Поведение роя

Стайное поведение , или роение , — это коллективное поведение, демонстрируемое существами, особенно животными, одинакового размера, которые собираются вместе, возможно, слоняясь в одном и том же месте или, возможно, перемещаясь в массе или мигрируя в каком-то направлении. Это очень междисциплинарная тема. [1]
Термин « роение» применяется, в частности, к насекомым, но его также можно применять к любому другому объекту или животному, проявляющему поведение стаи. Термин «стайное» или «шумение» может относиться конкретно к стаевому поведению птиц, стадному поведению четвероногих , а стайному поведению или стайному поведению рыб. Фитопланктон также собирается в огромные стаи, называемые цветками , хотя эти организмы являются водорослями и не обладают самодвижением, как животные. В более широком смысле термин «рой» применяется также к неодушевленным объектам, которые демонстрируют параллельное поведение, например, рой роботов , рой землетрясений или рой звезд.
С более абстрактной точки зрения, поведение роя — это коллективное движение большого количества самодвижущихся объектов . [2] С точки зрения разработчика математического моделирования, это возникающее поведение, возникающее из простых правил, которым следуют люди, и не требует какой-либо центральной координации. Поведение роя также изучается физиками активной материи как явление, которое не находится в термодинамическом равновесии и как таковое требует разработки инструментов, выходящих за рамки тех, которые доступны в статистической физике систем, находящихся в термодинамическом равновесии. В этом отношении роение сравнивают с математикой сверхтекучести , особенно в контексте стад скворцов (мурмурация). [3]
Поведение роя было впервые смоделировано на компьютере в 1986 году с помощью программы моделирования boids . [4] Эта программа имитирует простых агентов (боидов), которым разрешено передвигаться в соответствии с набором основных правил. Первоначально модель была разработана для имитации стайного поведения птиц, но ее также можно применять к стайным рыбам и другим стайным существам.
Модели [ править ]
В последние десятилетия ученые обратились к моделированию поведения стаи, чтобы глубже понять это поведение.
Математические модели [ править ]
| Внешние изображения | |
|---|---|
В ранних исследованиях поведения стаи использовались математические модели для моделирования и понимания поведения. Простейшие математические модели стад животных обычно представляют отдельных животных согласно трем правилам:
- Двигайтесь в том же направлении, что и их соседи.
- Оставайтесь рядом со своими соседями
- Избегайте столкновений с соседями
Компьютерная программа boids , созданная Крейгом Рейнольдсом в 1986 году, имитирует поведение стаи, следуя вышеуказанным правилам. [4] Многие последующие и нынешние модели используют вариации этих правил, часто реализуя их посредством концентрических «зон» вокруг каждого животного. В «зоне отталкивания», очень близко к животному, очаговое животное будет стремиться дистанцироваться от соседей, чтобы избежать столкновения. Чуть дальше, в «зоне выравнивания», фокусное животное будет стремиться выровнять направление своего движения с соседями. В самой внешней «зоне притяжения», которая простирается настолько далеко от фокусного животного, насколько это возможно, фокусное животное будет стремиться переместиться к соседу.
На форму этих зон обязательно будут влиять сенсорные возможности данного животного. Например, поле зрения птицы не распространяется за ее тело. Рыбы полагаются как на зрение, так и на гидродинамические восприятия, передаваемые через их боковые линии , тогда как антарктический криль полагается как на зрение, так и на гидродинамические сигналы, передаваемые через усики .
Однако недавние исследования стад скворцов показали, что каждая птица меняет свое положение относительно шести или семи животных, непосредственно окружающих ее, независимо от того, насколько близко или насколько далеко эти животные находятся. [5] Таким образом, взаимодействие между стайными скворцами основано на топологическом , а не на метрическом правиле. Еще неизвестно, применимо ли это к другим животным. Другое недавнее исследование, основанное на анализе записей стад над Римом, снятых высокоскоростной камерой, и предполагающее минимальные правила поведения, убедительно смоделировало ряд аспектов поведения стад. [6] [7] [8] [9]
модели Эволюционные
Чтобы понять, почему у животных развивается роевое поведение, ученые обратились к эволюционным моделям, которые моделируют популяции развивающихся животных. Обычно в этих исследованиях используется генетический алгоритм для моделирования эволюции на протяжении многих поколений. В этих исследованиях был изучен ряд гипотез, пытающихся объяснить, почему животные развивают стадное поведение, например, теория эгоистичного стада. [10] [11] [12] [13] [14] эффект замешательства хищника, [15] [16] эффект разбавления, [17] [18] и теория многих глаз. [19]
Агенты [ править ]
- Мах, Роберт; Швейцер, Франк (2003). «Мультиагентная модель биологического роения». Достижения в области искусственной жизни . Конспекты лекций по информатике . Том. 2801. стр. 810–820. CiteSeerX 10.1.1.87.8022 . дои : 10.1007/978-3-540-39432-7_87 . ISBN 978-3-540-20057-4 .
Самоорганизация [ править ]

Появление [ править ]
Концепция эмерджентности – что свойства и функции, обнаруженные на иерархическом уровне, отсутствуют и не имеют значения на более низких уровнях – часто является основным принципом, лежащим в основе самоорганизующихся систем . [20] Примером самоорганизации в биологии , приводящей к появлению в мире природы, являются колонии муравьев. Королева не отдает прямых приказов и не указывает муравьям, что им делать. [ нужна ссылка ] Вместо этого каждый муравей реагирует на раздражители в виде химических запахов личинок, других муравьев, злоумышленников, еды и скоплений отходов и оставляет после себя химический след, который, в свою очередь, стимулирует других муравьев. Здесь каждый муравей представляет собой автономную единицу, которая реагирует только в зависимости от своего местного окружения и генетически закодированных правил его разновидности. Несмотря на отсутствие централизованного принятия решений, колонии муравьев демонстрируют сложное поведение и даже смогли продемонстрировать способность решать геометрические задачи. Например, колонии обычно находят максимальное расстояние от всех входов в колонию, чтобы избавиться от трупов.
Стигмергия [ править ]
Еще одной ключевой концепцией в области роевого интеллекта является стигмергия . [21] [22] Стигмергия — это механизм непрямой координации между агентами или действиями. Принцип заключается в том, что след, оставленный в окружающей среде действием, стимулирует выполнение следующего действия тем же или другим агентом. Таким образом, последующие действия имеют тенденцию усиливать и опираться друг на друга, что приводит к спонтанному возникновению последовательной, очевидно, систематической деятельности. Стигмергия – это форма самоорганизации. Он создает сложные, казалось бы, разумные структуры без необходимости какого-либо планирования, контроля или даже прямого общения между агентами. Таким образом, он поддерживает эффективное сотрудничество между чрезвычайно простыми агентами, которым не хватает памяти, интеллекта или даже знания друг друга. [22]
Роевой интеллект [ править ]
интеллект — это коллективное поведение децентрализованных Роевой , самоорганизующихся систем, естественных или искусственных. Концепция используется в работе над искусственным интеллектом . Это выражение было введено Херардо Бени и Цзин Ваном в 1989 году в контексте клеточных роботизированных систем. [23]
Системы роевого интеллекта обычно состоят из популяции простых агентов, таких как боиды, локально взаимодействующих друг с другом и с окружающей средой. Агенты следуют очень простым правилам, и хотя не существует централизованной структуры управления, диктующей, как должны вести себя отдельные агенты, локальные и в определенной степени случайные взаимодействия между такими агентами приводят к возникновению разумного глобального поведения, неизвестного отдельным агентам.
Исследования роевого интеллекта носят междисциплинарный характер. Его можно разделить на исследования естественного роя, изучающие биологические системы, и исследования искусственного роя, изучающие человеческие артефакты. Существует также научное направление, пытающееся смоделировать сами роевые системы и понять лежащие в их основе механизмы, а также инженерное направление, ориентированное на применение идей, полученных в результате научного направления, для решения практических проблем в других областях. [24]
Алгоритмы [ править ]
Роевые алгоритмы следуют лагранжевому подходу или эйлерову подходу. [25] Эйлеров подход рассматривает рой как поле , работая с плотностью роя и получая средние свойства поля. Это гидродинамический подход, который может быть полезен для моделирования общей динамики крупных роев. [26] [27] [28] Однако большинство моделей работают с лагранжевым подходом, который представляет собой агентную модель, отслеживающую отдельных агентов (точек или частиц), составляющих рой. Модели отдельных частиц могут отслеживать информацию о направлении и расстоянии, которая теряется в эйлеровом подходе. [25] [29]
колонии Оптимизация муравьев
| Внешний образ | |
|---|---|
Оптимизация колонии муравьев — это широко используемый алгоритм, вдохновленный поведением муравьев и эффективно решающий проблемы дискретной оптимизации, связанные с роением. [31] Алгоритм был первоначально предложен Марко Дориго в 1992 году. [32] [33] и с тех пор был диверсифицирован для решения более широкого класса числовых задач. У видов, у которых есть несколько маток, королева может покинуть гнездо вместе с некоторыми рабочими, чтобы основать колонию на новом месте, - процесс, аналогичный роению медоносных пчел . [34] [35]
- Муравьи поведенчески бесхитростны; коллективно они выполняют сложные задачи. Муравьи обладают высокоразвитой сложной системой общения, основанной на знаках.
- Муравьи общаются с помощью феромонов; проложены тропы, по которым могут идти другие муравьи.
- Муравьи, испытывающие проблемы с маршрутизацией, бросают различные феромоны, используемые для расчета «кратчайшего» пути от источника к пункту назначения.
- Раух, Э.М.; Миллонас, ММ; Чиалво, Д.Р. (1995). «Формирование шаблонов и функциональность в роевых моделях». Буквы по физике А. 207 (3–4): 185. arXiv : adap-org/9507003 . Бибкод : 1995PhLA..207..185R . дои : 10.1016/0375-9601(95)00624-c . S2CID 120567147 .
Самодвижущиеся частицы [ править ]
| Внешние видео | |
|---|---|
- нужна Java |
Концепция самодвижущихся частиц (SPP) была введена в 1995 году Тамашем Вичеком и др. [37] как частный случай модели боидов, предложенной Рейнольдсом в 1986 году. [4] Рой SPP моделируется набором частиц, которые движутся с постоянной скоростью и реагируют на случайные возмущения, принимая при каждом приращении среднее направление движения других частиц в их локальной окрестности. [38]
Моделирование показывает, что подходящее «правило ближайшего соседа» в конечном итоге приводит к тому, что все частицы собираются вместе или движутся в одном направлении. Это возникает, несмотря на отсутствие централизованной координации и несмотря на то, что соседи для каждой частицы постоянно меняются с течением времени. [37] Модели SPP предсказывают, что роящиеся животные обладают определенными свойствами на уровне группы, независимо от типа животных в стае. [39] Роевые системы порождают новые модели поведения , которые проявляются в самых разных масштабах, некоторые из которых являются одновременно универсальными и устойчивыми. В теоретической физике стало непросто найти минимальные статистические модели, отражающие такое поведение. [40] [41]
роя Оптимизация частиц
Оптимизация роя частиц — еще один алгоритм, широко используемый для решения проблем, связанных с роями. Он был разработан в 1995 году Кеннеди и Эберхартом и изначально был направлен на моделирование социального поведения и хореографии птичьих стад и рыбных косяков. [42] [43] Алгоритм был упрощен, и было замечено, что он выполняет оптимизацию. Система первоначально засеивает совокупность случайными решениями. Затем он ищет в проблемном пространстве последовательные поколения, используя стохастическую оптимизацию, чтобы найти лучшие решения. Решения, которые он находит, называются частицами . Каждая частица хранит свое положение, а также лучшее решение, которого она достигла на данный момент. Оптимизатор роя частиц отслеживает лучшее локальное значение, полученное на данный момент любой частицей в локальной окрестности. Остальные частицы затем движутся через проблемное пространство, следуя за оптимальными частицами. На каждой итерации оптимизатор роя частиц ускоряет каждую частицу к ее оптимальному местоположению в соответствии с простыми математическими правилами . Оптимизация роя частиц применяется во многих областях. У него мало параметров для настройки, и версия, которая хорошо работает для конкретных приложений, также может хорошо работать с небольшими изменениями в ряде связанных приложений. [44] В книге Кеннеди и Эберхарта описаны некоторые философские аспекты приложений оптимизации роя частиц и роевого интеллекта. [45] Poli провел обширный обзор приложений. [46] [47]
Альтруизм [ править ]
Исследователи из Швейцарии разработали алгоритм, основанный на Гамильтона правиле родственного отбора . Алгоритм показывает, как альтруизм в стае сущностей может со временем развиваться и приводить к более эффективному поведению роя. [48] [49]
роение Биологическое

Самые ранние свидетельства роевого поведения животных датируются примерно 480 миллионами лет назад. окаменелости трилобита Ampyx priscus Недавно было описано, что сгруппированы в ряды вдоль дна океана. Все животные были взрослыми взрослыми особями и смотрели в одном направлении, как если бы они образовали линию конга или пелотон . Было высказано предположение, что они выстраиваются в очередь для миграции, подобно тому, как лангусты мигрируют очередями по одному; [50] также было высказано предположение, что образование является предшественником спаривания, [51] как и с мухой Leptoconops torrens . Результаты показывают, что коллективное поведение животных имеет очень раннее эволюционное происхождение. [52]
Примеры биологического роения встречаются в птичьих стаях . [53] рыбные косяки , [54] [55] стаи насекомых , [56] рои бактерий , [57] [58] формы, [59] молекулярные моторы , [60] четвероногие стада [61] и люди. [62] [63] [64] [65]
Общественные насекомые [ править ]
Поведение общественных насекомых (насекомых, живущих колониями , таких как муравьи, пчелы, осы и термиты) всегда было источником интереса для детей, натуралистов и художников. Отдельные насекомые, кажется, занимаются своими делами без какого-либо централизованного контроля, однако колония в целом ведет себя очень скоординировано. [66] Исследователи обнаружили, что сотрудничество на уровне колонии в значительной степени самоорганизуется . Возникающая групповая координация часто является лишь следствием взаимодействия особей в колонии. Эти взаимодействия могут быть удивительно простыми, например, когда один муравей просто следует по следу, оставленному другим муравьем. Тем не менее, совокупный эффект такого поведения может решить очень сложные проблемы, такие как поиск кратчайшего маршрута в сети возможных путей к источнику пищи. Организованное поведение, возникающее таким образом, иногда называют роевым интеллектом , формой биологического возникновения . [66]
Муравьи [ править ]
Отдельные муравьи не демонстрируют сложного поведения, однако колония муравьев коллективно выполняет сложные задачи, такие как строительство гнезд, забота о потомстве, строительство мостов и поиск пищи. Колония муравьев может коллективно выбирать (т. е. направлять большинство рабочих к) лучший или ближайший источник пищи из нескольких находящихся поблизости. [67] Такие коллективные решения достигаются с использованием механизмов положительной обратной связи. Муравьи выбирают лучший источник пищи, следуя двум простым правилам. Сначала муравьи, нашедшие пищу, возвращаются в гнездо, откладывая химический феромон . Больше феромонов заложено для более качественных источников пищи. [68] Таким образом, если одновременно будут найдены два равноудаленных источника пищи разного качества, след феромонов к лучшему будет сильнее. Муравьи в гнезде следуют еще одному простому правилу: в среднем предпочитают более прочные следы. Затем больше муравьев следуют по более сильному следу, поэтому больше муравьев достигают высококачественного источника пищи, и обеспечивается цикл положительной обратной связи, что приводит к коллективному решению о лучшем источнике пищи. Если есть два пути от муравейника к источнику пищи, то колония обычно выбирает более короткий путь. Это связано с тем, что муравьи, которые первыми возвращаются в гнездо из источника пищи, с большей вероятностью выберут более короткий путь. Затем еще больше муравьев идут по более короткому пути, усиливая след феромонов. [69]
Армейские муравьи , в отличие от большинства видов муравьев, не строят постоянных гнезд; Колония армейских муравьев почти непрерывно перемещается на протяжении всего времени своего существования, оставаясь по сути в постоянном состоянии роения. Несколько линий независимо развили один и тот же базовый поведенческий и экологический синдром, часто называемый «поведением легионеров», и может быть примером конвергентной эволюции . [70]
Успешные методы, используемые колониями муравьев, изучались в области информатики и робототехники для создания распределенных и отказоустойчивых систем для решения проблем. Эта область биомиметики привела к исследованиям передвижения муравьев, поисковым системам, использующим «тропы поиска пищи», отказоустойчивым хранилищам и сетевым алгоритмам . [71]
Медоносные пчелы [ править ]

В умеренном климате медоносные пчелы обычно образуют рой поздней весной. Рой обычно содержит около половины рабочих вместе со старой маткой, а новая королева остается с оставшимися рабочими в исходном улье. Когда медоносные пчелы выходят из улья, чтобы сформировать рой, они могут собраться на ветке дерева или кусте всего в нескольких метрах от улья. Пчелы группируются вокруг королевы и отправляют 20–50 разведчиков на поиски новых подходящих мест для гнезда. Разведчики — самые опытные собиратели в скоплении. Если разведчик находит подходящее место, он возвращается в кластер и продвигает его, танцуя версию танца виляния . Этот танец передает информацию о качестве, направлении и расстоянии до нового участка. Чем больше она воодушевлена своими открытиями, тем энергичнее она танцует. Если ей удастся убедить других, они могут отправиться и проверить найденный ею сайт. Если они одобряют, они также могут продвигать это. В процессе принятия решения скауты проверяют несколько сайтов, часто отказываясь от своего исходного сайта, чтобы продвигать лучший сайт другого скаута. Сначала разные скауты могут продвигать несколько разных сайтов. Через несколько часов, а иногда и дней, в результате процесса принятия решений в конечном итоге выявляется предпочтительное место. Когда все разведчики договорились об окончательном месте, вся группа взлетает и устремляется к нему. Иногда, если решение не принято, рой разделяется, и некоторые пчелы направляются в одном направлении; другие идут в другой. Обычно это приводит к неудаче, когда обе группы погибают. Новое место обычно находится в километре или более от исходного улья, хотя некоторые виды, например, Апис дорсата , [72] могут создавать новые колонии всего в 500 метрах от родного гнезда. Этот процесс коллективного принятия решений чрезвычайно успешен в определении наиболее подходящего нового места для гнезда и сохранении роя в целости и сохранности. Хорошее место для улья должно быть достаточно большим, чтобы вместить рой (около 15 литров в объеме), быть хорошо защищенным от непогоды, получать оптимальное количество солнечного света, находиться на некоторой высоте над землей, иметь небольшой вход и быть способным противостоять заражению муравьями – именно поэтому часто выбирают дупла деревьев. [73] [74] [75] [76] [77]
Несоциальные насекомые [ править ]
В отличие от социальных насекомых, стаи несоциальных насекомых, которые были изучены, по-видимому, в первую очередь функционируют в таких контекстах, как спаривание, питание, избегание хищников и миграция.
Мотыльки [ править ]
Мотыльки могут демонстрировать синхронное спаривание, во время которого феромоны, выделяемые самками, инициируют у самцов поисковое и роевое поведение. [78] Самцы чувствуют феромоны чувствительными усиками и могут отслеживать самок на расстоянии нескольких километров. [79] Спаривание в стае предполагает выбор самки и конкуренцию самцов. Только один самец в рое — обычно первый — успешно совокупляется. [80] Женщины максимизируют преимущества фитнеса и минимизируют затраты, регулируя начало и величину выброса феромонов. Слишком малое количество феромона не привлечет партнера, слишком большое позволяет менее приспособленным самцам почувствовать сигнал. [81] После совокупления самки откладывают яйца на растение-хозяин. Качество растения-хозяина может быть фактором, влияющим на место роения и откладки яиц. В одном случае исследователи наблюдали розово-полосатую бабочку-дубокрылку ( Anisota Virginiensis ), роящуюся на месте падали , где разложение, вероятно, повышало уровень питательных веществ в почве и качество растений-хозяев. [82]
Мухи [ править ]
Мошки, такие как Tokunagayusurika akamusi , образуют стаи, танцующие в воздухе. Роение служит нескольким целям, включая облегчение спаривания путем привлечения самок к рою - явление, известное как спаривание лек . Такие облачные стаи часто образуются ранним вечером, когда солнце садится низко, на кончике куста, на вершине холма, над лужей воды или даже иногда над человеком. Формирование таких стай происходит не инстинктивно, а в результате адаптивного поведения – «консенсуса» – между особями внутри стаи. Также предполагается, что роение является ритуалом , поскольку самец мошки редко бывает сам по себе, а не в рое. Это могло образоваться из-за преимущества снижения инбридинга за счет сбора самцов с различными генами в одном месте. [83] Род Culicoides , также известный как мокрецы, демонстрирует роевое поведение, которое, как полагают, сбивает с толку хищников. [84]
Тараканы [ править ]
Тараканы оставляют химические следы в своих фекалиях, а также выделяют в воздух феромоны для спаривания. Другие тараканы будут следовать по этим тропам, чтобы обнаружить источники еды и воды, а также обнаружить, где прячутся другие тараканы. Таким образом, группы тараканов могут проявлять эмерджентное поведение . [85] при котором поведение группы или стаи возникает из простого набора индивидуальных взаимодействий.
Тараканы в основном ведут ночной образ жизни и убегают при воздействии света. Исследование проверило гипотезу о том, что тараканы используют только две части информации, чтобы решить, куда идти в таких условиях: насколько темно и сколько там других тараканов. Исследование, проведенное Хосе Халлоем и его коллегами из Свободного университета Брюсселя и других европейских учреждений, создало набор крошечных роботов , которые кажутся тараканам другими тараканами и, таким образом, могут изменить восприятие тараканами критической массы . Роботы также были специально ароматизированы, чтобы их приняли настоящие тараканы. [86]
Саранча [ править ]

Саранча — это фаза роения короткорогих кузнечиков семейства Acrididae . Некоторые виды могут быстро размножаться при подходящих условиях и впоследствии стать стадными и мигрирующими. они образуют стаи Будучи нимфами, , а во взрослом состоянии – стаи – и те, и другие могут перемещаться на большие расстояния, быстро опустошая поля и сильно повреждая посевы . Самые большие стаи могут занимать сотни квадратных миль и содержать миллиарды саранчи. Саранча может съедать растения собственного веса (около 2 граммов) каждый день. Это означает, что один миллион саранчи может съедать более одной тонны пищи каждый день, а самые крупные стаи могут потреблять более 100 000 тонн еды каждый день. [87]
Было обнаружено, что роение саранчи связано с повышенным уровнем серотонина , из-за которого саранча меняет цвет, поедает гораздо больше, становится взаимно привлекательным и гораздо легче размножается. Исследователи предполагают, что роевое поведение является реакцией на перенаселенность, а исследования показали, что повышенная тактильная стимуляция задних ног или, у некоторых видов, просто встреча с другими особями вызывает повышение уровня серотонина. Превращение саранчи в роящуюся разновидность может быть вызвано несколькими контактами в минуту в течение четырехчасового периода. [88] [89] [90] [91] Примечательно, что врожденная предрасположенность к агрегации была обнаружена у птенцов пустынной саранчи Schistocerca gregaria , независимо от их родительской фазы. [92]
Реакция отдельной саранчи на потерю выравнивания в группе, по-видимому, увеличивает хаотичность ее движения до тех пор, пока снова не будет достигнуто выравнивание. Такое выравнивание, вызванное шумом, по-видимому, является внутренней характеристикой коллективного когерентного движения. [93]
поведение Миграционное

Миграция насекомых — сезонное перемещение насекомых , особенно видов стрекоз , жуков , бабочек и мотыльков . Расстояние может варьироваться от вида к виду, но в большинстве случаев в этих перемещениях участвуют большие количества особей. В некоторых случаях особи, мигрирующие в одном направлении, могут не вернуться, и вместо этого следующее поколение может мигрировать в противоположном направлении. В этом существенное отличие от миграции птиц .
Бабочки-монархи особенно известны своей длительной ежегодной миграцией. В Северной Америке они совершают массовые миграции на юг, начиная с августа и до первых заморозков. Весной происходит миграция на север. Монарх — единственная бабочка, которая регулярно мигрирует как на север, так и на юг, как это делают птицы. Но ни один человек не совершает весь путь туда и обратно. Во время этих миграций самки-монархи откладывают яйца для следующего поколения. [94] Продолжительность этих путешествий превышает нормальную продолжительность жизни большинства монархов, которая для бабочек, родившихся в начале лета, составляет менее двух месяцев. Последнее поколение летних вступает в нерепродуктивную фазу, известную как диапауза , и может жить семь месяцев и более. [95] Во время диапаузы бабочки перелетают в одно из многочисленных мест зимовки. Зимующее поколение обычно не размножается до тех пор, пока не покинет место зимовки где-то в феврале и марте. Весной на свои северные территории в США и Канаде возвращаются второе, третье и четвертое поколения. Как этому виду удается вернуться в одни и те же места зимовки через несколько поколений, все еще остается предметом исследований; схемы полета, по-видимому, унаследованы в зависимости от комбинации положения солнца на небе [96] и солнечный компас с компенсацией по времени, который зависит от циркадных часов , расположенных в их антеннах. [97] [98]
Птицы [ править ]

 Жужжание скворцов
Жужжание скворцов Миграция птиц [ править ]
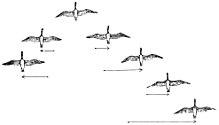
Примерно 1800 из 10 000 видов птиц в мире являются мигрирующими на большие расстояния. [100] Основной мотивацией миграции, по-видимому, является еда; например, некоторые колибри предпочитают не мигрировать, если их кормят зимой. Кроме того, более длинные дни северного лета дают размножающимся птицам больше времени, чтобы выкормить птенцов. Это помогает дневным птицам производить более крупные кладки , чем родственные немигрирующие виды, которые остаются в тропиках. Осенью дни становятся короче, и птицы возвращаются в более теплые регионы, где доступный запас пищи мало меняется в зависимости от сезона. Эти преимущества компенсируют высокий стресс, затраты на физические нагрузки и другие риски миграции, такие как хищничество.
Многие птицы мигрируют стаями. Предполагается, что для более крупных птиц полет стаями снижает затраты энергии. Часто предполагается, что V-образное формирование повышает эффективность и дальность полета птиц, особенно на длинных миграционных маршрутах. Все птицы, кроме первой, летят вверх от одного из вихрей на кончиках крыльев птицы впереди. Восходящий поток помогает каждой птице поддерживать собственный вес в полете, точно так же, как планер может бесконечно подниматься или сохранять высоту в восходящем воздухе. Гуси, летящие V-образным строем, экономят энергию, летая в восходящем потоке вихря на кончиках крыльев, создаваемого предыдущим животным в строю. Таким образом, птицам, летящим сзади, не нужно прилагать столько усилий, чтобы добиться подъемной силы. Исследования показывают, что птицы в V-образном строю располагаются примерно на оптимальном расстоянии, предсказанном простой аэродинамической теорией. [101] Гуси в V-образном строю могут сэкономить 12–20% энергии, необходимой им для полета в одиночку. [102] [103] Радиолокационные исследования показали, что красные сучки и чернозобики летят стаями на 5 км в час быстрее, чем когда они летят поодиночке. [104] Птицы, летящие на концах и спереди, своевременно чередуются, чтобы равномерно распределить усталость от полета среди членов стаи. Такая формация также облегчает общение и позволяет птицам поддерживать визуальный контакт друг с другом.
| Внешние видео | |
|---|---|
- из «Испытаний жизни» |
Другие животные могут использовать аналогичные методы рисования при миграции. Омары , например, мигрируют тесными гуськами «поездами омаров», иногда на сотни миль.
Средиземное и другие моря представляют собой серьезное препятствие для парящих птиц, которым приходится пересекать их в самых узких местах. Огромное количество крупных хищников проходит через такие районы, как Гибралтар , Фальстербо и Босфор и аистов во время миграции . Более распространенные виды, такие как европейский осоед , осенью исчисляются сотнями тысяч. Другие барьеры, такие как горные хребты, также могут вызывать перемещение, особенно крупных дневных мигрантов. Это заметный фактор, препятствующий миграционному процессу в Центральной Америке . Такая концентрация птиц во время миграции может поставить виды под угрозу. Некоторые впечатляющие мигранты уже вымерли, наиболее заметным из которых является странствующий голубь . Во время миграции стаи имели ширину в милю (1,6 км) и длину 300 миль (500 км), их прохождение занимало несколько дней и содержало до миллиарда птиц.
Морская жизнь [ править ]
Рыба [ править ]

| Внешний образ | |
|---|---|
Термин «косяк» можно использовать для описания любой группы рыб, включая группы смешанных видов, тогда как «стайка» используется для более сплоченных групп одного и того же вида, плавающих высоко синхронизированно и поляризованно.
Рыбы получают множество преимуществ от мелководного поведения, включая защиту от хищников (за счет лучшего обнаружения хищников и снижения шансов на поимку), более высокий успех в поисках пищи и более высокий успех в поиске партнера. [106] Также вероятно, что рыба выигрывает от членства в мелководье за счет повышения гидродинамической эффективности. [107]
Рыбы используют множество признаков для выбора товарищей по мелководью. Обычно они предпочитают более крупные косяки, косяков своего вида, косяков, похожих на них по размеру и внешнему виду, здоровую рыбу и родственников (если их узнают). «Эффект странности» утверждает, что любой член косяка, выделяющийся внешним видом, будет предпочтительно атакован хищниками. Это может объяснить, почему рыбы предпочитают скапливаться с особями, похожими на них. Таким образом, эффект странности будет иметь тенденцию к гомогенизации отмелей. [108]
Одним из загадочных аспектов отбора косяков является то, как рыба может присоединиться к косяку животных, похожих на нее, учитывая, что она не может знать свой собственный внешний вид. Эксперименты с рыбками данио показали, что предпочтение мелководья — это приобретенная способность, а не врожденная. Рыбка данио имеет тенденцию общаться с косяками, напоминающими косяки, в которых она была выращена, - это форма импринтинга . [109]
Другие открытые вопросы о поведении косяков включают определение того, какие особи несут ответственность за направление движения косяков. В случае миграционных перемещений большинство членов косяка, по-видимому, знают, куда они направляются. Что касается кормового поведения, то косяки золотого сияния (разновидность гольяна ) возглавляются небольшим количеством опытных особей, которые знали, когда и где можно найти пищу. [110]
По оценкам Радакова, косяки сельди в Северной Атлантике могут занимать до 4,8 кубических километров (1,2 кубических миль) с плотностью рыбы от 0,5 до 1,0 особей на кубический метр, что в общей сложности составляет несколько миллиардов рыб в одном косяке. [111]
- Партридж Б.Л. (1982) «Структура и функции рыбных косяков» Scientific American , июнь: 114–123.
- Пэрриш Дж.К., Вискидо С.В., Грюнбаум Д. (2002). «Самоорганизованные рыбные косяки: исследование новых свойств» (PDF) . Биол. Бык . 202 (3): 296–305. CiteSeerX 10.1.1.116.1548 . дои : 10.2307/1543482 . JSTOR 1543482 . ПМИД 12087003 . S2CID 377484 . [ постоянная мертвая ссылка ]
Миграция рыб [ править ]
| Внешний образ | |
|---|---|
В период с мая по июль огромное количество сардин нерестится в прохладных водах банки Агульяс , а затем следует по течению холодной воды на север вдоль восточного побережья Южной Африки. Эта великая миграция, называемая бегом сардин , создает впечатляющее безумие кормления вдоль береговой линии, поскольку морские хищники, такие как дельфины, акулы и олуши, нападают на косяки.
Krill[editКриль
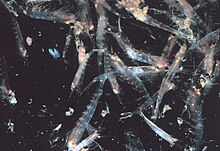
Большинство криля , мелких ракообразных , похожих на креветок , образуют большие стаи, плотность которых иногда достигает 10 000–60 000 особей на кубический метр. [113] [114] [115] Роение — это защитный механизм, сбивающий с толку более мелких хищников, которые хотели бы выбрать отдельных особей. Самые крупные стаи видны из космоса, и их можно отследить с помощью спутника. [116] Было замечено, что один рой покрыл площадь океана в 450 квадратных километров (175 квадратных миль) на глубине 200 метров (650 футов) и, по оценкам, содержал более 2 миллионов тонн криля. [117] Недавние исследования показывают, что криль не просто пассивно дрейфует в этих течениях, а фактически изменяет их. [117] Криль обычно совершает суточную вертикальную миграцию . Двигаясь вертикально через океан с 12-часовым циклом, стаи играют важную роль в смешивании более глубокой, богатой питательными веществами воды с бедной питательными веществами водой на поверхности. [117] До недавнего времени предполагалось, что они проводят день на большей глубине, а ночью поднимаются к поверхности. Было обнаружено, что чем глубже они погружаются, тем больше они снижают свою активность. [118] очевидно, чтобы уменьшить количество встреч с хищниками и сохранить энергию.
Более поздние исследования показали, что плавательная активность криля зависит от наполненности желудка. Сытые животные, питавшиеся у поверхности, плавают менее активно и поэтому опускаются ниже перемешанного слоя. [119] Когда они тонут, они производят фекалии, что может означать, что они играют важную роль в антарктическом углеродном цикле. Было обнаружено, что криль с пустым желудком плавает более активно и поэтому направляется к поверхности. Это означает, что вертикальная миграция может происходить два или три раза в день. Некоторые виды в течение дня образуют стаи на поверхности для кормления и размножения, хотя такое поведение опасно, поскольку делает их чрезвычайно уязвимыми для хищников. [120] Плотные стаи могут спровоцировать безумное кормление рыб, птиц и хищников-млекопитающих, особенно у поверхности. Если их потревожить, рой разбегается, а некоторые особи даже мгновенно линяют , оставляя эксувию в качестве приманки. [121] В 2012 году Гандоми и Алави представили, по всей видимости, успешный стохастический алгоритм для моделирования поведения стаев криля. Алгоритм основан на трех основных факторах: «(i) движение, вызванное присутствием других особей, (ii) деятельность по поиску пищи и (iii) случайное распространение». [122]
Копеподы [ править ]

Веслоногие ракообразные — это группа крошечных ракообразных, обитающих в морях и озерах. Многие виды являются планктонными (дрейфующими в морских водах), а другие — бентосными (живущими на дне океана). Веслоногие рачки обычно имеют длину от 1 до 2 миллиметров (от 0,04 до 0,08 дюйма), с телом каплевидной формы и большими усиками . Хотя, как и у других ракообразных, у них есть бронированный экзоскелет , они настолько малы, что у большинства видов этот тонкий панцирь и все тело почти полностью прозрачны. У веслоногих есть сложный срединный одиночный глаз, обычно ярко-красный, в центре прозрачной головы.
Копеподы также роятся. Например, моноспецифические стаи регулярно наблюдались вокруг коралловых рифов и морской травы , а также в озерах. Плотность стаи составляла около миллиона копепод на кубический метр. Типичные рои имели диаметр один-два метра, но некоторые превышали 30 кубических метров. Копеподам необходим визуальный контакт, чтобы держаться вместе, и ночью они расходятся. [123]
Весна вызывает цветение роящегося фитопланктона , который является пищей для веслоногих ракообразных. Планктонные копеподы обычно являются доминирующими представителями зоопланктона и , в свою очередь, являются основными пищевыми организмами для многих других морских животных. В частности, копеподы являются добычей кормовой рыбы и медуз , которые могут собираться в огромные, миллионные стаи. Некоторые веслоногие ракообразные очень быстро реагируют на побег , когда чувствуют хищника, и могут прыгать с высокой скоростью на несколько миллиметров (см. анимированное изображение ниже).
 На фото: Стая сельдей, питающаяся стаей веслоногих ракообразных.
На фото: Стая сельдей, питающаяся стаей веслоногих ракообразных. Анимация, показывающая, как синхронно охотящиеся сельди могут поймать очень бдительного и уклончивого копепода (нажмите, чтобы просмотреть).
Анимация, показывающая, как синхронно охотящиеся сельди могут поймать очень бдительного и уклончивого копепода (нажмите, чтобы просмотреть). Стаи медуз также охотятся на веслоногих ракообразных.
Стаи медуз также охотятся на веслоногих ракообразных.



Планктонные копеподы играют важную роль в углеродном цикле . Некоторые учёные утверждают, что они образуют самую большую животную биомассу на Земле. [124] За это звание они конкурируют с антарктическим крилем . Однако из-за своих меньших размеров и относительно более высоких темпов роста, а также из-за того, что они более равномерно распределены по большей части мирового океана, копеподы почти наверняка вносят гораздо больший вклад во вторичную продуктивность мировых океанов и в глобальный поглотитель углерода океана, чем криля и, возможно, больше, чем все другие группы организмов вместе взятые. В настоящее время считается, что поверхностные слои океанов являются крупнейшим в мире поглотителем углерода, поглощая около 2 миллиардов тонн углерода в год, что эквивалентно, возможно, трети выбросов углерода человеком , тем самым снижая их воздействие. Многие планктонные копеподы питаются ночью у поверхности, а днем погружаются в более глубокую воду, чтобы избежать визуальных хищников. Их линяющие экзоскелеты, фекальные шарики и дыхание на глубине приносят углерод в глубокое море.
Цветение водорослей [ править ]
Многие одноклеточные организмы, называемые фитопланктоном, обитают в океанах и озерах. При наличии определенных условий, таких как высокий уровень питательных веществ или света, эти организмы размножаются взрывным образом. Образующийся в результате плотный рой фитопланктона называется цветением водорослей . Цветение может занимать сотни квадратных километров, и его легко увидеть на спутниковых изображениях. Отдельный фитопланктон редко живет более нескольких дней, но цветение может длиться неделями. [125] [126]
Растения [ править ]
Ученые уже сотни лет приписывают растениям роевое поведение. В своей книге 1800 года «Фитология: или Философия сельского хозяйства и садоводства » Эразм Дарвин писал, что рост растений напоминает рои, наблюдаемые в других местах в природе. [127] Хотя он имел в виду более широкие наблюдения за морфологией растений и сосредоточился на поведении как корней, так и побегов, недавние исследования подтвердили это утверждение.
В частности, корни демонстрируют заметное роевое поведение, разрастаясь по шаблонам, которые превышают статистический порог случайной вероятности, и указывают на наличие связи между отдельными верхушками корней. Основная функция корней растений — поглощение питательных веществ из почвы, и именно эта цель определяет поведение стаи. Растения, растущие в непосредственной близости, адаптировали свой рост, чтобы обеспечить оптимальную доступность питательных веществ. Это достигается за счет роста в направлении, которое оптимизирует расстояние между близлежащими корнями, тем самым увеличивая их шансы на использование неиспользованных запасов питательных веществ. Действие такого поведения принимает две формы: максимизация расстояния и отталкивание от соседних верхушек корней. [128] Переходная зона кончика корня в значительной степени отвечает за мониторинг присутствия почвенных гормонов, сигнализируя при необходимости об ответных моделях роста. Реакция станции часто бывает сложной и включает в себя множество входных данных для формирования автономной реакции. Дополнительные факторы, определяющие рост стаи, включают свет и гравитацию, оба из которых также контролируются в переходной зоне верхушки корня. [129] Эти силы действуют, чтобы информировать любое количество растущих «основных» корней, которые демонстрируют свои собственные независимые выбросы ингибирующих химических веществ, чтобы установить соответствующее расстояние, тем самым способствуя модели поведения роя. Горизонтальный рост корней, будь то в ответ на высокое содержание минералов в почве или в результате роста столонов , приводит к разветвленному росту, который также образует свои собственные, независимые корневые рои. [130]
Бактерии [ править ]
Роение также описывает группировки некоторых видов хищных бактерий, таких как миксобактерии . Миксобактерии собираются в «волчьи стаи», активно перемещаясь с помощью процесса, известного как бактериальное скольжение , и держась вместе с помощью межклеточных молекулярных сигналов . [57] [131]
Млекопитающие [ править ]


- Пэрриш Дж. К., Эдельштейн-Кешет Л. (1999). «Сложность, закономерности и эволюционные компромиссы в агрегации животных» (PDF) . Наука . 284 (5411): 99–101. Бибкод : 1999Sci...284...99P . CiteSeerX 10.1.1.560.5229 . дои : 10.1126/science.284.5411.99 . ПМИД 10102827 . Архивировано из оригинала (PDF) 20 июля 2011 года.
Люди [ править ]

| Внешние изображения | |
|---|---|
Группа людей также может проявлять поведение стаи, например пешеходы. [134] или солдаты, кишащие на парапетах [ сомнительно – обсудить ] . В Кёльне, Германия, два биолога из Университета Лидса продемонстрировали стадоподобное поведение у людей. Группа людей демонстрировала модель поведения, похожую на стаю: если пять процентов стаи изменят направление, остальные последуют за ней. Если один человек был обозначен как хищник, а все остальные должны были его избегать, стая вела себя очень похоже на косяк рыб. [135] [136] Понимание того, как люди взаимодействуют в толпе, важно, если управление толпой позволит эффективно избежать жертв на футбольных полях, музыкальных концертах и станциях метро. [137]
Математическое моделирование поведения стай — распространенная технология, которая нашла применение в анимации. Имитация флокирования использовалась во многих фильмах. [138] создавать толпы , которые двигаются реалистично. Тима Бертона «Возвращение Бэтмена» был первым фильмом, в котором для рендеринга использовалась технология роя, реалистично изображающая движения группы летучих мышей с использованием системы boids . «Властелин колец» В трилогии фильма , использовалась подобная технология, известная как Massive во время батальных сцен. Технология Swarm особенно привлекательна, поскольку она дешева, надежна и проста.
Компьютерное моделирование на основе муравьев с использованием только шести правил взаимодействия также использовалось для оценки поведения при посадке самолета. [139] Авиакомпании также используют маршрутизацию на основе муравьев при назначении прибытия самолетов к выходам на посадку в аэропорту. Система авиакомпаний, разработанная Дугласом А. Лоусоном, использует теорию роя, или роевой интеллект — идею о том, что колония муравьев работает лучше, чем один. Каждый пилот действует как муравей, ищущий лучший выход на посадку в аэропорту. «Пилот на своем опыте узнает, что для него лучше, и оказывается, что это лучшее решение для авиакомпании», — объясняет Лоусон. В результате «колония» пилотов всегда выходит к воротам, они могут быстро прилететь и улететь. Программа может даже предупредить пилота о появлении резервного самолета еще до того, как оно произойдет. «Мы можем предвидеть, что это произойдет, поэтому у нас будут свободные ворота», — говорит Лоусон. [140]
Поведение роя возникает также в динамике транспортных потоков , например, в волне движения . Двунаправленное движение можно наблюдать по муравьиным тропам. [141] [142] В последние годы это поведение было исследовано для понимания моделей пешеходов и дорожного движения. [143] [144] Моделирование, основанное на моделях пешеходов, также применялось к толпам, которые бросились в панику из-за паники. [145]
Стадное поведение в маркетинге использовалось для объяснения зависимостей взаимного поведения клиентов. The Economist сообщил о недавней конференции в Риме , посвященной моделированию адаптивного поведения человека. [146] Он поделился механизмами, позволяющими увеличить импульсивные покупки и заставить людей «покупать больше, играя на стадном инстинкте». Основная идея заключается в том, что люди будут покупать больше продуктов, которые считаются популярными, и упоминаются несколько механизмов обратной связи для предоставления потребителям информации о популярности продукта, включая технологию смарт-карт и использование технологии радиочастотной идентификации . предложил модель «толпового движения» Исследователь Технологического института Флориды , которая привлекательна для супермаркетов, поскольку может «увеличить продажи без необходимости предоставлять людям скидки».
- Хелбинг Д., Кельч Дж., Молнар П. (1997). «Моделирование эволюции систем человеческих следов». Природа . 388 (6637): 47–50. arXiv : cond-mat/9805158 . Бибкод : 1997Natur.388...47H . дои : 10.1038/40353 . ПМИД 9214501 . S2CID 4364517 .
- Хелбинг Д., Фаркас И., Вичек Т. (2000). «Моделирование динамических особенностей побега паники». Природа . 407 (6803): 487–490. arXiv : cond-mat/0009448 . Бибкод : 2000Natur.407..487H . дои : 10.1038/35035023 . ПМИД 11028994 . S2CID 310346 .
- Хелбинг Д., Фаркас И.Дж., Вичек Т. (2000). «Замораживание при нагреве в приводной мезоскопической системе». Письма о физических отзывах . 84 (6): 1240–1243. arXiv : cond-mat/9904326 . Бибкод : 2000PhRvL..84.1240H . дои : 10.1103/PhysRevLett.84.1240 . ПМИД 11017488 . S2CID 18649078 .
Робототехника [ править ]

Применение принципов роя к роботам называется роевой робототехникой , а роевой интеллект относится к более общему набору алгоритмов.
| Внешние видео | |
|---|---|
Частично вдохновленные колониями насекомых, таких как муравьи и пчелы, исследователи моделируют поведение стаи тысяч крошечных роботов, которые вместе выполняют полезную задачу, например, находят что-то спрятанное, убирают или шпионят. Каждый робот довольно прост, но возникающее поведение роя более сложное. [1] Всю совокупность роботов можно рассматривать как одну распределенную систему, точно так же, как колонию муравьев можно считать суперорганизмом , обладающим роевым интеллектом . Самым большим роем, созданным на данный момент, является рой роботов-килоботов из 1024 человек. [148] Другие крупные стаи включают рой iRobot , проект SRI International /ActivMedia Robotics Centibots , [149] и рой проекта микророботов с открытым исходным кодом, который используется для исследования коллективного поведения. [150] [151] Рои также более устойчивы к сбоям. В то время как один большой робот может потерпеть неудачу и разрушить миссию, рой может продолжить работу, даже если несколько роботов потерпят неудачу. Это может сделать их привлекательными для миссий по исследованию космоса, где неудача обычно обходится чрезвычайно дорого. [152] Помимо наземной техники, роевая робототехника включает в себя также исследование стаев воздушных роботов. [147] [153] и разнородные группы наземных и воздушных транспортных средств. [154] [155]
В отличие от макроскопических роботов, коллоидные частицы на микромасштабе также могут быть использованы в качестве агентов для коллективного поведения при выполнении задач с использованием механических и физических подходов, таких как реконфигурируемый микророй, похожий на торнадо. [156] имитируя стайную рыбу, [157] иерархические виды частиц [158] имитация хищнического поведения млекопитающих, манипулирование микрообъектами с помощью трансформируемого микророя. [159] Изготовление таких коллоидных частиц обычно основано на химическом синтезе.
Военный [ править ]

Военное роение — это поведение, при котором автономные или частично автономные боевые единицы атакуют врага с нескольких разных направлений, а затем перегруппировываются. Пульсация , при которой отряды смещают точку атаки, также является частью военного роения. Военное роение предполагает использование децентрализованных сил против противника таким образом, чтобы упор делался на мобильность, связь, автономию подразделений, а также координацию или синхронизацию. [160] Исторически вооруженные силы использовали принципы роения, не исследуя их подробно, но сейчас активные исследования сознательно изучают военные доктрины, которые черпают идеи из роения.
Просто потому, что несколько юнитов сходятся к цели, они не обязательно сбиваются в кучу. Осадные операции не включают в себя роение, поскольку нет маневра; схождение есть но на осажденном укреплении. также не Засады партизан представляют собой стаи, поскольку они представляют собой «наезды и бегства». Несмотря на то, что засада может иметь несколько точек атаки на противника, партизаны отступают, когда они либо нанесли достаточный урон, либо когда им угрожает опасность.
США В 2014 году Управление военно-морских исследований опубликовало видео, демонстрирующее испытания группы небольших автономных ударных катеров-дронов, которые могут управлять и предпринимать скоординированные наступательные действия в группе. [161]
Галерея [ править ]
 Рой мигрирующих сельдей
Рой мигрирующих сельдей Рой пчел
Рой пчел![Сальпы, выстроенные цепочками, образуют огромные стаи.[162]](//upload.wikimedia.org/wikipedia/commons/thumb/6/63/Salp.jpg/120px-Salp.jpg)
![Люди, толпящиеся через выход, не всегда ведут себя как жидкость.[163][164]](//upload.wikimedia.org/wikipedia/commons/thumb/5/54/Crowd_04378.JPG/120px-Crowd_04378.JPG)
 Рой божьих коровок
Рой божьих коровок Рой роботов
Рой роботов Рой землетрясений
Рой землетрясений Рой древних звезд
Рой древних звезд


![Сальпы, выстроенные цепочками, образуют огромные стаи.[162]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/63/Salp.jpg/120px-Salp.jpg)
![Люди, толпящиеся через выход, не всегда ведут себя как жидкость.[163][164]](http://upload.wikimedia.org/wikipedia/commons/thumb/5/54/Crowd_04378.JPG/120px-Crowd_04378.JPG)




Мифы [ править ]
- Существует популярный миф о том, что лемминги совершают массовое самоубийство , сбегая со скал во время миграции. Движимые сильными биологическими побуждениями, некоторые виды леммингов могут мигрировать большими группами, когда плотность популяции становится слишком большой. Лемминги умеют плавать и могут пересечь водоем в поисках новой среды обитания. В таких случаях многие могут утонуть, если водоем настолько широк, что их физические возможности исчерпаны. Этот факт в сочетании с некоторыми необъяснимыми колебаниями численности норвежских леммингов породил миф. [165]
- Пираньи имеют репутацию бесстрашных рыб, которые сбиваются в свирепые и хищные стаи. Однако недавнее исследование, которое началось «с предпосылки, что они стайиваются как средство совместной охоты», обнаружило, что на самом деле они были довольно пугливыми рыбами, как и другие рыбы, которые обучались для защиты от своих хищников, таких как бакланы, кайманы и дельфины. Исследователь описал их как «по сути обычных рыб с большими зубами». [166]
См. также [ править ]
- Активная материя - поведение материи в масштабе системы.
- Рой Дайсона – гипотетическая мегаструктура вокруг звезды.
- Список собирательных существительных в английском языке
- Юбилей Мобил-Бэй - природное явление, происходящее в Мобил-Бэй, штат Алабама, США.
- Популяционный цикл
- Swarm (симуляция) – набор инструментов для моделирования на основе агентов с открытым исходным кодом.
- Свирлонное состояние - недавно (2020 г.) обнаружено новое состояние самодвижущихся частиц.
- Волна трафика - Тип загруженности шоссе.
- Свармалаторы — агенты, которые одновременно выполняют роение и синхронизацию .
Ссылки [ править ]
- ↑ Перейти обратно: Перейти обратно: а б Буффанае, Роланд (2016). Проектирование и управление динамикой роя . SpringerBriefs in Complexity (первое изд.). Спрингер. дои : 10.1007/978-981-287-751-2 . ISBN 978-981-287-750-5 .
- ^ О'Лоан; Эванс (1998). «Переменное устойчивое состояние при одномерном флокинге». Журнал физики A: Математический и общий . 32 (8): Л99–Л105. arXiv : cond-mat/9811336 . Бибкод : 1999JPhA...32L..99O . дои : 10.1088/0305-4470/32/8/002 . S2CID 7642063 .
- ^ Аттанаси, А.; Каванья, А.; Дель Кастелло, Л.; Джардина, И.; Григера, Т.С.; Джелич, А.; Мелилло, С.; Паризи, Л.; Пол, О.; Шен, Э.; Виале, М. (сентябрь 2014 г.). «Передача информации и поведенческая инерция в стаях скворцов» . Физика природы . 10 (9): 691–696. arXiv : 1303.7097 . Бибкод : 2014NatPh..10..691A . дои : 10.1038/nphys3035 . ПМЦ 4173114 . ПМИД 25264452 .
- ↑ Перейти обратно: Перейти обратно: а б с Рейнольдс CW (1987). «Стада, стада и школы: распределенная поведенческая модель». Материалы 14-й ежегодной конференции «Компьютерная графика и интерактивные технологии» . Том. 21. С. 25–34. CiteSeerX 10.1.1.103.7187 . дои : 10.1145/37401.37406 . ISBN 978-0-89791-227-3 . S2CID 546350 .
{{cite book}}:|journal=игнорируется ( помогите ) - ↑ Перейти обратно: Перейти обратно: а б Баллерини М, Кабиббо Н, Кандельер Р, Каванья А, Сисбани Е, Джардина И, Лекомт В, Орланди А, Паризи Г, Прокаччини А, Виале М, Здравкович В (2008). «Взаимодействие, управляющее коллективным поведением животных, зависит от топологического, а не метрического расстояния: данные полевого исследования» . Учеб. Натл. акад. наук. США . 105 (4): 1232–7. arXiv : 0709.1916 . Бибкод : 2008PNAS..105.1232B . дои : 10.1073/pnas.0711437105 . ПМК 2234121 . ПМИД 18227508 .
- ^ Хильденбрандт Х., Карере С., Хемельрейк К.К. (2010). «Самоорганизованные воздушные демонстрации тысяч скворцов: модель» . Поведенческая экология . 21 (6): 1349–1359. arXiv : 0908.2677 . дои : 10.1093/beheco/arq149 .
- ^ Хемельрейк К.К., Хильденбрандт Х (2011). «Некоторые причины изменчивой формы стай птиц» . ПЛОС ОДИН . 6 (8): e22479. Бибкод : 2011PLoSO...622479H . дои : 10.1371/journal.pone.0022479 . ПМК 3150374 . ПМИД 21829627 .
- ^ «Стаи и школы - Роение - Постоянная экспозиция - Посетите наши выставки и семинары! - Наука LinX - Университет Гронингена» . 10 ноября 2007 г.
- ^ «Исследования на факультете математики и естественных наук - Факультет математики и естественных наук - О нас - Университет Гронингена» . 25 октября 2012 г.
- ^ Ян, В.; Шмикл, Т. (2019). «Коллективное движение как конечный эффект в переполненных эгоистичных стадах» . Научные отчеты . 9 (1): 6618. Бибкод : 2019NatSR...9.6618Y . дои : 10.1038/s41598-019-43179-6 . ПМЦ 6488663 . ПМИД 31036873 .
- ^ Олсон Р.С., Кнестер Д.Б., Адами С. (2013). «Критическое взаимодействие между хищничеством, зависящим от плотности, и эволюцией эгоистичного стада» . Материалы 15-й ежегодной конференции по генетическим и эволюционным вычислениям . Гекко '13. стр. 247–254. дои : 10.1145/2463372.2463394 . ISBN 9781450319638 . S2CID 14414033 .
- ^ Уорд Ч.Р., Гобет Ф., Кендалл Дж. (2001). «Эволюционирование коллективного поведения в искусственной экологии» . Искусственная жизнь . 7 (2): 191–209. CiteSeerX 10.1.1.108.3956 . дои : 10.1162/106454601753139005 . ПМИД 11580880 . S2CID 12133884 .
- ^ Релуга ТК, Вискидо С (2005). «Имитация эволюции эгоистичного стадного поведения». Журнал теоретической биологии . 234 (2): 213–225. Бибкод : 2005JThBi.234..213R . дои : 10.1016/j.jtbi.2004.11.035 . ПМИД 15757680 .
- ^ Вуд А.Дж., Экленд Г.Дж. (2007). «Эволюция эгоистичного стада: появление различных стратегий агрегирования в индивидуальной модели» . Учебник по биологическим наукам . 274 (1618): 1637–1642. дои : 10.1098/rspb.2007.0306 . ПМК 2169279 . ПМИД 17472913 .
- ^ Олсон Р.С., Хинтце А., Дайер ФК, Кнестер Д.Б., Адами С. (2013). «Замешательства хищников достаточно, чтобы развить роевое поведение» . JR Soc. Интерфейс . 10 (85): 20130305. doi : 10.1098/rsif.2013.0305 . ПМК 4043163 . ПМИД 23740485 .
- ^ Демсар Дж., Хемельрейк К.К., Хильденбрандт Х., Баец И.Л. (2015). «Имитация нападения хищников на школы: развитие комплексной тактики» (PDF) . Экологическое моделирование . 304 : 22–33. doi : 10.1016/j.ecolmodel.2015.02.018 . hdl : 11370/0bfcbb69-a101-4ec1-833a-df301e49d8ef . S2CID 46988508 .
- ^ Тош ЧР (2011). «Какие условия способствуют отбору, зависящему от отрицательной плотности скоплений добычи?» (PDF) . Журнал теоретической биологии . 281 (1): 24–30. Бибкод : 2011JThBi.281...24T . дои : 10.1016/j.jtbi.2011.04.014 . ПМИД 21540037 .
- ^ Иоанну К.С., Гуттал В., Кузин И.Д. (2012). «Выбор хищных рыб для скоординированного коллективного движения виртуальной добычи» . Наука . 337 (6099): 1212–1215. Бибкод : 2012Sci...337.1212I . дои : 10.1126/science.1218919 . ПМИД 22903520 . S2CID 10203872 .
- ^ Олсон Р.С., Хейли П.Б., Дайер ФК, Адами С. (2015). «Изучение эволюции компромисса между бдительностью и поиском пищи у групповых организмов» . Королевское общество открытой науки . 2 (9): 150135. arXiv : 1408.1906 . Бибкод : 2015RSOS....250135O . дои : 10.1098/rsos.150135 . ПМЦ 4593673 . ПМИД 26473039 .
- ^ «Иерархия жизни» . 14 сентября 2008 г. Архивировано из оригинала 3 июля 2016 г. Проверено 6 октября 2009 г.
- ^ Парунак, Х. против Д. (2003). «Как создать роение» В: Материалы конференции по роению и сетевому командованию, контролю, связи, компьютерам, разведке, наблюдению и разведке (C4ISR), Маклин, Вирджиния, США, 3 января 2003 г.
- ↑ Перейти обратно: Перейти обратно: а б Марш Л.; Оноф К. (2008). «Стигмергическая эпистемология, стигмергическое познание» (PDF) . Исследование когнитивных систем . 9 (1): 136–149. дои : 10.1016/j.cogsys.2007.06.009 . S2CID 23140721 .
- ^ Бени, Г., Ван, Дж. Роевой интеллект в клеточных робототехнических системах, Продолжайте. Расширенный семинар НАТО по роботам и биологическим системам, Тоскана, Италия, 26–30 июня (1989 г.)
- ^ Дориго, М; Бираттари, М (2007). «Ройовой интеллект» . Схоларпедия . 2 (9): 1462. Бибкод : 2007SchpJ ...2.1462D . дои : 10.4249/scholarpedia.1462 .
- ↑ Перейти обратно: Перейти обратно: а б Ли, YX; Люкман, Р; Эдельштейн-Кешет, Л. (2007). «Минимальные механизмы формирования школ самодвижущихся частиц» (PDF) . Физика D: Нелинейные явления . 237 (5): 699–720. Бибкод : 2008PhyD..237..699L . дои : 10.1016/j.physd.2007.10.009 .
- ^ Тонер Дж и Ту Ю (1995) «Дальний порядок в двумерной модели xy: как птицы летают вместе» Physical Revue Letters, 75 (23) (1995), 4326–4329.
- ^ Топаз С, Бертоцци А (2004). «Схемы роения в двумерной кинематической модели биологических групп». SIAM J Appl Math . 65 (1): 152–174. Бибкод : 2004APS..MAR.t9004T . CiteSeerX 10.1.1.88.3071 . дои : 10.1137/S0036139903437424 . S2CID 18468679 .
- ^ Топаз С., Бертоцци А., Льюис М. (2006). «Нелокальная континуальная модель биологической агрегации». Бычья Математика Биол . 68 (7): 1601–1623. arXiv : q-bio/0504001 . дои : 10.1007/s11538-006-9088-6 . ПМИД 16858662 . S2CID 14750061 .
- ^ Каррильо, Дж; Форназье, М; Тоскани, Дж. (2010). «Частичные, кинетические и гидродинамические модели роения» (PDF) . Математическое моделирование коллективного поведения в социально-экономических науках и науках о жизни . Моделирование и симуляция в науке, технике и технологиях. Том. 3. С. 297–336. CiteSeerX 10.1.1.193.5047 . дои : 10.1007/978-0-8176-4946-3_12 . ISBN 978-0-8176-4945-6 .
- ^ «Проект Сварманоид» . [ постоянная мертвая ссылка ]
- ^ Оптимизация колонии муравьев . Проверено 15 декабря 2010 г.
- ^ А. Колорни, М. Дориго и В. Маньеццо, Распределенная оптимизация муравьиных колоний , материалы первой европейской конференции по искусственной жизни, Париж, Elsevier Publishing, 134–142, 1991.
- ^ М. Дориго, Оптимизация, обучение и естественные алгоритмы , докторская диссертация, Миланский политехнический университет, Италия, 1992.
- ^ Хёльдоблер и Уилсон (1990), стр. 143–179.
- ^ ДОРИГО, М.; ДИ КАРО, Г.; ГАМБЕРЕЛЛА, LM (1999). Муравьиные алгоритмы дискретной оптимизации искусственной жизни . МТИ Пресс.
- ^ Модель самоуправляемых частиц. Архивировано 14 октября 2012 г. в интерактивном моделировании Wayback Machine , 2005 г., Университет Колорадо. Проверено 10 апреля 2011 г.
- ↑ Перейти обратно: Перейти обратно: а б Вичек Т. , Чирок А., Бен-Джейкоб Э., Коэн И., Шошет О. (1995). «Новый тип фазового перехода в системе самодвижущихся частиц». Письма о физических отзывах . 75 (6): 1226–1229. arXiv : cond-mat/0611743 . Бибкод : 1995PhRvL..75.1226V . дои : 10.1103/PhysRevLett.75.1226 . ПМИД 10060237 . S2CID 15918052 .
- ^ Чирок А, Вичек Т (2006). «Коллективное поведение взаимодействующих самодвижущихся частиц». Физика А. 281 (1–4): 17–29. arXiv : cond-mat/0611742 . Бибкод : 2000PhyA..281...17C . дои : 10.1016/S0378-4371(00)00013-3 . S2CID 14211016 .
- ^ Буль Дж., Самптер Д.Д.Т., Кузен Д., Хейл Дж.Дж., Деспланд Э., Миллер Э.Р., Симпсон С.Дж. и др. (2006). «От беспорядка к порядку в марширующей саранче» (PDF) . Наука . 312 (5778): 1402–1406. Бибкод : 2006Sci...312.1402B . дои : 10.1126/science.1125142 . ПМИД 16741126 . S2CID 359329 . Архивировано из оригинала (PDF) 29 сентября 2011 года . Проверено 13 апреля 2011 г.
- ^ Тонер Дж., Ту Ю., Рамасвами С. (2005). «Гидродинамика и фазы скоплений» (PDF) . Анналы физики . 318 (1): 170–244. Бибкод : 2005АнФиз.318..170Т . дои : 10.1016/j.aop.2005.04.011 . Архивировано из оригинала (PDF) 18 июля 2011 года . Проверено 13 апреля 2011 г.
- ^ Бертен, Э; Дроз; Грегуар, Дж. (2009). «Гидродинамические уравнения для самодвижущихся частиц: микроскопический вывод и анализ устойчивости». Дж. Физ. А. 42 (44): 445001. arXiv : 0907.4688 . Бибкод : 2009JPhA...42R5001B . дои : 10.1088/1751-8113/42/44/445001 . S2CID 17686543 .
- ^ Кеннеди, Дж.; Эберхарт, Р. (1995). «Оптимизация роя частиц». Материалы Международной конференции IEEE по нейронным сетям . Том. IV. стр. 1942–1948.
- ^ Кеннеди, Дж. (1997). «Рой частиц: социальная адаптация знаний». Материалы Международной конференции IEEE по эволюционным вычислениям . стр. 303–308.
- ^ Hu X Оптимизация роя частиц: Учебное пособие . Проверено 15 декабря 2010 г.
- ^ Кеннеди, Дж.; Эберхарт, Р.К. (2001). Роевой интеллект . Морган Кауфманн. ISBN 978-1-55860-595-4 .
- ^ Поли, Р. (2007). «Анализ публикаций по приложениям оптимизации роя частиц» (PDF) . Технический отчет CSM-469 . Архивировано из оригинала (PDF) 16 июля 2011 года . Проверено 15 декабря 2010 г.
- ^ Поли, Р. (2008). «Анализ публикаций по применению оптимизации роя частиц» (PDF) . Журнал искусственной эволюции и приложений . 2008 : 1–10. дои : 10.1155/2008/685175 .
- ↑ Альтруизм помогает роящимся роботам летать лучше. Архивировано 15 сентября 2012 г. на сайте Wayback Machine Genevalunch.com , 4 мая 2011 г.
- ^ Вайбель, М; Флореано, Д; Келлер, Л. (2011). «Количественная проверка правила Гамильтона для эволюции альтруизма» . ПЛОС Биология . 9 (5): 1000615. doi : 10.1371/journal.pbio.1000615 . ПМК 3086867 . ПМИД 21559320 .
- ^ Хернкинд, W (1969). «Очередное поведение лангустов». Наука . 164 (3886): 1425–1427. Бибкод : 1969Sci...164.1425H . дои : 10.1126/science.164.3886.1425 . ПМИД 5783720 . S2CID 10324354 .
- ↑ Ископаемые «линии конги» раскрывают происхождение стаи животных National Geographic , 17 октября 2019 г.
- ^ Ваннье, Дж; Видаль, М; Маршан, Р; Эль Харири, К.; Курайс, К; Питтет, Б; Эль Альбани, А; Мазурье, А; Мартин, Э (2019). «Коллективное поведение членистоногих-трилобитов возрастом 480 миллионов лет из Марокко» . Научные отчеты . 9 (1): 14941. Бибкод : 2019НатСР...914941В . дои : 10.1038/s41598-019-51012-3 . ПМК 6797724 . ПМИД 31624280 .
- ^ Fear C (1984) Скворец , Издательство Оксфордского университета. ISBN 978-0-19-217705-6 .
- ^ Куропатка Б.Л. (1982). «Структура и функции рыбных косяков» (PDF) . Научный американец . Том. 246, нет. 6. С. 114–123. Бибкод : 1982SciAm.246f.114P . doi : 10.1038/scientificamerican0682-114 . ПМИД 7201674 . Архивировано из оригинала (PDF) 3 июля 2011 года.
- ^ Хаббард С., Бабак П., Сигурдссон С., Магнуссон К. (2004). «Модель формирования рыбных косяков и миграций рыб». Экол. Модель . 174 (4): 359–374. doi : 10.1016/j.ecolmodel.2003.06.006 .
- ^ Раух Э., Миллонас М., Чиалво Д. (1995). «Формирование шаблонов и функциональность в роевых моделях». Буквы по физике А. 207 (3–4): 185–193. arXiv : adap-org/9507003 . Бибкод : 1995PhLA..207..185R . дои : 10.1016/0375-9601(95)00624-C . S2CID 120567147 .
- ↑ Перейти обратно: Перейти обратно: а б Эллисон С., Хьюз С. (1991). «Бактериальное роение: пример прокариотической дифференциации и многоклеточного поведения». Научный прогресс . 75 (298, ч. 3–4): 403–422. ПМИД 1842857 .
- ^ Бен-Джейкоб Э., Коэн И., Шошет О., Чирок А., Вичек Т. (1995). «Кооперативное формирование хиральных паттернов при росте бактериальных колоний». Письма о физических отзывах . 75 (15): 2899–2902. Бибкод : 1995PhRvL..75.2899B . doi : 10.1103/PhysRevLett.75.2899 . ПМИД 10059433 .
- ^ Раппель В.Дж., Никол А., Саркисян А., Левин Х., Лумис В.Ф. (1999). «Самоорганизованное вихревое состояние в двумерной динамике Dictyostelium». Письма о физических отзывах . 83 (6): 1247–1250. arXiv : patt-sol/9811001 . Бибкод : 1999PhRvL..83.1247R . дои : 10.1103/PhysRevLett.83.1247 . S2CID 1590827 .
- ^ Чоудхури, Д. (2006). «Коллективные эффекты во внутриклеточном молекулярном транспорте: координация, сотрудничество и конкуренция». Физика А. 372 (1): 84–95. arXiv : физика/0605053 . Бибкод : 2006PhyA..372...84C . дои : 10.1016/j.physa.2006.05.005 . S2CID 14822256 .
- ^ Пэрриш Дж. К. и Хамнер В. М. (редакторы) (1997) Группы животных в трех измерениях Издательство Кембриджского университета. ISBN 978-0-521-46024-8 .
- ^ Хелбинг Д., Кельч Дж., Молнар П. (1997). «Моделирование эволюции систем человеческих следов». Природа . 388 (6637): 47–50. arXiv : cond-mat/9805158 . Бибкод : 1997Natur.388...47H . дои : 10.1038/40353 . ПМИД 9214501 . S2CID 4364517 .
- ^ Хелбинг Д., Фаркас И., Вичек Т. (2000). «Моделирование динамических особенностей побега паники». Природа . 407 (6803): 487–490. arXiv : cond-mat/0009448 . Бибкод : 2000Natur.407..487H . дои : 10.1038/35035023 . ПМИД 11028994 . S2CID 310346 .
- ^ Хелбинг Д., Фаркас И.Дж., Вичек Т. (2000). «Замораживание при нагреве в приводной мезоскопической системе». Письма о физических отзывах . 84 (6): 1240–1243. arXiv : cond-mat/9904326 . Бибкод : 2000PhRvL..84.1240H . дои : 10.1103/PhysRevLett.84.1240 . ПМИД 11017488 . S2CID 18649078 .
- ^
- Теория роя National Geographic . Тематическая статья, июль 2007 г.
- Бикман М., Сворд Г.А. и Симпсон С.К. (2008) Биологические основы роевого интеллекта . В книге «Ройовый интеллект: введение и применение» , Эдс Блюм С. и Меркл Д. Спрингер, Япония, К.К. ISBN 978-3-540-74088-9
- Пэрриш Дж. К., Эдельштейн-Кешет Л. (1999). «Сложность, закономерности и эволюционные компромиссы в агрегации животных» (PDF) . Наука . 284 (5411): 99–101. Бибкод : 1999Sci...284...99P . CiteSeerX 10.1.1.560.5229 . дои : 10.1126/science.284.5411.99 . ПМИД 10102827 . Архивировано из оригинала (PDF) 20 июля 2011 года.
- ↑ Перейти обратно: Перейти обратно: а б Бонабо Э. и Тераулаз Дж. (2008) «Рой ум». В вашем будущем с роботами . Специальные издания Scientific American.
- ^ Чачкес, Ти Джей; Грютер, К.; Ратниекс, FLW (2015). «Следовые феромоны: интегративный взгляд на их роль в организации колонии» . Ежегодный обзор энтомологии . 60 : 581–599. doi : 10.1146/annurev-ento-010814-020627 . ПМИД 25386724 . S2CID 37972066 .
- ^ Беккерс, Р.; Денебур, JL; Госс, С. (1993). «Модуляция прокладки следов у муравья Lasius niger (Hymenoptera: Formicidae) и его роль в коллективном выборе источника пищи». Журнал поведения насекомых . 6 (6): 751–759. Бибкод : 1993JIBeh...6..751B . CiteSeerX 10.1.1.161.9541 . дои : 10.1007/BF01201674 . S2CID 14670150 .
- ^ Госс, С.; Арон, С.; Денебур, JL; Пастилс, Дж. М. (1989). «Самоорганизованные ярлыки у аргентинского муравья» (PDF) . Naturwissenschaften . 76 (12): 579–581. Бибкод : 1989NW.....76..579G . дои : 10.1007/BF00462870 . S2CID 18506807 .
- ^ Брэди, Шон Г. (2003). «Эволюция синдрома армейского муравья: происхождение и долгосрочный эволюционный застой комплекса поведенческих и репродуктивных адаптаций» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (11): 6575–9. Бибкод : 2003PNAS..100.6575B . дои : 10.1073/pnas.1137809100 . ПМК 164488 . ПМИД 12750466 .
- ^ Дикке Э., Байд А., Клифф Д., Лэйзелл П. (2004). «Муравьиный метод проектирования сетей хранения данных». В AJ Ispeert, М. Мурата, Н. Вакамия (ред.). Труды биологических подходов к передовым информационным технологиям: первый международный семинар, BioADIT 2004 LNCS 3141 . Спрингер. стр. 364–379 . ISBN 9783540233398 .
- ^ Олдройд, БП (1998). «Сродство колоний в скоплениях Apis dorsata Fabricius (Hymenoptera, Apidae)». Общество насекомых . 47 : 94–95. дои : 10.1007/s000400050015 . S2CID 40346464 .
- ^ Милиус, Сьюзен Swarm Savvy. Архивировано 27 сентября 2012 г. в Wayback Machine . Как пчелы, муравьи и другие животные избегают глупых коллективных решений; Новости науки, 9 мая 2009 г.; Том 175 #10 (с. 16)
- ^ Стаи пчел следуют за высокоскоростными пчелами-стрикерами, чтобы найти новое гнездо ; ScienceDaily (24 ноября 2008 г.)
- ^ Сили, Томас Д.; Вишер, П. Кирк (2003). «Выбор дома: как разведчики в рое медоносных пчел воспринимают завершение группового принятия решения» (PDF) . Поведенческая экология и социобиология . 54 (5): 511–520. дои : 10.1007/s00265-003-0664-6 . S2CID 16948152 . Архивировано из оригинала (PDF) 31 января 2009 года . Проверено 14 декабря 2010 г.
- ^ Морс, РА (1963). «Ориентация роя у медоносных пчел». Наука . 141 (3578): 357–358. Бибкод : 1963Sci...141..357M . дои : 10.1126/science.141.3578.357 . ПМИД 17815993 . S2CID 46516976 .
- ^ Сили, Томас (2003). «Достижение консенсуса при выборе места для гнезда в стаях медоносных пчел: прекращение разногласий». Поведенческая экология и социобиология . 53 (6): 417–424. дои : 10.1007/s00265-003-0598-z . S2CID 34798300 .
- ^ Степьен, ТЛ; Змурчок, С.; Хенгениус, Дж.Б.; Каха Ривера, РМ; Д'Орсонья, MR; Линдси, А.Е. (2000). «Спаривание бабочек: моделирование призыва женских феромонов и стратегий навигации самцов для оптимизации репродуктивного успеха» . Прикладные науки . 10 (18): 6543. doi : 10.3390/app10186543 .
- ^ Бадеке, Б.; Хаверкамп, А.; Саксе, ЮАР (2016). «Задача для самца совки? Различение женского полового феромона на фоне летучих веществ растений» . Границы в физиологии . 7 : 143. doi : 10.3389/fphys.2016.00143 . ПМЦ 4843018 . ПМИД 27199761 .
- ^ Гринфилд, доктор медицины (1981). «Половые феромоны моли: эволюционная перспектива». Энтомолог Флориды . 64 (1): 4–17. дои : 10.2307/3494597 . JSTOR 3494597 .
- ^ Умберс, КДЛ; Саймондс, MRE; Кокко, Х. (2015). «Мотематика передачи сигналов женских феромонов: стратегии старения девственниц». Американский натуралист . 185 (3): 417–432. дои : 10.1086/679614 . hdl : 1885/13166 . ПМИД 25674695 . S2CID 13846468 .
- ^ Мейсон, Д.С.; Баруцци, К. (2019). «Любовь в странных местах» . Границы в экологии и окружающей среде . 17 (3): 184. Бибкод : 2019FrEE...17..184M . дои : 10.1002/плата.2027 .
- ^ «Мошки» . MDC Откройте для себя природу . Проверено 19 ноября 2019 г.
- ^ Киркеби, Карстен (30 июня 2018 г.). «Наблюдения за роевым поведением самок и смешанного пола у Culicoides LATREILLE, 1809 (Diptera: Ceratopogonidae)» . Польский журнал энтомологии . 87 (2): 191–197. дои : 10.2478/pjen-2018-0014 .
- ^ Дженнифер Вьегас. «Тараканы принимают групповые решения» . Канал Дискавери . Архивировано из оригинала 4 июля 2008 года . Проверено 10 июня 2006 г.
- ^ Лемоник, Майкл Д. (15 ноября 2007 г.). «Роботы-тараканы делают свое дело» . Время . Архивировано из оригинала 16 ноября 2007 года.
- ^ Саранча Locustidae National Geographic . Проверено 12 декабря 2010 г.
- ^ «Саранча полна серотонина» . 29 января 2009 г. – через BBC.
- ^ Роджерс С.М., Мэтисон Т., Деспланд Э., Доджсон Т., Берроуз М., Симпсон С.Дж. (ноябрь 2003 г.). «Механосенсорно-индуцированная поведенческая агрегация у пустынной саранчи Schistocerca gregaria » . Дж. Эксп. Биол . 206 (Часть 22): 3991–4002. дои : 10.1242/jeb.00648 . ПМИД 14555739 .
- ^ Стивенсон, Пенсильвания (2009). «Ключ от ящика Пандоры». Наука . 323 (5914): 594–5. дои : 10.1126/science.1169280 . ПМИД 19179520 . S2CID 39306643 .
- ↑ Блокирующее химическое вещество «счастье» может предотвратить нашествие саранчи , New Scientist , 29 января 2009 г., по состоянию на 31 января 2009 г.
- ^ Моше Гершон; Амир Аяли (май 2012 г.). «Врожденное фазовое поведение пустынной саранчи Schistocerca gregaria » . Наука о насекомых . 19 (6): 649–656. Бибкод : 2012InsSc..19..649G . дои : 10.1111/j.1744-7917.2012.01518.x . S2CID 83412818 .
- ^ Йейтс, Калифорния; Эрбан, Р; Эскудеро, К; Кузен, ID; Буль, Дж; Кеврекидис, И.Г.; Майни, ПК; Самптер, диджей (2009). «Собственный шум может способствовать слаженности коллективного движения стаи» . Учеб. Натл. акад. наук. США . 106 (14): 5464–9. Бибкод : 2009PNAS..106.5464Y . дои : 10.1073/pnas.0811195106 . ПМК 2667078 . ПМИД 19336580 .
- ^ Пайл, Роберт Майкл, «Полевой справочник Национального общества Одюбона по бабочкам Северной Америки», стр. 712-713, Альфред А. Кнопф, Нью-Йорк, ISBN 0-394-51914-0
- ^ «Монарх, Данай Плексипп» . Архивировано из оригинала 15 декабря 2012 года . Проверено 27 августа 2008 г.
- ^ Гульотта, Гай (2003): Бабочки, управляемые биологическими часами, ученые Солнца проливают свет на паломничество монархов. Архивировано 5 марта 2006 г. в Wayback Machine . Вашингтон Пост , 23 мая 2003 г., стр. A03. Проверено 7 января 2006 г.
- ^ Мерлин С., Гегир Р.Дж., Репперт С.М. (2009). «Усиковые циркадные часы координируют ориентацию по солнечному компасу у мигрирующих бабочек-монархов» . Наука . 325 (5948): 1700–1704. Бибкод : 2009Sci...325.1700M . дои : 10.1126/science.1176221 . ПМЦ 2754321 . ПМИД 19779201 .
- ^ Кириаку КП (2009). «Разоблачающее путешествие». Наука . 325 (5948): 1629–1630. дои : 10.1126/science.1178935 . ПМИД 19779177 . S2CID 206522416 .
- ^ Надь, М; Акос Зс, Биро Д; Вичек, Т (2010). «Иерархическая групповая динамика в голубиных стаях» (PDF) . Природа . 464 (7290): 890–893. arXiv : 1010.5394 . Бибкод : 2010Natur.464..890N . дои : 10.1038/nature08891 . ПМИД 20376149 . S2CID 4430488 . Архивировано из оригинала (PDF) 6 июля 2010 г. Дополнительный PDF-файл.
- ^ Секерджиоглу, Швейцария (2007). «Экология сохранения: территория превосходит мобильность в результате фрагментарного вымирания птиц» . Современная биология . 17 (8): Р283–Р286. дои : 10.1016/j.cub.2007.02.019 . ПМИД 17437705 . S2CID 744140 .
- ^ Уменьшение сопротивления при полете строем. Летающие самолеты в птичьих формированиях могут значительно увеличить дальность полета ; Центр оборонной технической информации; апрель 2002 г.; Проверено 27 февраля 2008 г.
- ^ Хаммель Д.; Бойкенберг М. (1989). «Аэродинамические интерференционные эффекты в групповом полете птиц». Дж. Орнитол . 130 (1): 15–24. дои : 10.1007/BF01647158 . S2CID 823269 .
- ^ Каттс, CJ и JR Speakman (1994). «Энергосбережение при групповом полете розовоногих гусей» (PDF) . Дж. Эксп. Биол . 189 (1): 251–261. дои : 10.1242/jeb.189.1.251 . ПМИД 9317742 .
- ^ Ньютон, И. (2008). Миграционная экология птиц . Эльсельвир. ISBN 978-0-12-517367-4 .
- ^ Питчер и др. 1982.
- ^ Питчер Т.Дж. и Пэриш Дж.К. (1993) «Функции стайного поведения костистых рыб» В: Питчер Т.Дж. (ред.) Поведение костистых рыб . Чепмен и Холл, Нью-Йорк, стр. 363–440.
- ^ Hoare DJ, Krause J, Peuhkuri N и Godin JGJ (2000) Размер тела и стайность рыб в Journal of Fish Biology, 57 (6) 1351-1366.
- ^ Снексер Дж.Л., Рул Н., Бауэр К., МакРоберт С.П. (2010). «Влияние пола и фенотипа на решения об обмелении рыбок данио» (PDF) . Международный журнал сравнительной психологии . 23 : 70–81. дои : 10.46867/IJCP.2010.23.01.04 . Архивировано из оригинала (PDF) 25 июля 2011 года.
- ^ Энгесзер Р.Э., Райан М.Дж., Паричи Д.М. (2004). «Обучение социальным предпочтениям у рыбок данио» (PDF) . Современная биология . 14 (10): 881–884. дои : 10.1016/j.cub.2004.04.042 . ПМИД 15186744 . S2CID 18741014 .
- ^ Рибс, С.Г. (2000). «Может ли меньшинство информированных лидеров определить перемещения косяка рыб в поисках пищи?». Поведение животных . 59 (2): 403–9. дои : 10.1006/anbe.1999.1314 . ПМИД 10675263 . S2CID 4945309 .
- ^ Радаков Д.В. (1973) Обучение экологии рыб. Израильская программа научного перевода, перевод Mill H. Halsted Press, Нью-Йорк. ISBN 978-0-7065-1351-6
- ^ Фотограф: Марк ван Коллер
- ^ Хамнер, В.М.; Хамнер, ПП; Стрэнд, Юго-Запад; Гилмер, Р.В. (1983). «Поведение антарктического криля Euphausia superba : хеморецепция, питание, стайное обучение и линька ». Наука . 220 (4595): 433–5. Бибкод : 1983Sci...220..433H . дои : 10.1126/science.220.4595.433 . ПМИД 17831417 . S2CID 22161686 .
- ^ У. Килс; П. Маршалл (1995). «Криль, как он плавает и питается – новые знания с помощью новых методов (« Антарктический криль – как он плавает и питается – новые знания с помощью новых методов »)». У И. Гемпеля; Г. Хемпель (ред.). биологии полярных океанов Опыт и . результаты Издательство Фишер . стр. 201–210. ISBN 978-3-334-60950-7 .
- ^ Р. Пайпер (2007). Необыкновенные животные: Энциклопедия любопытных и необычных животных . Гринвуд Пресс. ISBN 978-0-313-33922-6 .
- ^ Хоар, Бен (2009). Миграция животных. Лондон: Музей естественной истории. п. 107. ISBN 978-0-565-09243-6 .
- ↑ Перейти обратно: Перейти обратно: а б с Хоар, Бен (2009). Миграция животных. Лондон: Музей естественной истории. п. 107. ISBN 978-0-565-09243-6
- ^ Дж. С. Яффе; доктор медицинских наук Оманн; А. де Робертис (1999). «Сонарные оценки уровней дневной активности Euphausia pacifica в заливе Саанич» (PDF) . Канадский журнал рыболовства и водных наук . 56 (11): 2000–10. doi : 10.1139/cjfas-56-11-2000 . S2CID 228567512 . Архивировано из оригинала (PDF) 20 июля 2011 года.
- ^ Герайнт А. Тарлинг и Магнус Л. Джонсон (2006). «Насыщение вызывает у криля чувство погружения» . Современная биология . 16 (3): 83–4. дои : 10.1016/j.cub.2006.01.044 . ПМИД 16461267 .
- ^ Ховард, Д.: « Криль », стр. 133–140 в книге Karl, HA et al. (ред.): За Золотыми воротами – океанография, геология, биология и проблемы окружающей среды в заливе Фараллон , циркуляр Геологической службы США № 1198, 2001 г. Последний раз доступ к URL-адресам осуществлялся 4 июня 2010 г.
- ^ Д. Ховард. «Криль в Национальном морском заповеднике Корделл-Бэнк» . НОАА . Проверено 15 июня 2005 г.
- ^ Гандоми, АХ; Алави, АХ (2012). «Алгоритм стада криля: новый алгоритм оптимизации, основанный на биотехнологиях». Коммуникации в нелинейной науке и численном моделировании . 17 (12): 4831–4845. Бибкод : 2012CNSNS..17.4831G . дои : 10.1016/j.cnsns.2012.05.010 .
- ^ Хамнер, В.М.; Карлтон, Дж. Х. (1979). «Рой веслоногих ракообразных: характеристики и роль в экосистемах коралловых рифов» . Лимнол. Океаногр . 24 (1): 1–14. Бибкод : 1979LimOc..24....1H . дои : 10.4319/lo.1979.24.1.0001 .
- ^ Йоханнес Дюрбаум и Торстен Кюннеманн (5 ноября 1997 г.). «Биология копепод: Введение» . Ольденбургский университет Карла фон Осецкого . Архивировано из оригинала 26 мая 2010 года . Проверено 8 декабря 2009 г.
- ^ Линдси Р. и Скотт М. (2010) Что такое фитопланктон, Земная обсерватория НАСА .
- ^ Вредное цветение водорослей в Великих озерах. Архивировано 16 июня 2010 г. в Wayback Machine , 2009 г., NOAA , Центр передового опыта в области Великих озер и здоровья человека.
- ^ Дарвин, Эразм (1 января 1800 г.). Фитология: или Философия сельского хозяйства и садоводства. С теорией осушения болот и с усовершенствованной конструкцией бурового плуга . П. Бирн.
- ^ Чишак, Маржена; Компарини, Диего; Маццолаи, Барбара; Балушка, Франтишек; Арекки, Ф. Тито; Вичек, Тамаш; Манкузо, Стефано (17 января 2012 г.). «Роение в корнях растений» . ПЛОС ОДИН . 7 (1): e29759. Бибкод : 2012PLoSO...729759C . дои : 10.1371/journal.pone.0029759 . ISSN 1932-6203 . ПМК 3260168 . ПМИД 22272246 .
- ^ Балушка, Франтишек; Манкузо, Стефано; Фолькманн, Дитер; Барлоу, Питер В. (1 июля 2010 г.). «Переходная зона верхушки корня: связь сигнал-реакция в корне». Тенденции в науке о растениях . 15 (7): 402–408. doi : 10.1016/j.tplants.2010.04.007 . ПМИД 20621671 .
- ^ Дж., Тревавас А. (2014). Поведение и интеллект растений . Издательство Оксфордского университета. ISBN 9780199539543 . OCLC 961862730 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Райхенбах Х (2001). «Миксобактерии – продуценты новых биологически активных веществ» . J Ind Microbiol Biotechnol . 27 (3): 149–56. дои : 10.1038/sj.jim.7000025 . ПМИД 11780785 . S2CID 34964313 .
- ^ Фаркас И., Хелбинг Д., Вичек Т. (2002). «Мексиканские волны в возбудимой среде» (PDF) . Природа . 419 (6903): 131–132. arXiv : cond-mat/0210073 . Бибкод : 2002Natur.419..131F . дои : 10.1038/419131a . ПМИД 12226653 . S2CID 4309609 . Архивировано из оригинала (PDF) 10 июля 2007 года.
- ^ Неда З., Равас Э., Бреше Ю., Вичек Т., Барабаси А.Л. (2002). «Физика ритмических аплодисментов» (PDF) . Физический обзор E . 61 (6): 6987–6992. arXiv : cond-mat/0006423 . Бибкод : 2000PhRvE..61.6987N . дои : 10.1103/physreve.61.6987 . ПМИД 11088392 . S2CID 14135891 . Архивировано из оригинала (PDF) 11 июня 2011 года.
- ^ Хелбинг, Д; Кельч, Дж; Мольнар, П. (1997). «Моделирование эволюции систем человеческих следов». Природа . 388 (6637): 47–50. arXiv : cond-mat/9805158 . Бибкод : 1997Natur.388...47H . дои : 10.1038/40353 . ПМИД 9214501 . S2CID 4364517 .
- ^ " http://psychcentral.com/news/2008/02/15/herd-mentality-explained/1922.html Архивировано 29 ноября 2014 г. в Wayback Machine ". Проверено 31 октября 2008 г.
- ^ «Численная опасность во время хаджа» . Национальный. 6 сентября 2009 г.
- ^ Кузен И.Д., Краузе Дж. (2003). Самоорганизация и коллективное поведение у позвоночных (PDF) . Достижения в изучении поведения. Том. 32. стр. 1–75. дои : 10.1016/S0065-3454(03)01001-5 . ISBN 978-0-12-004532-7 . Архивировано из оригинала (PDF) 13 марта 2012 года . Проверено 14 апреля 2011 г.
- ^ Габбай, JME (2005). Сложность и аэрокосмическая промышленность: понимание возникновения путем связи структуры с производительностью с использованием мультиагентных систем (Диссертация). Манчестер: Докторская диссертация Манчестерского университета. Архивировано из оригинала 19 декабря 2014 года . Проверено 11 июля 2009 г.
- ^ Ливермор Р. (2008) «Мультиагентный системный подход к симуляционному исследованию, сравнивающий эффективность посадки на борт самолета с использованием заранее назначенных мест и стратегий свободного доступа для всех». Архивировано 13 августа 2011 г. в Wayback Machine Открытом университете , Технический отчет №. 2008/25.
- ^ «Самолеты, поезда и муравейники: ученые-компьютерщики моделируют деятельность муравьев, чтобы уменьшить задержки рейсов». Архивировано 24 ноября 2010 г. в Wayback Machine Science Daily , 1 апреля 2008 г.
- ^ Берд, Мартин; Н. Аранвела (февраль 2003 г.). «Частота встреч в лоб и скорость ходьбы собирателей в движении муравьев-листорезов». Общество насекомых . 50 (1): 3–8. дои : 10.1007/s000400300001 . S2CID 23876486 .
- ^ Рибейро, Педро; Андре Фрасао Элен; Жилберто Ксавьер; Карлос Навас; Фернандо Лейте Рибейру (1 апреля 2009 г.). Дорнхаус, Анна (ред.). «Муравьи могут научиться добывать пищу на тропах с односторонним движением» . ПЛОС ОДИН . 4 (4): е5024. Бибкод : 2009PLoSO...4.5024R . дои : 10.1371/journal.pone.0005024 . ПМЦ 2659768 . ПМИД 19337369 .
- ^ Джон, Александр; Андреас Шадшнайдер; Дебашиш Чоудхури; Кацухиро Нишинари (март 2008 г.). «Характеристики муравьиного транспортного потока». Роевой интеллект . 2 (1): 25–41. arXiv : 0903.1434 . дои : 10.1007/s11721-008-0010-8 . S2CID 18350336 .
- ^ Мы уже почти у цели? Автомобилистам есть чему поучиться у муравьев The Economist , 10 июля 2009 г.
- ^ Хелбинг, Дирк; Фаркас, Элайджа; Вичек, Тамаш (2000). «Моделирование динамических особенностей побега паники». Природа . 407 (6803): 487–490. arXiv : cond-mat/0009448 . Бибкод : 2000Natur.407..487H . дои : 10.1038/35035023 . ПМИД 11028994 . S2CID 310346 .
- ^ «Роение на полках: как магазины могут использовать стадный менталитет людей для увеличения продаж?». Экономист . 11 ноября 2006 г. с. 90.
- ↑ Перейти обратно: Перейти обратно: а б Кушлеев, Алекс; Меллингер, Дэниел; Пауэрс, Кейтлин; Кумар, Виджай (2013). «К рою маневренных микроквадроторов». Автономные роботы . 35 (4): 287–300. дои : 10.1007/s10514-013-9349-9 . S2CID 18340816 .
- ^ «Группа исследования самоорганизующихся систем» . Архивировано из оригинала 26 октября 2014 года.
- ^ «Совместный разведывательный проект роботов Centibots 100» . АктивМедиа Робототехника. Архивировано из оригинала 14 июля 2011 года.
- ^ «Проект микророботов с открытым исходным кодом» . Проверено 28 октября 2007 г.
- ^ «Рой» . Корпорация АйРобот. Архивировано из оригинала 27 сентября 2007 года . Проверено 28 октября 2007 г.
- ^ Кнапп, Луиза (21 декабря 2000 г.). «Смотри, в небе: Робофлай» . Проводной . Проверено 25 сентября 2008 г.
- ^ Саска, Мартин; Ян, Вакула; Либор, Преусил (2014). Стаи микролетательных аппаратов стабилизировались при визуальной относительной локализации . Международная конференция IEEE по робототехнике и автоматизации (ICRA).
- ^ Саска, Мартин; Вонасек, Войтех; Крайник, Томас; Преусил, Либор (2014). «Координация и навигация гетерогенных образований MAV–UGV, локализованных с помощью подхода «ястребий глаз», в рамках модельной схемы прогнозирующего управления» (PDF) . Международный журнал исследований робототехники . 33 (10): 1393–1412. дои : 10.1177/0278364914530482 . S2CID 1195374 .
- ^ Саска, Мартин; Вонасек, Войтех; Крайник, Томас; Преусил, Либор (2012). Координация и навигация разнородных групп БПЛА-УГВ, локализованных методом «ястребиного глаза» . Международная конференция IEEE/RSJ по интеллектуальным роботам и системам (IROS).
- ^ Цзи, Фэнтун; Джин, Дондон; Ван, Бен; Чжан, Ли (23 июня 2020 г.). «Световое зависание магнитного микророя в жидкости» . АСУ Нано . 14 (6): 6990–6998. дои : 10.1021/acsnano.0c01464 . ISSN 1936-0851 . ПМИД 32463226 . S2CID 218976382 .
- ^ Хьюз, Робин (22 февраля 2007 г.), Barracuda Tornado , получено 7 февраля 2022 г.
- ^ Моу, Фанчжи; Ли, Сяофэн; Се, Ци; Чжан, Цзяньхуа; Сюн, Канг; Сюй, Лейлей; Гуань, Цзяньго (20 декабря 2019 г.). «Активные микромоторные системы, построенные из пассивных частиц с биомиметическими взаимодействиями хищник-жертва» . АСУ Нано . 14 (1): 406–414. дои : 10.1021/acsnano.9b05996 . ISSN 1936-0851 . ПМИД 31860277 . S2CID 209435036 .
- ^ Ю, Цзянфань; Ван, Бен; Ду, Синчжоу; Ван, Цяньцянь; Чжан, Ли (21 августа 2018 г.). «Сверхрасширяемый лентообразный магнитный микророй» . Природные коммуникации . 9 (1): 3260. Бибкод : 2018NatCo...9.3260Y . дои : 10.1038/s41467-018-05749-6 . ISSN 2041-1723 . ПМК 6104072 . ПМИД 30131487 .
- ^ Эдвардс, Шон Дж. А. (2000). Роение на поле боя: прошлое, настоящее и будущее . Монография Рэнда MR-1100. Корпорация Рэнд. ISBN 978-0-8330-2779-5 .
- ↑ ВМС США могут «наводнить» противников лодками-роботами , CNN , 13 октября 2014 г.
- ^ «Ныряй и открывай: Научная экспедиция 10: Антарктида» . Проверено 3 сентября 2008 г.
- ↑ Моделирование толпы: моделирование поведения толпы людей или стаи животных имеет как легкомысленное, так и важное применение The Economist , 5 марта 2009 г.
- ^ Фишер, Лен (2009) Идеальный рой: наука о сложности в повседневной жизни. Страница 57. Основные книги. ISBN 978-0-465-01884-0
- ^ Вудфорд, Райли. «Миф о самоубийстве леммингов: фальшивое поведение диснеевского фильма» . Архивировано из оригинала 3 января 2010 года.
{{cite web}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Краснобрюхая пиранья действительно желтая New York Times , 24 мая 2005 г.
Источники [ править ]
- Блюм С. и Меркл Д. (2008) Ройовый интеллект: введение и применение Springer. ISBN 978-3-540-74088-9 .
- Камазин С., Денебур Дж.Л., Фрэнкс Н.Р. , Снейд Дж., Тераулаз Г. и Бонабо Э. (2003) Самоорганизация в биологических системах Издательство Принстонского университета. ISBN 978-0-691-11624-2 .
- Фишер Л. (2009) Идеальный рой: наука о сложности в повседневной жизни . Основные книги. ISBN 978-0-465-01884-0 .
- Кеннеди Дж. Ф., Кеннеди Дж., Эберхарт Р. К. и Ши Ю (2001) Роевой интеллект Морган Кауфманн. ISBN 978-1-55860-595-4 .
- Краузе, Дж. (2005) Жизнь в группах Oxford University Press. ISBN 978-0-19-850818-2
- Лим С.П., Джейн Л.С. и Дехури С. (2009) Инновации в роевом интеллекте Springer. ISBN 978-3-642-04224-9 .
- Миллер, Питер (2010) Умный рой: как понимание стад, школ и колоний может помочь нам лучше общаться, принимать решения и добиваться результатов Пингвин, ISBN 978-1-58333-390-7
- Неджа Н. и Мурель ЛдМ (2006) Интеллектуальные системы Swarm Springer. ISBN 978-3-540-33868-0 .
- Самптер, Дэвид Дж. Т. (2010) Коллективное поведение животных, издательство Принстонского университета. ISBN 978-0-691-14843-4 .
- Вичек А., Зафейрис А. (2012). «Коллективное движение». Отчеты по физике . 517 (3–4): 71–140. arXiv : 1010.5017 . Бибкод : 2012ФР...517...71В . дои : 10.1016/j.physrep.2012.03.004 . S2CID 119109873 .
Внешние ссылки [ править ]
- Статья New York Times о расследовании роения
- Из демонстрационного проекта Wolfram – требуется проигрыватель CDF (бесплатно) :
| Биологическое роение |  | |
|---|---|---|
| Миграция животных | ||
| Роевые алгоритмы | ||
| Коллективное движение | ||
| Роевая робототехника | ||
| Связанные темы | ||