Тецелозавр
| Тецелозавр Временной диапазон: поздний мел ,
| |
|---|---|

| |
| Реконструированный скелет, Музей естественной истории Берпи. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Клэйд : | † Неорнитишия |
| Семья: | † Тесцелозавриды |
| Подсемейство: | † Тесцелозаврины |
| Род: | † Тесцелозавр Гилмор , 1913 год. [ 1 ] |
| Типовой вид | |
| † Тесцелозавр заброшен Гилмор, 1913 год.
| |
| Другие виды | |
| Синонимы | |
Тесцелозавр ( / ˌ θ ɛ s ɪ l ə ˈ s ɔː r ə s / THESS -il -ə- SOR -əs ; древнегреческий θέσκελος - ( theskelos - ) означает «богоподобный», «чудесный» или «чудесный» и σαυρος ( саурос ) «ящерица») [ 5 ] — род неорнитишевых , динозавров появившийся в самом конце позднего мелового периода в Северной Америке. Он был представителем последней фауны динозавров перед мел-палеогеновым вымиранием около 66 миллионов лет назад. Сохранность и полнота многих экземпляров указывают на то, что он, возможно, предпочитал жить возле ручьев.
Этот двуногий неорнитиш известен по нескольким частичным скелетам и черепам, которые указывают на то, что его длина в среднем составляла от 2,5 до 4,0 метров (от 8,2 до 13,1 футов). У него были крепкие задние конечности, маленькие широкие руки и голова с вытянутой заостренной мордой. Форма . зубов и челюстей позволяет предположить, что это преимущественно травоядное животное Этот род динозавров считается специализированным неорнитишием, традиционно описываемым как гипсилофодонт , но совсем недавно признанным отличным от гипсилофодона . Для этого рода было предложено несколько видов. В настоящее время признаны действительными три вида : типовой вид T. ignoreus , T. garbanii и T. assiniboiensis .
Этот род привлек внимание средств массовой информации в 2000 году, когда экземпляр, обнаруженный в 1993 году в Южной Дакоте , США, был интерпретирован как включающий окаменелое сердце. Было много дискуссий по поводу того, принадлежали ли останки сердцу. Многие учёные сейчас сомневаются в идентификации объекта и последствиях такой идентификации.
Открытие, история и виды
[ редактировать ]
Типовой экземпляр Thescelosaurus и Уильямом ( USNM 7757) был обнаружен в 1891 году палеонтологами Джоном Беллом Хэтчером Х. Аттербеком в отложениях верхнемеловой маастрихта в позднего формации Ланс округе Ниобрара (в то время входившей в состав округа Конверс ), штат Вайоминг. , США . Скелет, однако, оставался в транспортировочных ящиках в течение многих лет, пока Чарльз У. Гилмор из Смитсоновского института не Национального музея естественной истории подготовил его и не описал в короткой статье в 1913 году, назвав его T. ignoreus ( ignoreus : "). пренебречь»). В то время он думал, что это связано с камптозавром . [ 1 ] В 1915 году он представил подробную монографию , описывающую хорошо сохранившийся скелет. [ 6 ] Типовой экземпляр был найден в основном с естественным сочленением, и отсутствовали только голова и шея, которые были потеряны из-за эрозии . [ 6 ] Название происходит от удивления, которое испытал Гилмор, обнаружив такой хороший экземпляр, который так долго оставался без присмотра. Он считал его легким, подвижным существом и относил к Hypsilophodontidae — семейству мелких двуногих динозавров. [ 6 ]

Другие останки подобных животных находили на протяжении конца 19 и 20 веков. Другой хорошо сохранившийся скелет из немного более старой формации Хорсшу-Каньон в Альберте , Канада , был назван T. warreni Уильямом Парксом в 1926 году. [ 7 ] Этот скелет имел заметные отличия от T. ignoreus , и поэтому Чарльз М. Штернберг в 1937 году поместил его в новый род Parksosaurus . [ 8 ] Штернберг также назвал дополнительный вид, T. edmontonensis , на основе другого сочлененного скелета, на этот раз включающего частичный череп ( NMC 8537), и обратил внимание на тяжелое телосложение и толстые кости этого рода. Из-за этих отличий от обычного легкого строения гипсилофодонтов он предположил, что этот род заслуживает подсемейства отдельного Thescelosaurinae . [ 9 ] T. edmontonensis , начиная с обзора Питера Гальтона 1974 года, обычно считался более устойчивой особью (возможно, противоположного пола типовой особи). [ 10 ] пренебречь Т. [ 11 ] [ 12 ] Однако Бойд и его коллеги обнаружили, что они не могут отнести его ни к одному из действующих видов Thescelosaurus, и посчитали, что этот экземпляр относится к неопределённому роду. [ 4 ] Другим предметом разногласий относительно T. edmontonensis является его лодыжка, которая, по утверждению Гальтона, была повреждена и неверно истолкована, но которую Уильям Дж. Моррис (1976) считал действительно отличной от T. ignoreus . [ 2 ]

В своей статье Моррис описал экземпляр ( SDSM 7210), состоящий из частичного черепа с тяжелыми гребнями на нижней челюсти и щеке, четырех частичных позвонков и двух костей пальцев , как неопознанный вид Thescelosaurus -Крик позднего маастрихта. , из Хелл Образование округа Хардинг , Южная Дакота , США. Он обратил внимание на предчелюстные зубы и глубоко посаженные зубы, которые, по его мнению, подтверждают наличие мускулистых щек. Моррис также указал на расширяющуюся наружу предчелюстную кость (что давало ей широкий клюв) и большие глазные яблоки. [ 2 ] Этот череп на протяжении многих лет признавался безымянным гипсилофодонтом. [ 12 ] пока Гальтон не сделал его типовым экземпляром нового рода и вида Bugenasaura infernalis («крупнощекая ящерица, принадлежащая нижним регионам», infernalis является отсылкой к формации Хелл-Крик). [ 13 ] Моррис также назвал новый возможный вид Thescelosaurus для экземпляра LACM 33542: ? T. garbanii (со знаком вопроса, поскольку он не был уверен в принадлежности этого рода). LACM 33542 включал большую частичную заднюю конечность («на треть крупнее, чем описанные экземпляры T. ignoreus и Parksosaurus, или почти в два раза больше, чем Hypsilophodon »), включая ступню, предплюсну , кости голени и части бедренной кости, а также пять шейных (шейных) костей. ) и одиннадцать спинных (задних) позвонков из формации Хелл-Крик округа Гарфилд , штат Монтана , США. Образец был обнаружен палеонтологом- любителем Харли Гарбани , отсюда и название. T. garbanii имел длину около 4,5 метров (15 футов), что больше, чем у средних экземпляров T. ignoreus . Помимо размера, Моррис обратил внимание на конструкцию лодыжки, которую он считал уникальной, за исключением Thescelosaurus edmontonensis , которую он считал отдельным видом. Потому что Моррис считал, что лодыжки T. garbanii . T. garbanii выгодно отличаются от лодыжек T. edmontonensis , он предварительно отнес его к Thescelosaurus . [ 2 ] Однако в научной литературе поддерживается точка зрения Гальтона о том, что T. edmontonensis не отличается от T. ignoreus (см. выше). В той же статье, в которой он описал Bugenasaura , Гальтон продемонстрировал, что черты, которые, по мнению Морриса, связывали T. garbanii и T. edmontonensis, были результатом повреждения лодыжки последнего, поэтому T. garbanii также можно считать отличным от Thescelosaurus . Чтобы лучше приспособить этот вид, Гальтон предположил, что он принадлежал к его новому роду Bugenasaura как B. garbanii , хотя он также отметил, что он мог принадлежать пахицефалозавриду аналогичного размера Stygimoloch или быть частью третьего, неизвестного динозавра. [ 13 ]

Клинт Бойд и его коллеги опубликовали повторную оценку Thescelosaurus , Bugenasaura и Parksosaurus в 2009 году, используя в качестве отправной точки новый черепной материал. Они обнаружили, что Parksosaurus действительно отличался от Thescelosaurus , и что череп Bugenasaura infernalis был по существу таким же, как череп, найденный с посткраниальным скелетом, который соответствовал Thescelosaurus . Поскольку B. infernalis нельзя было отличить от Thescelosaurus , они считали этот род синонимом Thescelosaurus , вид сомнительным , а SDSM 7210 — примером T. sp. Они обнаружили, что LACM 33542, хотя и фрагментарный, был экземпляром Thescelosaurus , и согласились с Моррисом, что структура лодыжки была отчетливой, вернув ее T. garbanii . Наконец, они отметили, что другой экземпляр, RSM P.1225.1, отличается от T. ignores в некоторых анатомических деталях и может представлять собой новый вид. Таким образом, Thescelosaurus согласно Boyd et al. (2009) представлен как минимум двумя, а возможно, и тремя действительными видами: типовыми видами T. ignoreus , T. garbanii и возможный безымянный вид. [ 4 ] В декабре 2011 года RSM P.1225.1 был отнесен к собственному виду Thescelosaurus assiniboiensis . Он был назван Калебом М. Брауном, Клинтом А. Бойдом и Энтони П. Расселом и известен только по своему голотипу, небольшому, сочленённому и почти полному скелету из формации Френчмен (поздний маастрихтский этап) Саскачевана . [ 3 ] В апреле 2022 года сообщалось, что экземпляр тесцелозавра был найден на местонахождении окаменелостей Танис , предположительно датируемый точным днем вымирания K-Pg , что сделало его первой окаменелостью нептичьего динозавра, обнаруженной с этой даты. [ 14 ] [ 15 ]
Описание
[ редактировать ]
В целом анатомия скелета этого рода хорошо документирована, а реставрации были опубликованы в нескольких статьях, включая реставрации скелета. [ 4 ] [ 6 ] [ 10 ] [ 16 ] и модели. [ 9 ] [ 6 ] Скелет известен настолько хорошо, что была проведена детальная реконструкция мышц бедра и задних конечностей. [ 17 ] Размер животного оценивается в диапазоне 2,5–4,0 м по длине (8,2–13,1 фута). [ 10 ] для различных экземпляров и весом 200–300 килограммов (450–660 фунтов ), [ 18 ] с крупным типовым экземпляром T. garbanii, длина которого оценивается в 4–4,5 метра (13,1–14,8 футов). [ 2 ] Как более подробно обсуждалось в разделе « Открытие, история и виды », он мог быть сексуально диморфным : один пол крупнее другого. [ 10 ] Молодые останки известны в нескольких местах, в основном это зубы. [ 19 ] [ 20 ]
Тесцелозавр был массивным двуногим животным, вероятно, травоядным, но потенциально нет. [ 11 ] имелся выступающий гребень . По длине обеих верхних челюстей (несущие зубы «щечные» кости) [ 4 ] и гребень на обеих зубных железах (зубная кость нижней челюсти). [ 21 ] Хребты [ 21 ] и положение зубов, расположенных глубоко внутри внешней поверхности черепа, интерпретируются как свидетельство наличия мускулистых щек. [ 2 ] Помимо длинного узкого клюва, череп также имел зубы в предчелюстной кости , или верхнем клюве (примитивный признак среди неорнитиший). [ 22 ] длинные стержнеобразные кости, называемые пальпебралами , из-за чего у животного были тяжелые костлявые брови. Над глазами располагались [ 22 ] Его зубы были двух типов : маленькие заостренные предчелюстные зубы и листовидные щечные зубы. [ 9 ] В обеих предчелюстных костях имелось по шесть мелких зубов, а на кончике клюва имелся беззубый участок. [ 4 ]

У тесцелозавров были короткие, широкие, пятипалые руки, четырехпалые ступни с пальцев копытеобразными кончиками и длинный хвост, укрепленный окостеневшими сухожилиями от середины до кончика, что уменьшало гибкость хвоста. [ 6 ] Грудная клетка была широкой, что обеспечивало широкую спину, а конечности были крепкими. [ 9 ] Животные, возможно, были способны передвигаться на четвереньках , учитывая их довольно длинные руки и широкие кисти. [ 10 ] но эта идея не получила широкого обсуждения в научной литературе, хотя и встречается в популярных работах. [ 23 ] [ 24 ] Чарльз М. Штернберг реконструировал его, расположив плечо почти перпендикулярно телу. [ 9 ] еще одна идея, которая ушла на второй план. Как отметил Питер Гальтон , кость плеча большинства птицетазовых животных сочленялась с плечом посредством суставной поверхности, которая состояла из всего конца кости, а не из отдельного шара и гнезда, как у млекопитающих. Ориентация суставной поверхности плеча также указывает на вертикальное, а не горизонтальное плечо динозавров. [ 25 ]
Рядом с бортами ребер обнаружены крупные тонкие плоские минерализованные пластины. [ 26 ] Их функция неизвестна; они, возможно, играли роль в дыхании . [ 27 ] Однако на пластинах не обнаружено мышечных рубцов или других признаков прикрепления, что свидетельствует против дыхательной функции. Недавнее гистологическое исследование слоистых пластинок вероятного юноши показало, что они, возможно, начинались как хрящи , а по мере старения животного превращались в кости. [ 28 ] Такие пластинки известны у ряда других церапод . [ 29 ]
На протяжении большей части истории природа покровов этого рода , будь то чешуя или что-то еще, оставалась неизвестной. Чарльз Гилмор описал участки карбонизированного материала возле плеч как возможный эпидермис с «проколотой» текстурой, но без регулярного рисунка. [ 6 ] в то время как Уильям Дж. Моррис предположил, что панцирь присутствовал в виде небольших щитков, которые он интерпретировал как расположенные по крайней мере вдоль средней линии шеи одного экземпляра. [ 2 ] Однако щитки не были обнаружены у других членистоногих экземпляров Thescelosaurus , и щитки Морриса могли быть крокодиловыми по происхождению. [ 29 ] В своем документальном фильме 2022 года «Динозавры: Последний день » сэр Дэвид Аттенборо сообщил об образце тесцелозавра , предположительно убитом в день вымирания K-Pg , покрытом отпечатками кожи, в том числе удлиненными чешуйками на ногах. Один из палеонтологов, проводивший раскопки, предположил, что они выполняли камуфляжную функцию. В последующем интервью Пол Барретт отметил, что это означает, что тесцелозавр был не таким оперенным, как предполагалось для других мелких неорнитиший. [ 30 ]
Классификация
[ редактировать ]
Тесцелозавр обычно был связан с гипсилофодоном и другими мелкими неорнитисхиями как гипсилофодонтид , хотя он выделялся среди них своим крепким телосложением, необычными задними конечностями и, в последнее время, необычно длинным черепом. [ 11 ] [ 9 ] [ 10 ] [ 12 ] Питер Гальтон в 1974 году представил один из вариантов классической конструкции, предположив, что из-за строения задних конечностей и тяжелого телосложения (не бегающего и не созданного для бега, по его определению), его следует включить в Iguanodontidae . Этого не последовало, поскольку Моррис решительно выступал против схемы классификации Гальтона. [ 2 ] В любом случае, Iguanodontidae Гальтона были полифилетическими , а не естественной группой, и поэтому не могли быть признаны в современном кладистическом использовании.
Хотя в начале 1990-х годов Hypsilophodontidae интерпретировались как естественная группа, [ 12 ] [ 31 ] эта гипотеза потеряла популярность, и было обнаружено, что Hypsilophodontidae представляют собой противоестественное семейство, состоящее из множества животных, более или менее тесно связанных с Iguanodontia ( парафилия ), с различными небольшими кладами близкородственных таксонов. [ 4 ] [ 11 ] [ 32 ] [ 33 ] [ 34 ] «Гипсилофодонтиды» и «гипсилофодонты» лучше понимать как неофициальные термины, обозначающие эволюционный уровень , а не настоящую кладу. Тесцелозавр считался одновременно очень базальным [ 31 ] и очень производное [ 11 ] среди гипсилофодонтов. Одна из проблем, которая потенциально может помешать классификации Thescelosaurus, заключается в том, что не все останки, отнесенные к T. ignoreus, обязательно принадлежат ему. [ 35 ] Клинт Бойд и его коллеги обнаружили, что, хотя клада Thescelosaurus включала род Bugenasaura существовало по крайней мере два, а возможно, и три вида и виды, которые были отнесены к этому роду, внутри Thescelosaurus отнесены к T. ignoresaurus. , и несколько экземпляров, ранее отнесенных к T. ignoresus, еще не могли быть отнесен к виду внутри рода. [ 4 ] Похоже, он тесно связан с паркзозавром . [ 4 ] [ 11 ] [ 33 ] [ 3 ] [ 36 ]
За исчезновением Hypsilophodontidae последовало признание отдельного семейства Thescelosauridae. Эта область генеалогического древа динозавров исторически была осложнена отсутствием исследований, но статьи Клинта Бойда и его коллег [ 4 ] и Калеб Браун и коллеги [ 3 ] [ 36 ] специально обратились к этим динозаврам. Бойд и др. (2009) и Браун и др. (2011) обнаружили, что североамериканские «гипсилофодонты» мелового периода разделились на два родственных кластера: один состоит из ородромеуса , ориктодромеуса и зефирозавра , а другой — из паркзозавра и тесцелозавра . [ 4 ] [ 3 ] Браун и др. (2013) получили аналогичные результаты с добавлением нового рода Albertadromeus к кладе Orodromeus и нескольких длиннорылых азиатских форм (ранее описанных как Jeholosauridae ). [ 32 ] к кладе Тесцелозавров . Они также формально определили Thescelosauridae ( Thescelosaurus ignoreus , Orodromeus makelai , их самого недавнего общего предка и всех потомков) и более мелкие клады Orodrominae и Thescelosaurinae. Приведенная ниже кладограмма принадлежит Brown et al. . [ 36 ]

| Тесцелозавриды |
| ||||||||||||||||||||||||||||||||||||||||||
Палеобиология
[ редактировать ]
Тецелозавр бродил на расстоянии примерно первого метра от земли, питаясь избирательно. [ 11 ] при удерживании пищи во рту за щеки во время пережевывания. [ 10 ] Тесцелозавр, вероятно, был медленнее других гипсилофодонтов из-за более тяжелого телосложения и строения ног. По сравнению с ними у него были необычные задние конечности, поскольку верхняя нога была длиннее голени , в противоположность гипсилофодону и бегущим животным вообще. [ 9 ] Известно, что у одного экземпляра была патология костей: длинные кости правой стопы срослись наверху, что затрудняло быстрое движение. [ 37 ] Исследование зубов Thescelosaurus и сравнение с современным пахицефалозавром Stegoceras позволяют предположить, что Thescelosaurus питался избирательно, в то время как Stegoceras питался более неразборчиво, позволяя обоим животным жить в одной и той же среде, не конкурируя за пищу. [ 38 ]
Исследования черепа тесцелозавра , в частности экземпляра «Вилло», проведенные Баттоном и Занно и др., Опубликованные в 2023 году, показывают, что его мозг был небольшим по сравнению с другими неорнитисхическими динозаврами, что позволяет предположить, что его когнитивные способности находились в пределах современных рептилий. Дополнительный анализ показал, что у него был плохой слух и было развито обоняние и чувство равновесия. В сочетании с особенностями посткраниального скелета, такими как сильные передние конечности, предполагается, что тесцелозавр , возможно, вел полуфоссорный образ жизни и, возможно, рыл норы под землей. С другой стороны, анализ может предложить филогенетически унаследованные черты от роющих предков. [ 39 ] [ 40 ]
Предполагаемое окаменелое сердце
[ редактировать ]
В 2000 году скелет этого рода (образец NCSM 15728), неофициально известный как «Вилло», ныне выставленный в Музее естественных наук Северной Каролины , был описан как включающий остатки четырехкамерного сердца и аорты . Первоначально он был обнаружен в 1993 году на северо-западе Южной Дакоты. Авторы обнаружили внутренние детали с помощью изображений компьютерной томографии (КТ). Они предположили, что сердце было омылено (превращено в могильный воск ) в безвоздушных условиях захоронения, а затем изменено на гетит , железный минерал, путем замены исходного материала. Авторы интерпретировали структуру сердца как указание на повышенную скорость метаболизма тесцелозавра на , а не хладнокровность рептилий . [ 26 ]
Их выводы были оспорены; вскоре после первоначального описания другие исследователи опубликовали статью, в которой утверждали, что сердце на самом деле представляет собой конкремент . Как они отметили, приведенная для объекта анатомия неверна (например, "аорта" сужается, переходя в "сердце" и лишена идущих от него артерий ), частично захватывает одно из ребер и имеет внутреннее строение из концентрических слоев. кое-где и сзади правой ножки сохранился еще один конкремент. [ 41 ] Первоначальные авторы отстаивали свою позицию; они согласились, что это был тип конкремента, но он сформировался вокруг и частично сохранил более мышечные части сердца и аорты. [ 42 ]

Исследование, опубликованное в 2011 году, применило несколько направлений исследования к вопросу идентичности объекта, включая более продвинутое компьютерное сканирование, гистологию , рентгеновскую дифракцию , рентгеновскую фотоэлектронную спектроскопию и сканирующую электронную микроскопию. В результате этих методов авторы установили следующее: внутренняя структура объекта не включает камеры, а состоит из трех несоединенных областей материала меньшей плотности и не сравнима со структурой страусиного сердца ; «стены» состоят из осадочных минералов, которые, как известно, не образуются в биологических системах, таких как гетит, полевого шпата минералы , кварц и гипс , а также некоторых фрагментов растений; углерод , азот и фосфор в их образцах отсутствовали — химические элементы, важные для жизни; клеточные структуры сердца отсутствовали. Был один возможный участок с клеточными структурами животных. Авторы обнаружили, что их данные подтверждают идентификацию как конкрецию песка из среды захоронения, а не сердца, с возможностью сохранения отдельных участков тканей. [ 43 ]
Однако вопрос о том, как эта находка отражает скорость метаболизма и внутреннюю анатомию динозавра, остается спорным, независимо от личности объекта. [ 43 ] И современные крокодилы , и птицы , ближайшие ныне живущие родственники нептичьих динозавров, имеют четырёхкамерное сердце (хотя и видоизменённое у крокодилов), поэтому они, вероятно, были и у нептичьих динозавров; структура не обязательно связана со скоростью метаболизма. [ 44 ]
Палеоэкология
[ редактировать ]Временной и географический диапазон
[ редактировать ]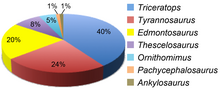
Настоящие останки тесцелозавра точно известны только из пород позднего маастрихта , из Альберты ( формация Сколлард ) и Саскачевана ( формация Френчмен ), Канады и Вайоминга ( формация Лэнс ), Южной Дакоты ( формация Хелл-Крик ), Монтаны (формация Хелл-Крик), и Колорадо ( формация Ларами ), США. [ 4 ] [ 11 ] За исключением птиц, это был один из последних родов динозавров, его останки были обнаружены всего в 3 метрах от пограничной глины, содержащей слой иридия , замыкающей меловой период. [ 45 ] Формация Ларами - самая старая формация, Тесцелозавр из которой известен , а магнитостратиграфия предполагает возраст формации Ларами 69–68 млн лет назад. [ 46 ] Имеются сообщения о зубах из более древних пород кампанского периода , особенно из формации Парка динозавров в Альберте. [ 47 ] но эти экземпляры не принадлежат Тесцелозавру , а скорее принадлежат Ородромею . [ 13 ] Для этого рода известно больше экземпляров, чем официально описано, например, экземпляр Триболда, [ 48 ] который послужил источником нескольких слепков скелетов для музеев.
Когда Гальтон вновь посетил Thescelosaurus и Bugenasaura в 1999 году, он описал зубной зуб UCMP 46911 из верхней юры Уэймута Англия , , как ср. Бугенасавр . [ 21 ] Если это действительно зуб животного, похожего на тесцелозавра, это значительно расширит стратиграфический диапазон группы.
среда обитания
[ редактировать ]
Были сделаны противоречивые сообщения относительно его предпочтительной среды обитания ; две статьи предполагают, что он предпочитал русла поймам рек , [ 49 ] [ 50 ] но другой предполагает, что они предпочли противоположное. [ 51 ] Возможное предпочтение каналов основано на относительном обилии окаменелостей тесцелозавров в песчаниках , представляющих среду русла, по сравнению с аргиллитами , представляющими среду поймы. [ 50 ] или скоплениях нескольких особей пока не О костных слоях сообщалось. Дейл Рассел в популярной работе отметил, что Тесцелозавр был самым распространенным мелким травоядным в формации Хелл-Крик в районе Форт-Пек . Он описал окружающую среду того времени как плоскую пойму с относительно сухим субтропическим климатом, в котором произрастали разнообразные растения, от покрытосеменных деревьев до лысого кипариса , папоротников и гинкго . Хотя большинство скелетов динозавров из этой области неполные, возможно, из-за низкого потенциала сохранения лесов, скелеты тесцелозавра гораздо более полные, что позволяет предположить, что этот род часто посещал ручьи ручьев. Таким образом, когда тесцелозавр умер, он мог находиться в реке или рядом с ней, что облегчило его захоронение и сохранение для последующего окаменения. Рассел предварительно сравнил его с капибарами и тапирами . [ 52 ]
Другие динозавры, разделившие одно и то же время и место, включают цератопсидов трицератопса и торозавра , гадрозаврида эдмонтозавра , анкилозавра анкилозавра , пахицефалозавра пахицефалозавра и тероподов орнитомима , троодона и тираннозавра . [ 53 ] [ 54 ] Тесцелозавр также был в изобилии в формации Ланс. Кости пальцев ног этого рода являются наиболее распространенными находками после окаменелостей трицератопса и эдмонтозавра , и, возможно, это был самый распространенный динозавр в жизни, если бы формация Ланс имела предвзятое отношение к сохранению мелких животных. [ 55 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б Гилмор, Чарльз В. (май 1913 г.). «Новый динозавр из формации Лэнс в Вайоминге» . Разные коллекции Смитсоновского института . 61 (5): 1–5.
- ^ Jump up to: а б с д и ж г час Моррис, Уильям Дж. (1976). «Гипсилофодонтные динозавры: новый вид и комментарии к их систематике» . В Черчере, CS (ред.). Атлон . Торонто: Королевский музей Онтарио. стр. 93–113 . ISBN 978-0-88854-157-4 .
- ^ Jump up to: а б с д и Браун, Калеб М.; Бойд, Клинт А.; Рассел, Энтони П. (2011). «Новый базальный динозавр-орнитопод (формация Френчмен, Саскачеван, Канада) и его значение для разнообразия орнитисхий позднего Маастрихта в Северной Америке» . Зоологический журнал Линнеевского общества . 163 (4): 1157–1198. дои : 10.1111/j.1096-3642.2011.00735.x .
- ^ Jump up to: а б с д и ж г час я дж к л Бойд, Клинт А.; Браун, Калеб М.; Шитц, Родни Д.; Кларк, Джулия А. (2009). «Таксономическая ревизия базальных неорнитишских таксонов Thescelosaurus и Bugenasaura ». Журнал палеонтологии позвоночных . 29 (3): 758–770. Бибкод : 2009JVPal..29..758B . дои : 10.1671/039.029.0328 . S2CID 84273584 .
- ^ Лидделл и Скотт (1980). Греко-английский лексикон, сокращенное издание . Издательство Оксфордского университета, Оксфорд, Великобритания. ISBN 978-0-19-910207-5 .
- ^ Jump up to: а б с д и ж г Гилмор, Чарльз В. (1915). «Остеология тесцелозавра , динозавра-ортопеда из формации Лэнс в Вайоминге» (PDF) . Труды Национального музея США . 49 (2127): 591–616. дои : 10.5479/si.00963801.49-2127.591 .
- ^ Паркс, Уильям А. (1926). « Thescelosaurus warreni , новый вид ортоподных динозавров из Эдмонтонской формации в Альберте». Исследования Университета Торонто (геологическая серия) . 21 : 1–42.
- ^ Штернберг, Чарльз М. (1937). «Классификация тесцелозавра с описанием нового вида». Труды Геологического общества Америки за 1936 год : 365.
- ^ Jump up to: а б с д и ж г Штернберг, Чарльз М. (1940). « Thescelosaurus edmontonensis , n. sp., и классификация Hypsilophodontidae». Журнал палеонтологии . 14 (5): 481–494.
- ^ Jump up to: а б с д и ж г Гальтон, Питер М. (1974). «Заметки о тесцелозавре , консервативном динозавре-орнитоподе из верхнего мела Северной Америки, с комментариями по классификации орнитопод». Журнал палеонтологии . 48 (5): 1048–1067.
- ^ Jump up to: а б с д и ж г час Норман, Дэвид Б .; Сьюс, Ханс-Дитер; Уитмер, Ларри М.; Кориа, Родольфо А. (2004). «Базальные орнитоподы». В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 393–412. ISBN 978-0-520-24209-8 .
- ^ Jump up to: а б с д Сьюс, Ханс-Дитер; Норман, Дэвид Б. (1990). «Гипсилофодонтиды, Тенонтозавры , Дриозавриды». В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия (1-е изд.). Беркли: Издательство Калифорнийского университета. стр. 498–509. ISBN 978-0-520-06727-1 .
- ^ Jump up to: а б с Гальтон, Питер М. (1995). «Вид базального гипсилофодонтидного динозавра Thescelosaurus Gilmore (Ornithischia: Ornithopoda) из позднего мела Северной Америки». Новогодний альбом по геологии и палеонтологии, Трактаты . 198 (3): 297–311. дои : 10.1127/njgpa/198/1995/297 .
- ^ Амос, Джонатан (6 апреля 2022 г.). «Танис: найдены окаменелости динозавра, погибшего в результате удара астероида, утверждают ученые» . Новости Би-би-си . Проверено 25 октября 2022 г.
- ^ Мартин, Салин. «Окаменелости динозавра, погибшего в результате удара астероида, обнаружены в Северной Дакоте, говорят ученые» . физ.орг . Проверено 25 октября 2022 г.
- ^ Бретт-Сурман, Майкл К. (1997). «Орнитоподы». В Фарлоу, Джеймс О.; Бретт-Сурман, Майкл К. (ред.). Полный динозавр . Блумингтон и Индианаполис: Издательство Университета Индианы. стр. 330–346 . ISBN 978-0-253-33349-0 .
- ^ Ромер, Альфред С. (1927). «Тазовая мускулатура птицетазовых динозавров». Акта Зоология . 8 (2–3): 225–275. дои : 10.1111/j.1463-6395.1927.tb00653.x .
- ^ Эриксон, Брюс Р. (2003). Динозавры Музея науки Миннесоты . Сент-Пол, Миннесота: Музей науки Миннесоты. п. 31.
- ^ Карпентер, Кеннет (1982). «Детеныши динозавров из позднемеловых формаций Ланс и Хелл-Крик и описание нового вида теропод». Вклад в геологию . 20 (2): 123–134.
- ^ Рассел, Дейл А .; Манабе, Макото (2002). «Краткий обзор скопления динозавров Хелл-Крик (верхний меловой период)». В Хартмане, Джозеф Х.; Джонсон, Кирк Р.; Николс, Дуглас Дж. (ред.). Формация Хелл-Крик и граница мелового и третичного периодов на Северных Великих равнинах: интегрированные континентальные записи конца мелового периода . Специальный доклад Геологического общества Америки , 361. Боулдер, Колорадо: Геологическое общество Америки. стр. 169–176. ISBN 978-0-8137-2361-7 .
- ^ Jump up to: а б с Гальтон, Питер М. (1999). «Краниальная анатомия гипсилофодонтного динозавра Bugenasaura infernalis (Ornithischia: Ornithopoda) из верхнего мела Северной Америки». Revue Paléobiologie, Женева . 18 (2): 517–534.
- ^ Jump up to: а б Гальтон, Питер М. (1997). «Краниальная анатомия базального гипсилофодонтидного динозавра Thescelosaurus ignores Gilmore (Ornithischia; Ornithopoda) из верхнего мела Северной Америки». Revue Paléobiologie, Женева . 16 (1): 231–258.
- ^ Ламберт, Дэвид; Группа Диаграмм (1990). «Тецелозавриды» . Книга данных о динозаврах . Нью-Йорк: Книги Эйвона. п. 153 . ISBN 978-0-380-75896-8 .
- ^ Лессем, Дональд ; Перенасыщение, Дональд Ф. (1993). Энциклопедия динозавров Общества динозавров . Random House, Inc. с. 475 . ISBN 978-0-679-41770-5 .
- ^ Гальтон, Питер М. (1970). «Поза гадрозавровых динозавров». Журнал палеонтологии . 44 (3): 464–473.
- ^ Jump up to: а б Фишер, Пол Э.; Рассел, Дейл А.; Стоскопф, Майкл К.; Баррик, Риз Э.; Хаммер, Майкл; Кузьмиц, Эндрю А. (апрель 2000 г.). «Сердечно-сосудистые доказательства средней или более высокой скорости метаболизма у птицетазовых динозавров». Наука . 288 (5465): 503–505. Бибкод : 2000Sci...288..503F . дои : 10.1126/science.288.5465.503 . ПМИД 10775107 .
- ^ Новас, Фернандо Э.; Камбьясо, Андреа В.; Амбриозо, Альфредо (2004). «Новый базальный игуанодонт (Dinosauria, Ornithischia) из верхнего мела Патагонии». Амегиниана . 41 (1): 75–82.
- ^ Бойд, Клинт А.; Клеланд, Тимоти П. (2008). «Морфология и гистология грудных пластин неорнитишевых динозавров» . Аннотация с программами - Геологическое общество Америки; Юго-восточная секция, 57-е ежегодное собрание . 40 (2): 2. Архивировано из оригинала 8 июня 2011 г. Проверено 13 июня 2008 г.
- ^ Jump up to: а б Батлер, Ричард Дж .; Гальтон, Питер М. (2008). «Кожный доспех» динозавра-орнитопода Hypsilophodon из Уилдена (ранний мел: баррем) острова Уайт: переоценка». Меловые исследования . 29 (4): 636–642. Бибкод : 2008CrRes..29..636B . дои : 10.1016/j.cretres.2008.02.002 .
- ^ «Танис: найдены окаменелости динозавра, погибшего в результате удара астероида, утверждают ученые» . Новости Би-би-си . 6 апреля 2022 г.
- ^ Jump up to: а б Вейшампель, Дэвид Б .; Генрих, Рональд Э. (1992). «Систематика Hypsilophodontidae и базальных Iguanodontia (Dinosauria: Ornithopoda)» (PDF) . Историческая биология . 6 (3): 159–184. Бибкод : 1992HBio....6..159W . дои : 10.1080/10292389209380426 . Архивировано из оригинала (PDF) 30 сентября 2007 г. Проверено 10 марта 2007 г.
- ^ Jump up to: а б Хан, Фэн-Лу; Барретт, Пол М.; Батлер, Ричард Дж.; Сюй, Син (2012). «Посткраниальная анатомия Jeholosaurus shangyuanensis (Dinosauria, Ornithischia) из нижнемеловой формации Исянь Китая». Журнал палеонтологии позвоночных . 32 (6): 1370–1395. Бибкод : 2012JVPal..32.1370H . дои : 10.1080/02724634.2012.694385 . S2CID 86754247 .
- ^ Jump up to: а б Вейшампель, Дэвид Б .; Цзяну, Коралия-Мария; Чики, З.; Норман, Дэвид Б. (2003). «Остеология и филогения Залмокса (нг), необычного динозавра-эуорнитопода из позднего мела Румынии». Журнал систематической палеонтологии . 1 (2): 1–56. Бибкод : 2003JSPal...1...65W . дои : 10.1017/S1477201903001032 . S2CID 86339025 .
- ^ Варриккио, Дэвид Дж.; Мартин, Энтони Дж.; Кацура, Ёсихиро (2007). «Первые следы и окаменелости тела роющего динозавра» . Труды Королевского общества B: Биологические науки . 274 (1616): 1361–1368. дои : 10.1098/rspb.2006.0443 . ПМК 2176205 . ПМИД 17374596 .
- ^ Батлер, Ричард Дж.; Апчерч, Пол; Норман, Дэвид Б. (2008). «Филогения птицетазовых динозавров». Журнал систематической палеонтологии . 6 (1): 1–40. Бибкод : 2008JSPal...6....1B . дои : 10.1017/S1477201907002271 . S2CID 86728076 .
- ^ Jump up to: а б с Браун, Калеб Маршалл; Эванс, Дэвид С.; Райан, Майкл Дж.; Рассел, Энтони П. (2013). «Новые данные о разнообразии и численности мелких орнитопод (Dinosauria, Ornithischia) из группы реки Белли (кампан) Альберты». Журнал палеонтологии позвоночных . 33 (3): 495–520. Бибкод : 2013JVPal..33..495B . дои : 10.1080/02724634.2013.746229 . S2CID 129160518 .
- ^ Эриксон, Брюс Р. (2003). Динозавры Музея науки Миннесоты . Сент-Пол, Миннесота: Музей науки Миннесоты. стр. 31–32.
- ^ Нэйлор Хаджинс, Майкл; Карри, Филип Дж.; Салливан, Корвин (2021). «Стоматологическая оценка Stegoceras validum (Ornithischia: Pachycephalosauridae) и Thescelosaurus ignores (Ornithischia: Thescelosauridae): палеоэкологические выводы». Меловые исследования . 130 : 105058. doi : 10.1016/j.cretres.2021.105058 . S2CID 239253658 .
- ^ Баттон, Дэвид Дж.; Занно, Линдси Э. (6 ноября 2023 г.). «Нейроанатомия позднемелового Thescelosaurus ignores (Neornithischia: Thescelosauridae) раскрывает новые экологические специализации внутри динозавров» . Научные отчеты . 13 (1): 19224. Бибкод : 2023NatSR..1319224B . дои : 10.1038/s41598-023-45658-3 . ПМЦ 10628235 . ПМИД 37932280 .
- ^ Пик, Трейси (7 ноября 2023 г.). «Копайте это: у «заброшенного» динозавра были сверхчувства» . Новости штата Северная Каролина .
- ^ Роу, Тимоти; Макбрайд, Эрл Ф.; Серено, Пол К. (февраль 2001 г.). «Технический комментарий: динозавр с каменным сердцем». Наука . 291 (5505): 783а. дои : 10.1126/science.291.5505.783a . ПМИД 11157158 .
- ^ Рассел, Дейл А .; Фишер, Пол Э.; Баррик, Риз Э.; Стоскопф, Майкл К. (февраль 2001 г.). «Ответ: динозавр с каменным сердцем». Наука . 291 (5505): 783а. дои : 10.1126/science.291.5505.783a . ПМИД 11157158 .
- ^ Jump up to: а б Клеланд, Тимоти П.; Стоскопф, Майкл К.; Швейцер, Мэри Х. (2011). «Гистологическое, химическое и морфологическое исследование «сердца» небольшого позднемелового тесцелозавра ». Naturwissenschaften . 98 (3): 203–211. Бибкод : 2011NW.....98..203C . дои : 10.1007/s00114-010-0760-1 . ПМИД 21279321 . S2CID 2408562 .
- ^ Чинсами, Анусуя; Хиллениус, Виллем Дж. (2004). «Физиология нептичьих динозавров». Динозаврия (2-е изд.). стр. 643–659.
- ^ Карпентер, Кеннет ; Брейтаупт, Брент Х. (1986). «Последние меловые появления анкилозавров-нодозавров (Dinosauria, Ornithischia) в западной части Северной Америки и постепенное вымирание динозавров». Журнал палеонтологии позвоночных . 6 (3): 251–257. Бибкод : 1986JVPal...6..251C . дои : 10.1080/02724634.1986.10011619 .
- ^ * Хикс, Дж. Ф., Джонсон, К. Р., Обрадович, Дж. Д., Миггинс, Д. П. и Токс, Л. 2003. Магнитостратиграфия от верхнего мела (маастрихта) до нижнего эоцена в бассейне Денвера, Колорадо. В книге К.Р. Джонсона, Р.Г. Рейнольдса и М.Л. Рейнольдса (редакторы), «Палеонтология и стратиграфия слоев ларамида в бассейне Денвера», Pt. II., Геология Скалистых гор 38: 1-27.
- ^ Сахни, Ашок (1972). «Фауна позвоночных формации Джудит-Ривер, Монтана» (бесплатный PDF-файл, может не загружаться) . Бюллетень Американского музея естественной истории . 147 : 321–412. http://hdl.handle.net/2246/1099 . Проверено 10 марта 2007 г.
- ^ «Тецелозавр» . Триболд Палеонтология, Инк . Проверено 1 июня 2016 г.
- ^ Пирсон, Дин А.; Шефер, Терри; Джонсон, Кирк Р.; Николс, Дуглас Дж.; и Хантер, Джон П. (2002). «Биостратиграфия позвоночных формации Хелл-Крик на юго-западе Северной Дакоты и северо-западе Южной Дакоты». Формация Хелл-Крик и граница мелового и третичного периодов на Северных Великих равнинах: интегрированные континентальные записи конца мелового периода. 145–167.
- ^ Jump up to: а б Лайсон, Тайлер Р.; Лонгрич, Николас Р. (2011). «Пространственное разделение ниш у динозавров позднего мела (маастрихта) Северной Америки» . Труды Королевского общества Б. 278 (1709): 1158–1164. дои : 10.1098/rspb.2010.1444 . ПМК 3049066 . ПМИД 20943689 .
- ^ Карпентер, Кеннет ; Молодой; Д. Брюс (2002). «Динозавры позднего мела из бассейна Денвера, штат Колорадо» (PDF) . Геология Скалистых гор . 37 (2): 237–254. Бибкод : 2002RMGeo..37..237C . дои : 10.2113/11 .
- ^ Рассел, Дейл А. (1989). Одиссея во времени: Динозавры Северной Америки . Миноква, Висконсин: NorthWord Press, Inc., стр. 175–176. ISBN 978-1-55971-038-1 .
- ^ Вейшампель, Дэвид Б.; Барретт, Пол М.; Кориа, Родольфо А.; ЛеЛёфф, Жан; Сюй Син; Чжао Сицзинь; Сахни, Ашок; Гомани, Элизабет, член парламента; и Ното, Кристофер Р. (2004). «Распространение динозавров». Динозаврия (2-я). 517–606.
- ^ Бигелоу, Филипп. «Меловые фаунистические фации Хелл-Крик»; поздний маастрихт» . Архивировано из оригинала 24 января 2007 г. Проверено 26 января 2007 г.
- ^ Дерстлер, Крейг (1994). «Динозавры формации Ланс в восточном Вайоминге». В Нельсоне, Джеральд Э. (ред.). Динозавры Вайоминга . Путеводитель геологической ассоциации Вайоминга, 44-я ежегодная полевая конференция. Геологическая ассоциация Вайоминга. стр. 127–146.
Внешние ссылки
[ редактировать ]- Уилло, динозавр с сердцем - Официальный сайт «Вилло» из Музея естественных наук Северной Каролины.
- Тецелозавриды
- Окаменелости Канады
- Позднемеловые динозавры Северной Америки.
- Ископаемые таксоны описаны в 1913 году.
- Таксоны, названные Чарльзом В. Гилмором
- Ланс фауна
- Фауна Хелл-Крик
- Фауна Сколларда
- Палеонтология в Южной Дакоте
- Палеонтология в Вайоминге
- Палеонтология в Альберте
- Формация Ларами
- Орнитишские роды
- Меловая Южная Дакота
- Меловой Вайоминг
- Меловая Альберта
- Позднемеловые птицетазовые
- Орнитишии Северной Америки
- Многовидовые нептичьи роды динозавров