EGF-подобный домен
В этой статье отсутствует информация о консенсусном цистинном повторении, положении диффульфидной связи (остатки, Intra/Intra?). ( Март 2019 г. ) |
| EGF-подобный домен | |||
|---|---|---|---|
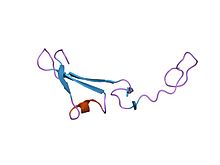 Структура эпидермального фактора роста, подобного домену хрена-альфа. [ 1 ] | |||
| Идентификаторы | |||
| Символ | Egf | ||
| Pfam | PF00008 | ||
| PFAM клан | CL0001 | ||
| Эко | 389.1.1 | ||
| InterPro | IPR000742 | ||
| PROSITE | PDOC00021 | ||
| Краткое содержание | 1apo / scope / supfam | ||
| CDD | CD00053 | ||
| |||
| EGF-подобный домен, внеклеточный | |||
|---|---|---|---|
 Кристаллическая структура внеклеточного сегмента интегрина альфавбета3 | |||
| Идентификаторы | |||
| Символ | EGF_2 | ||
| Pfam | PF07974 | ||
| PFAM клан | CL0001 | ||
| InterPro | IPR013111 | ||
| CDD | CD00054 | ||
| |||
EGF -подобный домен представляет собой эволюционный консервативный белковый домен , который получает свое название от эпидермального фактора роста , где он был впервые описан. Он состоит из от 30 до 40 аминокислотных остатков и был обнаружен в большом количестве в основном животных белков. [ 2 ] [ 3 ] Большинство случаев EGF-подобного домена обнаружены во внеклеточном домене мембранных белков или в белках, которые, как известно, секретируются . Исключением из этого является простагландин-эндопероксида синтаза . EGF-подобный домен включает в себя 6 остатков цистеина , которые в эпидермальном факторе роста, как было показано, образуют 3 дисульфидные связи . Структуры 4-дисульфидных доменов EGF были решены из белков ламинина и интегрина . Основной структурой EGF-подобных доменов представляет собой двухцепочечный β-лист с последующей петлей до короткого C-концевого двухцепочечного β-листа. Эти два β-листа обычно обозначаются как основные (N-концевые) и незначительные (С-концевые) листы. [ 4 ] EGF-подобные домены часто встречаются в многочисленных тандемных копиях в белках: эти повтора обычно складываются вместе, образуя один линейный блок соленоидного домена в качестве функциональной единицы.
Подтипы
[ редактировать ]Несмотря на сходство доменов EGF, были идентифицированы отдельные доменные подтипы. [ 5 ] Два основных предложенных типа EGF-подобных доменов-это домен, похожий на EGF (HEGF), и домен C1R-подобного (CEGF) комплемента (CEGF), [ 4 ] который был впервые идентифицирован в протеазе человека C1R. [ 5 ] C1R - очень специфическая сериновая протеаза, инициирующая классический путь активации комплемента во время иммунного ответа. [ 6 ] И HEGF-, и CEGF-подобные домены содержат три дисульфида и происходят от общего предка, который нес четыре дисульфида, из которых один был потерян во время эволюции. Кроме того, CEGF-подобные домены можно разделить на два подтипа (1 и 2), тогда как все HEGF-подобные домены принадлежат одному подтипу. [ 4 ]
Дифференциация CEGF-подобных и HEGF-подобных доменов и их подтипов основана на структурных особенностях и связности их дисульфидных связей. CEGF- и HEGF-подобные домены имеют особую форму и ориентацию незначительного листа, а одна С-концевая полуцистина имеет другую позицию. Потерянные цистеины общего предка различаются между доменами CEGF и HEGF, и, следовательно, эти типы различаются по их дисульфидным связям. Дифференциация CEGF в подтип 1 и 2, которые, вероятно, произошли после его распределения от HEGF, основана на различных числах остатков между отдельными полусустинами. N-терминал, расположенный мотив связывания кальция, можно найти в HEGF-, а также в CEGF-подобных доменах и, следовательно, не подходит для их отделения. [ 4 ]
hEGF- and cEGF-like domains also contain post-translational modifications, which are often unusual and differ between hEGF- and cEGF-like domains. These post-translational modifications include O-glycosylations, mostly O-fucose modifications, and β-hydroxylation of aspartate and asparagine residues. O-fucose modifications have only been detected in hEGF-like domains and they are important for the proper folding of the hEGF-like domain. β-Hydroxylation appears in hEGF- and cEGF-like domains, the former is hydroxylated on an aspartic acid while the latter is hydroxylated on an asparagine residue. The biological role of this post-translational modification is unclear,[4] but mice with a knockout of the aspartyl-β-hydroxylation enzyme show developmental defects.[7]
Proteins containing EGF-like domains are widespread and can be exclusively hEGF- or cEGF-like, or contain a mix of both. In many mitogenic and developmental proteins such as Notch and Delta the EGF-like domains are only of the hEGF type. Other proteins contain only cEGF such as thrombomodulin and the LDL-receptor. In mixed EGF-proteins the hEGF- and cEGF-like domains are grouped together with the hEGFs always being N-terminal of the cEGFs. Such proteins are involved in blood coagulation or are components of the extracellular matrix like fibrillin and LTBP-1 (Latent-transforming growth factor beta-binding protein 1). In addition to the aforementioned three disulfide hEGF- and cEGF-like types, there are proteins carrying a four-disulfide EGF-like domain like laminin and integrin.[4]
The two main EGF-like domain subtypes hEGF and cEGF are not just distinct in their structure and conformation but also have different functions. This hypothesis is substantiated by research on LTBP-1. LTBP-1 anchors the transforming growth factor β (TGF-β) to the extracellular matrix. hEGF-like domains play a role in targeting the LTBP-1/TGF-β assembly to the extracellular matrix. Once attached to the extracellular matrix, TGF-β dissociates from hEGF subunits to allow its subsequent activation. cEGF-like domains seem to play an unspecific role in this activation by promoting the cleavage of LTBP-1 from TGF-β by various proteases.[4]
In conclusion, although distinct EGF-like domains are grouped, subtypes can be clearly separated by their sequence, conformation and, most importantly, their function.
Role in the immune system and apoptosis
[edit]Selectins, a group of proteins that are involved in leukocyte rolling towards a source of inflammation, contain an EGF-like domain along with a lectin domain and short consensus repeats (SCRs).[8][9] The functions of the EGF-like domain vary between different selectin types. Kansas and co-workers were able to show that the EGF-like domain is not required for maximal cellular adhesion in L-selectin (expressed on lymphocytes). However, it is involved in both ligand recognition and adhesion in P-selectin (expressed on platelets) and may also be involved in protein-protein interactions. It has been suggested that the interactions between lectin domains and carbohydrate ligands might be calcium-dependent.[8]
Immature human dendritic cells appear to require interactions with the EGF-like domains of selectins during their maturation process. Blocking of this interaction with monoclonal anti-EGF-like domain antibodies prevents dendritic cell maturation. The immature cells fail to activate T-cells and produce less interleukin 12 than wild-type dendritic cells.[10]
Phan et al. could show that the artificial insertion of an N-glycosylation site into the EGF-like domains in P- and L-selectins increased the affinities of selectins to their ligands and led to slower rolling.[9] Therefore, EGF-like domains seem to play a crucial role in leukocyte movements towards inflammatory stimuli.
The EGF-like domain is also part of laminins, an important group of extracellular proteins. The EGF-like domains are usually masked in intact membranes, but become exposed when the membrane is destroyed, e.g. during inflammation, thereby stimulating membrane growth and restoring damaged membrane parts.[11]
Moreover, the EGF-like domain repeats of the stabilin-2 domain have been shown to specifically recognize and bind apoptotic cells, probably by recognizing phosphatidylserine, an apoptotic cell marker (“eat me-signal”).[12] Park et al. further demonstrated that the domains are able to competitively impair recognition of apoptotic cells by macrophages.
In conclusion, the EGF-like domain appears to play a vital role in immune responses as well as in eliminating dead cells in the organism.
Calcium-binding
[edit]Calcium-binding EGF-like domains (cbEGF-like domains) play a seminal role in diseases such as the Marfan syndrome[13] or the X-chromosome linked hemorrhagic disorder hemophilia B[14] and are among the most abundant extracellular calcium-binding domains.[15] Importantly, cbEGF- like domains impart specific functions to a variety of proteins in the blood clotting cascade. Examples include the coagulation factors VII, IX and X, protein C and its cofactor protein S.[15]
Calcium-binding EGF-like domains are typically composed of 45 amino acids, arranged as two antiparallel beta sheets.[15] Several cysteine residues within this sequence form disulfide bridges.
cbEGF-like domains show no significant structural deviations from EGF-like domains; however, as the name suggests, cbEGF-like domains bind a single calcium ion. The binding affinity to calcium varies widely and often depends on adjacent domains.[15] The consensus motif for calcium binding is Asp-Leu/Ile-Asp-Gln-Cys. Coordination of calcium strongly correlates with an unusual posttranslational modification of cbEGF-like domains: either an asparagine or aspartate is beta-hydroxylated giving rise to erythro-beta-hydroxyasparagine (Hyn) or erythro-beta-hydroxyaspartic acid (Hya), respectively. Hya can be found in the N-terminal cbEGF module (see below) of factors IX, X, and protein C. The Hyn modification appears to be more prevalent than Hya and has been shown to occur in fibrillin-1, an extracellular matrix protein.[16] Both modifications are catalyzed by the dioxygenase Asp/Asn-beta-hydroxylase,[17] and are unique to EGF domains in eukaryotes.[15]
Further posttranslational modifications have been reported. Glycosylation in the form of O-linked di- or trisaccharides may occur at a serine residue between the first two cysteines of blood coagulation factors VII and IX.[18][19][20] Factor VII exhibits an O-linked fucose at Ser60.[20]
Multiple cbEGF domains are often connected by one or two amino acids to form larger, repetitive arrays, here referred to as 'cbEGF modules'. In the blood-clotting cascade, coagulation factors VII, IX and X and protein C contain a tandem of two cbEGF modules, whereas protein S has four. Impressively, in fibrillin-1 and fibrillin-2, 43 cbEGF modules have been found.[21] The modularity of these proteins adds complexity to protein-protein but also module-module interaction. In factors VII, IX and X, the two cbEGF modules are preceded by an N-terminal gamma-carboxyglutamic acid (Gla) containing module (the Gla module).[15] In vitro studies on the Gla-cbEGF tandem isolated from factor X revealed a Kd-value of 0.1 mM for calcium binding [18] with the free calcium blood plasma concentrations being approximately 1.2 mM. Surprisingly, in the absence of the Gla module, the cbEGF module exhibits a Kd-value of 2.2 mM for calcium.[17] Thus, the presence of the Gla module increases calcium affinity 20-fold. Similarly, the activity of Gla and serine protease modules are modified by the cbEGF modules. In the absence of calcium, the Gla and cbEGF modules are highly mobile. As the cbEGF module associates with calcium, however, movement of the Gla module is significantly restricted because the cbEGF module now adopts a conformation that locks the neighboring Gla module in a fixed position.[22][23] Therefore, calcium coordination induces conformational changes which, in turn, might modulate enzymatic activity.
Impaired coordination of calcium can result in serious disorders. Defective calcium binding to coagulation factor IX contributes to the development of hemophilia B. Individuals with this hereditary disease tend to develop hemorrhages, potentially leading to life-threatening conditions. The cause of hemophilia B is decreased activity or deficiency of blood coagulation factor IX. Point mutations resulting in decreased affinity of factor IX to calcium are thought to be implicated in this bleeding disorder.[15] On a molecular basis, it appears that hemophilia B can be the result of an impaired ability to localize the Gla module efficiently, as it usually occurs after calcium coordination by the cbEGF module in fully functional factor IX.[15] This defect is thought to impair the biological function of factor IX. A similar problem occurs in patients with hemophilia B and carrying a mutation (Glu78Lys) in factor IX that prevents interaction of the two cbEGF modules with one another.[15] Conversely, in healthy individuals, Glu78 in the first cbEGF-module contacts Arg94 in the second cbEGF module and thereby aligns both modules.[24] Thus, domain-domain interactions (partially facilitated by calcium coordination) are crucial for the catalytic activity of proteins involved in the blood-clotting cascade.
Proteins containing this domain
[edit]Below is a list of human proteins containing the EGF-like domain:
- AGC1; AGRIN; AREG; ATRN; ATRNL1;
- BCAN; BMP1; BTC;
- C1S; CASPR4; CD248; CD93; CELSR1; CELSR2; CELSR3; CLEC14A; CNTNAP1; CNTNAP2; CNTNAP3; CNTNAP4; CNTNAP5; COMP; COX-2; CRB1; CRB2; CSPG3; CUBN;
- DLK1; DLL1; DLL3; DLL4; DNER;
- EDIL3; EGF; EGFL11; EGFL8; EGFL9; EGFLAM; EPGN; EREG;
- F7; F9; F10; F12; FAT; FAT2; FAT4; FBN1; FBN2; FBN3;
- GAS6;
- HABP2; HBEGF; HEG1; HGFAC; HMCN1; HSPG2;
- ITGB5;
- JAG1; JAG2;
- LDLR; LRP1; LRP10; LRP1B; LRP2; LRP4; LRP5; LRP6; LRP8; LTBP1; LTBP2; LTBP3; LTBP4;
- MATN1; MATN2; MATN3; MATN4; MEGF12; MEGF6; MEP1A; MEP1B; MFGE8; MMRN1; MMRN1; MUC4;
- NAGPA; NID1; NID2; NOTCH1; NOTCH2; NOTCH2NL; NOTCH3; NOTCH4; NRG1; NRG2; NRG3; NRG4; NRXN1; NRXN2; NRXN3; NTNG2;
- ODZ1; ODZ2; OIT3;
- PLAT; PP187; PROC; PROS1; PROZ; PTGS1; PTGS2;
- RAMP;
- SCUBE1; SCUBE2; SCUBE3; SEL-OB; SELE; SELL; SELP; SLIT1; SLIT2; SLIT3; SNED1; STAB1; STAB2; SVEP1;
- TECTA; TGFA; THBD; THBS1; THBS2; THBS4; TIE1; TLL1; TLL2; TMEFF1; TMEFF2; TNC; TNXB;
- UMOD;
- VASN; VCAN; VLDLR; VWA2;
- WIF1;
- ZAN;
See also
[edit]References
[edit]- ^ Nagata K, Kohda D, Hatanaka H, et al. (August 1994). "Solution structure of the epidermal growth factor-like domain of heregulin-alpha, a ligand for p180erbB-4". EMBO J. 13 (15): 3517–23. doi:10.1002/j.1460-2075.1994.tb06658.x. PMC 395255. PMID 8062828.
- ^ Downing AK, Knott V, Werner JM, Cardy CM, Campbell ID, Handford PA (May 1996). "Solution structure of a pair of calcium-binding epidermal growth factor-like domains: implications for the Marfan syndrome and other genetic disorders". Cell. 85 (4): 597–605. doi:10.1016/S0092-8674(00)81259-3. PMID 8653794. S2CID 15410014.
- ^ Bork P, Downing AK, Kieffer B, Campbell ID (May 1996). "Structure and distribution of modules in extracellular proteins". Q. Rev. Biophys. 29 (2): 119–67. doi:10.1017/S0033583500005783. PMID 8870072. S2CID 6104446.
- ^ Jump up to: a b c d e f g Wouters MA, Rigoutsos I, Chu CK, Feng LL, Sparrow DB, Dunwoodie SL (2005). "Evolution of distinct EGF domains with specific functions". Protein Science. 14 (4): 1091–103. doi:10.1110/ps.041207005. PMC 2253431. PMID 15772310.
- ^ Jump up to: a b Bersch B, Hernandez JF, Marion D, Arlaud GJ (1998). "Solution Structure of the Epidermal Growth Factor (EGF)-like Module of Human Complement Protease C1r, an Atypical Member of the EGF Family". Biochemistry. 37 (5): 1204–14. doi:10.1021/bi971851v. PMID 9477945.
- ^ Circolo A, Garnier G, Volanakis JE (2003). "A novel murine complement-related gene encoding a C1r-like serum protein". Molecular Immunology. 39 (14): 899–906. doi:10.1016/S0161-5890(02)00283-3. PMID 12686506.
- ^ Stenflo J, Ohlin AK, Owen WG, Schneider WJ (1988). "beta-Hydroxyaspartic acid or beta-hydroxyasparagine in bovine low density lipoprotein receptor and in bovine thrombomodulin". Journal of Biological Chemistry. 263 (1): 21–24. doi:10.1016/S0021-9258(19)57350-1. PMID 2826439.
- ^ Jump up to: a b Канзас Г.С., Сондерс К.Б., Лей К. и др. (1994). «Роль эпидермального фактора роста, подобного домену P-селектина в распознавании лиганда и клеточной адгезии» . J Cell Biol . 124 (4): 609–18. doi : 10.1083/jcb.124.4.609 . PMC 2119911 . PMID 7508943 .
- ^ Jump up to: а беременный Фан Ут, Уолдрон Т.Т., Спрингер Т.А. (2006). «Реконструкция границы раздела лектин-EGF-подобного домена в P- и L-селектине увеличивает адгезивность и устойчивость к сдвигу при гидродинамической силе» . Nat Immunol . 7 (8): 883–9. doi : 10.1038/ni1366 . PMC 1764822 . PMID 16845394 .
- ^ Zhou T, Zhang Y, Sun G, et al. (2006). «Моноклональное антитело домена анти-P-селектина Lectin-EGF ингибирует созревание незрелых дендритных клеток человека». Exp Mol Pathol . 80 (2): 171–6. doi : 10.1016/j.yexmp.2005.10.004 . PMID 16413535 .
- ^ Löffler, G; Петридес, PE; Heinrich, PC (1997). Биохимия и патобиохимия (5 -е изд.). Берлин, Гейдельберг: Springer-Verlag. п. 747. ISBN 3-540-59006-4 .
- ^ Park Sy, Kim Sy, Jung My, et al. (2008). «Эпидермальный фактор роста, подобный доменному доменному повторению табилина-2, распознает фосфатидилсерин во время клиренса клеточного трупа» . Мол -клеточная биол . 28 (17): 5288–98. doi : 10.1128/mcb.01993-07 . PMC 2519725 . PMID 18573870 .
- ^ Хандфорд П.А., Даунинг А.К., Рао З., Хьюетт Д.Р., Сайкес Б.К., Килти С.М. (1991). «Свойства связывания кальция и молекулярная организация эпидермальных доменов, подобных факторам роста, у фибриллина-1 человека» . Дж. Биол. Химический 270 (12): 6751–6. doi : 10.1074/jbc.270.12.6751 . PMID 7896820 .
- ^ Хандфорд П.А., Мэйхью М., Барон М., Виншип П.Р., Кэмпбелл И.Д., Браунли Г.Г. (1991). «Ключевые остатки, участвующие в мотивах связывания кальция в EGF-подобных доменах». Природа . 351 (6322): 164–7. Bibcode : 1991natur.351..164H . doi : 10.1038/351164A0 . PMID 2030732 . S2CID 4338236 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Stenflo J, Stenberg Y, Muranyi A (2000). «Кальций-связывающие EGF-подобные модули в коагуляционных протеиназах: функция иона кальция в модульных взаимодействиях». Biochimica et Biophysica Acta (BBA) - структура белка и молекулярная ферма . 1477 (1–2): 51–63. doi : 10.1016/s0167-4838 (99) 00262-9 . PMID 10708848 .
- ^ Glanville RW, Qian RQ, McClure DW, Maslen CL, et al. (1994). «Связывание кальция, гидроксилирование и гликозилирование эпидермальных факторов роста предшественника, подобных фактору роста, фибриллина-1, белка гена Марфана» . Дж. Биол. Химический 269 (43): 26630–4. doi : 10.1016/s0021-9258 (18) 47065-2 . PMID 7929395 .
- ^ Jump up to: а беременный Jia S, Vandusen WJ, Diehl Re, et al. (1992). «Клонирование кДНК и экспрессия бычьего аспартил (аспарагинил) бета-гидроксилазы» . Дж. Биол. Химический 267 (20): 14322–7. doi : 10.1016/s0021-9258 (19) 49715-9 . PMID 1378441 .
- ^ Jump up to: а беременный Valcarce C, Selander-Sunnerhagen M, Tämlitz AM, Drakenberg T, Björk I, Stenflo J (1996). «Аффинность кальция NH2-концевого эпидермального фактора роста, подобного фактору, фактора X» . Дж. Биол. Химический 268 (35): 26673–8. doi : 10.1016/s0021-9258 (19) 74365-8 . PMID 8253800 .
- ^ Nishimura H, Kawabata S, Kisiel W, et al. (1989). «Идентификация дисахаридного (XYL-GLC) и трисахаридного (XYL2-GLC) O-гликозидного, связанного с сериновым остатком в первом эпидермальном факторе роста, подобном домену человеческих факторов VII и IX и белка Z и бычьего белка Z» . Дж. Биол. Химический 264 (34): 20320–5. doi : 10.1016/s0021-9258 (19) 47065-8 . PMID 2511201 .
- ^ Jump up to: а беременный Bjoern S, Foster D, Thim L, et al. (1991). «Человеческая плазма и рекомбинантный фактор VII» . Дж. Биол. Химический 266 (17): 11051–7. Doi : 10.1016/s0021-9258 (18) 99126-x . PMID 1904059 .
- ^ Piha-Gossack A, Sossin W, Reinhardt DT, et al. (2012). «Эволюция внеклеточных фибриллинов и их функциональных доменов» . Plos один . 7 (3): 33560. Bibcode : 2012ploso ... 733560p . doi : 10.1371/journal.pone.0033560 . PMC 3306419 . PMID 22438950 .
- ^ Sunnerhagen M, Forsen S, Hoffren A, Drakenberg T, Teleman O, Stenflo J (1995). «Структура Ca (2+)-свободный домен GLA проливает свет на мембрановое связывание белков коагуляции крови». Природа структурная и молекулярная биология . 2 (6): 504–9. doi : 10.1038/nsb0695-504 . PMID 7664114 . S2CID 8570806 .
- ^ Sunnerhagen M, Olah GA, Stenflo J, Forsen S, Drakenberg T, Trewhella J (1996). «Относительная ориентация доменов GLA и EGF в коагуляционном факторе x изменяется при связывании Ca2+ с первым доменом EGF. Объединенное исследование рентгеновского рассеяния NMR-Small Angle» ». Биохимия . 35 (36): 11547–59. doi : 10.1021/bi960633j . PMID 8794734 .
- ^ Кристоф О.Д., одолженность П.Дж., Колкман Дж.А., Браунли Г.Г., Мертенс К. (1988). «Остатки коагуляции крови IX Glu78 и Arg94 обеспечивают связь между обоими эпидермальными доменами, подобными факторам роста, которая имеет решающее значение при взаимодействии с легкой цепью фактора VIII» . Дж. Биол. Химический 273 (1): 222–27. doi : 10.1074/jbc.273.1.222 . PMID 9417068 .