Ангиогенез
| Ангиогенез | |
|---|---|
 Ангиогенез после васкулогенез | |
| Идентификаторы | |
| Сетка | D000096482 |
| Анатомическая терминология | |

Ангиогенез -это физиологический процесс, посредством которого новые кровеносные сосуды образуются из ранее существовавших сосудов, [ 1 ] [ 2 ] [ 3 ] сформировано на более ранней стадии васкулогенез . Ангиогенез продолжает рост сосудистой сети в основном путем процессов прорастания и расщепления, но такие процессы, как ангиогенез коалерорования , [ 4 ] Удлинение судов и сосуды также играют роль. [ 2 ] Васулогенез является эмбриональным образованием эндотелиальных клеток из мезодермных клеток, предшественников [ 5 ] и от неоваскуляризации , хотя дискуссии не всегда являются точными (особенно в старых текстах). Первые сосуды в развивающемся эмбрионе формируются посредством васкулогенез, после чего ангиогенез отвечают за большинство, если не для всех, рост кровеносных сосудов во время развития и при заболевании. [ 6 ] [ 7 ] [ 8 ]
Ангиогенез является нормальным и жизненно важным процессом в росте и развитии, а также в заживлении ран и в образовании грануляционной ткани . Тем не менее, это также является фундаментальным шагом в переходе опухолей из доброкачественного состояния к злокачественному , что приводит к использованию ингибиторов ангиогенеза при лечении рака . [ 9 ] Основная роль ангиогенеза в росте опухоли была впервые предложена в 1971 году Иудеи Фолькман , который описал опухоли как «горячие и кровавые». [ 10 ] Иллюстрируя, что, по крайней мере, для многих типов опухолей, перфузия промывки и даже гиперемия являются характерными.
Типы
[ редактировать ]Прорастание ангиогенеза
[ редактировать ]Прорастающий ангиогенез был первой идентифицированной формой ангиогенеза, и из -за этого он гораздо более понятен, чем интуисцептивный ангиогенез. Это происходит на нескольких хорошо охарактеризованных этапах. Первоначальный сигнал поступает из областей ткани, лишенных сосудистой сети. Гипоксия , которая отмечена в этих областях , заставляет ткани требовать присутствия питательных веществ и кислорода, которые позволят ткани выполнять метаболическую активность. Из-за этого паренхимальные клетки будут выделять сосудистый эндотелиальный фактор роста ( VEGF-A ), который является проангиогенным фактором роста. [ 11 ] Эти биологические сигналы активируют рецепторы на эндотелиальных клетках, присутствующих в ранее существовавших кровеносных сосудах. Во -вторых, активированные эндотелиальные клетки, также известные как клетки наконечника , [ 12 ] Начните высвобождать ферменты , называемые протеазами , которые разлагают базарную мембрану , чтобы позволить эндотелиальным клеткам сбежать от исходных (родительских) стен сосудов. эндотелиальные клетки Затем пролиферируются в окружающую матрицу и образуют твердые ростки, соединяющие соседние сосуды. Клетки, которые пролиферируют, расположены за клетками кончика и известны как стеб -клетки . [ 12 ] Пролиферация этих клеток позволяет капиллярной ростке расти в длину одновременно.
Поскольку ростки распространяются на источник ангиогенного стимула, эндотелиальные клетки мигрируют в тандеме , используя молекулы адгезии, называемые интегринами . Эти ростки затем образуют петли, чтобы стать полноценным просветом сосуда , когда клетки мигрируют в место ангиогенеза. Прорастание происходит со скоростью несколько миллиметров в день и позволяет новым сосудам расти между пробелами в сосудистой сети . Он заметно отличается от разделения ангиогенеза, потому что он формирует совершенно новые сосуды, а не расщепление существующих сосудов.
Интуисцептивный ангиогенез
[ редактировать ]Интуисцептивный ангиогенез , также известный как разделение ангиогенеза , представляет собой образование нового кровеносного сосуда путем разделения существующего кровеносного сосуда на два.
Инвабеция впервые наблюдалась у неонатальных крыс. В этом типе формирования сосудов капиллярная стена простирается в просвет, чтобы разделить один сосуд на два. Существует четыре фазы интуисцептивного ангиогенеза. Во -первых, две противоположные капиллярные стены устанавливают зону контакта. Во -вторых, разъединения эндотелиальных клеток реорганизуются, а бислой сосуда перфорирован, чтобы позволить факторам роста и клетки проникать в просвет. В -третьих, между двумя новыми сосудами в зоне контакта образуется ядро, которая заполнена перицитами и миофибробластами . Эти клетки начинают класть коллагеновые волокна в ядро, чтобы обеспечить внеклеточный матрикс для роста просвета сосуда. Наконец, ядро не было изменена в основной структуре. Инвабеция важна, потому что это реорганизация существующих клеток. Это позволяет значительно увеличить количество капилляров без соответствующего увеличения количества эндотелиальных клеток . Это особенно важно для эмбрионального развития, так как нет достаточно ресурсов, чтобы создать богатых Микропаскулятура с новыми клетками каждый раз, когда развивается новый сосуд. [ 13 ]
Коалесцентный ангиогенез
[ редактировать ]Ангиогенез коалерования - это способ ангиогенеза, который считается противоположностью интуисцептивному ангиогенезу, где капилляры сливаются или объединяются, чтобы сделать более крупный кровенсен, тем самым увеличивая кровоток и кровообращение. [ 14 ] Коалесцентный ангиогенез вышел из области эмбриологии. Предполагается, что он играет роль в формировании неоваскулятуры, например, в опухоли. [ 15 ]
Физиология
[ редактировать ]Механическая стимуляция
[ редактировать ]Механическая стимуляция ангиогенеза не хорошо охарактеризована. Существует значительное количество споров в отношении стресса сдвига , действующего на капилляры, вызвав ангиогенез, хотя современные знания предполагают, что увеличение сокращений мышц может увеличить ангиогенез. [ 16 ] Это может быть связано с увеличением производства оксида азота во время упражнений. Оксид азота приводит к вазодилатации кровеносных сосудов.
Химическая стимуляция
[ редактировать ]Химическая стимуляция ангиогенеза выполняется различными ангиогенными белками, например, интегринами и простаглантами, включая несколько факторов роста, например, VEGF, FGF.
Обзор
[ редактировать ]| Стимулятор | Механизм |
|---|---|
| FGF | Способствует пролиферации и дифференцировке эндотелиальных клеток, клеток гладких мышц и фибробластов |
| Овощ | Влияет на проницаемость |
| VEGFR и NRP-1 | Интегрировать сигналы выживания |
| Ang1 и Ang2 | Стабилизировать сосуды |
| PDGF (BB-Homomomer) и PDGFR | вернуть клетки гладких мышц |
| Рецепторы TGF-β , эндоглин и TGF-β | ↑ внеклеточного матрикса Производство |
| CCL2 | Рекрутирует лимфоциты на участки воспаления |
| Гистамин | |
| Integrins A V B 3 , A V B 5 (? [ 17 ] ) 5 B и 1 | Связывать матрицу макромолекулы и протеиназы |
| VE-кадгерин и CD31 | Эндотелиальные молекулы соединения |
| Эфрин | Определить формирование артерий или вен |
| активаторы плазминогена | Преодолевает внеклеточный матрикс , выпускает и активирует факторы роста |
| Ингибитор активатора плазминогена-1 | Стабилизирует близлежащие суда |
| ENOS и COX-2 | |
| AC133 | регулирует ангиобластов дифференциацию |
| Id1 / id3 | Регулирует эндотелиальную трансдифференцировку |
| Класс 3 семафорины | Модулирует адгезию эндотелиальных клеток, миграцию, пролиферацию и апоптоз. Изменяет сосудистую проницаемость [ 18 ] |
| Nogo-A | Регулирует миграцию и пролиферацию эндотелиальных клеток. [ 19 ] Изменяет сосудистую проницаемость. [ 20 ] |
FGF
[ редактировать ]Семейство фактора роста фибробластов (FGF) с его прототипом членов FGF-1 (кислота FGF) и FGF-2 (базовый FGF) состоит на сегодняшний день не менее 22 известных членов. [ 21 ] Большинство из них представляют собой одноцепочечные пептиды 16-18 кДа и демонстрируют высокую аффинность к гепарину и гепаран сульфат. В целом, FGF стимулируют различные клеточные функции путем связывания с FGF-рецепторами клеточной поверхности в присутствии гепариновых протеогликанов. Семейство FGF-рецепторов состоит из семи членов, и все рецепторные белки представляют собой одноцепочечные рецепторные тирозинкиназы, которые активируются посредством аутофосфорилирования, индуцированного механизмом FGF-опосредованной димеризации рецепторов. Активация рецептора приводит к каскаду трансдукции сигнала, которая приводит к активации генов и разнообразным биологическим реакциям, включая дифференцировку клеток, пролиферацию и растворение матрицы, что инициируя процесс митогенной активности, критический для роста эндотелиальных клеток, фибробластов и гладких мышечных клеток Полем FGF-1, уникальный среди всех 22 членов семейства FGF, может связываться со всеми семью подтипами FGF-рецептора, что делает его самым широким членом семейства FGF, и мощный митоген для разнообразных типов клеток, необходимых для установки ангиогенного Ответ в поврежденных (гипоксических) тканях, где происходит активация FGF-рецепторов. [ 22 ] FGF-1 стимулирует пролиферацию и дифференцировку всех типов клеток, необходимых для создания артериального сосуда, включая эндотелиальные клетки и клетки гладких мышц; Этот факт отличает FGF-1 от других проангиогенных факторов роста , таких как фактор роста эндотелиального эндотелия сосудов (VEGF), который в первую очередь способствует формированию новых капилляров. [ 23 ] [ 24 ]
Помимо FGF-1, одной из наиболее важных функций фактора роста фибробластов-2 (FGF-2 или BFGF ) является продвижение пролиферации эндотелиальных клеток и физическая организация эндотелиальных клеток в трубчатые структуры, что способствует ангиогенезу. FGF-2 является более сильным ангиогенным фактором, чем VEGF или PDGF ( фактор роста, полученный из тромбоцитов ); Тем не менее, это менее мощно, чем FGF-1. Помимо стимулирования роста кровеносных сосудов, AFGF (FGF-1) и BFGF (FGF-2) являются важными игроками в заживлении ран. Они стимулируют пролиферацию фибробластов и эндотелиальных клеток, которые вызывают ангиогенез и развивающуюся грануляционную ткань; Оба увеличивают кровоснабжение и заполняют рану/полость в начале процесса заживления ран.
Овощ
[ редактировать ]Было продемонстрировано, что фактор роста эндотелиального эндотелия (VEGF) является основным участником ангиогенеза, увеличивая количество капилляров в данной сети. Первоначальные исследования in vitro продемонстрировали, что бычьи капиллярные эндотелиальные клетки будут пролиферировать и показывать признаки структур труб при стимуляции VEGF и BFGF , хотя результаты были более выражены с VEGF. [ 25 ] Повышенная регуляция VEGF является основным компонентом физиологического ответа на физические упражнения, и его роль в ангиогенезе является возможным лечением в сосудистых травмах. [ 26 ] [ 27 ] [ 28 ] [ 29 ] Исследования in vitro ясно демонстрируют, что VEGF является мощным стимулятором ангиогенеза, потому что в присутствии этого фактора роста выселенные эндотелиальные клетки будут пролиферируются и мигрируют, в конечном итоге образуя структуры труб, напоминающие капилляры. [ 16 ] VEGF вызывает массивный сигнальный каскад в эндотелиальных клетках. Binding to VEGF receptor-2 (VEGFR-2) starts a tyrosine kinase signaling cascade that stimulates the production of factors that variously stimulate vessel permeability (eNOS, producing NO), proliferation/survival (bFGF), migration (ICAMs/VCAMs/MMPs) и, наконец, дифференциация в зрелые кровеносные сосуды. Механически, VEGF активируется с сокращениями мышц в результате увеличения кровотока в пораженные области. Повышенный поток также вызывает значительное увеличение продукции мРНК рецепторов VEGF 1 и 2. Увеличение выработки рецептора означает, что сокращения мышц могут вызвать повышение регуляции сигнального каскада, относящегося к ангиогенезу. В рамках ангиогенного сигнального каскада, широко считается, что нет основного фактора ангиогенного ответа, поскольку ингибирование NO существенно снижает эффекты ангиогенных факторов роста. Однако ингибирование NO во время упражнений не ингибирует ангиогенез, указывая на то, что в ангиогенном ответе есть и другие факторы. [ 16 ]
Ангиопоэтины
[ редактировать ]Ангиопоэтины исследованиями , Ang1 и Ang2, необходимы для образования зрелых кровеносных сосудов, как продемонстрировано мыши . [ 30 ] ANG1 и ANG2 являются факторами роста белка, которые действуют путем связывания их рецепторов, TIE-1 и TIE-2 ; Хотя это несколько противоречиво, кажется, что клеточные сигналы передаются в основном TIE-2 ; Хотя некоторые документы показывают физиологическую сигнализацию через TIE-1 и . Этими рецепторами являются тирозинкиназы . Таким образом, они могут инициировать передачу сигналов клеток , когда связывание лиганда вызывает димеризацию, которая инициирует фосфорилирование на ключевых тирозинах.
Ммп
[ редактировать ]Другим важным участником ангиогенеза является матричная металлопротеиназа (MMP). MMP помогают разгрозить белки, которые сохраняют твердые стенки сосуда. Этот протеолиз позволяет эндотелиальным клеткам сбежать в интерстициальную матрицу, как видно при прорастании ангиогенеза. Ингибирование MMP предотвращает образование новых капилляров . [ 31 ] Эти ферменты высоко регулируются во время процесса образования сосудов, потому что разрушение внеклеточного матрикса уменьшит целостность микропаскулятуры. [ 16 ]
DLL4
[ редактировать ]Дельта-подобный лиганд 4 (DLL4) является белком с отрицательным регуляторным эффектом на ангиогенез. [ 32 ] [ 33 ] DLL4 является трансмембранным лигандом, для семейства рецепторов Notch . Было проведено много исследований, которые послужили для определения последствий дельта-подобного лиганда 4. Одно исследование, в частности, оценивало влияние DLL4 на сосудистость и рост опухоли. [ 34 ] Чтобы опухоль расти и развиваться, она должна иметь правильную сосудистую сеть. Путь VEGF жизненно важен для развития сосудистой сети, которая, в свою очередь, помогает расти опухоли. Комбинированная блокада VEGF и DLL4 приводит к ингибированию прогрессирования опухоли и ангиогенеза на протяжении всей опухоли. Это связано с препятствием передачи сигналов в передаче сигналов эндотелиальных клеток, которая отключает пролиферацию и прорастание этих эндотелиальных клеток. С этим торможением клетки неконтролируемо растут, поэтому рак останавливается на этом этапе. Если блокада, однако, должна была быть поднята, клетки снова начнут свою пролиферацию. [ 35 ]
Класс 3 семафорины
[ редактировать ]Семфорины класса 3 (SEMA3S) регулируют ангиогенез путем модуляции адгезии эндотелиальных клеток , миграции, пролиферации, выживания и рекрутирования перицитов . [ 18 ] Кроме того, семафорины могут мешать VEGF-опосредованному ангиогенезу, поскольку как SEMA3, так и VEGF-A конкурируют за связывание рецептора нейропилина в эндотелиальных клетках. [ 36 ] [ 37 ] Следовательно, относительные уровни экспрессии SEMA3 и VEGF-A могут быть важны для ангиогенеза. [ 18 ]
Химическое запрещение
[ редактировать ]Ингибитор ангиогенеза может быть эндогенным или выходить извне в качестве лекарственного средства или диетического компонента .
Применение в медицине
[ редактировать ]Ангиогенез как терапевтическая цель
[ редактировать ]Ангиогенез может быть мишенью для борьбы с такими заболеваниями, как болезни сердца, характеризующиеся либо плохой васкуляризацией, либо аномальной сосудистой сети. [ 38 ] Применение конкретных соединений, которые могут ингибировать или вызывать создание новых кровеносных сосудов в организме, может помочь бороться с такими заболеваниями. Наличие кровеносных сосудов, где не должно быть ни одного, может повлиять на механические свойства ткани, увеличивая вероятность неудачи. Отсутствие кровеносных сосудов в восстановительной или иной метаболически активной ткани может ингибировать восстановление или другие важные функции. Несколько заболеваний, такие как ишемические хронические раны , являются результатом неудачи или недостаточного образования кровеносных сосудов и могут лечить местным расширением кровеносных сосудов, тем самым привлекая новые питательные вещества на участок, способствуя восстановлению. Другие заболевания, такие как возрастная дегенерация желтого пятна , могут быть созданы путем локальной экспансии кровеносных сосудов, мешающих нормальным физиологическим процессам.
Современное клиническое применение принципа ангиогенеза можно разделить на две основные области: антиангиогенные методы лечения, с которой началось ангиогенные исследования и проангиогенные терапии. Принимая во внимание, что антиангиогенная терапия используется для борьбы с раком и злокачественными новообразованиями, [ 39 ] [ 40 ] которые требуют пролиферации кислорода и питательных веществ, про-ангиогенная терапия изучается как варианты лечения сердечно-сосудистых заболеваний , что является причиной смерти в западном мире . Одним из первых применений проангиогенных методов у людей было немецкое исследование с использованием фактора роста фибробластов 1 (FGF-1) для лечения заболевания коронарной артерии. [ 23 ] [ 41 ] [ 42 ] [ 43 ]
Что касается механизма действия , проангиогенные методы могут быть дифференцированы на три основные категории: генная терапия , нацеливание генов, представляющих интерес для усиления или ингибирования; Заместительная терапия белка , которая в первую очередь манипулирует ангиогенным фактором роста, такими как FGF-1 или фактор роста эндотелия сосудов , VEGF; и клеточная терапия, которая включает в себя имплантацию специфических типов клеток.
Есть все еще серьезные, нерешенные проблемы, связанные с генной терапией. Трудности включают эффективную интеграцию терапевтических генов в геном клеток -мишеней, снижая риск нежелательного иммунного ответа, потенциальной токсичности, иммуногенности , воспалительных реакций и онкогенеза , связанных с вирусными векторами, используемыми при имплантационных генах и чистой сложности генетики Основа ангиогенеза. Наиболее часто встречающиеся расстройства у людей, такие как болезни сердца, высокое кровяное давление, диабет и болезнь Альцгеймера , скорее всего, вызваны комбинированными эффектами вариаций во многих генах, и, таким образом, инъекция одного гена может не быть значительно полезными в такие заболевания. [ Цитация необходима ]
Напротив, проангиогенная белковая терапия использует четко определенные, точно структурированные белки, с ранее определенными оптимальными дозами отдельного белка для болезненных состояний и с хорошо известными биологическими эффектами. [ 1 ] С другой стороны, препятствие белковой терапии - это способ доставки. Устные, внутривенные, внутриартериальные или внутримышечные маршруты введения белка не всегда являются такими эффективными, как терапевтический белок может метаболизировать или очистить, прежде чем он сможет попасть в ткань-мишени. Проангиогенная терапия на основе клеток все еще остается на ранних стадиях исследований, и многие открытые вопросы, касающиеся лучших типов клеток и дозировки для использования.
Ангиогенез опухоли
[ редактировать ]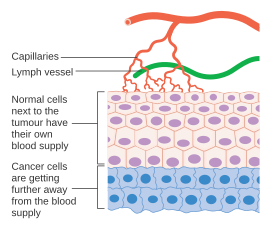
Раковые клетки - это клетки, которые потеряли способность делиться контролируемым образом. Злокачественная опухоль состоит из популяции быстро делящихся и растущих раковых клеток, которые постепенно начисляют мутации . Тем не менее, опухоли нуждаются в выделенном кровоснабжении, чтобы обеспечить кислород и другие необходимые питательные вещества, которые им требуются, чтобы расти до определенного размера (обычно 1–2 мм 3 ). [ 44 ] [ 45 ]
Опухоли вызывают рост кровеносных сосудов (ангиогенез), секретируя различные факторы роста (например, VEGF ) и белки. Факторы роста, такие как BFGF и VEGF, могут вызвать рост капилляров в опухоль, что некоторые исследователи подозревают, что требуется питательные вещества, что позволяет расширить опухоль. В отличие от нормальных кровеносных сосудов, опухолевые кровеносные сосуды расширяются нерегулярной формой. [ 46 ] Другие клиницисты считают, что ангиогенез действительно служит путем отходов, убирая биологические конечные продукты, секретируемые быстро делящимися раковыми клетками. В любом случае ангиогенез является необходимым и необходимым шагом для перехода от небольшого безвредного кластера клеток, которые часто говорят, что размером с металлического шарика в конце шариковой ручки к большой опухоли. Ангиогенез также требуется для распространения опухоли или метастазирования . [ 9 ] Одиночные раковые клетки могут оторваться от установленной твердой опухоли, попасть в кровеносные сосуды и переносить в отдаленное место, где они могут имплантировать и начать рост вторичной опухоли. В настоящее время данные свидетельствуют о том, что кровеносные сосуды в данной солидной опухоле, на самом деле, могут быть мозаичными сосудами, состоящими из эндотелиальных клеток и опухолевых клеток. [ 9 ] Эта мозаичность позволяет значительно выбросить опухолевые клетки в сосудистую сеть, что, возможно, способствует появлению циркулирующих опухолевых клеток в периферической крови пациентов со злокачественными новообразованиями. [ 47 ] Последующий рост таких метастазов также потребует поставки питательных веществ и кислорода и пути утилизации отходов.
Эндотелиальные клетки давно считаются генетически более стабильными, чем раковые клетки. Эта геномная стабильность дает преимущество для нацеливания эндотелиальных клеток с использованием антиангиогенной терапии по сравнению с химиотерапией , направленной на раковые клетки, которые быстро мутируют и получают лекарственную устойчивость к лечению. По этой причине считается, что эндотелиальные клетки являются идеальной мишенью для терапии, направленной против них. [ 48 ]
Образование опухолевых кровеносных сосудов
[ редактировать ]Механизм образования кровеносных сосудов с помощью ангиогенеза инициируется спонтанным разделением опухолевых клеток из -за мутации. Ангиогенные стимуляторы затем высвобождаются опухолевыми клетками. Затем они отправляются в уже установленные, близлежащие кровеносные сосуды и активируют свои рецепторы эндотелиальных клеток. Это вызывает высвобождение протеолитических ферментов из сосудистой сети. Эти ферменты нацелены на определенную точку на кровеносном сосуде и вызывают образование пор. Это тот момент, откуда будет расти новый кровеносный сосуд. Причина, по которой опухолевые клетки нуждаются в кровоснабжении, заключается в том, что они не могут расти более чем на 2-3 миллиметра в диаметре без установленного кровоснабжения, что эквивалентно около 50-100 клеток. [ 49 ] Некоторые исследования показали, что сосуды, образованные внутри опухолевой ткани, имеют более высокую нерегулярность и больше по размеру, что также связано с более низким прогнозом. [ 50 ] [ 51 ]
Ангиогенез при сердечно -сосудистых заболеваниях
[ редактировать ]Ангиогенез представляет собой превосходную терапевтическую мишень для лечения сердечно -сосудистых заболеваний. Это мощный, физиологический процесс, который лежит в основе естественного способа, которым наши тела реагируют на уменьшение кровоснабжения на жизненно важные органы, а именно неоангиогенез : производство новых коллатеральных сосудов для преодоления ишемического оскорбления. [ 23 ] Большое количество доклинических исследований было проведено с помощью белковой, генной и клеточной терапии на животных моделях ишемии сердца, а также моделями заболевания периферической артерии. Воспроизводимые и заслуживающие доверия успехи в этих ранних исследованиях на животных привели к большому энтузиазму, что этот новый терапевтический подход может быть быстро переведен в клиническую пользу для миллионов пациентов в западном мире с этими расстройствами. Десятилетие клинического тестирования как генной, так и белковой терапии, предназначенных для стимуляции ангиогенеза в недостаточных тканях и органах, привело к одному разочарованию к другому. Хотя все эти доклинические показания, которые дали большие перспективы для перехода ангиогенеза от животных к людям, были в том или ином виде, включенные в клинические испытания на ранней стадии, FDA на сегодняшний день настаивала на том, что первичные Конечная точка для одобрения ангиогенного агента должна быть улучшением физических упражнений у лечащихся пациентов. [ 52 ]
Эти сбои предполагали, что либо это неправильные молекулярные мишени, которые вызывают неоваскуляризацию, что их можно эффективно использовать только в том случае, если они сформулированы и введены правильно, или что их представление в контексте общего клеточного микросреда может играть жизненно важную роль в их полезности. Может потребоваться представить эти белки таким образом, чтобы имитировать естественные сигнальные события, включая концентрацию , пространственные и временные профили, а также их одновременное или последовательное представление с другими соответствующими факторами. [ 53 ]
Упражнение
[ редактировать ]Ангиогенез обычно связан с аэробными упражнениями и упражнениями на выносливость . В то время как артериогенез производит изменения сети, которые позволяют значительно увеличить количество общего потока в сети, ангиогенез вызывает изменения, которые позволяют обеспечить большую доставку питательных веществ в течение длительного периода времени. Капилляры предназначены для обеспечения максимальной эффективности доставки питательных веществ, поэтому увеличение количества капилляров позволяет сети доставлять больше питательных веществ за то же количество времени. Большее количество капилляров также обеспечивает больший обмен кислородом в сети. Это жизненно важно для обучения на выносливость, потому что это позволяет человеку продолжать тренироваться в течение длительного периода времени. Тем не менее, никакие экспериментальные данные не свидетельствуют о том, что в упражнениях на выносливость требуется увеличение капиллярности, чтобы увеличить максимальную доставку кислорода. [ 16 ]
Макулярная дегенерация
[ редактировать ]Сверхэкспрессия VEGF вызывает повышенную проницаемость в кровеносных сосудах в дополнение к стимулирующему ангиогенезу. При влажной макулярной дегенерации VEGF вызывает пролиферацию капилляров в сетчатку. Поскольку увеличение ангиогенеза также вызывает отек , кровь и другие жидкости сетчатки протекают в сетчатку , вызывая потерю зрения. Антиангиогенные препараты, нацеленные на пути VEGF, теперь успешно используются для лечения этого типа макулярной дегенерации
Тканевые конструкции
[ редактировать ]Ангиогенез сосудов из тела хозяина в имплантированные ткани, инженерированные конструкции. Успешная интеграция часто зависит от тщательной васкуляризации конструкции, поскольку она обеспечивает кислород и питательные вещества и предотвращает некроз в центральных областях имплантата. [ 54 ] Было показано, что PDGF стабилизирует васкуляризацию в коллагенгликозаминогликановых каркасах . [ 55 ]
История
[ редактировать ]Первый отчет об ангиогенезе можно проследить до книги «Трактат о крови, воспалении и ранних ранх», опубликованных в 1794 году, где шотландского анатомиста Джона Хантера были составлены результаты исследований . В своем исследовании Хантер наблюдал процесс роста новых кровеносных сосудов у кроликов. Тем не менее, он не пропустил термин «ангиогенез», который в настоящее время широко используется учеными. Охотник также ошибочно приписал процесс роста новых кровеносных сосудов с эффектом врожденного жизненно важного принципа в крови. Считается, что термин «ангиогенез» появился только до 1900 -х годов. Начало современного исследования ангиогенеза отмечено докладом Иудеи Фолькман о ключевой роли ангиогенеза в росте опухоли. [ 10 ] [ 56 ] [ 57 ]
Количественная оценка
[ редактировать ]Количественная оценка параметров сосудистой сети, таких как микрососудистая плотность, имеет различные осложнения из -за преимущественного окрашивания или ограниченного представления тканей гистологическими срезами. Недавние исследования показали полную трехмерную реконструкцию сосудистой структуры опухоли и количественную оценку структур сосудов во всех опухолях на животных моделях. [ 58 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный Santulli G, ed. (2013). Ангиогенез понимания систематического обзора Нью -Йорк: Nova Science. ISBN 978-1-62618-114-4 .
- ^ Jump up to: а беременный Dudley AC, Griffioen AW (август 2023 г.). «Патологический ангиогенез: механизмы и терапевтические стратегии» . Ангиогенез . 26 (3): 313–347. doi : 10.1007/s10456-023-09876-7 . PMC 10105163 . PMID 37060495 .
- ^ Birbrair A, Zhang T, Wang ZM, Messi ML, Olson JD, Mintz A, Delbono O (июль 2014 г.). «Перициты типа 2 участвуют в нормальном и опухолевом ангиогенезе» . Американский журнал физиологии. Клеточная физиология . 307 (1): C25 - C38. doi : 10.1152/ajpcell.00084.2014 . PMC 4080181 . PMID 24788248 .
- ^ Nitzsche B, Rong WW, Goede A, Hoffmann B, Scarpa F, Kuebler WM, et al. (Февраль 2022 г.). «Коалценнительное ангиогенез-очевидность для новой концепции созревания сосудистой сети» . Ангиогенез . 25 (1): 35–45. doi : 10.1007/s10456-021-09824-3 . PMC 8669669 . PMID 34905124 .
- ^ Risau W, Flamme I (1995). «Васулогенез». Ежегодный обзор биологии клеток и развития . 11 : 73–91. doi : 10.1146/annurev.cb.11.110195.000445 . PMID 8689573 .
- ^ Flamme I, Frölich T, Risau W (ноябрь 1997 г.). «Молекулярные механизмы васкулогенез и эмбрионального ангиогенеза». Журнал клеточной физиологии . 173 (2): 206–210. doi : 10.1002/(SICI) 1097-4652 (199711) 173: 2 <206 :: AID-JCP22> 3.0.co; 2-c . PMID 9365523 . S2CID 36723610 .
- ^ Arrabi N, Torabi M, Fassihi A, Ghasemi F (2024). «Идентификация ингибиторов рецепторов рецепторов эндотелиального эндотелиального эндотелия сосудов посредством моделирования обучения на основе деревьев и моделирования молекулярной стыковки». Хемометрика . 1 (1): 1. DOI : 10.1002/cem.3545 .
- ^ Тораби М., Ясами-Хабабани С., Сардари С., Гасеми Ф. (2024). «Выявление новых потенциальных кандидатов для ингибирования EGF через алгоритм машинного обучения». Европейский журнал фармакологии . 963 (15): 9759–9815. doi : 10.1016/j.ejphar.2023.176176 . PMID 38000720 .
- ^ Jump up to: а беременный в Milosevic V, Edelmann RJ, Fosse JH, Ostman A, Akslen LA (2022). «Молекулярные фенотипы эндотелиальных клеток в злокачественных опухолях». В Akslen La, Watnick Rs (Eds.). Биомаркеры микроокружения опухоли . Cham: Springer International Publishing. С. 31–52. doi : 10.1007/978-3-030-98950-7_3 . ISBN 978-3-030-98950-7 .
- ^ Jump up to: а беременный Penn JS (11 марта 2008 г.). Сетчатка и хориоидальный ангиогенез . Спрингер. С. 119–. ISBN 978-1-4020-6779-2 Полем Получено 26 июня 2010 года .
- ^ Adair TH, Montani JP. Ангиогенез. Сан -Рафаэль (Калифорния): науки о жизни Morgan & Claypool; 2010. Глава 1, Обзор ангиогенеза. Доступно по адресу: https://www.ncbi.nlm.nih.gov/books/nbk53238/
- ^ Jump up to: а беременный Уиверс H, Skaer H (июль 2014 г.). «Клетки кончиков: основные регуляторы тубулогенез?» Полем Семинары в биологии клеток и развития . 31 (100): 91–99. doi : 10.1016/j.semcdb.2014.04.009 . PMC 4071413 . PMID 24721475 .
- ^ Burri PH, Hlushchuk R, Joonov V (ноябрь 2004 г.). «Интуисцептивный ангиогенез: его появление, его характеристики и его значение» . Динамика развития . 231 (3): 474–488. doi : 10.1002/dvdy.20184 . PMID 15376313 . S2CID 35018922 .
- ^ Nitzsche B, Rong WW, Goede A, Hoffmann B, Scarpa F, Kuebler WM, et al. (Февраль 2022 г.). «Коалценнительное ангиогенез-очевидность для новой концепции созревания сосудистой сети» . Ангиогенез . 25 (1): 35–45. doi : 10.1007/s10456-021-09824-3 . PMC 8669669 . PMID 34905124 .
- ^ Pezzella F, Kerbel RS (февраль 2022 г.). «Об ангиогенезе коалса и замечательной гибкости кровеносных сосудов» . Ангиогенез . 25 (1): 1–3. doi : 10.1007/s10456-021-09825-2 . PMID 34993716 . S2CID 254188870 .
- ^ Jump up to: а беременный в дюймовый и Приор BM, Yang HT, Terjung RL (сентябрь 2004 г.). «Что заставляет сосуды расти с тренировками?». Журнал прикладной физиологии . 97 (3): 1119–1128. doi : 10.1152/japplphysiol.00035.2004 . PMID 15333630 .
- ^ Возможно, ингибитор ангиогенеза: Sheppard D (октябрь 2002 г.). «Эндотелиальные интегрины и ангиогенез: больше не так просто» . Журнал клинических исследований . 110 (7): 913–914. doi : 10.1172/jci16713 . PMC 151161 . PMID 12370267 .
- ^ Jump up to: а беременный в Mecollari V, Nieuwenhuis B, Verhaagen J (2014). «Перспектива роли передачи сигналов семафорина класса III в травме центральной нервной системы» . Границы в клеточной нейробиологии . 8 : 328. doi : 10.3389/fncel.2014.00328 . PMC 4209881 . PMID 25386118 .
- ^ Rust R, Grönnert L, Gantner C, Enzler A, Mulders G, Weber RZ, et al. (Июль 2019). «Целевая терапия Nogo-A способствует восстановлению сосудов и функциональному восстановлению после инсульта» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (28): 14270–14279. Bibcode : 2019pnas..11614270R . doi : 10.1073/pnas.1905309116 . PMC 6628809 . PMID 31235580 .
- ^ Rust R, Weber RZ, Grönnert L, Mulders G, Maurer MA, Hofer AS, et al. (Декабрь 2019). «Антитела против Nogo-A предотвращают утечку сосудов и действуют как проангиогенные факторы после инсульта» . Научные отчеты . 9 (1): 20040. BIBCODE : 2019NATSR ... 920040R . doi : 10.1038/s41598-019-56634-1 . PMC 6934709 . PMID 31882970 .
- ^ Орниц Д.М., Ито Н. (2001). «Факторы роста фибробластов» . Биология генома . 2 (3): обзоры3005. doi : 10.1186/gb-2001-2-3-reviews3005 . PMC 138918 . PMID 11276432 .
- ^ Blaber M, Disalvo J, Thomas Ka (февраль 1996 г.). «Рентгеновская кристаллическая структура человеческого фактора роста фибробластов человека». Биохимия . 35 (7): 2086–2094. Citeseerx 10.1.1.660.7607 . doi : 10.1021/bi9521755 . PMID 8652550 .
- ^ Jump up to: а беременный в Stegmann TJ (декабрь 1998 г.). «FGF-1: фактор роста человека в индукции неоангиогенеза». Экспертное мнение по расследованию лекарств . 7 (12): 2011–2015. doi : 10.1517/13543784.7.12.2011 . PMID 15991943 .
- ^ Khurana R, Simons M (апрель 2003 г.). «Понимание испытаний ангиогенеза с использованием фактора роста фибробластов для прогрессирующего артериосклеротического заболевания». Тенденции в сердечно -сосудистой медицине . 13 (3): 116–122. doi : 10.1016/S1050-1738 (02) 00259-1 . PMID 12691676 .
- ^ Goto F, Goto K, Weindel K, Folkman J (ноябрь 1993 г.). «Синергетические эффекты фактора роста эндотелиального эндотелия и основного фактора роста фибробластов на пролиферацию и образование пуповины эндотелиальных клеток бычьего капилляра в коллагеновых гелях». Лабораторное расследование; Журнал технических методов и патологии . 69 (5): 508–517. PMID 8246443 .
- ^ Ding YH, Luan XD, Li J, Rafols JA, Guthinkonda M, Diaz FG, Ding Y (декабрь 2004 г.). «Сверхэкспрессия ангиогенных факторов, вызванная физическими упражнениями, и снижение ишемии/реперфузионного повреждения при инсульте» . Современные нервно -сосудистые исследования . 1 (5): 411–420. doi : 10.2174/1567202043361875 . PMID 16181089 . S2CID 22015361 . Архивировано из оригинала 19 апреля 2012 года.
{{cite journal}}: Cs1 maint: непредвзятый URL ( ссылка ) - ^ Гэвин Т.П., Робинсон К.Б., Йегер Р.К., Англия Дж.А., Нифонг Л.В., Хикнер Р.К. (январь 2004 г.). «Реакция ангиогенного фактора роста на острые системные упражнения в скелетных мышцах человека». Журнал прикладной физиологии . 96 (1): 19–24. doi : 10.1152/japplphysiol.00748.2003 . PMID 12949011 . S2CID 12750224 .
- ^ Kraus RM, Stallings HW, Yeager RC, Gavin TP (апрель 2004 г.). «Циркулирующая реакция в плазме VEGF на физические упражнения у сидячих и выносливых людей». Журнал прикладной физиологии . 96 (4): 1445–1450. doi : 10.1152/japplphysiol.01031.2003 . PMID 14660505 . S2CID 21090407 .
- ^ Ллойд П.Г., Приор Б.М., Ян Х.Т., Тержунг Р.Л. (май 2003). «Экспрессия ангиогенного фактора роста в скелетных мышцах крысы в ответ на тренировку». Американский журнал физиологии. Сердечная и циркуляторная физиология . 284 (5): H1668 - H1678. doi : 10.1152/ajpheart.00743.2002 . PMID 12543634 .
- ^ Терстон Г (октябрь 2003 г.). «Роль ангиопоэтинов и рецепторных тирозинкиназ в ангиогенезе и лимфангиогенезе». Клеточные и тканевые исследования . 314 (1): 61–68. doi : 10.1007/s00441-003-0749-6 . PMID 12915980 . S2CID 2529783 .
- ^ Haas TL, Milkiewicz M, Davis SJ, Zhou AL, Egginton S, Brown MD, et al. (Октябрь 2000). «Активность металлопротеиназы матрицы требуется для ангиогенеза, вызванного активностью, в скелетных мышцах крысы». Американский журнал физиологии. Сердечная и циркуляторная физиология . 279 (4): H1540 - H1547. doi : 10.1152/ajpheart.2000.279.4.h1540 . PMID 11009439 . S2CID 2543076 .
- ^ Лобов И.Б., Ренард Р.А., Пападопулос Н., Гейл Н.В., Терстон Г., Янкопулос Г.Д., Виганд С.Дж. (февраль 2007 г.). «Delta-подобный лиганд 4 (DLL4) индуцируется VEGF как негативный регулятор ангиогенного прорастания» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (9): 3219–3224. Bibcode : 2007pnas..104.3219L . doi : 10.1073/pnas.0611206104 . PMC 1805530 . PMID 17296940 .
- ^ Hellström M, Phng LK, Hofmann JJ, Wallgard E, Coultas L, Lindblom P, et al. (Февраль 2007 г.). «Передача сигналов DLL4 через Notch1 регулирует образование клеток наконечника во время ангиогенеза». Природа . 445 (7129): 776–780. Bibcode : 2007natur.445..776h . doi : 10.1038/nature05571 . PMID 17259973 . S2CID 4407198 .
- ^ Segarra M, Williams CK, Sierra Mde L, Bernardo M, McCormick PJ, Maric D, et al. (Сентябрь 2008 г.). «Активация передачи сигналов DLL4 снижает опухолевую сосудистость и ингибирует рост опухоли» . Кровь . 112 (5): 1904–11. doi : 10.1182/blood-2007-11-126045 . PMC 2518892 . PMID 18577711 .
- ^ Lee D, Kim D, Choi YB, Kang K, Sung ES, Ahn JH, et al. (Июль 2016 г.). «Одновременная блокада VEGF и DLL4 с помощью HD105, биспецифического антитела, ингибирует прогрессирование опухоли и ангиогенез» . Мабс . 8 (5): 892–904. doi : 10.1080/19420862.2016.1171432 . PMC 4968104 . PMID 27049350 .
- ^ Soker S, Takashima S, Miao HQ, Neufeld G, Klagsbrun M (март 1998 г.). «Нейропилин-1 экспрессируется эндотелиальными и опухолевыми клетками в качестве изоформ-специфического рецептора для фактора роста эндотелия сосудов» . Клетка . 92 (6): 735–745. doi : 10.1016/s0092-8674 (00) 81402-6 . PMID 9529250 . S2CID 547080 .
- ^ Herzog B, Pellet-Many C, Britton G, Hartzoulakis B, Zachary IC (август 2011 г.). «Связывание VEGF с NRP1 необходимо для стимуляции VEGF миграции эндотелиальных клеток, образования комплекса между NRP1 и VEGFR2 и передачи сигналов через фосфорилирование FAK TYR407» . Молекулярная биология клетки . 22 (15): 2766–2776. doi : 10.1091/mbc.e09-12-1061 . PMC 3145551 . PMID 21653826 .
- ^ Ferrara N, Kerbel RS (декабрь 2005 г.). «Ангиогенез как терапевтическая цель». Природа . 438 (7070): 967–974. Bibcode : 2005natur.438..967f . doi : 10.1038/nature04483 . PMID 16355214 . S2CID 1183610 .
- ^ Folkman J, Klagsbrun M (январь 1987 г.). «Ангиогенные факторы». Наука . 235 (4787): 442–447. Bibcode : 1987sci ... 235..442f . doi : 10.1126/science.2432664 . PMID 2432664 .
- ^ Folkman J (сентябрь 1996 г.). «Борьба с раком, нападая на его кровоснабжение». Scientific American . 275 (3): 150–154. Bibcode : 1996sciam.275c.150f . doi : 10.1038/Scientificamerican0996-150 . PMID 8701285 .
- ^ Стегманн Т.Дж., Хопперт Т., Шнайдер А., Гемейнхардт С., Кечер М., Ибинг Р., Струпп Г. (сентябрь 2000 г.). «[Индукция индукции инурогенеза миокарда с помощью факторов роста человека. Новый терапевтический подход при коронарной болезни сердца]». Герц (на немецком языке). 25 (6): 589–599. doi : 10.1007/pl00001972 . PMID 11076317 . S2CID 21240045 .
- ^ Folkman J (февраль 1998 г.). «Ангиогенная терапия сердца человека» . Циркуляция . 97 (7): 628–629. doi : 10.1161/01.cir.97.7.628 . PMID 9495294 .
- ^ Zarei P, Ghasemi F (2024). «Применение искусственного интеллекта и репозиции лекарственного средства для выявления ингибиторов рецепторов рецептора фактора роста фибробластов: обзор» . Усовершенствованные биомедицинские исследования . 13 (15): 9759–9815. doi : 10.4103/abr.abr_170_23 . PMC 10958741 . PMID 38525398 .
- ^ McDougall SR, Anderson AR, капеллан MA (август 2006 г.). «Математическое моделирование динамического адаптивного опухолевого ангиогенеза: клинические последствия и терапевтические стратегии нацеливания». Журнал теоретической биологии . 241 (3): 564–589. Bibcode : 2006jthbi.241..564m . doi : 10.1016/j.jtbi.2005.12.022 . PMID 16487543 .
- ^ Пролив Ф., Герреро П., Аларкон Т., Мейни П.К., Бирн Х.М. (февраль 2015 г.). «Мезоскопическое и континуум моделирование ангиогенеза» . Журнал математической биологии . 70 (3): 485–532. Arxiv : 1401.5701 . doi : 10.1007/s00285-014-0771-1 . PMC 5320864 . PMID 24615007 .
- ^ Гонсалес-Перес Р.Р., Руэда Б.Р. (2013). Регуляторы ангиогенеза опухоли (первое изд.). Бока Ратон: Тейлор и Фрэнсис. п. 347. ISBN 978-1-4665-8097-8 Полем Получено 2 октября 2014 года .
- ^ Аллард В.Дж., Матера Дж., Миллер М.К., Репуллет М., Коннелли М.К., Рао С. и др. (Октябрь 2004). «Опухолевые клетки циркулируют в периферической крови всех основных карциномов, но не у здоровых субъектов или пациентов с незвуковыми заболеваниями» . Клиническое исследование рака . 10 (20): 6897–6904. doi : 10.1158/1078-0432.ccr-04-0378 . PMID 15501967 .
- ^ Bagri A, Kouros-Mehr H, Leong KG, Plowman GD (март 2010 г.). «Использование анти-VEGF-адъювантной терапии при раке: проблемы и обоснование». Тенденции в молекулярной медицине . 16 (3): 122–132. doi : 10.1016/j.molmed.2010.01.004 . PMID 20189876 .
- ^ Nishida N, Yano H, Nishida T, Kamura T, Kojiro M (сентябрь 2006 г.). «Ангиогенез при раке» . Сосудистое здоровье и управление рисками . 2 (3): 213–219. doi : 10.2147/vhrm.2006.2.3.213 . PMC 1993983 . PMID 17326328 .
- ^ Milosevic V, Edelmann RJ, Winge I, Strell C, Mezheyeuuski A, Knutsvik G, et al. (Июль 2023 г.). «Размер сосуда как маркер выживаемости при положительном раке молочной железы рецептора эстрогена» . Исследования и лечения рака молочной железы . 200 (2): 293–304. doi : 10.1007/s10549-023-06974-4 . PMC 10241708 . PMID 37222874 .
- ^ Mikalsen LT, Dhakal HP, Bruland Øs, Naume B, Borgen E, Nesland JM, Olsen DR (2013-10-11). Аоки I (ред.). «Клиническое воздействие среднего размера сосуда и пользы у пациентов с раком молочной железы» . Plos один . 8 (10): E75954. BIBCODE : 2013PLOSO ... 875954M . doi : 10.1371/journal.pone.0075954 . PMC 3795733 . PMID 24146798 .
- ^ Hariawala MD, Sellke FW (июнь 1997 г.). «Ангиогенез и сердце: терапевтические последствия» . Журнал Королевского общества медицины . 90 (6): 307–311. doi : 10.1177/014107689709000604 . PMC 1296305 . PMID 9227376 .
- ^ Cao L, Mooney DJ (ноябрь 2007 г.). «Пространственно -временный контроль над передачей сигналов фактора роста для терапевтической неоваскуляризации» . Расширенные обзоры доставки наркотиков . 59 (13): 1340–1350. doi : 10.1016/j.addr.2007.08.012 . PMC 2581871 . PMID 17868951 .
- ^ Rouwkema J, Khademhosseini A (сентябрь 2016 г.). «Васуляризация и ангиогенез в тканевой инженерии: помимо создания статических сетей» . Тенденции в биотехнологии . 34 (9): 733–745. doi : 10.1016/j.tibtech.2016.03.002 . PMID 27032730 .
- ^ Do Amaral RJ, Cavanagh B, O'Brien FJ, Kearney CJ (февраль 2019). «Фактор роста, полученный из тромбоцитов, стабилизирует васкуляризацию в коллагеновых гликозаминогликановых каркасах in vitro» . Журнал тканевой инженерии и регенеративной медицины . 13 (2): 261–273. doi : 10.1002/срок.2789 . PMID 30554484 . S2CID 58767660 .
- ^ Lenzi P, Bocci G, Natale G (апрель 2016 г.). «Джон Хантер и происхождение термина« ангиогенез » ». Ангиогенез . 19 (2). Springer Science and Business Media LLC: 255–256. doi : 10.1007/s10456-016-9496-7 . HDL : 11568/795270 . PMID 26842740 . S2CID 254189385 .
- ^ Adair TH, Montani JP (2010). «История» . Ангиогенез . Morgan & Claypool Life Sciences. doi : 10.4199/c00017ed1v01y201009isp009 (неактивный 2024-09-12). PMID 21452444 . Получено 2023-07-20 .
{{cite book}}: CS1 Maint: doi неактивен по состоянию на сентябрь 2024 года ( ссылка ) - ^ Chien CC, Kempson IM, Wang CL, Chen HH, Hwu Y, Chen NY, et al. (Май -июнь 2013). «Полное микромасштабное профилирование опухолевого микроангиогенеза: микрорадиологическая методология выявляет фундаментальные аспекты ангиогенеза опухоли и дает множество количественных параметров для ее характеристики». Биотехнологические достижения . 31 (3): 396–401. doi : 10.1016/j.biotechadv.2011.12.001 . PMID 22193280 .