therapsida , в которую входили ближайшие родственники млекопитающих. Ромер и Прайс поместили еще одну группу ранних синапсид, называемых варанопидами, в состав Sphenacodontia, считая их более примитивными, чем другие сфенакодонты, такие как Dimetrodon . [ 66 ] Они считали, что варанопиды и диметродоноподобные сфенакодонты были тесно связаны между собой, поскольку обе группы были плотоядными, хотя варанопиды намного меньше и больше похожи на ящериц, у них нет парусов.
Современный взгляд на взаимоотношения синапсид был предложен палеонтологом Робертом Р. Рейсом в 1986 году, чье исследование включало особенности, обнаруженные в основном в черепе, а не в посткраниальном скелете. [ 67 ] Диметродон по-прежнему считается сфенакодонтом в рамках этой филогении , но варанодонтиды теперь считаются скорее базальными синапсидами, выходящими за пределы клады Sphenacodontia. В состав Sphenacodontia входит группа Sphenacodontoidea , которая, в свою очередь, включает Sphenacodontidae и Therapsida . Sphenacodontidae — это группа, включающая Dimetrodon и несколько других синапсидов с парусной спинкой, таких как Sphenacodon и Secodontosaurus , в то время как Therapsida включает млекопитающих и их в основном пермских и триасовых родственников.
Ниже представлена кладограмма Clade Synapsida, которая соответствует этой филогении Synapsida , измененной на основе анализа Бенсона (2012). [ 66 ]
| Амниота |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
На приведенной ниже кладограмме показаны взаимоотношения нескольких видов Dimetrodon из Brink et al. , (2015). [ 68 ]
| Сфенакодонтиды |
| ||||||||||||||||||||||||||||||
Палеобиология
[ редактировать ]Функция нервных отростков
[ редактировать ]
Палеонтологи предложили множество способов использования паруса в жизни. Некоторые из первых, кто задумался о его предназначении, предположили, что парус мог служить камуфляжем среди тростника, пока диметродон ждал добычу, или как настоящий парус, похожий на лодку, чтобы ловить ветер, пока животное было в воде. [ 69 ] Другая причина заключается в том, что длинные нервные отростки могли бы стабилизировать туловище, ограничивая движения вверх и вниз, что позволило бы более эффективно перемещаться из стороны в сторону во время ходьбы. [ 27 ]
Терморегуляция
[ редактировать ]В 1940 году Альфред Ромер и Ллевеллин Айвор Прайс предположили, что парус выполняет функцию терморегуляции, позволяя людям согревать свое тело Солнцем. В последующие годы было создано множество моделей для оценки эффективности терморегуляции у диметродона . Например, в статье 1973 года в журнале Nature палеонтологи С. Д. Брэмвелл и П. Б. Феллгетт подсчитали, что человеку весом 200 кг (440 фунтов) потребовалось около полутора часов, чтобы температура его тела поднялась с 26 до 32 °C (79 до 90 °F). [ 70 ] В 1986 году Стивен К. Хаак пришел к выводу, что потепление происходит медленнее, чем считалось ранее, и что этот процесс, вероятно, занял четыре часа. Используя модель, основанную на множестве факторов окружающей среды и гипотетических физиологических аспектах диметродона , Хаак обнаружил, что парус позволяет диметродону быстрее согреваться утром и достигать немного более высокой температуры тела в течение дня, но он неэффективен в выделении избыточного тепла. и не позволял Диметродону сохранять более высокую температуру тела в ночное время. [ 71 ] В 1999 году группа инженеров-механиков создала компьютерную модель для анализа способности паруса регулировать температуру тела в разные времена года и пришла к выводу, что парус полезен для улавливания и выделения тепла в любое время года. [ 72 ]

Большинство этих исследований указывают на две терморегулирующие функции паруса диметродона : одну — как средство быстрого согревания по утрам, а другую — как способ охладиться, когда температура тела становится высокой. Предполагается, что диметродон и все другие наземные позвоночные ранней перми были хладнокровными или пойкилотермными и полагались на солнце для поддержания высокой температуры тела. Из-за своих больших размеров диметродон обладал высокой тепловой инерцией , а это означает, что изменения температуры тела у него происходили медленнее, чем у животных с меньшим телом. Поскольку по утрам температура повышалась, маленькая добыча диметродона могла согреть свое тело гораздо быстрее, чем существо размером с диметродон . Многие палеонтологи, в том числе Хаак, предположили, что парус Диметродона , возможно, позволял ему быстро согреться утром, чтобы не отставать от добычи. [ 71 ] Большая площадь поверхности паруса также означала, что тепло могло быстро рассеиваться в окружающей среде, что было полезно, если животному нужно было высвободить избыточное тепло, вырабатываемое в процессе метаболизма или поглощаемое солнцем. Диметродон, возможно, отклонил свой парус от солнца, чтобы охладиться, или ограничил приток крови к парусу, чтобы поддерживать тепло ночью. [ 69 ]
В 1986 году Дж. Скотт Тернер и К. Ричард Трейси предположили, что эволюция паруса у Dimetrodon была связана с эволюцией теплокровности у предков млекопитающих. Они думали, что парус Диметродона позволял ему быть гомеотермным , поддерживая постоянную, хотя и низкую температуру тела. Млекопитающие также гомойотермны, хотя они отличаются от диметродона тем, что они эндотермичны и контролируют температуру своего тела изнутри посредством усиленного метаболизма. Тернер и Трейси отметили, что ранние терапсиды, более развитая группа синапсидов, тесно связанных с млекопитающими, имели длинные конечности, которые могли выделять тепло таким же образом, как парус Диметродона . Гомеотермия, развившаяся у таких животных, как диметродон , возможно, перешла и на терапсидов посредством модификации формы тела, которая в конечном итоге развилась в теплокровность млекопитающих. [ 73 ]

Недавние исследования паруса диметродона и других сфенакодонтид подтверждают утверждение Хаака 1986 года о том, что парус плохо приспособлен к выделению тепла и поддержанию стабильной температуры тела. Наличие парусов у видов Dimetrodon с мелким телом, таких как D. milleri и D. teutonis, не соответствует идее о том, что целью паруса была терморегуляция, поскольку паруса меньшего размера менее способны передавать тепло и потому что маленькие тела могут легко поглощать и выделять тепло. самостоятельно. Более того, близкие родственники диметродона, такие как сфенакодон, имеют очень низкие гребни, которые были бы бесполезны в качестве устройств терморегуляции. [ 29 ] Считается, что большой парус Диметродона развился постепенно из этих меньших гребней, а это означает, что на протяжении большей части эволюционной истории паруса терморегуляция не могла выполнять важную функцию. [ 74 ]
Хотя функция его паруса остается неясной, диметродон и другие сфенакодонтиды, вероятно, были эндотермами всего тела, характеризующимися высоким энергетическим метаболизмом ( тахиметаболизмом ) и, вероятно, способностью поддерживать высокую и стабильную температуру тела. Этот вывод был частью исследования, охватывающего все амниоты , которое показало, что тахиметаболическая эндотермия широко распространена повсюду и, вероятно, плезиоморфна как синапсидам , так и зауропсидам . В случае диметродона доказательством был размер отверстий, через которые кровь доставлялась к трубчатым костям, а также высокое кровяное давление, которое было необходимо для обеспечения кровью верхушек хорошо васкуляризированных шипов, поддерживающих парус. [ 75 ]
У экземпляров Dimetrodon с более крупным телом паруса больше по сравнению с их размером, что является примером положительной аллометрии . Положительная аллометрия может способствовать терморегуляции, поскольку это означает, что по мере того, как люди становятся крупнее, площадь поверхности увеличивается быстрее, чем масса. Животные с более крупным телом выделяют большое количество тепла в результате метаболизма, и количество тепла, которое должно рассеиваться с поверхности тела, значительно больше, чем то, которое должно рассеиваться животными с меньшим телом. Эффективное рассеивание тепла можно предсказать у многих разных животных, используя единую зависимость между массой и площадью поверхности. Однако исследование аллометрии диметродона в 2010 году выявило другую взаимосвязь между его парусом и массой тела: фактический показатель масштабирования паруса был намного больше, чем показатель, ожидаемый у животного, приспособленного к рассеиванию тепла. Исследователи пришли к выводу, что парус диметродона рос гораздо быстрее, чем это было необходимо для терморегуляции, и предположили, что половой отбор было основной причиной его эволюции. [ 74 ]
Половой отбор
[ редактировать ]Аллометрический показатель высоты паруса аналогичен масштабированию межвидовой длины рогов к высоте плеч у оленьих . Более того, как заметил Баккер (1970) на примере Dimetrodon , у многих видов ящериц поднимается дорсальный гребень кожи во время демонстрации угрозы и ухаживания, а у современных ящериц присутствуют положительно аллометрические, сексуально диморфные оборки и подвесы (Echelle et al. 1978; Кристиан и др. 1995). Имеются также свидетельства полового диморфизма как по прочности скелета, так и по относительной высоте шипов D.limbatus (Ромер и Прайс, 1940). [ 74 ]
Половой диморфизм
[ редактировать ]Диметродон , возможно, имел половой диморфизм , а это означает, что самцы и самки имели немного разные размеры тела. некоторые экземпляры диметродона Предполагается, что относятся к самцам, поскольку у них более толстые кости, большие паруса, более длинные черепа и более выраженные верхнечелюстные «ступени», чем у других. Основываясь на этих различиях, установленные скелеты в Американском музее естественной истории (AMNH 4636) и Полевом музее естественной истории могут быть мужчинами, а скелеты в Денверском музее природы и науки ( MCZ 1347) и Музее Мичиганского университета естествознания могут быть женщинами. [ 44 ]


Палеоэкология
[ редактировать ]
Окаменелости диметродона известны из США (Техас, Оклахома, Нью-Мексико, Аризона, Юта и Огайо) и Германии, территорий, которые были частью суперконтинента Еврамерика в ранней перми. В Соединенных Штатах почти весь материал, приписываемый диметродону, поступил из трех геологических групп в северо-центральном Техасе и южно-центральной Оклахоме: группа Клир-Форк , группа Уичито и группа Пиз-Ривер . [ 76 ] [ 77 ] Большинство находок окаменелостей являются частью равнинных экосистем, которые в пермский период представляли собой обширные водно-болотные угодья. В частности, Красные пласты Техаса — это территория огромного разнообразия ископаемых четвероногих или четырехногих позвоночных. Помимо диметродона , наиболее распространенными четвероногими в Красных пластах и во всех раннепермских отложениях на юго-западе США являются земноводные Archeria , Diplocaulus , Eryops и Trimerorhachis , рептилиоморфа Seymouria , рептилия Captorhinus и синапсиды Ophiacodon и Edaphosaurus. . Эти четвероногие составляли группу животных, которую палеонтолог Эверетт К. Олсон назвал «пермско-каменноугольной хронофауной», фауной , которая доминировала в континентальной евроамериканской экосистеме в течение нескольких миллионов лет. [ 78 ] Судя по геологии таких отложений, как Красные пласты, считается, что фауна населяла хорошо заросшую равнинную дельтовую экосистему . [ 79 ]
Пищевая сеть
[ редактировать ]
Олсон сделал множество выводов о палеоэкологии красных отложений Техаса и роли диметродона в его экосистеме. Он предложил несколько основных типов экосистем, в которых жили древнейшие четвероногие. Диметродон принадлежал к самой примитивной экосистеме, развившейся из водных пищевых сетей. В нем водные растения были основными продуцентами и в основном питались рыбой и водными беспозвоночными. Большинство наземных позвоночных питались этими водными основными потребителями. Диметродон, вероятно, был главным хищником экосистемы Красных пластов, питаясь различными организмами, такими как акула Xenacanthus. [ нужна ссылка ] , водные амфибии Trimerorhachis и Diplocaulus , а также наземные четвероногие Seymouria и Trematops . Насекомые известны из красных пластов ранней перми и, вероятно, в некоторой степени были вовлечены в ту же пищевую сеть, что и диметродон , питаясь мелкими рептилиями, такими как Captorhinus . Комплекс Red Beds также включал некоторых из первых крупных наземных травоядных, таких как Edaphosaurus и Diadectes . Питаясь в основном наземными растениями, эти травоядные животные не получали энергию из водных пищевых сетей. По мнению Олсона, лучшим современным аналогом экосистемы, в которой обитает диметродон, является Эверглейдс . [ 79 ] Точный образ жизни диметродона (от земноводного до наземного) долгое время был спорным, но микроанатомия костей подтверждает наземный образ жизни. [ 80 ] это означает, что он питался в основном на суше, на берегу или на очень мелководье. Также существуют доказательства того, что диметродон охотился на спящего диплокаулуса во время засухи: три частично съеденных молодых диплокаулуса в норе из восьми со следами зубов диметродона , который их раскопал и убил. [ 81 ]
Единственный вид диметродона , обнаруженный за пределами юго-запада США, — это D. teutonis из Германии. Его останки были найдены в формации Тамбах на окаменелом участке, называемом местонахождением Бромакер. Бромакера Ассоциация раннепермских четвероногих необычна тем, что здесь мало синапсид с крупным телом, выполняющих роль высших хищников. По оценкам, D. teutonis имел длину всего 1,7 метра (5,6 фута), что слишком мало, чтобы охотиться на крупных диадектидных травоядных животных, которые в изобилии встречаются в сообществе Бромакера. Скорее всего, он питался мелкими позвоночными и насекомыми. Только три окаменелости можно отнести к крупным хищникам, и считается, что это были либо крупные варанопиды , либо мелкие сфенакодонты , оба из которых потенциально могли охотиться на D. teutonis . В отличие от равнинных дельтовых красных пластов Техаса, отложения Бромакера, как полагают, представляют собой горную среду без водных видов. Вполне возможно, что крупные хищники не входили в состав группы Бромакера, поскольку они зависели от крупных водных животных. земноводные для еды. [ 12 ]
См. также
[ редактировать ]- Секодонтозавр - вымерший род синапсид.
- Динозавры в Парке Юрского периода
- Эдафозавр - вымерший род синапсид.
Ссылки
[ редактировать ]- ^ «Диметродон» . Lexico Британский словарь английского языка . Издательство Оксфордского университета . Архивировано из оригинала 22 марта 2020 г.
- ^ «диметродон» . Dictionary.com Полный (онлайн). нд . Проверено 12 февраля 2018 г.
- ^ «Диметродон» . Палеобиологическая база данных . Архивировано из оригинала 2 октября 2013 года . Проверено 23 августа 2012 г.
- ^ Jump up to: а б с д Ангельчик, К.Д. (2009). «Диметродон — не динозавр: использование древовидного мышления для понимания древних родственников млекопитающих и их эволюции» . Эволюция: образование и информационно-пропагандистская деятельность . 2 (2): 257–271. дои : 10.1007/s12052-009-0117-4 .
- ^ Хаттенлокер, АК; Рега, Э. (2012). «Палеобиология и костная микроструктура синапсид пеликозаврианского класса». В Чинсами, А. (ред.). Предшественники млекопитающих: радиация, гистология, биология . Издательство Университета Индианы. стр. 90–119. ISBN 978-0-253-35697-0 .
- ^ «Знаменитые доисторические животные, которые на самом деле не были динозаврами» . 17 февраля 2021 г. Архивировано из оригинала 7 апреля 2021 г.
- ^ Блэк, Райли. «Диметродон в вашем генеалогическом древе» . Смитсоновский журнал . Проверено 5 декабря 2021 г.
- ^ Клаудсли-Томпсон, Дж. Л. (19 января 2005 г.). Экология и поведение мезозойских рептилий . Springer Science & Business Media. ISBN 978-3-540-22421-1 .
- ^ Фиеста, Энрике; Дэвидсон, Джон (10 января 2015 г.). Диметродон — пермский хищник . Книги Мендон Коттедж. ISBN 978-1-310-19617-1 .
- ^ Захос, Фрэнк; Ашер, Роберт (22 октября 2018 г.). Эволюция, разнообразие и систематика млекопитающих . Вальтер де Грюйтер ГмбХ & Ко КГ. ISBN 978-3-11-034155-3 .
- ^ Гоне, Иордания; Барден, Жереми; Жирондо, Марк; Хатчинсон, Джон Р.; Лорен, Мишель (2023). «Раскрытие постурального разнообразия млекопитающих: вклад поперечных сечений плечевой кости в палеобиологические выводы» . Журнал эволюции млекопитающих . 30 (2): 321–337. doi : 10.1007/s10914-023-09652-w . S2CID 256788973 .
- ^ Jump up to: а б с д и ж Берман, Д.С.; Рейс, РР; Мартенс, Т.; Хенричи, AC (2001). «Новый вид Dimetrodon (Synapsida: Sphenacodontidae) из нижней перми Германии регистрирует первое появление этого рода за пределами Северной Америки» (PDF) . Канадский журнал наук о Земле . 38 (5): 803–812. Бибкод : 2001CaJES..38..803B . doi : 10.1139/cjes-38-5-803 .
- ^ Jump up to: а б с д Фрёбиш, Дж.; Шох, Р.Р.; Мюллер, Дж.; Шиндлер, Т.; Швайс, Д. (2011). «Новый базальный сфенакодонтидный синапсид из позднего карбона бассейна Саар-Наэ, Германия» (PDF) . Acta Palaeontologica Polonica . 56 (1): 113–120. дои : 10.4202/app.2010.0039 . S2CID 45410472 .
- ^ Олсон, ЕС; Бирбауэр, младший (1953). «Формация Сан-Анджело, пермь Техаса, и ее позвоночные». Журнал геологии . 61 (5): 389–423. Бибкод : 1953JG.....61..389O . дои : 10.1086/626109 . S2CID 128681671 .
- ^ Олсон, ЕС (1955). «Параллелизм в эволюции пермских рептильных фаун Старого и Нового Света» . Филдиана . 37 (13): 385–401.
- ^ Штернберг, CW (1942). «Скелет неполовозрелого пеликозавра Dimetrodon cf. grandis из перми Техаса». Журнал палеонтологии . 16 (4): 485–486. JSTOR 1298848 .
- ^ Лорин, М.; Рейс, Р.Р. (2012). «Синапсиды: млекопитающие и их вымершие родственники» . Веб-проект «Древо жизни» . Архивировано из оригинала 7 декабря 2012 года . Проверено 24 августа 2012 г.
- ^ «Экспонат: Диметродон» . Американский музей естественной истории . Архивировано из оригинала 4 июля 2012 года . Проверено 2 июля 2012 года .
- ^ Jump up to: а б Баур, Г.; Кейс, ЕС (1899 г.). «История пеликозаврий с описанием рода Dimetrodon , Cope». Труды Американского философского общества . 20 (1): 5–62. дои : 10.2307/1005488 . hdl : 2027/uc1.32106020416696 . JSTOR 1005488 .
- ^ Jump up to: а б Аблер, В.Л. 2001. Модель зубцов тираннозавра, сделанная с помощью пропила и сверла. п. 84-89. В: Жизнь мезозойских позвоночных . Под ред. Танке, Д.Х., Карпентер, К., Скрепник, М.В., Издательство Университета Индианы.
- ^ «Великий энергетический вызов» . Среда .
- ^ Махо, Чай; Махо, Сиги; Скотт, Дайанн; Рейс, Роберт Р. (19 августа 2022 г.). «Пермские гиперплотоядные животные свидетельствуют о сложностях с зубами среди ранних амниот» . Природные коммуникации . 13 (1): 4882. Бибкод : 2022NatCo..13.4882M . дои : 10.1038/s41467-022-32621-5 . ПМЦ 9391490 . ПМИД 35986022 .
- ^ Кемп, Т.С. (2006). «Происхождение и раннее распространение терапсидных млекопитающих рептилий: палеобиологическая гипотеза» . Журнал эволюционной биологии . 19 (4): 1231–1247. дои : 10.1111/j.1420-9101.2005.01076.x . ПМИД 16780524 . S2CID 3184629 .
- ^ Лорин, М.; Рейс, Р.Р. (1997). «Аутапоморфии основных клад синапсидов» . Веб-проект «Древо жизни» . Проверено 24 августа 2012 г.
- ^ Ромер, А.С. (1927). «Заметки о пермско-каменноугольной рептилии диметродоне ». Журнал геологии . 35 (8): 673–689. Бибкод : 1927JG.....35..673R . дои : 10.1086/623462 . JSTOR 30060393 . S2CID 140679339 .
- ^ Jump up to: а б Рега, Э.А.; Норьега, К.; Сумида, СС; Хаттенлокер, А.; Ли, А.; Кеннеди, Б. (2012). «Зажившие переломы нервных отростков связанного с ним скелета диметродона: последствия для морфологии и функции спинного паруса». Филдиана Науки о жизни и Земле . 5 : 104–111. дои : 10.3158/2158-5520-5.1.104 . S2CID 108887164 .
- ^ Jump up to: а б с Рега, Э.; Сумида, С.; Норьега, К.; Пелл, К.; Ли, А. (2005). «Доказательная палеопатология I: онтогенетические и функциональные последствия спинных парусов диметродона ». Журнал палеонтологии позвоночных . 25 (С3): 103А. дои : 10.1080/02724634.2005.10009942 . S2CID 220413556 .
- ^ Сумида, С.; Рега, Э.; Норьега, К. (2005). «Доказательная палеопатология II: Влияние на филогенетический анализ рода Dimetrodon ». Журнал палеонтологии позвоночных . 25 (С3): 120А. дои : 10.1080/02724634.2005.10009942 . S2CID 220413556 .
- ^ Jump up to: а б с д и ж г Хаттенлокер, АК; Рега, Э.; Сумида, СС (2010). «Сравнительная анатомия и остеогистология гиперудлиненных нервных отростков сфенакодонтид Sphenacodon и Dimetrodon (Amniota: Synapsida)». Журнал морфологии . 271 (12): 1407–1421. дои : 10.1002/jmor.10876 . ПМИД 20886514 . S2CID 40899700 .
- ^ Рега, Э.А.; Норьега, К.; Сумида, С.; Ли, А. (2004). «Гистологический анализ травматического повреждения нескольких нервных отростков связанного с ним скелета диметродона : последствия для реакции заживления, морфологии спинного паруса и возраста смерти в синапсиде нижней перми». Интегративная и сравнительная биология . 44 : 628.
- ^ Константинович Чудинов Петр (1965). «Новые факты о фауне верхней перми СССР». Журнал геологии . 73 (1): 117–30. Бибкод : 1965JG.....73..117C . дои : 10.1086/627048 . S2CID 129428120 .
- ^ Бота-Бринк, Дж.; Модесто, СП (2007). «Агрегация пеликозавров разного возраста из Южной Африки: самые ранние свидетельства родительской заботы об амниотах?» . Труды Королевского общества Б. 274 (1627): 2829–2834. дои : 10.1098/rspb.2007.0803 . ПМК 2288685 . ПМИД 17848370 .
- ^ Недзведский, Г.; Бояновский, М. (2012). «Предполагаемый отпечаток тела эвпеликозавра из ранней перми Внутрисудетского бассейна, Польша». Ихнос . 19 (3): 150–155. Бибкод : 2012Ично..19..150Н . дои : 10.1080/10420940.2012.702549 . S2CID 129567176 .
- ^ Шпиндлер, Фредерик; Вернебург, Ральф; Шнайдер, Йорг В.; Лутхардт, Людвиг; Аннакер, Волкер; Рёсслер, Ронни (2018). «Первые древесные« пеликозавры »(Synapsida: Varanopidae) из ранней перми Хемница ископаемого Лагерштетте, Юго-Восточная Германия, с обзором филогении варанопидов». ПалЗ . 92 (2): 315–364. Бибкод : 2018PalZ...92..315S . дои : 10.1007/s12542-018-0405-9 . S2CID 133846070 .
- ^ Модесто, Шон П. (январь 2020 г.). «Боюсь об отношениях рептилий» . Экология и эволюция природы . 4 (1): 10–11. дои : 10.1038/s41559-019-1074-0 . ISSN 2397-334X . ПМИД 31900449 . S2CID 209672518 .
- ^ Форд, Дэвид П.; Бенсон, Роджер Би Джей (2019). «Переописание Orovenator mayorum (Sauropsida, Diapsida) с использованием микроКТ высокого разрешения и последствия для ранней филогении амниот» . Статьи по палеонтологии . 5 (2): 197–239. Бибкод : 2019PPal....5..197F . дои : 10.1002/spp2.1236 . ISSN 2056-2802 . S2CID 92485505 .
- ^ Спалдинг, ДЭЭ (1995). « Батигнат , первый «динозавр» Канады ». В Сарджанте, БЫЛО (ред.). Окаменелости позвоночных и эволюция научных концепций . Тейлор и Фрэнсис США. стр. 245–254. ISBN 2881249965 .
- ^ Спалдинг, ДЭЭ (1995). « Батигнат , первый «динозавр» Канады ». В Сарджанте, БЫЛО (ред.). Окаменелости позвоночных и эволюция научных концепций . Тейлор и Фрэнсис США. стр. 245–254. ISBN 2881249965 .
- ^ «Канукозавр! Первым канадским «динозавром» стал Dimetrodon Borealis» . физ.орг .
- ^ Мэтью, WD (1908). «Обзор «Ревизии пеликозавров Северной Америки» Кейса » . Наука . 27 (699): 816–818. Бибкод : 1908Sci....27..816M . дои : 10.1126/science.27.699.816 .
- ^ Jump up to: а б с д и ж г час я Кейс, ЕС (1907). Ревизия пеликозавров Северной Америки . Вашингтон, округ Колумбия: Вашингтонский Институт Карнеги. стр. 1 –176.
- ^ Jump up to: а б с д и Коуп, ЭД (1878). «Описания вымерших батрахий и рептилий из пермских формаций Техаса». Труды Американского философского общества . 17 (101): 505–530. JSTOR 982652 .
- ^ Jump up to: а б Коуп, ЭД (1877). «Описания вымерших позвоночных из пермских и триасовых формаций США». Труды Американского философского общества . 17 (100): 182–193. JSTOR 982295 .
- ^ Jump up to: а б с д и ж г час Ромер, А.С.; Прайс, Л.И. (1940). «Обзор пеликозавров». Специальный доклад Геологического общества Америки . Специальные статьи Геологического общества Америки. 28 : 1–538. дои : 10.1130/spe28-p1 .
- ^ Вон, ПП (1966). «Сравнение фаун раннепермских позвоночных региона Четырех углов и северо-центрального Техаса» (PDF) . Вклад в науку. 105 . Музей естественной истории округа Лос-Анджелес: 1–13. Архивировано из оригинала (PDF) 10 января 2017 г. Проверено 21 апреля 2016 г.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Вон, ПП (1969). «Раннепермские позвоночные из южного Нью-Мексико и их палеозогеографическое значение» (PDF) . Вклад в науку. 166 . Музей естественной истории округа Лос-Анджелес: 1–22. Архивировано из оригинала (PDF) 10 января 2017 г. Проверено 21 апреля 2016 г.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Олсон, ЕС (1975). «Пермско-каменноугольная палеоэкология и морфотипический ряд» . Американский зоолог . 15 (2): 371–389. дои : 10.1093/icb/15.2.371 .
- ^ Jump up to: а б с Берман, Д.С. (1977). «Новый вид Dimetrodon (Reptilia, Pelycosauria) из недельтовой фации нижней перми северо-центральной части Нью-Мексико». Журнал палеонтологии . 51 (1): 108–115. JSTOR 1303466 .
- ^ Jump up to: а б с Мадалена, К.; Сумида, С.; Зейглер, К.; Рега, Э. (2007). «Новая находка синапсида раннепермского уровня пеликозавров Dimetrodon (Eupelycosauria: Sphenacodontidae) из группы нижнего Катлера (ранняя пермь) Хемез-Пуэбло, северо-центральная часть Нью-Мексико». Журнал палеонтологии позвоночных . 27 (3, Доп.): 110А. дои : 10.1080/02724634.2007.10010458 .
- ^ Вон, ПП (1973). Позвоночные животные из группы Катлера в Долине Монументов и ее окрестностях (PDF) . Путеводитель геологического общества Нью-Мексико. Том. 24. Геологическое общество Нью-Мексико. стр. 99–105.
- ^ Коуп, ЭД (1881). «О некоторых новых батрахиях и рептилиях из пермских красных пластов Техаса». Бюллетень Геологической службы США . 6 : 79–82.
- ^ Кейс, EC (1903). «Остеология Embolophorus dolovianus , Cope, с попыткой реставрации» . Журнал геологии . 11 (1): 1–28. Бибкод : 1903JG.....11....1C . дои : 10.1086/621055 . JSTOR 30056735 .
- ^ Кейс, ЕС (1904 г.). «Остеология черепа рода пеликозавров диметродона » . Журнал геологии . 12 (4): 304–311. Бибкод : 1904JG.....12..304C . дои : 10.1086/621157 . JSTOR 30055825 .
- ^ Гилмор, CW (1919). «Смонтированный скелет Dimetrodon gigas в Национальном музее США с примечаниями по анатомии скелета» (PDF) . Труды Национального музея США . 56 (2300): 525–539. дои : 10.5479/si.00963801.56-2300.525 .
- ^ Хенричи, AC; Берман, Д.С.; Лукас, СГ; Хекерт, AB; Райнхарт, ЛФ; Зейглер, К.Э. (2005). «Запястье и предплюсна раннепермского синапсида Sphenacodonferox (Eupelycosauria: Sphenacodontidae)» (PDF) . В Лукасе, СГ; Зейглер, К.Е. (ред.). Неморская пермь . Бюллетень Музея естественной истории и науки Нью-Мексико. Том. 30. Альбукерке: Музей естественной истории и науки Нью-Мексико. стр. 106–110.
- ^ Jump up to: а б с д и Ромер, А.С. (1937). «Новые роды и виды рептилий-пеликозавров» (PDF) . Труды Зоологического клуба Новой Англии . 16 : 89–97.
- ^ Олсон, ЕС (1962). «Позднепермские наземные позвоночные, США и СССР». Труды Американского философского общества . 52 (2): 1–224. дои : 10.2307/1005904 . JSTOR 1005904 .
- ^ Баттейл, Б. (2000). «Сравнение фауны амниот поздней перми Гондваны и Лавразии». Журнал африканских наук о Земле . 31 (1): 165–174. Бибкод : 2000JAfES..31..165B . дои : 10.1016/S0899-5362(00)00081-6 .
- ^ Коуп, ЭД (1878). «Тероморфные рептилии» . Американский натуралист . 12 (12): 829–830. дои : 10.1086/272251 .
- ^ Коуп, ЭД (1880). «Второй вклад в историю позвоночных пермской формации Техаса». Труды Американского философского общества . 19 (107): 38–58. JSTOR 982605 .
- ^ Коуп, ЭД (1886). «Длинноиглая тероморфа пермской эпохи» . Американский натуралист . 20 (6): 544–545. дои : 10.1086/274275 .
- ^ Коуп, ЭД (1882). «Третий вклад в историю позвоночных пермской формации Техаса». Труды Американского философского общества . 20 (112): 447–461. JSTOR 982692 .
- ^ Уиллистон, Юго-Запад (1916). «Остеология некоторых американских пермских позвоночных, II». Вклад Музея Уокера . 1 : 165–192.
- ^ Рейс, РР; Берман, Д.С.; Скотт, Д. (1992). «Черепная анатомия и взаимоотношения секодонтозавра, необычной млекопитающей рептилии (Synapsida: Sphenacodontidae) из ранней перми Техаса». Зоологический журнал Линнеевского общества . 104 (2): 127–184. дои : 10.1111/j.1096-3642.1992.tb00920.x . S2CID 56425294 .
- ^ Появляются млекопитающие
- ^ Jump up to: а б Бенсон, Р.Дж. (2012). «Взаимоотношения базальных синапсид: краниальные и посткраниальные морфологические перегородки предполагают разные топологии». Журнал систематической палеонтологии . 10 (2): 601–624. Бибкод : 2012JSPal..10..601B . дои : 10.1080/14772019.2011.631042 . S2CID 84706899 .
- ^ Рейс, Р.Р. (1986). «Пеликозаврия». В Сьюсе, Х.-Д. (пшеница.). Справочник по палеогерпетологии . Том. 17А. Густав Фишер Верлаг. стр. 1–102. ISBN 978-3-89937-032-4 . ОЛ 12985656М .
- ^ Бринк, Кирстин С.; Мэддин, Хиллари С.; Эванс, Дэвид С.; Рейс, Роберт Р.; Сьюс, Ханс-Дитер (2015). «Переоценка исторической канадской окаменелости Bathygnathus Borealis из ранней перми острова Принца Эдуарда» . Канадский журнал наук о Земле . 52 (12): 1109–1120. Бибкод : 2015CaJES..52.1109B . doi : 10.1139/cjes-2015-0100 .
- ^ Jump up to: а б Флоридес, Джорджия; Калогиру, ЮАР; Тассу, ЮАР; Врубель, Л. (2001). «Природная среда и термическое поведение Dimetrodonlimbatus ». Журнал термической биологии . 26 (1): 15–20. Бибкод : 2001JTBio..26...15F . дои : 10.1016/S0306-4565(00)00019-X . ПМИД 11070340 .
- ^ Брамвелл, CD; Феллгетт, П.Б. (1973). «Терморегуляция у парусных ящериц». Природа . 242 (5394): 203–205. Бибкод : 1973Natur.242..203B . дои : 10.1038/242203a0 . S2CID 4159825 .
- ^ Jump up to: а б Хаак, Южная Каролина (1986). «Тепловая модель парусного пеликозавра». Палеобиология . 12 (4): 450–458. Бибкод : 1986Pbio...12..450H . дои : 10.1017/S009483730000316X . S2CID 124339088 .
- ^ Флоридес, Джорджия; Врубель, ЛК; Калогиру, ЮАР; Тассу, ЮАР (1999). «Тепловая модель рептилий и пеликозавров». Журнал термической биологии . 24 (1): 1–13. Бибкод : 1999JTBio..24....1F . дои : 10.1016/S0306-4565(98)00032-1 .
- ^ Тернер, Дж.С.; Трейси, ЧР (1986). «Размер тела, гомеотермия и контроль теплообмена у млекопитающих рептилий» (PDF) . В Хоттоне, Северная Каролина III; Маклин, доктор медицинских наук; Рот, Джей-Джей; Рот, ЕС (ред.). Экология и биология млекопитающих рептилий . Вашингтон, округ Колумбия: Издательство Смитсоновского института. стр. 185–194. Архивировано из оригинала (PDF) 12 апреля 2016 г. Проверено 26 июля 2012 г.
- ^ Jump up to: а б с Томкинс, Дж.Л.; ЛеБас, Северная Каролина; Уиттон, член парламента; Мартилл, DM; Хамфрис, С. (2010). «Положительная аллометрия и предыстория полового отбора» (PDF) . Американский натуралист . 176 (2): 141–148. дои : 10.1086/653001 . ПМИД 20565262 . S2CID 36207 . Архивировано из оригинала (PDF) 12 апреля 2016 г.
- ^ Григг, Гордон; Новак, Джулия; Бикудо, Хосе Эдуардо Перейра Вилкен; Бал, Нареш Чандра; Вудворд, Холли Н.; Сеймур, Роджер С. (2022). «Энотермия всего тела: древняя, гомологичная и широко распространенная среди предков млекопитающих, птиц и крокодилов» . Биологические обзоры . 97 (2): 766–801. дои : 10.1111/brv.12822 . hdl : 2440/134060 . ISSN 1464-7931 . ПМК 9300183 . ПМИД 34894040 . S2CID 245021195 .
- ^ Лукас, СГ; Шпильманн, Дж.А.; Райнхарт, ЛФ; Мартенс, Т. (2009). Диметродон (Amniota: Synapsida: Sphenacodontidae) из формации Або нижней перми, округ Сокорро, Нью-Мексико (PDF) . Путеводитель геологического общества Нью-Мексико. Том. 60. Геологическое общество Нью-Мексико. стр. 281–284.
- ^ Нельсон, Джон В., Роберт В. Хук и Дэн С. Чейни (2013). Литостратиграфия формации Clear Fork нижней перми (леонарда) Северо-Центрального Техаса в период перехода от карбона к перми: Бюллетень 60, под ред. Спенсер Г. Лукас и др. Музей естественной истории и науки Нью-Мексико, стр. 286-311. Проверено 28 декабря 2017 г.
- ^ Салливан, К.; Рейс, РР; Мэй, WJ (2000). «Крупные диссорофоидные элементы скелета из трещин отрога Ричардса в нижней перми, Оклахома, и их палеоэкологические последствия». Журнал палеонтологии позвоночных . 20 (3): 456–461. doi : 10.1671/0272-4634(2000)020[0456:LDSEFT]2.0.CO;2 . JSTOR 4524117 . S2CID 140709673 .
- ^ Jump up to: а б Олсон, ЕС (1966). «Эволюция сообщества и происхождение млекопитающих». Экология . 47 (2): 291–302. Бибкод : 1966Ecol...47..291O . дои : 10.2307/1933776 . JSTOR 1933776 .
- ^ Крылов А.; Жермен, Д.; Кановиль, А.; Винсент, П.; Саш, М.; Лаурин, М. (2008). «Эволюция костной микроанатомии большеберцовой кости четвероногих и ее использование в палеобиологических выводах» . Журнал эволюционной биологии . 21 (3): 807–826. дои : 10.1111/j.1420-9101.2008.01512.x . ПМИД 18312321 . S2CID 6102313 .
- ^ «Плавниковое чудовище отрубало головы древним земноводным» . Живая наука . 31 октября 2013 г.
Внешние ссылки
[ редактировать ] Определения из Викисловаря
Определения из Викисловаря  СМИ из Commons
СМИ из Commons  Таксоны из Wikispecies
Таксоны из Wikispecies  Данные из Викиданных
Данные из Викиданных
- Страница Dimetrodon Palaeos на Dimetrodon
- Введение на веб-страницу палеонтологического музея Калифорнийского университета пеликозавров , посвященную ранним синапсидам, включая диметродон.
- «Диметродон: наш самый маловероятный предок» . PBS Эоны . 21 августа 2017 г. — через YouTube .






- Пермская Германия
- Сфенакодонтиды
- Цизуральские синапсиды Европы
- Цизуральские синапсиды Северной Америки
- Роды доисторических синапсид
- Переходные окаменелости
- Таксоны, названные Эдвардом Дринкером Коупом
- Ископаемые таксоны, описанные в 1878 г.
- Первые появления цизуральского рода
- Вымирание приуральских родов
- Высшие хищники