Мелководье и школьное обучение


В биологии любая группа рыб , которая держится вместе по социальным причинам, считается стайной , и если группа скоординированно плывет в одном направлении, они собираются в стаю . [1] В обиходе эти термины иногда используются довольно широко. [1] Около четверти видов рыб стайиваются всю жизнь, а около половины часть жизни стайиваются. [2]
Рыбы получают множество преимуществ от мелководного поведения, включая защиту от хищников (за счет лучшего обнаружения хищников и снижения шансов на поимку отдельной особи), повышение успеха в поисках пищи и более высокий успех в поиске партнера. Также вероятно, что рыба выигрывает от членства в мелководье за счет повышения гидродинамической эффективности.
Рыбы используют множество признаков для выбора товарищей по мелководью. Обычно они предпочитают более крупные косяки, товарищей по мелководью своего вида, товарищей по мелководью, похожих на них по размеру и внешнему виду, здоровую рыбу и родственников (если их распознают).
Эффект странности утверждает, что любой член косяка, выделяющийся внешним видом, будет в первую очередь объектом нападения хищников. Это может объяснить, почему рыбы предпочитают скапливаться с особями, похожими на них самих. Таким образом, эффект странности имеет тенденцию к гомогенизации косяков.
Обзор
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( январь 2021 г. ) |

Скопление рыб — общий термин для обозначения любой группы рыб, собравшейся в некоторой местности. Агрегаты рыб могут быть структурированными и неструктурированными. Неструктурированная агрегация может представлять собой группу представителей разных видов и размеров, которые случайным образом собрались возле какого-либо локального ресурса, например, еды или мест гнездования.
Если, кроме того, скопление объединяется интерактивным, социальным образом, их можно назвать мелководьем . [1] [а] Хотя стайные рыбы могут свободно взаимодействовать друг с другом, при этом каждая рыба плавает и добывает пищу в некоторой степени независимо, они, тем не менее, знают о других членах группы, о чем свидетельствует то, как они корректируют поведение, например плавание, чтобы оставаться рядом. к другой рыбе в группе. Мелководные группы могут включать рыб разного размера, а также могут включать подгруппы смешанных видов.
Если косяк становится более организованным, и рыбы синхронизируют свое плавание, так что все они движутся с одинаковой скоростью и в одном направлении, то можно сказать, что рыба стайная . [1] [3] [б] Стайные рыбы обычно принадлежат к одному и тому же виду и возрасту/размеру. Рыбные косяки движутся так, что отдельные члены находятся на точном расстоянии друг от друга. Школы предпринимают сложные маневры, как будто у школ есть собственное мнение. [4]
Тонкости стайного обучения еще далеко не до конца изучены, особенно энергетика плавания и кормления. Было предложено множество гипотез, объясняющих функцию стайного обучения, например, лучшая ориентация, синхронизированная охота, замешательство хищников и снижение риска быть обнаруженным. Школьное обучение также имеет недостатки, такие как накопление выделений в дыхательных путях, а также истощение кислорода и пищи. То, как рыбный массив в косяке, вероятно, дает преимущества в энергосбережении, хотя это спорно. [5]

Рыбы могут быть облигатными или факультативными (факультативными) косяками. [6] Облигатные косяки, такие как тунцы , сельди и анчоусы , проводят все свое время на мелководье или обучении в стаях и начинают волноваться, если их отделяют от группы. Факультативные косяки, такие как атлантическая треска , сайды и некоторые карангиды , сбиваются в мель лишь время от времени, возможно, в репродуктивных целях. [7]
Стайная рыба может превратиться в дисциплинированный и скоординированный косяк, а затем за считанные секунды снова превратиться в аморфный косяк. Такие сдвиги вызваны изменением активности: кормлением, отдыхом, путешествием или избеганием хищников. [4]
Когда стайные рыбы прекращают кормиться, они разбивают ряды и сбиваются в косяки. Мелководья более уязвимы для нападения хищников. Форма, которую принимает косяк или косяк, зависит от типа рыбы и того, что она делает. Перемещающиеся косяки могут образовывать длинные тонкие линии, квадраты, овалы или амебоидные формы. Быстро движущиеся косяки обычно образуют клинообразную форму, а кормящиеся косяки имеют тенденцию становиться круглыми. [4]
Кормовая рыба – это мелкая рыба, на которую охотятся более крупные хищники. Хищники включают других более крупных рыб, морских птиц и морских млекопитающих . Типичными океаническими кормовыми рыбами являются мелкие рыбы -фильтраторы, такие как сельдь , анчоусы и менхаден . Кормовые рыбы компенсируют свои небольшие размеры образованием косяков. Некоторые плавают в синхронизированных решетках с открытым ртом, чтобы эффективно фильтровать корм из планктона . [8] Эти косяки могут стать огромными, перемещаясь вдоль береговой линии и мигрируя через открытые океаны. Мелководья являются концентрированными пищевыми ресурсами для крупных морских хищников.

Эти порой огромные скопления питают пищевую сеть океана . Большинство кормовых рыб являются пелагическими рыбами , что означает, что они формируют свои косяки в открытой воде, а не на дне или вблизи него ( донные рыбы ). Кормовые рыбы недолговечны и остаются практически незамеченными для человека. Хищники пристально следят за косяками, прекрасно осведомлены об их численности и местонахождении и сами совершают миграции , часто в собственных косяках, которые могут простираться на тысячи миль, чтобы связаться с ними или оставаться на связи. [9]
Сельдь – одна из самых эффектных стайных рыб. Они собираются вместе в огромных количествах. Крупнейшие школы часто образуются в ходе миграций путем слияния с более мелкими школами. «Цепочки» косяков длиной в сто километров (60 миль) наблюдались у кефали, мигрирующей в Каспийском море . По оценкам Радакова, косяки сельди в Северной Атлантике могут занимать до 4,8 кубических километров (1,2 кубических миль) с плотностью рыбы от 0,5 до 1,0 рыбы на кубический метр ( 3 ⁄ 8 до 3 ⁄ рыбы на кубический ярд), всего около трех миллиардов рыб в одном косяке. [10] Эти косяки перемещаются вдоль береговой линии и пересекают открытые океаны. В целом школы сельди имеют очень точные механизмы, которые позволяют школе поддерживать относительно постоянную скорость движения. У сельди отличный слух, и их косяки очень быстро реагируют на хищника. Сельди держатся на определенном расстоянии от движущегося аквалангиста или крейсирующего хищника, такого как косатка, образуя вакуоль, похожую на пончик с самолета-корректировщика. [11]
Многие виды крупных хищных рыб также собираются в стаи, в том числе многие далеко мигрирующие рыбы , такие как тунец и некоторые океанские акулы . Китообразные, такие как дельфины, морские свиньи и киты, действуют организованными социальными группами, называемыми стадами .
«Поведение мелководья обычно описывается как компромисс между преимуществами проживания в группах против хищников и издержками усиления конкуренции за пищу». [12] Ланда (1998) утверждает, что совокупные преимущества обмеления, как подробно описано ниже, представляют собой сильные селективные стимулы для рыбы присоединяться к косякам. [13] Пэрриш и др. (2002) аналогичным образом утверждают, что стайное образование является классическим примером появления , когда существуют свойства, которыми обладает стайка, но не отдельная рыба. Эмерджентные свойства дают членам школы эволюционное преимущество, которого не получают лица, не являющиеся ее членами. [14]
Социальное взаимодействие
[ редактировать ]Поддержку социальной и генетической функции скоплений, особенно образованных рыбами, можно увидеть в нескольких аспектах их поведения. Например, эксперименты показали, что у отдельных рыб, удаленных из косяка, частота дыхания будет выше, чем у рыб, обнаруженных в косяке. [15] Этот эффект связывают со стрессом, и поэтому эффект от общения с представителями своего вида оказывается успокаивающим и мощной социальной мотивацией для пребывания в агрегации. [16] Например, сельдь станет очень возбужденной, если ее изолировать от сородичей. [7] Из-за адаптации к стайному поведению их редко показывают в аквариумах . Даже при самых лучших условиях, которые могут предложить аквариумы, они становятся хрупкими и медлительными по сравнению с их трепещущей энергией в диких стаях. [ нужна ссылка ]
Преимущества собирательства
[ редактировать ]
Также было высказано предположение, что плавание в группах повышает успех в поисках пищи. Эта способность была продемонстрирована Питчером и другими в их исследовании пищевого поведения мелководных карповых . [17] В этом исследовании было количественно оценено время, необходимое группам пескарей и золотых рыбок, чтобы найти кусок пищи. Численность рыб в группах варьировала, установлено статистически значимое уменьшение времени, необходимого более крупным группам для поиска пищи. Дополнительную поддержку расширению кормовой способности косяков можно увидеть в структуре косяков хищных рыб. Партридж и другие проанализировали структуру косяка атлантического голубого тунца по аэрофотоснимкам и обнаружили, что косяк принял параболическую форму, что наводит на мысль о совместной охоте этого вида. [18]
«Причиной этого является наличие множества глаз, ищущих пищу. Рыбы в косяках «делятся» информацией, внимательно наблюдая за поведением друг друга. Пищевое поведение одной рыбы быстро стимулирует поиск пищи у других. [19]
Плодородную кормовую среду для кормовых рыб обеспечивают океанские апвеллинги. Океанические круговороты — это крупномасштабные океанские течения, вызванные эффектом Кориолиса . Вызванные ветром поверхностные течения взаимодействуют с этими круговоротами и подводной топографией, такой как подводные горы , рыболовные отмели и края континентальных шельфов , вызывая нисходящие и апвеллинги . [20] Они могут переносить питательные вещества, необходимые для развития планктона. В результате могут появиться богатые кормовые угодья, привлекательные для кормовых рыб, питающихся планктоном. В свою очередь, кормовая рыба сама становится местом кормления более крупных рыб-хищников. Большинство апвеллингов являются прибрежными, и многие из них поддерживают одни из самых продуктивных рыболовных промыслов в мире. Регионы заметного апвеллинга включают прибрежные районы Перу , Чили , Аравийского моря , западную часть Южной Африки , восточную часть Новой Зеландии и побережье Калифорнии . [ нужна ссылка ]
Веслоногие ракообразные , основной зоопланктон , являются основным элементом меню кормовой рыбы. Это группа мелких ракообразных, обитающих в океанических и пресноводных средах обитания . Веслоногие рачки обычно имеют длину от одного миллиметра (0,04 дюйма) до двух миллиметров (0,08 дюйма) и имеют каплевидное тело. Некоторые учёные утверждают, что они образуют самую большую животную биомассу на планете. [21] Копеподы очень бдительны и уклончивы. У них большие усики (см. фото внизу слева). Расправляя усики, они чувствуют волну давления от приближающейся рыбы и прыгают с огромной скоростью на несколько сантиметров. Если концентрация копепод достигает высокого уровня, стайная сельдь применяет метод, называемый кормлением баранов . На фото ниже сельдевой баран питается стаей веслоногих ракообразных . Они плавают с широко открытым ртом и крышками . полностью раскрытыми [ нужна ссылка ]
 У этого копепода антенна расправлена (нажмите, чтобы увеличить). Антенна улавливает волну давления приближающейся рыбы.
У этого копепода антенна расправлена (нажмите, чтобы увеличить). Антенна улавливает волну давления приближающейся рыбы. Веслоногие ракообразные являются основным источником пищи для кормовых рыб, таких как атлантическая сельдь .
Веслоногие ракообразные являются основным источником пищи для кормовых рыб, таких как атлантическая сельдь . Стая баранов сельди, питающихся стаей копепод, с расширенными крышками , так что видны их красные жабры.
Стая баранов сельди, питающихся стаей копепод, с расширенными крышками , так что видны их красные жабры. Анимация, показывающая, как сельдь, охотясь синхронно, может поймать очень бдительного и уклончивого копепода.
Анимация, показывающая, как сельдь, охотясь синхронно, может поймать очень бдительного и уклончивого копепода.




Рыбы плавают по сетке, где расстояние между ними такое же, как длина прыжка их добычи, как показано на анимации выше справа. В анимации молодь сельди охотится на копепод синхронно . Копеподы чувствуют своими усиками волну давления приближающейся сельди и реагируют быстрым прыжком. Длина прыжка довольно постоянна. Рыбы выстраиваются в сетку с этой характерной длиной прыжка. Копепод может прыгнуть около 80 раз, прежде чем устанет. После прыжка ему требуется 60 миллисекунд, чтобы снова расправить усики, и эта временная задержка становится его гибелью, поскольку почти бесконечный поток сельди позволяет сельди в конечном итоге перехватить копепод. Одна молодь сельди никогда не могла поймать крупную копеподу. [8]
Репродуктивные преимущества
[ редактировать ]Третье предполагаемое преимущество групп рыб заключается в том, что они выполняют репродуктивную функцию. Они обеспечивают расширенный доступ к потенциальным партнерам, поскольку поиск партнера на мелководье не требует много энергии. А для мигрирующих рыб, которые перемещаются на большие расстояния в поисках нереста, вполне вероятно, что навигация по косяку при участии всех членов косяка будет лучше, чем у отдельной рыбы. [4] [ нужна страница ]
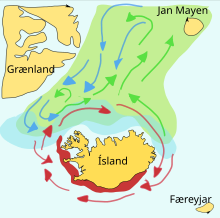
Кормовые рыбы часто совершают большие миграции между местами нереста, нагула и нагула. Школы определенного типа обычно путешествуют в треугольнике между этими территориями. Например, нерестилища одного стада сельди находятся на юге Норвегии , места нагула — в Исландии , а места нагула — в северной Норвегии. Подобные широкие треугольные путешествия могут быть важны, поскольку кормовая рыба во время кормления не может различить собственное потомство. [ нужна ссылка ]
Мойва – кормовая рыба семейства корюшка , обитающая в Атлантическом и Северном Ледовитом океанах. Летом они пасутся на густых стаях планктона на краю шельфового ледника. Более крупная мойва также поедает криль и других ракообразных . Мойва большими стаями перемещается к берегу для нереста и миграции весной и летом, чтобы накормиться в богатых планктоном районах между Исландией , Гренландией и Ян-Майеном . На миграцию влияют океанские течения . В окрестностях Исландии созревающая мойва весной и летом совершает крупные кормовые миграции на север. Обратная миграция происходит в сентябре-ноябре. Нерестовая миграция начинается к северу от Исландии в декабре или январе. [22]
Гидродинамический КПД
[ редактировать ]Эта теория утверждает, что группы рыб могут экономить энергию, плавая вместе, во многом так же, как велосипедисты могут тянуть друг друга в пелотоне . Считается, что гуси, летящие в строю Ви, экономят энергию, летая в восходящем потоке вихря на кончиках крыльев, создаваемого предыдущим животным в строю. [23] [24] Было предложено повысить эффективность группового плавания для косяков рыб и антарктического криля . [ нужна ссылка ]
Казалось бы, разумно предположить, что регулярное расположение и однородность размеров рыб в косяках приведет к гидродинамической эффективности. [12] Хотя ранние лабораторные эксперименты не смогли обнаружить гидродинамические преимущества, создаваемые соседями рыбы в косяке, [19] Считается, что повышение эффективности действительно происходит в дикой природе. Более поздние эксперименты с группами рыб, плавающих в лотках, подтверждают это: рыба снижает затраты на плавание на целых 20% по сравнению с тем, когда та же рыба плавает изолированно. [25] Ланда (1998) утверждал, что лидер стаи постоянно меняется, поскольку, хотя пребывание в корпусе стаи дает гидродинамическое преимущество, лидер первым получает пищу. [13] Более поздние исследования показывают, что после того, как люди, находящиеся в передней части школы, встречают и поглощают больше еды, они затем перемещаются дальше в пределах школы из-за двигательных ограничений, возникающих во время переваривания еды. [26]
Уклонение от хищников
[ редактировать ]

Обычно наблюдается, что стайная рыба подвергается особой опасности быть съеденной, если ее отделить от стаи. [4] несколько функций рыбных косяков по борьбе с хищниками Было предложено .
Одним из потенциальных методов, с помощью которых косяки рыб могут отпугивать хищников, является «эффект замешательства хищников», предложенный и продемонстрированный Милински и Хеллером (1978). [29] Эта теория основана на идее, что хищникам становится трудно выбирать отдельную добычу из групп, поскольку множество движущихся целей создают сенсорную перегрузку зрительного канала хищника. Выводы Милински и Хеллера были подтверждены экспериментами. [30] [31] и компьютерное моделирование. [32] [33] «Стайные рыбы одинакового размера и серебристого цвета, поэтому визуально ориентированному хищнику сложно выделить особь из массы извивающихся, сверкающих рыб и затем успеть схватить добычу до того, как она исчезнет в стае». [4]
Стайное поведение сбивает с толку орган боковой линии (LLO), а также электросенсорную систему (ESS) хищников. [34] [35] [36] Движения плавников отдельной рыбы действуют как точечный источник волн, излучающий градиент, с помощью которого хищники могут его локализовать. Поскольку поля многих рыб перекрываются, стайное обучение должно скрывать этот градиент, возможно, имитируя волны давления более крупного животного, и, скорее всего, сбивать с толку восприятие боковой линии. [34] LLO необходим на заключительных стадиях нападения хищника. [37] Электрорецептивные животные могут локализовать источник поля, используя пространственные неоднородности. Чтобы производить отдельные сигналы, отдельные жертвы должны находиться на расстоянии примерно пяти ширины тела друг от друга. Если объекты расположены слишком близко друг к другу, чтобы их можно было различить, они образуют размытое изображение. [38] На основании этого было высказано предположение, что стайное обучение может сбить с толку ЭСС хищников. [34]
Третий потенциальный эффект скоплений животных против хищников - это гипотеза «многих глаз». Эта теория утверждает, что по мере увеличения размера группы задача сканирования окружающей среды на наличие хищников может быть распределена между многими особями. по-видимому, не только Такое массовое сотрудничество, обеспечивает более высокий уровень бдительности, но и дает больше времени для индивидуального кормления. [39] [40]
Четвертая гипотеза антихищнического эффекта рыбных косяков - это эффект «разбавления при встрече». Эффект разбавления представляет собой уточнение безопасности в цифрах и взаимодействует с эффектом путаницы. [19] При данной атаке хищника большая косяк съедает меньшую часть, чем мелкий косяк. [41] Гамильтон предположил, что животные собираются в группы из-за «эгоистичного» избегания хищника, что, таким образом, является формой поиска укрытия. [42] Другая формулировка теории была дана Тернером и Питчером и рассматривалась как комбинация вероятностей обнаружения и атаки. [43] В компоненте теории обнаружения было высказано предположение, что потенциальная добыча может получить выгоду от совместного проживания, поскольку хищник с меньшей вероятностью наткнется на одну группу, чем на разбросанную группу. Что касается компонента нападения, считалось, что атакующий хищник с меньшей вероятностью съест определенную рыбу, когда присутствует большее количество рыбы. В целом, рыба имеет преимущество, если она находится в большей из двух групп, при условии, что вероятность обнаружения и нападения не увеличивается непропорционально с размером группы. [44]
Стайные кормовые рыбы подвергаются постоянным нападениям хищников. Примером могут служить нападения, произошедшие во время промысла африканских сардин . Миграция африканских сардин – это захватывающая миграция миллионов серебристых сардин вдоль южного побережья Африки. С точки зрения биомассы, популяция сардин может соперничать с огромной миграцией антилоп гну в Восточной Африке . [45] У сардин короткий жизненный цикл: они живут всего два или три года. Взрослые сардины в возрасте около двух лет собираются на берегу Агульяс , где они нерестятся весной и летом, выпуская в воду десятки тысяч икринок. Затем взрослые сардины сотнями косяков направляются к субтропическим водам Индийского океана . Более крупная отмель может иметь длину 7 километров (4,3 мили), ширину 1,5 километра (0,93 мили) и глубину 30 метров (98 футов). Огромное количество акул, дельфинов, тунцов, парусников, капских морских котиков и даже косаток собираются и следуют за косяками, создавая безумное кормление вдоль береговой линии. [46]

При угрозе сардины (и другие кормовые рыбы) инстинктивно группируются вместе и создают массивные комки с наживкой . Шары-приманки могут достигать 20 метров (66 футов) в диаметре. Они кратковременны и редко длятся более 20 минут.Икра рыб, оставленная на отмелях Агульяс, течением дрейфует на северо-запад в воды у западного побережья, где личинки развиваются в молодь рыбы. Когда они становятся достаточно взрослыми, они собираются в плотные косяки и мигрируют на юг, возвращаясь на берега Агульяса, чтобы возобновить цикл. [46]
Развитие стайного поведения, вероятно, было связано с повышением качества восприятия, хищническим образом жизни и механизмами сортировки по размеру во избежание каннибализма. [36] У предков, питавшихся фильтратором, до того, как развилось зрение и октаволатеральная система (OLS), риск нападения хищников был ограничен и в основном из-за беспозвоночных хищников. Следовательно, в то время безопасность численности, вероятно, не была основным стимулом для сбора в косяках или косяках. Развитие зрения и OLS позволило бы обнаружить потенциальную добычу. Это могло привести к увеличению вероятности каннибализма на мелководье. С другой стороны, повышение качества восприятия дало бы мелким особям шанс убежать или никогда не присоединяться к косяку с более крупной рыбой. Показано, что мелкие рыбы избегают присоединения к группе с более крупными рыбами, хотя и крупные рыбы не избегают присоединения к мелким сородичам. [47] Этот механизм сортировки, основанный на повышении качества восприятия, мог бы привести к однородности размеров рыб в косяках, что увеличило бы способность двигаться синхронно. [36]
Противодействие хищникам
[ редактировать ]
Хищники разработали различные контрмеры, чтобы подорвать защитные стайные и стайные маневры кормовой рыбы. Рыба -парусник поднимает парус, чтобы казаться намного больше, чтобы она могла пасти стаю рыб или кальмаров. Рыба-меч на высокой скорости несется сквозь стаи кормовых рыб, рубя мечами, чтобы убить или оглушить добычу. Затем они поворачиваются и возвращаются, чтобы съесть свой «улов». Акулы-молотилки используют свои длинные хвосты, чтобы оглушить стайную рыбу. Прежде чем нанести удар, акулы уплотняют стаи добычи, плавая вокруг нее и расплескивая воду хвостом, часто парами или небольшими группами. Молотилки плывут кругами, сбивая стайную добычу в компактную массу, а затем резко наносят ей удар верхней долей хвоста, чтобы оглушить ее. [48] [49] Акулы-спиннеры проносятся вертикально через косяк, вращаясь вокруг своей оси с открытой пастью и щелкая по сторонам. Инерция акулы в конце этого спирального движения часто поднимает ее в воздух. [50] [51]
 Стадо парусников с парусами.
Стадо парусников с парусами. Рыбы-меч рубят своими мечами.
Рыбы-меч рубят своими мечами. Акулы-молотилки наносят удары хвостами.
Акулы-молотилки наносят удары хвостами. Акулы-спиннеры вращаются вокруг своей длинной оси.
Акулы-спиннеры вращаются вокруг своей длинной оси.



![↑ Команда обыкновенных афалин вместе заставляет стайных рыб прыгать в воздух. В таком уязвимом положении рыба становится легкой добычей дельфинов.[52]](//upload.wikimedia.org/wikipedia/commons/thumb/2/2e/Barrier_feeding_in_bottlenose_dolphins.jpg/559px-Barrier_feeding_in_bottlenose_dolphins.jpg) ↑ Команда обыкновенных афалин вместе заставляет стайных рыб прыгать в воздух. В таком уязвимом положении рыба становится легкой добычей дельфинов. [52]
↑ Команда обыкновенных афалин вместе заставляет стайных рыб прыгать в воздух. В таком уязвимом положении рыба становится легкой добычей дельфинов. [52]
![↑ Команда обыкновенных афалин вместе заставляет стайных рыб прыгать в воздух. В таком уязвимом положении рыба становится легкой добычей дельфинов.[52]](http://upload.wikimedia.org/wikipedia/commons/thumb/2/2e/Barrier_feeding_in_bottlenose_dolphins.jpg/559px-Barrier_feeding_in_bottlenose_dolphins.jpg)
Некоторые хищники, например дельфины, охотятся отдельными группами. Одним из методов, используемых многими видами дельфинов, является стадное содержание , при котором стайка будет контролировать стаю рыб, в то время как отдельные члены по очереди пропахивают и питаются более плотно упакованной стаей (формация, широко известная как шарик с наживкой ). Загон — это метод, при котором рыбу преследуют на мелководье, где ее легче поймать. В Южной Каролине атлантическая афалина делает еще один шаг вперед, применяя так называемое кормление на берегу, когда рыбу загоняют на илистые отмели и извлекают оттуда. [53]
Обыкновенных афалин наблюдали с помощью другого метода. Один дельфин действует как «водитель» и гонит косяк рыб навстречу нескольким другим дельфинам, которые образуют барьер. Дельфин-поводырь хлопает лапой, и рыба подпрыгивает в воздух. Когда рыба прыгает, дельфин-водитель движется вместе с дельфинами-барьерами и ловит рыбу в воздухе. [52] Этот тип совместной ролевой специализации, по-видимому, более распространен у морских животных , чем у наземных животных , возможно, потому, что в океанах больше изменчивости в разнообразии добычи, биомассе и подвижности хищников. [52]
Во время вылова сардин около 18 000 дельфинов, ведущих себя как овчарки, сбивают сардины в шарики с приманкой или загоняют их в загон на мелководье. Как только шарики с наживкой собраны, дельфины и другие хищники по очереди пробираются сквозь них, поглощая рыбу, пока она проносится. Сверху на них нападают также морские птицы: стаи олушей , бакланов , крачек и чаек . Некоторые из этих морских птиц падают с высоты 30 метров (100 футов), ныряя в воду, оставляя за собой следы, похожие на пар, похожие на следы истребителей. [46] Олуши падают в воду со скоростью до 100 километров в час (60 миль в час). Под кожей на лице и груди у них есть воздушные мешки, которые действуют как пузырчатая пленка , смягчая удары водой.
Известно, что некоторые популяции афалин в Мавритании занимаются межвидовым совместным рыболовством с рыбаками-людьми. Дельфины гонят косяк рыб к берегу, где их ждут люди со своими сетями. В путанице забросных сетей дельфины ловят и большое количество рыбы. Также наблюдались внутривидовые методы совместного поиска пищи, и некоторые предполагают, что такое поведение передается культурными средствами. Ренделл и Уайтхед предложили структуру изучения культуры китообразных. [54]
Некоторые киты бросаются питаться шариками с наживкой. [55] Кормление с выпадом — это экстремальный метод кормления, при котором кит ускоряется из-под шарика с приманкой до высокой скорости, а затем открывает пасть под большим углом. Это создает давление воды, необходимое для расширения рта, поглощения и фильтрации огромного количества воды и рыбы. Кормление огромных рорквалов на выпаде считается крупнейшим биомеханическим событием на Земле. [56]
| Внешние видео | |
|---|---|
 Пара горбатых китов, разновидность рорквалов, кормится на выпаде.
Пара горбатых китов, разновидность рорквалов, кормится на выпаде. Олуши «пикирующие бомбы» на большой скорости
Олуши «пикирующие бомбы» на большой скорости


Как рыбная школа
[ редактировать ]
Косяки рыб плавают дисциплинированными фалангами, причем некоторые виды, например сельдь, способны двигаться вверх и вниз с впечатляющей скоростью, извиваясь туда-сюда и внося поразительные изменения в форму косяка без столкновений. Как будто их движения поставлены хореографически, хотя это не так. Чтобы рыба могла это сделать, должны быть системы очень быстрого реагирования. Молодые рыбы отрабатывают стайные техники в парах, а затем в больших группах по мере развития их техники и чувств. Стайное поведение развивается инстинктивно и не перенимается у более старых рыб. Чтобы учиться так, как они это делают, рыбам необходимы сенсорные системы, которые могут с большой скоростью реагировать на небольшие изменения в их положении относительно соседа. Большинство школ теряют способность к обучению с наступлением темноты и просто мелеют. Это указывает на то, что видение важно для школьного образования. О важности зрения говорит и поведение рыб, временно ослепших. У стайных видов глаза расположены по бокам головы, что означает, что они могут легко видеть своих соседей. Кроме того, стайные виды часто имеют «отметины стайки» на плечах или основании хвоста или визуально заметные полосы, которые служат ориентирами во время стайного обучения. [57] По функциям аналогичны пассивным маркерам при искусственном захвате движения. Однако рыбы без этих маркеров все равно будут вести стайное поведение. [58] хотя, возможно, не так эффективно. [ нужна ссылка ]
Другие чувства также используются. Феромоны или звук также могут играть свою роль, но подтверждающих доказательств пока не обнаружено. — Боковая линия это линия, проходящая по бокам рыбы от жаберных крышек до основания хвоста. В лабораторных экспериментах у стайных рыб удаляли боковые линии. Они подплыли ближе, что привело к теории о том, что боковые линии обеспечивают дополнительные стимулы, когда рыба подходит слишком близко. [57] Система боковой линии очень чувствительна к изменениям течения воды и вибрации воды. Он использует рецепторы, называемые невромастами , каждый из которых состоит из группы волосковых клеток. Волосы окружены выступающей желеобразной купулой , обычно длиной от 0,1 до 0,2 мм. Волосковые клетки боковой линии подобны волосковым клеткам внутреннего уха позвоночных, что указывает на общее происхождение боковой линии и внутреннего уха. [4]
Описание структуры мелководья
[ редактировать ]Трудно наблюдать и описывать трехмерную структуру реальных рыбных косяков из-за большого количества задействованных рыб. Методы включают использование последних достижений в области акустики рыболовства . [59]
Параметры, определяющие косяк рыб, включают:
- Размер косяка – количество рыб в косяке. На краю континентального шельфа у восточного побережья Северной Америки была использована техника дистанционного зондирования для получения изображений рыбных косяков. Косяки, состоящие, скорее всего, из атлантической сельди , щупа , хека и черного морского окуня, содержали «десятки миллионов» рыб и простирались на «многие километры». [60]
- Плотность. Плотность косяка рыбы равна количеству рыб, разделенному на объем, занимаемый косяком. Плотность не обязательно является постоянной во всей группе. Плотность рыб в косяках обычно составляет около одной рыбы на куб длины тела. [61]
 Низкая плотность
Низкая плотность Высокая плотность
Высокая плотность Низкая полярность
Низкая полярность Высокая полярность
Высокая полярность




- Полярность. Групповая полярность описывает степень, в которой все рыбы смотрят в одном направлении. Для определения этого параметра определяют среднюю ориентацию всех животных в группе. Затем для каждого животного находят угловую разницу между его ориентацией и групповой ориентацией. Групповая полярность представляет собой среднее из этих различий. [62]
- Расстояние до ближайшего соседа. Расстояние до ближайшего соседа (NND) описывает расстояние между центроидом одной рыбы (фокусной рыбы) и центроидом рыбы, ближайшей к фокусной рыбе. Этот параметр можно найти для каждой рыбы в агрегации и затем усреднить. Необходимо внимательно учитывать рыб, находящихся на краю скопления рыб, так как в одном направлении у этих рыб нет соседей. NND также связан с плотностью упаковки. У стайных рыб NND обычно составляет от половины до одной длины тела. [ нужна ссылка ]
- Позиция ближайшего соседа. В полярной системе координат позиция ближайшего соседа описывает угол и расстояние от ближайшего соседа до фокусной рыбы. [ нужна ссылка ]
- Фракция упаковки . Фракция упаковки — это параметр, заимствованный из физики для определения организации (или состояния, т. е. твердого, жидкого или газообразного) трехмерных групп рыб. Это альтернатива плотности. В этом параметре агрегация идеализируется как ансамбль твердых сфер, где каждая рыба находится в центре сферы. Фракция упаковки определяется как отношение общего объема, занимаемого всеми отдельными сферами, к общему объему агрегата. Значения варьируются от нуля до единицы, где небольшая доля упаковки представляет собой разбавленную систему, подобную газу. [63]
- Интегрированная условная плотность. Этот параметр измеряет плотность в различных масштабах длины и, следовательно, описывает однородность плотности по всей группе животных. [63]
- Функция парного распределения . Этот параметр обычно используется в физике для характеристики степени пространственного порядка в системе частиц. Она также описывает плотность, но эта мера описывает плотность на расстоянии от данной точки. Каванья и др. обнаружили, что стаи скворцов имеют больше структуры, чем газ, но меньше, чем жидкость. [63]
Моделирование школьного поведения
[ редактировать ]

![]() Моделирование Boids – требуется Java
Моделирование Boids – требуется Java
Математические модели
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( январь 2021 г. ) |
Наблюдательный подход дополняется математическим моделированием школ. Наиболее распространенные математические модели школ предписывают отдельным животным следовать трем правилам:
- Двигайтесь в том же направлении, что и ваш сосед
- Оставайтесь рядом с соседями
- Избегайте столкновений с соседями
Примером такого моделирования является программа boids , созданная Крейгом Рейнольдсом в 1986 году. [65] Другая модель — модель самодвижущихся частиц, предложенная Вичеком и др. в 1995 году [66] Во многих современных моделях используются вариации этих правил. Например, многие модели реализуют эти три правила посредством многослойных зон вокруг каждой рыбы.
- В зоне отталкивания очень близко к рыбе очаговая рыба будет стремиться дистанцироваться от соседей, чтобы избежать столкновения.
- В немного более удаленной зоне выравнивания рыба-фокус будет стремиться выровнять направление своего движения со своими соседями.
- В самой дальней зоне притяжения, которая простирается настолько далеко от фокусной рыбы, насколько она способна ее почувствовать, фокусная рыба будет стремиться переместиться к соседу.
На форму этих зон обязательно будут влиять сенсорные возможности рыбы. Рыбы полагаются как на зрение, так и на гидродинамические сигналы, передаваемые через боковую линию . Антарктический криль полагается на зрение и гидродинамические сигналы, передаваемые через его усики .
В магистерской диссертации, опубликованной в 2008 году, Моши Чарнелл описал школьное поведение без использования компонента соответствия поведения человека. [67] Его модель сводит три основных правила к следующим двум:
- Оставайтесь рядом с соседями
- Избегайте столкновений с соседями
В статье, опубликованной в 2009 году, исследователи из Исландии рассказывают о своем применении модели взаимодействующих частиц к запасам мойвы вокруг Исландии, успешно предсказывая маршрут нерестовой миграции на 2008 год. [68]
Эволюционные модели
[ редактировать ]Чтобы понять, почему у животных развивается роевое поведение , ученые обратились к эволюционным моделям, которые имитируют популяции эволюционирующих животных. Обычно в этих исследованиях используется генетический алгоритм для моделирования эволюции на протяжении многих поколений модели. В этих исследованиях был изучен ряд гипотез, объясняющих, почему животные развивают стадное поведение, например эгоистичная теория стада . [69] [70] [71] [72] эффект замешательства хищника, [33] [73] эффект разбавления, [74] [75] и теория многих глаз. [76]
Картирование формирования школ
[ редактировать ]В 2009 году, опираясь на последние достижения в области акустической визуализации, [59] [77] группа исследователей Массачусетского технологического института «впервые наблюдала образование и последующую миграцию огромного косяка рыб». [78] Результаты дают первое практическое подтверждение общих теорий о том, как ведут себя большие группы людей, от стай саранчи до стад птиц . [79]
Исследователи сфотографировали нерест атлантической сельди у отмели Жорж . Они обнаружили, что вечером рыбы собираются вместе из более глубокой воды, беспорядочно скапливаясь на мели. Цепная реакция запускается, когда плотность населения достигает критического значения, подобно волне зрителей , бегущей по спортивному стадиону. Затем происходит быстрый переход, и рыбы становятся сильно поляризованными и синхронизированными, подобно стайной рыбе. После перехода стаи начинают мигрировать, простираясь на расстояние до 40 километров (25 миль) через океан, на мелководные части берега. Там они нерестятся ночью. Утром косяк рыб снова возвращается на более глубокую воду, а затем распускается. Также были обнаружены небольшие группы лидеров, которые существенно влияли на гораздо более крупные группы. [79]
Лидерство и принятие решений
[ редактировать ]Рыбные косяки сталкиваются с решениями, которые они должны принять, если хотят остаться вместе. Например, решение может заключаться в том, в каком направлении плыть при столкновении с хищником, в каких местах остановиться и добыть корм или когда и куда мигрировать. [80]
определение кворума может функционировать как процесс коллективного принятия решений В любой децентрализованной системе . Реакция кворума определяется как «резкое увеличение вероятности того, что члены группы будут выполнять определенное поведение после превышения порогового минимального количества их товарищей по группе, уже совершающих такое поведение». [81] Недавнее исследование показало, что небольшие группы рыб использовали консенсус при принятии решений, решая, какой рыбной модели следовать. Рыбы сделали это по простому правилу кворума: люди наблюдали за решениями других, прежде чем принимать собственные решения. Эта техника обычно приводила к «правильным» решениям, но иногда приводила к «неправильным» решениям. Кроме того, по мере увеличения размера группы рыбы принимали более точные решения, следуя более привлекательной модели рыбы. [82] Принятие решений на основе консенсуса, форма коллективного разума , эффективно использует информацию из нескольких источников, чтобы в целом прийти к правильному выводу. Такое поведение было также продемонстрировано при стайном поведении трехиглой колюшки . [81]
Другие открытые вопросы о поведении косяков включают определение того, какие особи несут ответственность за направление движения косяков. В случае миграционных перемещений большинство членов косяка, по-видимому, знают, куда они направляются. Наблюдения за кормовым поведением содержащихся в неволе золотых гольянов (разновидность гольяна ) показали, что они образовывали косяки, которыми руководило небольшое количество опытных особей, которые знали, когда и где можно найти пищу. [83] Если все золотые фингеры в косяке обладают одинаковыми знаниями о наличии пищи, есть несколько особей, которые все же становятся естественными лидерами (чаще находясь впереди), и поведенческие тесты показывают, что они от природы смелее. [84] Меньшие золотые фингеры, кажется, охотнее, чем более крупные, находятся ближе к передней части косяка, возможно, потому, что они более голодны. [85] Наблюдения за обыкновенной плотвой показали, что особи, лишенные пищи, как правило, находятся в передней части косяка, где они получают больше еды. [86] [87] но там они также могут быть более уязвимы для хищников, устраивающих засады. [88] Особи, опасающиеся хищников, склонны искать более центральные позиции внутри косяков. [89]
Выбор мелководья
[ редактировать ]
Экспериментальные исследования предпочтений мелководья провести относительно легко. Аквариум, содержащий выбирающую рыбу, зажат между двумя аквариумами с разными косяками, и предполагается, что выбирающая рыба проводит больше времени рядом с косяком, который она предпочитает. Исследования такого рода выявили несколько факторов, важных для предпочтения мелководья. [ нужна ссылка ]
Рыбы обычно предпочитают более крупные косяки. [90] [91] Это имеет смысл, поскольку более крупный косяк обычно обеспечивает лучшую защиту от хищников. Действительно, предпочтение более крупных косяков кажется более сильным, когда поблизости находятся хищники. [92] [93] или у видов, которые больше полагаются на мелководье, чем на бронежилеты, защищаясь от хищников. [94] Более крупные косяки также могут быстрее находить пищу, хотя эту пищу придется разделить между большим количеством особей. Конкуренция может означать, что голодные особи могут предпочитать более мелкие косяки или проявлять меньшее предпочтение к очень большим косякам, как показано на примере колюшек. [95] [96]
Рыбы предпочитают скапливаться в стаях представителей своего вида. Иногда несколько видов могут смешиваться в одной косяке, но при попадании в такие косяки хищника рыбы перестраиваются так, что каждая особь оказывается ближе к представителям своего вида. [97]
Рыбы, как правило, предпочитают косяки, состоящие из особей, соответствующих их размеру. [98] [99] [100] Это имеет смысл, поскольку хищникам легче ловить особей, выделяющихся в стае. Некоторые рыбы могут даже предпочитать косяки других видов, если это означает, что они лучше соответствуют текущему размеру тела. [101] Однако что касается размера косяка, голод может повлиять на предпочтение рыбы аналогичного размера; например, крупная рыба может предпочесть общаться с более мелкой рыбой из-за конкурентного преимущества, которое они получат перед этими мелководными товарищами. У золотого шинера крупные сытые рыбы предпочитают общаться с другими крупными особями, а голодные предпочитают более мелкие косяки. [102]
Рыбы предпочитают скапливаться с особями, с которыми выбирающая рыба уже знакома. Это было продемонстрировано на гуппи . [103] [104] трехиглая колюшка , [105] полосатый киллифиш , [106] окунь для серфинга Embiotoca jacksoni , [107] Мексиканская тетра , [108] и различные пескари. [109] [110] Исследование гольяна с горы Белого Облака также показало, что выбирающие рыбу предпочитают стаи с особями, которые придерживаются той же диеты, что и они сами. [111]
Было показано, что колюшки и киллифиши предпочитают косяки, состоящие из здоровых особей, а не зараженных паразитами, на основании визуальных признаков паразитизма и аномального поведения паразитированных рыб. [112] [113] [114] [115] Рыбки данио предпочитают косяки, состоящие из сытой (с большей шириной желудка) рыбы, а не рыб, лишенных пищи. [116]
Трехиглая колюшка предпочитает присоединяться к косяку, состоящему из смелых особей, а не пугливых. [117] Рыбы-ангелы предпочитают стаи, состоящие из подчиненных, а не доминирующих особей. [118] Европейский гольян может различать косяки, состоящие из хороших и плохих конкурентов, даже при отсутствии очевидных признаков, таких как различия в агрессивности, размере или скорости кормления; они предпочитают сотрудничать с бедными конкурентами. [119] Все это предполагает стратегию добычи еды, поскольку более смелые люди с большей вероятностью найдут еду, в то время как подчиненные будут предлагать меньшую конкуренцию за найденную еду. [ нужна ссылка ]
Рыбы предпочитают объединяться в косяки, которые активно кормятся. [120] [121] Золотой фингал также может обнаруживать упреждающую активность косяков, которые ожидают скорого кормления, и предпочтительно присоединяться к таким косякам. [122] Рыбки данио также выбирают более активные косяки. [123]
Коммерческое рыболовство
[ редактировать ]Стайное поведение рыб используется в промышленных масштабах промысловым рыболовством . Огромные кошельковые сейнеры используют самолеты-корректировщики для обнаружения стайной рыбы, такой как тунец, треска, скумбрия и кормовая рыба. Они могут захватывать огромные косяки, быстро окружая их кошельковыми неводами с помощью быстрых вспомогательных лодок и сложного гидролокатора, способного отслеживать форму косяка. [ нужна ссылка ]
Дальнейшие примеры
[ редактировать ]
Рыбы -кузнецы живут рыхлыми косяками. У них есть симбиотические отношения с паразитом, питающимся рыбой-сеноритой. Когда они встречают косяк рыб-сенорит, они останавливаются, образуют плотный шар и висят вверх ногами (на фото), каждая рыба ждет своей очереди, чтобы ее почистили. Рыба-сенорита собирает мертвые ткани и внешних паразитов, таких как паразитические копекоды и изокоды, с кожи других рыб. [ нужна ссылка ]
Некоторые косяки занимаются моббинговым поведением . Например, синежабрники образуют большие гнездовые колонии и иногда нападают на щелкающих черепах . Это может служить для рекламы своего присутствия, изгнания хищника из этого района или помощи в культурной передаче признания хищника. [124]
Пираньи имеют репутацию бесстрашных рыб, охотящихся свирепыми стаями. Однако недавние исследования, которые «начались с предпосылки, что они стайиваются как средство совместной охоты», обнаружили, что на самом деле они были довольно пугливыми рыбами, как и другие рыбы, которые обучались для защиты от хищников, таких как бакланы и кайманы. и дельфины. Пираньи «по сути похожи на обычных рыб с большими зубами». [125]
Кальмары Гумбольдта — крупные хищные морские беспозвоночные, которые перемещаются стаями численностью до 1200 особей. Они плавают со скоростью до 24 километров в час (15 миль в час или 13 узлов), приводимые в движение водой, выбрасываемой через сифон и два треугольных плавника. На их щупальцах имеются присоски с острыми зубами, которыми они хватают добычу и тащат ее к большому острому клюву. В течение дня кальмары Гумбольдта ведут себя подобно мезопелагическим рыбам , обитающим на глубинах от 200 до 700 м (от 660 до 2300 футов). Электронное мечение показало, что они также подвергаются ежедневным вертикальным миграциям , которые приближают их к поверхности от заката до рассвета. [126] Они охотятся у поверхности по ночам, пользуясь темнотой, чтобы использовать свое острое зрение, чтобы добыть более обильную добычу. Кальмары питаются в основном мелкой рыбой , ракообразными , головоногими моллюсками и веслоногими раками и охотятся за своей добычей совместно, что является первым наблюдением такого поведения у беспозвоночных. [127] Также известно, что кальмары Гумбольдта быстро пожирают более крупную добычу при совместной охоте в группах. Кальмары Гумбольдта известны своей скоростью в поедании пойманной на крючок рыбы, акул и кальмаров, даже представителей своего вида и косяков. [128] и, как известно, нападали на рыбаков и дайверов. [129]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Другие собирательные существительные, используемые для обозначения рыбы, включают тягу рыбы, дрейф рыбы или чешую рыбы. , используемые для конкретных групп видов рыб или морских животных, включают измельчение черной рыбы, беспокойство золотой рыбки, сбор сельди, привязку или бег лосося, дрожь акул, лихорадку скатов, запах тилапии, парение форели и стручка Собирательные существительные киты. [ нужна ссылка ]
- ^ Обмеление — это особый случай агрегирования, а стайное образование — особый случай обмеления. часто рассматривают их как синонимы Хотя стайное обучение и обмеление в биологии означают разные вещи, неспециалисты : носители британского английского склонны использовать слово «обмеление» для описания любой группы рыб, тогда как носители американского английского склонны использовать «стайное обучение». так же свободно. [1]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и Питчер и Пэриш, 1993, стр. 365.
- ^ Шоу, Э. (1978). «Стульная рыбка». Американский учёный . 66 (2): 166–175. Бибкод : 1978AmSci..66..166S .
- ^ Хелфман Г., Коллетт Б. и Фейси Д.: Разнообразие рыб, Blackwell Publishing, стр. 375, 1997, ISBN 0-86542-256-7
- ^ Перейти обратно: а б с д и ж г Мойл П.Б. и Чех Дж.Дж. (2003) Рыбы, Введение в ихтиологию. 5-е изд, Бенджамин Каммингс. ISBN 978-0-13-100847-2
- ^ Питчер, Т.Дж. и Пэрриш, Дж.К. (1993) Поведение костистых рыб, Глава 12: Функции стайного поведения костистых рыб Springer. ISBN 978-0-412-42930-9
- ^ Бредер, К.М. младший (1967). «О значении выживания рыбных косяков». Зоология . 52 : 25–40.
- ^ Перейти обратно: а б Партридж, Б.; Питчер, Т.; Каллен, М.; Уилсон, Дж. (1980). «Трехмерная структура рыбных косяков». Поведение Экол Социобиол . 6 (4): 277–288. дои : 10.1007/BF00292770 . S2CID 8032766 .
- ^ Перейти обратно: а б Килс, У. (1992). «EcoSCOPE и dynIMAGE: микромасштабные инструменты для исследования взаимодействий хищник-жертва in situ». Арка Гидробиол Бей . 36 : 83–96.
- ^ Национальная коалиция по сохранению морской среды : Кормовая рыба
- ^ Радаков Д.В. (1973) Обучение экологии рыб. Израильская программа научного перевода, перевод Mill H. Halsted Press, Нью-Йорк. ISBN 978-0-7065-1351-6
- ^ Ноттестад, Л.; Аксельсен, Бельгия (1999). «Маневры по стайке сельди в ответ на нападения косаток» (PDF) . Канадский журнал зоологии . 77 (10): 1540–1546. дои : 10.1139/z99-124 . Архивировано из оригинала (PDF) 17 декабря 2008 года.
- ^ Перейти обратно: а б Хоар, диджей; Краузе, Дж.; Пеукури, Н.; Годин, JGJ (2000). «Размер тела и стайность рыб». Журнал биологии рыб . 57 (6): 1351–1366. Бибкод : 2000JFBio..57.1351H . дои : 10.1111/j.1095-8649.2000.tb02217.x .
- ^ Перейти обратно: а б Ланда, Дж.Т. (1998). «Биоэкономика стайных рыб: эгоистичные рыбы, квазибезбилетники и другие рыбные сказки». Экологическая биология рыб . 53 (4): 353–364. Бибкод : 1998EnvBF..53..353L . дои : 10.1023/А:1007414603324 . S2CID 12674762 .
- ^ Пэрриш, Дж. К. ; Вишедо, Южная Каролина; Грюнбаум, Д. (2002). «Самоорганизованные рыбные косяки: исследование новых свойств». Биологический бюллетень . 202 (3): 296–305. CiteSeerX 10.1.1.116.1548 . дои : 10.2307/1543482 . JSTOR 1543482 . ПМИД 12087003 . S2CID 377484 .
- ^ Надлер, Лорен Э.; Киллен, Шон С.; МакКлюр, Ева С.; Мандей, Филип Л.; Маккормик, Марк И. (15 сентября 2016 г.). «Обмеление снижает скорость метаболизма у стадных видов коралловых рифовых рыб» . Журнал экспериментальной биологии . 219 (18): 2802–2805. дои : 10.1242/jeb.139493 . ПМК 5047653 . ПМИД 27655821 .
- ^ Абрахамс, М.; Колган, П. (1985). «Риск хищничества, гидродинамическая эффективность и их влияние на структуру школы». Экологическая биология рыб . 13 (3): 195–202. Бибкод : 1985EnvBF..13..195A . дои : 10.1007/BF00000931 . S2CID 22329892 .
- ^ Питчер, Т.; Магурран, А.; Уинфилд, И. (1982). «Рыба в больших косяках быстрее находит пищу». Поведение. Экол. Социобиол . 10 (2): 149–151. дои : 10.1007/BF00300175 . S2CID 6340986 .
- ^ Партридж, Б.; Йоханссон, Дж.; Калиш, Дж. (1983). «Структура косяков гигантского голубого тунца в заливе Кейп-Код». Экологическая биология рыб . 9 (3–4): 253–262. Бибкод : 1983EnvBF...9..253P . дои : 10.1007/BF00692374 . S2CID 6799134 .
- ^ Перейти обратно: а б с Питчер и Пэриш, 1993 г.
- ^ «Ветровые поверхностные течения: апвеллинг и даунвеллинг» .
- ^ Биология веслоногих ракообразных. Архивировано 1 января 2009 г. в Wayback Machine в Ольденбургском университете Карла фон Осецкого.
- ^ Гьёсетер, Х. (1998). «Популяционная биология и промысел мойвы ( Mallotus villosus ) в Баренцевом море». Сарсия . 83 (6): 453–496. дои : 10.1080/00364827.1998.10420445 .
- ^ Фиш, FE (1995). «Кинематика плавания утят строем: последствия положения». Журнал экспериментальной зоологии . 273 (1): 1–11. дои : 10.1002/jez.1402730102 . S2CID 49732151 .
- ^ Александр, Р. Макнил (2004). «Гидродинамическое прицепление к подъемной силе – в плавании, полете и велоспорте» . Журнал биологии . 3 (2): 7. doi : 10.1186/jbiol5 . ПМК 416560 . ПМИД 15132738 .
- ^ Маррас, Стефано; Киллен, Шон С.; Линдстрем, Ян; Маккензи, Дэвид Дж.; Стеффенсен, Джон Ф.; Доменичи, Паоло (февраль 2015 г.). «Рыбы, плавающие стаями, экономят энергию независимо от их пространственного положения» . Поведенческая экология и социобиология . 69 (2): 219–226. дои : 10.1007/s00265-014-1834-4 . ПМЦ 4293471 . ПМИД 25620833 .
- ^ Маклин, Стефани; Перссон, Анна; Норин, Томми; Киллен, Шон С. (апрель 2018 г.). «Метаболические затраты на кормление прогнозируемо изменяют пространственное распределение особей в рыбных косяках» . Современная биология . 28 (7): 1144–1149.e4. Бибкод : 2018CBio...28E1144M . дои : 10.1016/j.cub.2018.02.043 . ПМИД 29576472 .
- ^ Перейти обратно: а б Апарна, Бхадури (2011) Сказки пересмешника: материалы по поведению животных Колледж OpenStax .
- ^ Хоар, диджей; Кузен, ID; Годин, Дж. Г.; Краузе, Дж. (2004). «Выбор размера группы рыб в зависимости от контекста». Поведение животных . 67 (1): 155–164. дои : 10.1016/j.anbehav.2003.04.004 . S2CID 2244463 .
- ^ Милински, Х.; Хеллер, Р. (1978). «Влияние хищника на оптимальное пищевое поведение колюшек». Природа . 275 (5681): 642–644. Бибкод : 1978Natur.275..642M . дои : 10.1038/275642a0 . S2CID 4184043 .
- ^ Йешке Дж. М., Толлриан Р.; Толлриан, Ральф (2007). «Роение добычи: какие хищники сбиваются с толку и почему?». Поведение животных . 74 (3): 387–393. дои : 10.1016/j.anbehav.2006.08.020 . S2CID 53163951 .
- ^ Иоанну CC; Тош ЧР; Невилл Л; Краузе Дж. (2008). «Эффект замешательства — от нейронных сетей до снижения риска нападения хищников» . Поведенческая экология . 19 (1): 126–130. дои : 10.1093/beheco/arm109 .
- ^ Кракауэр, округ Колумбия (1995). «Группы сбивают с толку хищников, используя узкие места восприятия: коннекционистская модель эффекта замешательства». Поведенческая экология и социобиология . 36 (6): 421–429. дои : 10.1007/BF00177338 . S2CID 22967420 .
- ^ Перейти обратно: а б Олсон Р.С.; Хинтце А; Дайер ФК; Кнестер Д.Б.; Адами С (2013). «Замешательства хищников достаточно, чтобы развить роевое поведение» . JR Soc. Интерфейс . 10 (85): 20130305.arXiv : 1209.3330 . дои : 10.1098/rsif.2013.0305 . ПМК 4043163 . ПМИД 23740485 .
- ^ Перейти обратно: а б с Ларссон, М. (2009). «Возможные функции октаволатеральной системы при стайке рыб» . Рыба и рыболовство . 10 (3): 344–355. Бибкод : 2009AqFF...10..344L . дои : 10.1111/j.1467-2979.2009.00330.x .
- ^ Ларссон, М (2011). «Случайные звуки передвижения в познании животных» . Познание животных . 15 (1): 1–13. дои : 10.1007/s10071-011-0433-2 . ПМК 3249174 . ПМИД 21748447 .
- ^ Перейти обратно: а б с Ларссон, М. (2012). «Почему рыбная школа?» . Современная зоология . 58 (1): 116–128. дои : 10.1093/czoolo/58.1.116 .
- ^ Нью, Дж.Г.; Фьюкс, Луизиана; Хан, АН (2001). «Ударное пищевое поведение при мускусе, масквинонгия Esox: вклад боковой линии и зрительных сенсорных систем». J Exp Biol . 204 (6): 1207–1221. дои : 10.1242/jeb.204.6.1207 . ПМИД 11222136 .
- ^ Бабино, Д; Льюис, Дж. Э.; Лонгтин, А (2007). «Пространственная острота и обнаружение добычи у слабоэлектрических рыб» . ПЛОС Компьютерная Биол . 3 (3): 402–411. Бибкод : 2007PLSCB...3...38B . дои : 10.1371/journal.pcbi.0030038 . ПМК 1808493 . ПМИД 17335346 .
- ^ Робертс, Дж. (1996). «Почему индивидуальная бдительность снижается по мере увеличения размера группы». Анимальное поведение . 51 (5): 1077–1086. CiteSeerX 10.1.1.472.7279 . дои : 10.1006/anbe.1996.0109 . S2CID 53202810 .
- ^ Лима, С. (1995). «Назад к основам бдительности против хищников: эффект размера группы». Поведение животных . 49 (1): 11–20. дои : 10.1016/0003-3472(95)80149-9 . S2CID 53205760 .
- ^ Морс, Д.Х. (1977). «Пищевое поведение и избегание хищников в гетероспецифичных группах». Бионаука . 27 (5): 332–339. дои : 10.2307/1297632 . JSTOR 1297632 .
- ^ Гамильтон, WD (1971). «Геометрия для эгоистичного стада». Дж. Теория биологии . 31 (2): 295–311. Бибкод : 1971JThBi..31..295H . дои : 10.1016/0022-5193(71)90189-5 . ПМИД 5104951 .
- ^ Тернер, Г.; Питчер, Т. (1986). «Снижение атак: модель групповой защиты путем комбинированного предотвращения и разбавления». Американский натуралист . 128 (2): 228–240. дои : 10.1086/284556 . S2CID 84738064 .
- ^ Краузе, Дж.; Ракстон, Г.; Рубинштейн, Д. (1998). «Всегда ли размер косяка влияет на успех охоты хищника?». Журнал биологии рыб . 52 (3): 494–501. дои : 10.1006/jfbi.1997.0595 .
- ^ «Морские ученые чешут головы над сардинами» . Архивировано из оригинала 25 сентября 2004 года.
- ^ Перейти обратно: а б с д и «Феномен безумного кормления акул сардинами в Африке» . Архивировано из оригинала 2 декабря 2008 года.
- ^ Лахлан, РФ; Крукс, Л; Лаланд, КН (1998). «Кто за кем следует? Мелководные предпочтения и социальное обучение гуппи поиску информации». Поведение животных . 56 (1): 181–190. дои : 10.1006/anbe.1998.0760 . ПМИД 9710476 . S2CID 30973104 .
- ^ Зейтц, JC Pelagic Thresher . Флоридский музей естественной истории. Проверено 22 декабря 2008 г.
- ^ Оливер, СП; Тернер, младший; Ганн, К; Сильвоса, М; Д'Урбан Джексон, Т (2013). «Акулы-молотилки используют удары хвостом в качестве охотничьей стратегии» . ПЛОС ОДИН . 8 (7): е67380. Бибкод : 2013PLoSO...867380O . дои : 10.1371/journal.pone.0067380 . ПМЦ 3707734 . ПМИД 23874415 .
- ^ Компаньо, LJV (1984). Акулы мира: аннотированный и иллюстрированный каталог известных на сегодняшний день видов акул . Рим: Продовольственная и сельскохозяйственная организация. стр. 466–468. ISBN 978-92-5-101384-7 .
- ^ « Carcharhinus brevipinna , акула-вертушка» . MarineBio.org . Проверено 9 мая 2009 г.
- ^ Перейти обратно: а б с Газда, СК; Коннор, RC; Эдгар, РК; Кокс, Ф (2005). «Разделение труда с ролевой специализацией на групповой охоте на афалин ( Tursiops truncatus ) у берегов Сидар-Ки, Флорида» . Труды Королевского общества Б. 272 (1559): 135–140. дои : 10.1098/rspb.2004.2937 . ПМК 1634948 . ПМИД 15695203 .
- ^ «Прибрежные запасы атлантического афалины: обзор состояния и управление», Материалы и рекомендации семинара, проведенного в Бофорте, Северная Каролина, 13 сентября 1993 г. - 14 сентября 1993 г. Министерство торговли США, Национальное управление океанических и атмосферных исследований, Национальная служба морского рыболовства. стр. 56–57.
- ^ Ренделл, Л.; Уайтхед, Х. (2001). «Культура китов и дельфинов». Поведенческие и мозговые науки . 24 (2): 309–382. дои : 10.1017/S0140525X0100396X . ПМИД 11530544 . S2CID 24052064 .
- ^ Ривз Р.Р., Стюарт Б.С., Клэпхэм П.Дж. и Пауэлл Дж.А. (2002) Руководство Национального общества Одюбона по морским млекопитающим World Chanticleer Press. ISBN 9780375411410 .
- ^ Потвин, Дж; Гольдбоген, Дж.А.; Шедвик, RE (2009). « Пассивное и активное поглощение: вердикт на основе моделирования траектории кормящихся финвалов Balaenoptera physalus» . JR Soc. Интерфейс . 6 (40): 1005–1025. дои : 10.1098/rsif.2008.0492 . ПМЦ 2827442 . ПМИД 19158011 .
- ^ Перейти обратно: а б Боун Кью и Мур Р.Х. (2008) Биология рыб, стр. 418–422, Taylor & Francisco Group. ISBN 978-0-415-37562-7
- ^ BBC News Online , Рыба-робоб принят косяком диких рыб , 09:54 по Гринвичу, четверг, 1 июля 2010 г., 10:54 Великобритания
- ^ Перейти обратно: а б «Одна рыба, две рыбы: новый датчик MIT улучшает подсчет рыбы» . Физика.орг . Проверено 13 сентября 2014 г.
- ^ Макрис, Северная Каролина; Ратилал, П.; Саймондс, DT; Джаганнатан, С.; Ли, С.; Неро, RW (2006). «Популяция и поведение рыб, выявленные с помощью мгновенной визуализации в масштабе континентального шельфа». Наука . 311 (5761): 660–663. Бибкод : 2006Sci...311..660M . дои : 10.1126/science.1121756 . ПМИД 16456080 . S2CID 140558930 .
- ^ Питчер, Ти Джей; Партридж, ТЛ (1979). «Плотность и объем косяка рыбы». Мар Биол . 54 (4): 383–394. Бибкод : 1979MarBi..54..383P . дои : 10.1007/BF00395444 . S2CID 84810154 .
- ^ Вишидо, С.; Пэрриш, Дж .; Грюнбаум, Д. (2004). «Индивидуальное поведение и возникающие свойства рыбных косяков: сравнение наблюдений и теории» (PDF) . Серия «Прогресс в области морской экологии» . 273 : 239–249. Бибкод : 2004MEPS..273..239В . дои : 10.3354/meps273239 .
- ^ Перейти обратно: а б с Каванья, А.; Чимарелли, Джардина; Орланди, Паризи; Прокаччини, Сантагати; Стефанини (2008). «Новые статистические инструменты для анализа структуры групп животных». Математические биологические науки . 214 (1–2): 32–37. дои : 10.1016/j.mbs.2008.05.006 . ПМИД 18586280 .
- ^ «Модель самодвижущихся частиц» . ФЕТ . Проверено 13 сентября 2014 г.
- ^ Рейнольдс, CW (1987). «Стада, стада и школы: распределенная поведенческая модель». Материалы 14-й ежегодной конференции по компьютерной графике и интерактивным технологиям - SIGGRAPH '87 . Том. 21. С. 25–34. CiteSeerX 10.1.1.103.7187 . дои : 10.1145/37401.37406 . ISBN 978-0897912273 . S2CID 546350 .
- ^ Вичек, Т ; Чирок, А; Бен-Джейкоб, Э; Коэн, я; Шошет, О (1995). «Новый тип фазового перехода в системе самодвижущихся частиц». Письма о физических отзывах . 75 (6): 1226–1229. arXiv : cond-mat/0611743 . Бибкод : 1995PhRvL..75.1226V . дои : 10.1103/PhysRevLett.75.1226 . ПМИД 10060237 . S2CID 15918052 .
- ^ Чарнелл, М. (2008) «Индивидуальное моделирование экологических систем и социальных агрегаций» . Скачать [ постоянная мертвая ссылка ]
- ^ Барбаро А., Эйнарссон Б., Бирнир Б., Сигурдссон С., Валдимарссон С., Палссон О.К., Свейнбьёрнссон С., Сигурдссон П. (2009). «Моделирование и моделирование миграции пелагических рыб» . Журнал морских наук ICES . 66 (5): 826–838. doi : 10.1093/icesjms/fsp067 .
- ^ Олсон Р.С.; Кнестер Д.Б.; Адами С (2013). «Критическое взаимодействие между хищничеством, зависящим от плотности, и эволюцией эгоистичного стада». Материалы 15-й ежегодной конференции по генетическим и эволюционным вычислениям . стр. 247–254. дои : 10.1145/2463372.2463394 . ISBN 9781450319638 . S2CID 14414033 .
- ^ Уорд ЧР; Гобет Ф; Кендалл Дж. (2001). «Эволюционирование коллективного поведения в искусственной экологии» . Искусственная жизнь . 7 (2): 191–209. CiteSeerX 10.1.1.108.3956 . дои : 10.1162/106454601753139005 . ПМИД 11580880 . S2CID 12133884 .
- ^ Релуга ТС, Вискидо С; Вискидо, Стивен (2005). «Имитация эволюции эгоистичного стадного поведения». Журнал теоретической биологии . 234 (2): 213–225. Бибкод : 2005JThBi.234..213R . дои : 10.1016/j.jtbi.2004.11.035 . ПМИД 15757680 .
- ^ Вуд А.Дж., Экленд Г.Дж.; Экланд, GJ (2007). «Эволюция эгоистичного стада: появление различных стратегий агрегирования в индивидуальной модели» . Учебник по биологическим наукам . 274 (1618): 1637–1642. дои : 10.1098/rspb.2007.0306 . ПМК 2169279 . ПМИД 17472913 .
- ^ Демсар Дж; Хемельрейк СК; Хильденбрандт Х и Баец, Иллинойс (2015). «Имитация нападения хищников на школы: развитие комплексной тактики» (PDF) . Экологическое моделирование . 304 : 22–33. Бибкод : 2015EcMod.304...22D . doi : 10.1016/j.ecolmodel.2015.02.018 . hdl : 11370/0bfcbb69-a101-4ec1-833a-df301e49d8ef . S2CID 46988508 .
- ^ Тош ЧР (2011). «Какие условия способствуют отбору, зависящему от отрицательной плотности скоплений добычи?» (PDF) . Журнал теоретической биологии . 281 (1): 24–30. Бибкод : 2011JThBi.281...24T . дои : 10.1016/j.jtbi.2011.04.014 . ПМИД 21540037 .
- ^ Иоанну CC; Гутталь В; Кузен И.Д. (2012). «Выбор хищных рыб для скоординированного коллективного движения виртуальной добычи» . Наука . 337 (6099): 1212–1215. Бибкод : 2012Sci...337.1212I . дои : 10.1126/science.1218919 . ПМИД 22903520 . S2CID 10203872 .
- ^ Олсон Р.С.; Хейли П.Б.; Дайер ФК и Адами С (2015). «Изучение эволюции компромисса между бдительностью и поиском пищи у групповых организмов» . Королевское общество открытой науки . 2 (9): 150135. arXiv : 1408.1906 . Бибкод : 2015RSOS....250135O . дои : 10.1098/rsos.150135 . ПМЦ 4593673 . ПМИД 26473039 .
- ^ Макрис, Северная Каролина; Ратилал, П; Саймондс, DT; Джаганнатан, С; Ли, С; Неро, RW (2006). «Популяция и поведение рыб, выявленные с помощью мгновенной визуализации в масштабе континентального шельфа». Наука . 311 (5761): 660–663. Бибкод : 2006Sci...311..660M . дои : 10.1126/science.1121756 . ПМИД 16456080 . S2CID 140558930 .
- ^ Макрис, Северная Каролина; Ратилал, П; Джаганнатан, С; Гонг, З; Эндрюс, М; Берцатос, я; Годё, Орегон; Нерон, RW; Джех, М; и др. (2009). «Критическая плотность населения вызывает быстрое образование огромных косяков океанических рыб». Наука . 323 (5922): 1734–1737. Бибкод : 2009Sci...323.1734M . дои : 10.1126/science.1169441 . ПМИД 19325116 . S2CID 6478019 .
- ^ Перейти обратно: а б «Ученые идентифицируют генезис моделей поведения животных» . Проверено 13 сентября 2014 г.
- ^ Самптер, Д. «Коллективное поведение» .
- ^ Перейти обратно: а б Уорд, Эй Джей; Краузе, Дж; Самптер, диджей (2012). «Кворум принятия решений в кормовых рыбных косяках» . ПЛОС ОДИН . 7 (3): e32411. Бибкод : 2012PLoSO...732411W . дои : 10.1371/journal.pone.0032411 . ПМК 3296701 . ПМИД 22412869 .
- ^ Самптер, Д.; Краузе, Дж; Джеймс, Р.; Кузен, И.; Уорд, А. (2008). «Принятие рыбами консенсусного решения» . Современная биология . 18 (22): 1773–1777. Бибкод : 2008CBio...18.1773S . дои : 10.1016/j.cub.2008.09.064 . ПМИД 19013067 . S2CID 11362054 .
- ^ Рибс, С.Г. (2000). «Может ли меньшинство информированных лидеров определить перемещения косяка рыб в поисках пищи?». Поведение животных . 59 (2): 403–409. дои : 10.1006/anbe.1999.1314 . ПМИД 10675263 . S2CID 4945309 .
- ^ Леблон, К.; Рибс, С.Г. (2006). «Индивидуальное лидерство и смелость в стаях золотых фингеров ( Notemigonus crisoleucas )». Поведение . 143 (10): 1263–1280. дои : 10.1163/156853906778691603 . S2CID 56117643 .
- ^ Рибс, С.Г. (2001). «Влияние размеров тела на лидерство в стаях золотистых фингеров Notemigonus Crysoleucas ». Поведение . 138 (7): 797–809. дои : 10.1163/156853901753172656 .
- ^ Краузе, Дж. (1993). «Взаимосвязь между добычей пищи и положением косяка в смешанной стае плотвы ( Rutilus rutilus ) и голавля ( Leuciscus leuciscus ): полевое исследование». Экология . 93 (3): 356–359. Бибкод : 1993Oecol..93..356K . дои : 10.1007/bf00317878 . ПМИД 28313435 . S2CID 13140673 .
- ^ Краузе, Дж.; Буманн, Д.; Тодт, Д. (1992). «Взаимосвязь между предпочтением позиции и состоянием питания особей в стаях молоди плотвы ( Rutilus rutilus )». Поведенческая экология и социобиология . 30 (3–4): 177–180. дои : 10.1007/bf00166700 . S2CID 32061496 .
- ^ Буманн, Д.; Краузе, Дж.; Рубинштейн, Д. (1997). «Риск смертности от пространственного положения в группах животных: опасность оказаться впереди». Поведение . 134 (13): 1063–1076. дои : 10.1163/156853997x00403 .
- ^ Краузе, Дж. (1993). «Влияние Шрекстоффа на мелководное поведение гольяна: проверка эгоистичной стадной теории Гамильтона» . Поведение животных . 45 (5): 1019–1024. дои : 10.1006/anbe.1993.1119 . S2CID 54287659 .
- ^ Кинлисайд, MHA (1955). «Некоторые аспекты стайного поведения рыб» (PDF) . Поведение . 8 : 183–248. дои : 10.1163/156853955x00229 . S2CID 86589740 .
- ^ Тедегер, RW; Краузе, Дж. (1995). «Зависимость от плотности и многочисленность в страхе стимулировали агрегационное поведение стайной рыбы». Философские труды Лондонского королевского общества Б. 350 (1334): 381–390. Бибкод : 1995РСТБ.350..381Т . дои : 10.1098/rstb.1995.0172 .
- ^ Хагер, MC; Хельфман, Г.С. (1991). «Безопасность в численности: выбор размера косяка гольянами в условиях угрозы хищников». Поведенческая экология и социобиология . 29 (4): 271–276. дои : 10.1007/bf00163984 . S2CID 30901973 .
- ^ Эшли, Э.Дж.; Кац, Л.Б.; Вулф, JW (1993). «Балансирование компромисса между риском и изменением размера косяка северного краснобрюхого ельца (Phoxinus eos)». Копейя . 1993 (2): 540–542. дои : 10.2307/1447157 . JSTOR 1447157 .
- ^ Краузе, Дж.; Годен, Ж.-Г.Дж.; Рубинштейн, Д. (1998). «Выбор группы как функция различий в размерах групп и времени оценки рыб: влияние уязвимости видов перед хищниками». Этология . 104 (1): 68–74. Бибкод : 1998Этол.104...68К . дои : 10.1111/j.1439-0310.1998.tb00030.x .
- ^ ван Гавр, Н.; Фитцджеральд, Дж.Дж. (1988). «Обмеление и распознавание родства у трехиглой колюшки (Gasterosteus aculeatus L.)». Биология поведения . 13 : 190–201.
- ^ Краузе, Дж. (1993). «Влияние голода на выбор размера косяка трехиглой колюшкой Gasterosteus aculeatus». Журнал биологии рыб . 43 (5): 775–780. Бибкод : 1993JFBio..43..775K . дои : 10.1111/j.1095-8649.1993.tb01154.x .
- ^ Аллан-младший; Питчер, Ти Джей (1986). «Сегрегация видов при бегстве хищников в косяках карповых рыб». Пресноводная биология . 16 (5): 653–659. Бибкод : 1986FrBio..16..653A . дои : 10.1111/j.1365-2427.1986.tb01007.x .
- ^ Ранта, Э.; Линдстрем, К.; Пеукури, Н. (1992). «Когда трехиглая колюшка идет в школу, размер имеет значение». Поведение животных . 43 : 160–162. дои : 10.1016/s0003-3472(05)80082-x . S2CID 53177367 .
- ^ Ранта, Э.; Ювонен, С.-К.; Пеукури, Н. (1992). «Дополнительные доказательства стайного обучения колюшек в зависимости от размера». Журнал биологии рыб . 41 (4): 627–630. Бибкод : 1992JFBio..41..627R . дои : 10.1111/j.1095-8649.1992.tb02689.x .
- ^ Краузе, Дж. (1994). «Влияние пищевой конкуренции и риска нападения хищников на стайку молоди голавля ( Leuciscus cephalus ) по размеру». Этология . 96 (2): 105–116. Бибкод : 1994Этол..96..105К . дои : 10.1111/j.1439-0310.1994.tb00886.x .
- ^ Краузе, Дж.; Годин (2010). «Выбор косяков у полосатой рыбы-убийцы ( Fundulusdiaphanus , Teleostei, Cyprinodontidae ): влияние риска нападения хищников, размера рыбы, видового состава и размера косяков». Этология . 98 (2): 128–136. дои : 10.1111/j.1439-0310.1994.tb01063.x .
- ^ Рибс, С.Г.; Солнье, Н. (1997). «Влияние голода на выбор косяков у золотых фингеров (Pisces: Cyprinidae, Notemigonusrysoleucas )». Этология . 103 (8): 642–652. Бибкод : 1997Этол.103..642Р . дои : 10.1111/j.1439-0310.1997.tb00175.x .
- ^ Магурран, А.Е.; Сегерс, Б.Х.; Шоу, П.В.; Карвальо, Греция (1994). «Школьные предпочтения знакомой рыбы гуппи Poecilia reticulata». Журнал биологии рыб . 45 (3): 401–406. дои : 10.1006/jfbi.1994.1142 .
- ^ Гриффитс, Юго-Запад; Магурран, А.Е. (1999). «Решения о школьном обучении гуппи ( Poecilia reticulata ) основаны на знакомстве, а не на распознавании родственников путем совпадения фенотипов». Поведенческая экология и социобиология . 45 (6): 437–443. дои : 10.1007/s002650050582 . S2CID 23085058 .
- ^ Барбер, И.; Ракстон, Джорджия (2000). «Важность стабильного школьного образования: держатся ли знакомые колюшки вместе?» . Труды Лондонского королевского общества Б. 267 (1439): 151–155. дои : 10.1098/rspb.2000.0980 . ПМК 1690514 . ПМИД 10687820 .
- ^ Ли-Дженкинс, SSY; Годен, Ж.-ГЖ (2010). «Социальное знакомство и образование косяков у молоди рыб». Журнал биологии рыб . 76 (3): 580–590. Бибкод : 2010JFBio..76..580L . дои : 10.1111/j.1095-8649.2009.02512.x . ПМИД 20666898 .
- ^ Сиккель, ПК; Фуллер, Калифорния (2010). «Выявление предпочтений и доказательства существования групп братьев и сестер молодыми черными окунями Embiotoca jacksoni ». Журнал биологии рыб . 76 (7): 1671–1681. Бибкод : 2010JFBio..76.1671S . дои : 10.1111/j.1095-8649.2010.02607.x . ПМИД 20557623 .
- ^ Де Фрайпон, М.; Тайнс, Г. (1986). «Реакция пещерной рыбы Astyanax mexicanus ( Anopticichthys antrobius ) на запах известных и неизвестных сородичей». Эксперименты . 42 (9): 1053–1054. дои : 10.1007/bf01940729 . S2CID 29725205 .
- ^ Браун, GE; Смит, RJF (1994). «Толстоголовые гольяны используют химические сигналы, чтобы отличить естественных обитателей мелководья от незнакомых сородичей» . Журнал химической экологии . 20 (12): 3051–3061. Бибкод : 1994JCEco..20.3051B . дои : 10.1007/bf02033710 . ПМИД 24241976 . S2CID 31326304 .
- ^ Фермер, Северная Каролина; Риббл, DO; Миллер, III (2004). «Влияние знакомства на поведение мелководья в Техасе и чернохвостых фингеров». Журнал биологии рыб . 64 (3): 776–782. Бибкод : 2004JFBio..64..776F . дои : 10.1111/j.1095-8649.2004.00332.x .
- ^ Вебстер, ММ; Адамс, Эл.; Лаланд, КН (2008). «Химические сигналы, специфичные для диеты, влияют на предпочтения ассоциации и использование участков добычи у стайной рыбы». Поведение животных . 76 : 17–23. дои : 10.1016/j.anbehav.2007.12.010 . S2CID 53175064 .
- ^ Дугаткин, Л.А.; Фитцджеральд, Дж.Дж.; Лавуа, Дж. (1994). «Молодые трехиглые колюшки избегают паразитированных сородичей». Экологическая биология рыб . 39 (2): 215–218. Бибкод : 1994EnvBF..39..215D . дои : 10.1007/bf00004940 . S2CID 39806095 .
- ^ Краузе, Дж.; Годин (2010). «Влияние паразитизма на выбор стаи у полосатой рыбы-убийцы ( Fundulusdiaphanus , Teleostei, Cyprinodontidae)». Этология . 102 : 40–49. дои : 10.1111/j.1439-0310.1996.tb01102.x .
- ^ Барбер, И.; Дауни, округ Колумбия; Брейтуэйт, Вирджиния (1998). «Паразитизм, странность и механизм выбора косяков». Журнал биологии рыб . 53 (6): 1365–1368. дои : 10.1006/jfbi.1998.0788 .
- ^ Уорд, AJW; Дафф, Эй Джей; Краузе, Дж.; Барбер, И. (2005). «Стайное поведение колюшек, зараженных микроспоридийным паразитом Glutea anomala ». Экологическая биология рыб . 72 (2): 155–160. Бибкод : 2005EnvBF..72..155W . CiteSeerX 10.1.1.460.7259 . дои : 10.1007/s10641-004-9078-1 . S2CID 21197916 .
- ^ Краузе, Дж.; Хартманн, Н.; Причард, В.Л. (1999). «Влияние состояния питания на выбор косяка у рыбок данио Danio rerio ». Поведение животных . 57 (4): 771–775. дои : 10.1006/anbe.1998.1010 . ПМИД 10202085 . S2CID 25036232 .
- ^ Харкорт, JL; Свитман, Г.; Джонстон, РА; Маника, А. (2009). «Личность имеет значение: влияние смелости на выбор косяка у трехиглой колюшки». Поведение животных . 77 (6): 1501–1505. дои : 10.1016/j.anbehav.2009.03.004 . S2CID 53254313 .
- ^ Гомес-Лаплаза, LM (2005). «Влияние социального статуса на предпочтения мелководья пресноводных скалярий ( Pterophyllum scalare )». Поведение . 142 (6): 827–844. дои : 10.1163/1568539054729141 . S2CID 145533152 .
- ^ Меткалф, Северная Каролина; Томсон, Британская Колумбия (1995). «Рыба распознает и предпочитает скапливаться у плохих конкурентов». Труды Лондонского королевского общества Б. 259 (1355): 207–210. Бибкод : 1995РСПСБ.259..207М . дои : 10.1098/rspb.1995.0030 . S2CID 85131321 .
- ^ Питчер, Ти Джей; Хаус, AC (1987). «Правила кормления групповыми кормушками: копирование кормовой площади зависит от плотности пищи у стайных золотых рыбок». Этология . 76 (2): 161–167. дои : 10.1111/j.1439-0310.1987.tb00681.x .
- ^ Краузе, Дж (1992). «Идеальное бесплатное распространение и механизм оценки рентабельности пятен у трехиглой колюшки ( Gasterosteus aculeatus )». Поведение . 123 (1–2): 27–37. дои : 10.1163/156853992x00093 .
- ^ Рибс, С.Г.; Галлант, BY (1997). «Активность, связанная с ожиданием еды, как сигнал о местном усилении у золотых сияющих рыб (Рыбы: Cyprinidae, Notemigonusrysoleucas )». Этология . 103 (12): 1060–1069. Бибкод : 1997Этол.103.1060Р . дои : 10.1111/j.1439-0310.1997.tb00148.x . S2CID 84055118 .
- ^ Причард, В.Л.; Лоуренс, Дж.; Батлин, РК; Краузе, Дж. (2001). «Выбор косяка у рыбок данио, Danio rerio : влияние размера и активности косяка». Поведение животных . 62 (6): 1085–1088. дои : 10.1006/anbe.2001.1858 . S2CID 53165127 .
- ^ Домини, Уоллес Дж. (1983). «Моббинг среди колониально гнездящихся рыб, особенно синежабрников, Lepomis macrochirus ». Копейя . 1983 (4): 1086–1088. дои : 10.2307/1445113 . JSTOR 1445113 .
- ^ Фонтан, Генри (24 мая 2005 г.). «Краснобрюхая пиранья на самом деле желтая» . Нью-Йорк Таймс . Проверено 13 сентября 2014 г.
- ^ Гилли, ВФ; Маркайда, У.; Бакстер, Швейцария; Блок, бакалавр; Бустани, А.; Зейдберг, Л.; Райзенбихлер, К.; Робисон, Б.; Баззино, Г.; Салинас, К. (2006). «Вертикальные и горизонтальные миграции гигантского кальмара Dosidicus gigas, выявленные с помощью электронной маркировки» (PDF) . Серия «Прогресс в области морской экологии» . 324 : 1–17. Бибкод : 2006MEPS..324....1G . дои : 10.3354/meps324001 .
- ^ Циммерманн, Тим (июль 2006 г.). «Вот кальмар Гумбольдта» . За пределами Интернета . Проверено 13 сентября 2014 г.
- ^ «Загадочная история кальмара-каннибала - Национальная федерация дикой природы» . Проверено 13 сентября 2014 г.
- ^ Томас, Пит (26 марта 2007 г.). «Предупредительные огни моря» . Лос-Анджелес Таймс .
Дальнейшее чтение
[ редактировать ]- Бонабо, Э; Дагорн, Л. (1995). «Возможная универсальность распределения рыбных косяков по размерам» (PDF) . Физический обзор . 51 (6): R5220–R5223. Бибкод : 1995PhRvE..51.5220B . дои : 10.1103/physreve.51.r5220 . ПМИД 9963400 .
- Боински С. и Гарбер П.А. (2000) В движении: как и почему животные путешествуют группами, University of Chicago Press. ISBN 978-0-226-06339-3
- Бредер, CM (1954). «Уравнения, описывающие косяки рыб и другие скопления животных». Экология . 35 (3): 361–370. Бибкод : 1954Экол...35..361Б . дои : 10.2307/1930099 . JSTOR 1930099 .
- Чилдресс С. (1981) Механика плавания и полета Издательство Кембриджского университета. ISBN 978-0-521-28071-6
- Камазин С., Денебур Дж.Л., Фрэнкс Н.Р. , Снейд Дж., Тераулаз Г. и Бонабо Э. (2003) Самоорганизация в биологических системах. Издательство Принстонского университета. ISBN 978-0-691-11624-2 - особенно глава 11
- Эванс, СР; Финния, М; Маника, А (2007). «Мелодые предпочтения десятиногих ракообразных». Поведение животных . 74 (6): 1691–1696. дои : 10.1016/j.anbehav.2007.03.017 . S2CID 53150496 .
- Делькур, Дж; Понсин, П. (2012). «Косяки и косяки: вернемся к эвристическим определениям и количественным ориентирам». Обзоры по биологии рыб и рыболовству . 22 (3): 595–619. Бибкод : 2012RFBF...22..595D . дои : 10.1007/s11160-012-9260-z . S2CID 18306602 .
- Готре Дж., Йост К. и Тераулаз Г. (2008) Ключевые поведенческие факторы в модели самоорганизующегося косяка рыб. Annales Zoologici Fennici 45: 415–428.
- Годин, Дж. Дж. (1997) Поведенческая экология костистых рыб, издательство Оксфордского университета. ISBN 978-0-19-850503-7
- Гош С. и Рамамурти К.В. (2004) Проектирование сетевых систем информационных технологий Springer. ISBN 978-0-387-95544-5
- Хагер, MC; Хельфман, Г.С. (1991). «Безопасность в численности: выбор размера косяка гольянами в условиях угрозы хищников». Поведенческая экология и социобиология . 29 (4): 271–276. дои : 10.1007/BF00163984 . S2CID 30901973 .
- Хемельрейк, СК; Хильденбрандт, Х; Рейндерс, Дж; Стамхейс, Э.Дж. (2010). «Появление продолговатой формы косяка: модели и эмпирические данные рыб» (PDF) . Этология . 116 (11): 1–14. Бибкод : 2010Ethol.116.1099H . дои : 10.1111/j.1439-0310.2010.01818.x .
- Хоар, диджей; Краузе, Дж (2003). «Социальная организация, структура косяка и передача информации». Рыба и рыболовство . 4 (3): 269–279. Бибкод : 2003AqFF....4..269H . дои : 10.1046/j.1467-2979.2003.00130.x .
- Инада Ю (2001) «Механизм управления рыбными косяками» Complexity International удостоверения личности , Том 8, Загрузка
- Инагаки, Т; Сакамото, В; Аоки, я (1976). «Исследования стайного поведения рыб - III Взаимосвязь между скоростью и формой стайного поведения» . Бюллетень Японского общества научного рыболовства . 42 (6): 629–635. дои : 10.2331/suisan.42.629 .
- Като Н. и Айерс Дж. (2004) Биомеханизмы плавания и полета спрингера. ISBN 978-4-431-22211-8
- Кеннеди Дж., Эберхарт Р.К. и Ши Ю. (2001) Роевой интеллект Морган Кауфманн. ISBN 978-1-55860-595-4
- Краузе, Дж. (2005) Жизнь в группах Oxford University Press. ISBN 978-0-19-850818-2
- Краузе, Дж (2005). «Позиционирование рыбных косяков: анализ затрат и выгод» . Журнал биологии рыб . 43 : 309–314. дои : 10.1111/j.1095-8649.1993.tb01194.x . Архивировано из оригинала 5 января 2013 года.
- Краузе, Дж; Ракстон, Джорджия; Рубинштейн, Д. (2005). «Всегда ли размер косяка влияет на успех охоты хищника?» . Журнал биологии рыб . 52 (3): 494–501. дои : 10.1111/j.1095-8649.1998.tb02012.x . [ мертвая ссылка ]
- Литвак, МК (1993). «Реакция стайной рыбы на угрозу воздушного нападения». Экологическая биология рыб . 36 (2): 183–192. Бибкод : 1993EnvBF..36..183L . дои : 10.1007/BF00002798 . S2CID 30214279 .
- Луртон X (2003) Подводная акустика Спрингер. ISBN 978-3-540-42967-8
- Мойл П.Б. и Ван Дайк К.М. (1995) Рыба: Путеводитель для энтузиастов Издательство Калифорнийского университета. ISBN 978-0-520-20165-1
- Пэрриш Дж.К. и Хамнер В.М. (1997) Группы животных в трех измерениях: как виды объединяются Издательство Кембриджского университета. ISBN 978-0-521-46024-8
- Пэрриш, Дж. К. ; Вишидо, СВ; Грюнбаумб, Д. (2002). «Самоорганизованные рыбные косяки: исследование новых свойств» (PDF) . Биол. Бык . 202 (3): 296–305. CiteSeerX 10.1.1.116.1548 . дои : 10.2307/1543482 . JSTOR 1543482 . ПМИД 12087003 . S2CID 377484 .
- Партридж, Б.Л. (1982). «Структура и функции рыбных косяков» (PDF) . Научный американец . Том. 246, нет. 6. С. 114–123. Бибкод : 1982SciAm.246f.114P . doi : 10.1038/scientificamerican0682-114 . ПМИД 7201674 . Архивировано из оригинала (PDF) 3 июля 2011 года.
- Питчер, Ти Джей (1983). «Эвристические определения поведения рыбы при стайке». Поведение животных . 31 (2): 611–613. дои : 10.1016/s0003-3472(83)80087-6 . S2CID 53195091 .
- Питчер Т.Дж. и Пэриш Дж.К. (1993) «Функции стайного поведения костистых рыб» В: Питчер Т.Дж. (ред.) Поведение костистых рыб . Чепмен и Холл, Нью-Йорк, стр. 363–440.
- Питчер, Ти Джей ; Магурран, А.Е.; Уинфилд, Эй-Джей (1982). «Рыба в больших косяках быстрее находит пищу». Поведенческая экология и социобиология . 10 (2): 149–151. дои : 10.1007/BF00300175 . S2CID 6340986 .
- Питчер Т.Дж. (2010) «Обучение рыб» В: Стил Дж.Х., Торп С.А. и Турекян К.К. (ред.) Морская биология, Academic Press, страницы 337–349. ISBN 978-0-08-096480-5 .
- Прайор К. и Норрис К.С. (1998) Общества дельфинов: открытия и загадки Издательство Калифорнийского университета. ISBN 0-520-21656-3 , ISBN 978-0-520-21656-3
- Росс Д.А. (2000) Книги о рыбацких океанских столбах. ISBN 978-0-8117-2771-6
- Скалабрин, К; Массе, Дж (1993). «Акустическое обнаружение пространственного и временного распределения рыбных косяков в Бискайском заливе» . Водные живые ресурсы . 6 (3): 269–283. Бибкод : 1993AqLR....6..269S . дои : 10.1051/алр:1993027 .
- Сено, Х; Накаи, К. (1995). «Математический анализ обмеления рыбы с помощью модели диффузии, зависящей от плотности». Экологическое моделирование . 79 (3): 149–157. Бибкод : 1995EcMod..79..149S . дои : 10.1016/0304-3800(93)E0143-Q .
- Симмондс Э.Дж. и МакЛеннан, Д.Н. (2005) Издательство Fisheries Acoustics Blackwell Publishing. ISBN 978-0-632-05994-2
- Суппи Р., Фернандес Д. и Люке Э. (2003) Рыбные школы: моделирование PDES и 3D-анимация в реальном времени в параллельной обработке и прикладной математике: 5-я Международная конференция, PPAM 2003, Springer. ISBN 978-3-540-21946-0
- Вичек, А; Зафейрис, А (2012). «Коллективное движение». Отчеты по физике . 517 (3–4): 71–140. arXiv : 1010.5017 . Бибкод : 2012ФР...517...71В . дои : 10.1016/j.physrep.2012.03.004 . S2CID 119109873 .
- Белый TI (2007) В защиту дельфинов Blackwell Publishing. ISBN 978-1-4051-5779-7
- Вольф, Н.Г. (1985). «Странные рыбы покидают смешанные группы видов, когда им угрожают». Поведенческая экология и социобиология . 17 (1): 47–52. дои : 10.1007/bf00299428 . S2CID 11935938 .
- Вуттон, Р.Дж. (1998) Экология костистых рыб Спрингер. ISBN 978-0-412-64200-5
Внешние ссылки
[ редактировать ]- Веб-сайт «Коллективное поведение животных», организованный на основе одноименной книги Дэвида Самптера (2008).
- Проект STARFLAG: Описание проекта по отлову скворцов
- Центр биологического дизайна Технологического института Джорджии
- Исследовательский сайт Дэвида Самптера
- Исследовательский сайт Иэна Кузена
- Веб-сайт Джулии Пэрриш, исследователя агрегации животных
- Программа исследований пелагического рыболовства (2002 г.) Текущее состояние и новые направления изучения стайного и агрегационного поведения пелагических рыб.
- Клевер, Чарльз (2008) Фиш умеет считать до четырех, но не выше Telegraph Media Group.
- Миграционное поведение сельди
- Пример моделирования школьного обучения
- Бхадури, Апарна (2010) Обучение в рыбном колледже OpenStax . Обновлено 16 июля 2010 г.
| О рыбе |  | ||||
|---|---|---|---|---|---|
| Анатомия и физиология | |||||
| сенсорный системы | |||||
| Воспроизведение | |||||
| Передвижение | |||||
| Другой поведение | |||||
| По среде обитания | |||||
| Другие типы | |||||
| Коммерция |
| ||||
| Основные группы | |||||
| Списки | |||||

о рыболовстве Темы науки |
|---|

