Поведение роя

Стайное поведение , или роение , — это коллективное поведение, демонстрируемое существами, особенно животными, одинакового размера, которые собираются вместе, возможно, слоняясь в одном и том же месте или, возможно, перемещаясь в массе или мигрируя в каком-то направлении. Это очень междисциплинарная тема. [1]
Термин « роение» применяется, в частности, к насекомым, но его также можно применять к любому другому объекту или животному, проявляющему поведение стаи. Термин «стайное» или «шумение» может относиться конкретно к стаевому поведению птиц, стадному поведению четвероногих , а стайному поведению или стайному поведению рыб. Фитопланктон также собирается в огромные стаи, называемые цветками , хотя эти организмы являются водорослями и не обладают самодвижением, как животные. В более широком смысле термин «рой» применяется также к неодушевленным объектам, которые демонстрируют параллельное поведение, например, рой роботов , рой землетрясений или рой звезд.
From a more abstract point of view, swarm behaviour is the collective motion of a large number of self-propelled entities.[2] From the perspective of the mathematical modeller, it is an emergent behaviour arising from simple rules that are followed by individuals and does not involve any central coordination. Swarm behaviour is also studied by active matter physicists as a phenomenon which is not in thermodynamic equilibrium, and as such requires the development of tools beyond those available from the statistical physics of systems in thermodynamic equilibrium. In this regard, swarming has been compared to the mathematics of superfluids, specifically in the context of starling flocks (murmuration).[3]
Swarm behaviour was first simulated on a computer in 1986 with the simulation program boids.[4] This program simulates simple agents (boids) that are allowed to move according to a set of basic rules. The model was originally designed to mimic the flocking behaviour of birds, but it can be applied also to schooling fish and other swarming entities.
Models
[edit]In recent decades, scientists have turned to modeling swarm behaviour to gain a deeper understanding of the behaviour.
Mathematical models
[edit]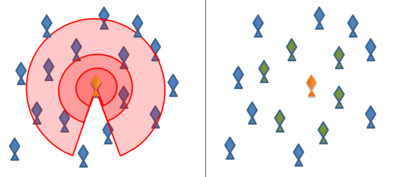
| External images | |
|---|---|
Early studies of swarm behaviour employed mathematical models to simulate and understand the behaviour. The simplest mathematical models of animal swarms generally represent individual animals as following three rules:
- Move in the same direction as their neighbours
- Remain close to their neighbours
- Avoid collisions with their neighbours
The boids computer program, created by Craig Reynolds in 1986, simulates swarm behaviour following the above rules.[4] Many subsequent and current models use variations on these rules, often implementing them by means of concentric "zones" around each animal. In the "zone of repulsion", very close to the animal, the focal animal will seek to distance itself from its neighbours to avoid collision. Slightly further away, in the "zone of alignment", the focal animal will seek to align its direction of motion with its neighbours. In the outermost "zone of attraction", which extends as far away from the focal animal as it is able to sense, the focal animal will seek to move towards a neighbour.
The shape of these zones will necessarily be affected by the sensory capabilities of a given animal. For example, the visual field of a bird does not extend behind its body. Fish rely on both vision and on hydrodynamic perceptions relayed through their lateral lines, while Antarctic krill rely both on vision and hydrodynamic signals relayed through antennae.
However recent studies of starling flocks have shown that each bird modifies its position, relative to the six or seven animals directly surrounding it, no matter how close or how far away those animals are.[5] Interactions between flocking starlings are thus based on a topological, rather than a metric, rule. It remains to be seen whether this applies to other animals. Another recent study, based on an analysis of high-speed camera footage of flocks above Rome and assuming minimal behavioural rules, has convincingly simulated a number of aspects of flock behaviour.[6][7][8][9]
Evolutionary models
[edit]In order to gain insight into why animals evolve swarming behaviours, scientists have turned to evolutionary models that simulate populations of evolving animals. Typically these studies use a genetic algorithm to simulate evolution over many generations. These studies have investigated a number of hypotheses attempting to explain why animals evolve swarming behaviours, such as the selfish herd theory[10][11][12][13][14] the predator confusion effect,[15][16] the dilution effect,[17][18] and the many eyes theory.[19]
Agents
[edit]- Mach, Robert; Schweitzer, Frank (2003). "Multi-Agent Model of Biological Swarming". Advances In Artificial Life. Lecture Notes in Computer Science. Vol. 2801. pp. 810–820. CiteSeerX 10.1.1.87.8022. doi:10.1007/978-3-540-39432-7_87. ISBN 978-3-540-20057-4.
Self-organization
[edit]
Emergence
[edit]The concept of emergence—that the properties and functions found at a hierarchical level are not present and are irrelevant at the lower levels–is often a basic principle behind self-organizing systems.[20] An example of self-organization in biology leading to emergence in the natural world occurs in ant colonies. The queen does not give direct orders and does not tell the ants what to do.[citation needed] Instead, each ant reacts to stimuli in the form of chemical scents from larvae, other ants, intruders, food and buildup of waste, and leaves behind a chemical trail, which, in turn, provides a stimulus to other ants. Here each ant is an autonomous unit that reacts depending only on its local environment and the genetically encoded rules for its variety. Despite the lack of centralized decision making, ant colonies exhibit complex behaviours and have even been able to demonstrate the ability to solve geometric problems. For example, colonies routinely find the maximum distance from all colony entrances to dispose of dead bodies.
Stigmergy
[edit]A further key concept in the field of swarm intelligence is stigmergy.[21][22] Stigmergy is a mechanism of indirect coordination between agents or actions. The principle is that the trace left in the environment by an action stimulates the performance of a next action, by the same or a different agent. In that way, subsequent actions tend to reinforce and build on each other, leading to the spontaneous emergence of coherent, apparently systematic activity. Stigmergy is a form of self-organization. It produces complex, seemingly intelligent structures, without need for any planning, control, or even direct communication between the agents. As such it supports efficient collaboration between extremely simple agents, who lack any memory, intelligence or even awareness of each other.[22]
Swarm intelligence
[edit]Swarm intelligence is the collective behaviour of decentralized, self-organized systems, natural or artificial. The concept is employed in work on artificial intelligence. The expression was introduced by Gerardo Beni and Jing Wang in 1989, in the context of cellular robotic systems.[23]
Swarm intelligence systems are typically made up of a population of simple agents such as boids interacting locally with one another and with their environment. The agents follow very simple rules, and although there is no centralized control structure dictating how individual agents should behave, local, and to a certain degree random, interactions between such agents lead to the emergence of intelligent global behaviour, unknown to the individual agents.
Swarm intelligence research is multidisciplinary. It can be divided into natural swarm research studying biological systems and artificial swarm research studying human artefacts. There is also a scientific stream attempting to model the swarm systems themselves and understand their underlying mechanisms, and an engineering stream focused on applying the insights developed by the scientific stream to solve practical problems in other areas.[24]
Algorithms
[edit]Swarm algorithms follow a Lagrangian approach or an Eulerian approach.[25] The Eulerian approach views the swarm as a field, working with the density of the swarm and deriving mean field properties. It is a hydrodynamic approach, and can be useful for modelling the overall dynamics of large swarms.[26][27][28] However, most models work with the Lagrangian approach, which is an agent-based model following the individual agents (points or particles) that make up the swarm. Individual particle models can follow information on heading and spacing that is lost in the Eulerian approach.[25][29]
Ant colony optimization
[edit]| External image | |
|---|---|
Ant colony optimization is a widely used algorithm which was inspired by the behaviours of ants, and has been effective solving discrete optimization problems related to swarming.[31] The algorithm was initially proposed by Marco Dorigo in 1992,[32][33] and has since been diversified to solve a wider class of numerical problems. Species that have multiple queens may have a queen leaving the nest along with some workers to found a colony at a new site, a process akin to swarming in honeybees.[34][35]
- Ants are behaviourally unsophisticated; collectively they perform complex tasks. Ants have highly developed sophisticated sign-based communication.
- Ants communicate using pheromones; trails are laid that can be followed by other ants.
- Routing problem ants drop different pheromones used to compute the "shortest" path from source to destination(s).
- Rauch, EM; Millonas, MM; Chialvo, DR (1995). "Pattern formation and functionality in swarm models". Physics Letters A. 207 (3–4): 185. arXiv:adap-org/9507003. Bibcode:1995PhLA..207..185R. doi:10.1016/0375-9601(95)00624-c. S2CID 120567147.
Self-propelled particles
[edit]| External videos | |
|---|---|
– needs Java |
The concept of self-propelled particles (SPP) was introduced in 1995 by Tamás Vicsek et al.[37] as a special case of the boids model introduced in 1986 by Reynolds.[4] An SPP swarm is modelled by a collection of particles that move with a constant speed and respond to random perturbations by adopting at each time increment the average direction of motion of the other particles in their local neighbourhood.[38]
Simulations demonstrate that a suitable "nearest neighbour rule" eventually results in all the particles swarming together, or moving in the same direction. This emerges, even though there is no centralized coordination, and even though the neighbours for each particle constantly change over time.[37] SPP models predict that swarming animals share certain properties at the group level, regardless of the type of animals in the swarm.[39] Swarming systems give rise to emergent behaviours which occur at many different scales, some of which are both universal and robust. It has become a challenge in theoretical physics to find minimal statistical models that capture these behaviours.[40][41]
Particle swarm optimization
[edit]Particle swarm optimization is another algorithm widely used to solve problems related to swarms. It was developed in 1995 by Kennedy and Eberhart and was first aimed at simulating the social behaviour and choreography of bird flocks and fish schools.[42][43] The algorithm was simplified and it was observed to be performing optimization. The system initially seeds a population with random solutions. It then searches in the problem space through successive generations using stochastic optimization to find the best solutions. The solutions it finds are called particles. Each particle stores its position as well as the best solution it has achieved so far. The particle swarm optimizer tracks the best local value obtained so far by any particle in the local neighbourhood. The remaining particles then move through the problem space following the lead of the optimum particles. At each time iteration, the particle swarm optimiser accelerates each particle toward its optimum locations according to simple mathematical rules. Particle swarm optimization has been applied in many areas. It has few parameters to adjust, and a version that works well for a specific applications can also work well with minor modifications across a range of related applications.[44] A book by Kennedy and Eberhart describes some philosophical aspects of particle swarm optimization applications and swarm intelligence.[45] An extensive survey of applications is made by Poli.[46][47]
Altruism
[edit]Researchers in Switzerland have developed an algorithm based on Hamilton's rule of kin selection. The algorithm shows how altruism in a swarm of entities can, over time, evolve and result in more effective swarm behaviour.[48][49]
Biological swarming
[edit]
The earliest evidence of swarm behaviour in animals dates back about 480 million years. Fossils of the trilobite Ampyx priscus have been recently described as clustered in lines along the ocean floor. The animals were all mature adults, and were all facing the same direction as though they had formed a conga line or a peloton. It has been suggested they line up in this manner to migrate, much as spiny lobsters migrate in single-file queues;[50] it has also been suggested that the formation is the precursor for mating,[51] as with the fly Leptoconops torrens. The findings suggest animal collective behaviour has very early evolutionary origins.[52]
Examples of biological swarming are found in bird flocks,[53] fish schools,[54][55] insect swarms,[56] bacteria swarms,[57][58] molds,[59] molecular motors,[60] quadruped herds[61] and people.[62][63][64][65]
Social insects
[edit]The behaviour of social insects (insects that live in colonies, such as ants, bees, wasps and termites) has always been a source of fascination for children, naturalists and artists. Individual insects seem to do their own thing without any central control, yet the colony as a whole behaves in a highly coordinated manner.[66] Researchers have found that cooperation at the colony level is largely self-organized. The group coordination that emerges is often just a consequence of the way individuals in the colony interact. These interactions can be remarkably simple, such as one ant merely following the trail left by another ant. Yet put together, the cumulative effect of such behaviours can solve highly complex problems, such as locating the shortest route in a network of possible paths to a food source. The organised behaviour that emerges in this way is sometimes called swarm intelligence, a form of biological emergence.[66]
Ants
[edit]Individual ants do not exhibit complex behaviours, yet a colony of ants collectively achieves complex tasks such as constructing nests, taking care of their young, building bridges and foraging for food. A colony of ants can collectively select (i.e. send most workers towards) the best, or closest, food source from several in the vicinity.[67] Such collective decisions are achieved using positive feedback mechanisms. Selection of the best food source is achieved by ants following two simple rules. First, ants which find food return to the nest depositing a pheromone chemical. More pheromone is laid for higher quality food sources.[68] Thus, if two equidistant food sources of different qualities are found simultaneously, the pheromone trail to the better one will be stronger. Ants in the nest follow another simple rule, to favor stronger trails, on average. More ants then follow the stronger trail, so more ants arrive at the high quality food source, and a positive feedback cycle ensures, resulting in a collective decision for the best food source. If there are two paths from the ant nest to a food source, then the colony usually selects the shorter path. This is because the ants that first return to the nest from the food source are more likely to be those that took the shorter path. More ants then retrace the shorter path, reinforcing the pheromone trail.[69]
Army ants, unlike most ant species, do not construct permanent nests; an army ant colony moves almost incessantly over the time it exists, remaining in an essentially perpetual state of swarming. Several lineages have independently evolved the same basic behavioural and ecological syndrome, often referred to as "legionary behaviour", and may be an example of convergent evolution.[70]
The successful techniques used by ant colonies have been studied in computer science and robotics to produce distributed and fault-tolerant systems for solving problems. This area of biomimetics has led to studies of ant locomotion, search engines that make use of "foraging trails", fault-tolerant storage and networking algorithms.[71]
Honey bees
[edit]
In temperate climates, honey bees usually form swarms in late spring. A swarm typically contains about half the workers together with the old queen, while the new queen stays back with the remaining workers in the original hive. When honey bees emerge from a hive to form a swarm, they may gather on a branch of a tree or on a bush only a few meters from the hive. The bees cluster about the queen and send out 20–50 scouts to find suitable new nest locations. The scouts are the most experienced foragers in the cluster. If a scout finds a suitable location, she returns to the cluster and promotes it by dancing a version of the waggle dance. This dance conveys information about the quality, direction, and distance of the new site. The more excited she is about her findings, the more vigorously she dances. If she can convince others they may take off and check the site she found. If they approve they may promote it as well. In this decision-making process, scouts check several sites, often abandoning their own original site to promote the superior site of another scout. Several different sites may be promoted by different scouts at first. After some hours and sometimes days, a preferred location eventually emerges from this decision-making process. When all scouts agree on the final location, the whole cluster takes off and swarms to it. Sometimes, if no decision is reached, the swarm will separate, some bees going in one direction; others, going in another. This usually results in failure, with both groups dying. A new location is typically a kilometre or more from the original hive, though some species, e.g., Apis dorsata,[72] may establish new colonies within as little as 500 meters from the natal nest. This collective decision-making process is remarkably successful in identifying the most suitable new nest site and keeping the swarm intact. A good hive site has to be large enough to accommodate the swarm (about 15 litres in volume), has to be well-protected from the elements, receive an optimal amount of sunshine, be some height above the ground, have a small entrance and be capable of resisting ant infestation - that is why tree cavities are often selected.[73][74][75][76][77]
Non-social insects
[edit]Unlike social insects, swarms of non-social insects that have been studied primarily seem to function in contexts such as mating, feeding, predator avoidance, and migration.
Moths
[edit]Moths may exhibit synchronized mating, during which pheromones released by females initiate searching and swarming behavior in males.[78] Males sense pheromones with sensitive antennae and may track females as far as several kilometers away.[79] Swarm mating involves female choice and male competition. Only one male in the swarm—typically the first—will successfully copulate.[80] Females maximize fitness benefits and minimize cost by governing the onset and magnitude of pheromone deployed. Too little pheromone will not attract a mate, too much allows less fit males to sense the signal.[81] After copulation, females lay the eggs on a host plant. Quality of host plant may be a factor influencing the location of swarming and egg-laying. In one case, researchers observed pink-striped oakworm moths (Anisota virginiensis) swarming at a carrion site, where decomposition likely increased soil nutrient levels and host plant quality.[82]
Flies
[edit]Midges, such as Tokunagayusurika akamusi, form swarms, dancing in the air. Swarming serves multiple purposes, including the facilitation of mating by attracting females to approach the swarm, a phenomenon known as lek mating. Such cloud-like swarms often form in early evening when the sun is getting low, at the tip of a bush, on a hilltop, over a pool of water, or even sometimes above a person. The forming of such swarms is not out of instinct, but an adaptive behavior – a "consensus" – between the individuals within the swarms. It is also suggested that swarming is a ritual, because there is rarely any male midge by itself and not in a swarm. This could have formed due to the benefit of lowering inbreeding by having males of various genes gathering in one spot.[83] The genus Culicoides, also known as biting midges, have displayed swarming behavior which are believed to cause confusion in predators.[84]
Cockroaches
[edit]Cockroaches leave chemical trails in their feces as well as emitting airborne pheromones for mating. Other cockroaches will follow these trails to discover sources of food and water, and also discover where other cockroaches are hiding. Thus, groups of cockroaches can exhibit emergent behaviour,[85] in which group or swarm behaviour emerges from a simple set of individual interactions.
Cockroaches are mainly nocturnal and will run away when exposed to light. A study tested the hypothesis that cockroaches use just two pieces of information to decide where to go under those conditions: how dark it is and how many other cockroaches there are. The study conducted by José Halloy and colleagues at the Free University of Brussels and other European institutions created a set of tiny robots that appear to the roaches as other roaches and can thus alter the roaches' perception of critical mass. The robots were also specially scented so that they would be accepted by the real roaches.[86]
Locusts
[edit]
Locusts are the swarming phase of the short-horned grasshoppers of the family Acrididae. Some species can breed rapidly under suitable conditions and subsequently become gregarious and migratory. They form bands as nymphs and swarms as adults—both of which can travel great distances, rapidly stripping fields and greatly damaging crops. The largest swarms can cover hundreds of square miles and contain billions of locusts. A locust can eat its own weight (about 2 grams) in plants every day. That means one million locusts can eat more than one tonne of food each day, and the largest swarms can consume over 100,000 tonnes each day.[87]
Swarming in locusts has been found to be associated with increased levels of serotonin which causes the locust to change colour, eat much more, become mutually attracted, and breed much more easily. Researchers propose that swarming behaviour is a response to overcrowding and studies have shown that increased tactile stimulation of the hind legs or, in some species, simply encountering other individuals causes an increase in levels of serotonin. The transformation of the locust to the swarming variety can be induced by several contacts per minute over a four-hour period.[88][89][90][91] Notably, an innate predisposition to aggregate has been found in hatchlings of the desert locust, Schistocerca gregaria, independent of their parental phase.[92]
An individual locust's response to a loss of alignment in the group appears to increase the randomness of its motion, until an aligned state is again achieved. This noise-induced alignment appears to be an intrinsic characteristic of collective coherent motion.[93]
Migratory behavior
[edit]
Insect migration is the seasonal movement of insects, particularly those by species of dragonflies, beetles, butterflies, and moths. The distance can vary from species to species, but in most cases these movements involve large numbers of individuals. In some cases the individuals that migrate in one direction may not return and the next generation may instead migrate in the opposite direction. This is a significant difference from bird migration.
Monarch butterflies are especially noted for their lengthy annual migration. In North America they make massive southward migrations starting in August until the first frost. A northward migration takes place in the spring. The monarch is the only butterfly that migrates both north and south as the birds do on a regular basis. But no single individual makes the entire round trip. Female monarchs deposit eggs for the next generation during these migrations.[94] The length of these journeys exceeds the normal lifespan of most monarchs, which is less than two months for butterflies born in early summer. The last generation of the summer enters into a non-reproductive phase known as diapause and may live seven months or more.[95] During diapause, butterflies fly to one of many overwintering sites. The generation that overwinters generally does not reproduce until it leaves the overwintering site sometime in February and March. It is the second, third and fourth generations that return to their northern locations in the United States and Canada in the spring. How the species manages to return to the same overwintering spots over a gap of several generations is still a subject of research; the flight patterns appear to be inherited, based on a combination of the position of the sun in the sky[96] and a time-compensated Sun compass that depends upon a circadian clock that is based in their antennae.[97][98]
Birds
[edit]
 Murmurations of starlings
Murmurations of starlings Bird migration
[edit]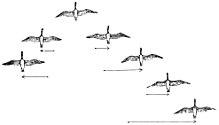
Approximately 1800 of the world's 10,000 bird species are long-distance migrants.[100] The primary motivation for migration appears to be food; for example, some hummingbirds choose not to migrate if fed through the winter. Also, the longer days of the northern summer provide extended time for breeding birds to feed their young. This helps diurnal birds to produce larger clutches than related non-migratory species that remain in the tropics. As the days shorten in autumn, the birds return to warmer regions where the available food supply varies little with the season. These advantages offset the high stress, physical exertion costs, and other risks of the migration such as predation.
Many birds migrate in flocks. For larger birds, it is assumed that flying in flocks reduces energy costs. The V formation is often supposed to boost the efficiency and range of flying birds, particularly over long migratory routes. All the birds except the first fly in the upwash from one of the wingtip vortices of the bird ahead. The upwash assists each bird in supporting its own weight in flight, in the same way a glider can climb or maintain height indefinitely in rising air. Geese flying in a V formation save energy by flying in the updraft of the wingtip vortex generated by the previous animal in the formation. Thus, the birds flying behind do not need to work as hard to achieve lift. Studies show that birds in a V formation place themselves roughly at the optimum distance predicted by simple aerodynamic theory.[101] Geese in a V-formation may conserve 12–20% of the energy they would need to fly alone.[102][103] Red knots and dunlins were found in radar studies to fly 5 km per hour faster in flocks than when they were flying alone.[104] The birds flying at the tips and at the front are rotated in a timely cyclical fashion to spread flight fatigue equally among the flock members. The formation also makes communication easier and allows the birds to maintain visual contact with each other.
| External videos | |
|---|---|
– from The Trials of Life |
Other animals may use similar drafting techniques when migrating. Lobsters, for example, migrate in close single-file formation "lobster trains", sometimes for hundreds of miles.
The Mediterranean and other seas present a major obstacle to soaring birds, which must cross at the narrowest points. Massive numbers of large raptors and storks pass through areas such as Gibraltar, Falsterbo, and the Bosphorus at migration times. More common species, such as the European honey buzzard, can be counted in hundreds of thousands in autumn. Other barriers, such as mountain ranges, can also cause funnelling, particularly of large diurnal migrants. This is a notable factor in the Central American migratory bottleneck. This concentration of birds during migration can put species at risk. Some spectacular migrants have already gone extinct, the most notable being the passenger pigeon. During migration the flocks were a mile (1.6 km) wide and 300 miles (500 km) long, taking several days to pass and containing up to a billion birds.
Marine life
[edit]Fish
[edit]
| External image | |
|---|---|
The term "shoal" can be used to describe any group of fish, including mixed-species groups, while "school" is used for more closely knit groups of the same species swimming in a highly synchronised and polarised manner.
Fish derive many benefits from shoaling behaviour including defence against predators (through better predator detection and by diluting the chance of capture), enhanced foraging success, and higher success in finding a mate.[106] It is also likely that fish benefit from shoal membership through increased hydrodynamic efficiency.[107]
Fish use many traits to choose shoalmates. Generally they prefer larger shoals, shoalmates of their own species, shoalmates similar in size and appearance to themselves, healthy fish, and kin (when recognised). The "oddity effect" posits that any shoal member that stands out in appearance will be preferentially targeted by predators. This may explain why fish prefer to shoal with individuals that resemble them. The oddity effect would thus tend to homogenise shoals.[108]
One puzzling aspect of shoal selection is how a fish can choose to join a shoal of animals similar to themselves, given that it cannot know its own appearance. Experiments with zebrafish have shown that shoal preference is a learned ability, not innate. A zebrafish tends to associate with shoals that resemble shoals in which it was reared, a form of imprinting.[109]
Other open questions of shoaling behaviour include identifying which individuals are responsible for the direction of shoal movement. In the case of migratory movement, most members of a shoal seem to know where they are going. In the case of foraging behaviour, captive shoals of golden shiner (a kind of minnow) are led by a small number of experienced individuals who knew when and where food was available.[110]
Radakov estimated herring schools in the North Atlantic can occupy up to 4.8 cubic kilometres (1.2 cu mi) with fish densities between 0.5 and 1.0 fish/cubic metre, totalling several billion fish in one school.[111]
- Partridge BL (1982) "The structure and function of fish schools" Scientific American, June:114–123.
- Parrish JK, Viscido SV, Grunbaum D (2002). "Self-Organized Fish Schools: An Examination of Emergent Properties" (PDF). Biol. Bull. 202 (3): 296–305. CiteSeerX 10.1.1.116.1548. doi:10.2307/1543482. JSTOR 1543482. PMID 12087003. S2CID 377484.[permanent dead link]
Fish migration
[edit]| External image | |
|---|---|
Between May and July huge numbers of sardines spawn in the cool waters of the Agulhas Bank and then follow a current of cold water northward along the east coast of South Africa. This great migration, called the sardine run, creates spectacular feeding frenzies along the coastline as marine predators, such as dolphins, sharks and gannets attack the schools.
Krill
[edit]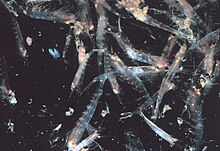
Most krill, small shrimp-like crustaceans, form large swarms, sometimes reaching densities of 10,000–60,000 individual animals per cubic metre.[113][114][115] Swarming is a defensive mechanism, confusing smaller predators that would like to pick out single individuals. The largest swarms are visible from space and can be tracked by satellite.[116] One swarm was observed to cover an area of 450 square kilometres (175 square miles) of ocean, to a depth of 200 meters (650 feet) and was estimated to contain over 2 million tons of krill.[117] Recent research suggests that krill do not simply drift passively in these currents but actually modify them.[117] Krill typically follow a diurnal vertical migration. By moving vertically through the ocean on a 12-hour cycle, the swarms play a major part in mixing deeper, nutrient-rich water with nutrient-poor water at the surface.[117] Until recently it has been assumed that they spend the day at greater depths and rise during the night toward the surface. It has been found that the deeper they go, the more they reduce their activity,[118] apparently to reduce encounters with predators and to conserve energy.
Later work suggested that swimming activity in krill varied with stomach fullness. Satiated animals that had been feeding at the surface swim less actively and therefore sink below the mixed layer.[119] As they sink they produce faeces which may mean that they have an important role to play in the Antarctic carbon cycle. Krill with empty stomachs were found to swim more actively and thus head towards the surface. This implies that vertical migration may be a bi- or tri-daily occurrence. Some species form surface swarms during the day for feeding and reproductive purposes even though such behaviour is dangerous because it makes them extremely vulnerable to predators.[120] Dense swarms may elicit a feeding frenzy among fish, birds and mammal predators, especially near the surface. When disturbed, a swarm scatters, and some individuals have even been observed to moult instantaneously, leaving the exuvia behind as a decoy.[121] In 2012, Gandomi and Alavi presented what appears to be a successful stochastic algorithm for modelling the behaviour of krill swarms. The algorithm is based on three main factors: " (i) movement induced by the presence of other individuals (ii) foraging activity, and (iii) random diffusion."[122]
Copepods
[edit]
Copepods are a group of tiny crustaceans found in the sea and lakes. Many species are planktonic (drifting in sea waters), and others are benthic (living on the ocean floor). Copepods are typically 1 to 2 millimetres (0.04 to 0.08 in) long, with a teardrop shaped body and large antennae. Although like other crustaceans they have an armoured exoskeleton, they are so small that in most species this thin armour, and the entire body, is almost totally transparent. Copepods have a compound, median single eye, usually bright red, in the centre of the transparent head.
Copepods also swarm. For example, monospecific swarms have been observed regularly around coral reefs and sea grass, and in lakes. Swarms densities were about one million copepods per cubic metre. Typical swarms were one or two metres in diameter, but some exceeded 30 cubic metres. Copepods need visual contact to keep together, and they disperse at night.[123]
Spring produces blooms of swarming phytoplankton which provide food for copepods. Planktonic copepods are usually the dominant members of the zooplankton, and are in turn major food organisms for many other marine animals. In particular, copepods are prey to forage fish and jellyfish, both of which can assemble in vast, million-strong swarms. Some copepods have extremely fast escape responses when a predator is sensed and can jump with high speed over a few millimetres (see animated image below).
 На фото: Стая сельдей, питающаяся стаей веслоногих ракообразных.
На фото: Стая сельдей, питающаяся стаей веслоногих ракообразных. Анимация, показывающая, как синхронно охотящиеся сельди могут поймать очень бдительного и уклончивого копепода (нажмите, чтобы просмотреть).
Анимация, показывающая, как синхронно охотящиеся сельди могут поймать очень бдительного и уклончивого копепода (нажмите, чтобы просмотреть). Стаи медуз также охотятся на веслоногих ракообразных.
Стаи медуз также охотятся на веслоногих ракообразных.



Планктонные копеподы играют важную роль в углеродном цикле . Некоторые учёные утверждают, что они образуют самую большую животную биомассу на Земле. [124] За это звание они конкурируют с антарктическим крилем . Однако из-за своего меньшего размера и относительно более высоких темпов роста, а также из-за того, что они более равномерно распределены по большей части мирового океана, копеподы почти наверняка вносят гораздо больший вклад во вторичную продуктивность мировых океанов и в глобальный сток углерода в океане, чем криля и, возможно, больше, чем все другие группы организмов вместе взятые. В настоящее время считается, что поверхностные слои океанов являются крупнейшим в мире поглотителем углерода, поглощая около 2 миллиардов тонн углерода в год, что эквивалентно, возможно, трети выбросов углерода человеком , тем самым снижая их воздействие. Многие планктонные копеподы питаются ночью у поверхности, а днем погружаются в более глубокую воду, чтобы избежать визуальных хищников. Их линяющие экзоскелеты, фекальные шарики и дыхание на глубине приносят углерод в глубокое море.
Цветение водорослей
[ редактировать ]Многие одноклеточные организмы, называемые фитопланктоном, обитают в океанах и озерах. При наличии определенных условий, таких как высокий уровень питательных веществ или света, эти организмы размножаются взрывным образом. Образующийся в результате плотный рой фитопланктона называется цветением водорослей . Цветение может занимать сотни квадратных километров, и его легко увидеть на спутниковых изображениях. Отдельный фитопланктон редко живет более нескольких дней, но цветение может длиться неделями. [125] [126]
Растения
[ редактировать ]Ученые уже сотни лет приписывают растениям роевое поведение. В своей книге 1800 года « Фитология: или Философия сельского хозяйства и садоводства » Эразм Дарвин писал, что рост растений напоминает рои, наблюдаемые в других местах в природе. [127] Хотя он имел в виду более широкие наблюдения за морфологией растений и сосредоточился на поведении как корней, так и побегов, недавние исследования подтвердили это утверждение.
Корни растений , в частности, демонстрируют заметное роевое поведение, растут по шаблонам, которые превышают статистический порог случайной вероятности, и указывают на наличие связи между отдельными верхушками корней . Основная функция корней растений — поглощение питательных веществ из почвы , и именно эта цель определяет поведение стаи. Растения, растущие в непосредственной близости, адаптировали свой рост, чтобы обеспечить оптимальную доступность питательных веществ. Это достигается за счет роста в направлении, которое оптимизирует расстояние между близлежащими корнями, тем самым увеличивая их шансы на использование неиспользованных запасов питательных веществ. Действие такого поведения принимает две формы: максимизация расстояния и отталкивание от соседних верхушек корней. [128] Переходная зона кончика корня в значительной степени отвечает за мониторинг присутствия почвенных гормонов, сигнализируя при необходимости об ответных моделях роста. Реакция станции часто бывает сложной и включает в себя множество входных данных для формирования автономной реакции. Дополнительные факторы, определяющие рост стаи, включают свет и гравитацию, оба из которых также контролируются в переходной зоне верхушки корня. [129] Эти силы действуют, чтобы информировать любое количество растущих «основных» корней, которые демонстрируют свои собственные независимые выбросы ингибирующих химических веществ, чтобы установить соответствующее расстояние, тем самым способствуя модели поведения роя. Горизонтальный рост корней, будь то в ответ на высокое содержание минералов в почве или в результате роста столонов , приводит к разветвленному росту, который также образует свои собственные, независимые корневые рои. [130]
Бактерии
[ редактировать ]Роение также описывает группировки некоторых видов хищных бактерий, таких как миксобактерии . Миксобактерии собираются в «волчьи стаи», активно перемещаясь с помощью процесса, известного как бактериальное скольжение , и держась вместе с помощью межклеточных молекулярных сигналов . [57] [131]
Млекопитающие
[ редактировать ]

- Пэрриш Дж. К., Эдельштейн-Кешет Л. (1999). «Сложность, закономерности и эволюционные компромиссы в агрегации животных» (PDF) . Наука . 284 (5411): 99–101. Бибкод : 1999Sci...284...99P . CiteSeerX 10.1.1.560.5229 . дои : 10.1126/science.284.5411.99 . ПМИД 10102827 . Архивировано из оригинала (PDF) 20 июля 2011 года.
Люди
[ редактировать ]
| Внешние изображения | |
|---|---|
Группа людей также может проявлять поведение стаи, например пешеходы. [134] или солдаты, кишащие на парапетах [ сомнительно – обсудить ] . В Кёльне, Германия, два биолога из Университета Лидса продемонстрировали стадоподобное поведение у людей. Группа людей демонстрировала модель поведения, похожую на стаю: если пять процентов стаи изменят направление, остальные последуют за ней. Если один человек был обозначен как хищник, а все остальные должны были его избегать, стая вела себя очень похоже на косяк рыб. [135] [136] Понимание того, как люди взаимодействуют в толпе, важно, если управление толпой позволит эффективно избежать жертв на футбольных полях, музыкальных концертах и станциях метро. [137]
Математическое моделирование поведения стай — распространенная технология, которая нашла применение в анимации. Имитация флокирования использовалась во многих фильмах. [138] создавать толпы , которые двигаются реалистично. Тима Бертона «Возвращение Бэтмена» был первым фильмом, в котором для рендеринга использовалась технология роя, реалистично изображающая движения группы летучих мышей с использованием системы boids . «Властелин колец» В трилогии фильма , использовалась подобная технология, известная как Massive во время батальных сцен. Технология Swarm особенно привлекательна, поскольку она дешева, надежна и проста.
Компьютерное моделирование на основе муравьев с использованием только шести правил взаимодействия также использовалось для оценки поведения при посадке самолета. [139] Авиакомпании также используют маршрутизацию на основе муравьев при назначении прибытия самолетов к выходам на посадку в аэропорту. Система авиакомпаний, разработанная Дугласом А. Лоусоном, использует теорию роя, или роевой интеллект — идею о том, что колония муравьев работает лучше, чем один. Каждый пилот действует как муравей, ищущий лучший выход на посадку в аэропорту. «Пилот на своем опыте узнает, что для него лучше, и оказывается, что это лучшее решение для авиакомпании», — объясняет Лоусон. В результате «колония» пилотов всегда выходит к воротам, они могут быстро прилететь и улететь. Программа может даже предупредить пилота о появлении резервного самолета еще до того, как оно произойдет. «Мы можем предвидеть, что это произойдет, поэтому у нас будут свободные ворота», — говорит Лоусон. [140]
Поведение роя возникает также в динамике транспортных потоков , например, в волне движения . Двунаправленное движение можно наблюдать по муравьиным тропам. [141] [142] В последние годы это поведение было исследовано для понимания моделей пешеходов и дорожного движения. [143] [144] Моделирование, основанное на моделях пешеходов, также применялось к толпам, которые бросились в панику из-за паники. [145]
Стадное поведение в маркетинге использовалось для объяснения зависимостей взаимного поведения клиентов. The Economist сообщил о недавней конференции в Риме , посвященной моделированию адаптивного поведения человека. [146] Он поделился механизмами, позволяющими увеличить импульсивные покупки и заставить людей «покупать больше, играя на стадном инстинкте». Основная идея заключается в том, что люди будут покупать больше продуктов, которые считаются популярными, и упоминаются несколько механизмов обратной связи для предоставления потребителям информации о популярности продукта, включая технологию смарт-карт и использование технологии радиочастотной идентификации . предложил модель «толпового движения» Исследователь Технологического института Флориды , которая привлекательна для супермаркетов, поскольку может «увеличить продажи без необходимости предоставлять людям скидки».
- Хелбинг Д., Кельч Дж., Молнар П. (1997). «Моделирование эволюции систем человеческих следов». Природа . 388 (6637): 47–50. arXiv : cond-mat/9805158 . Бибкод : 1997Natur.388...47H . дои : 10.1038/40353 . ПМИД 9214501 . S2CID 4364517 .
- Хелбинг Д., Фаркас И., Вичек Т. (2000). «Моделирование динамических особенностей побега паники». Природа . 407 (6803): 487–490. arXiv : cond-mat/0009448 . Бибкод : 2000Natur.407..487H . дои : 10.1038/35035023 . ПМИД 11028994 . S2CID 310346 .
- Хелбинг Д., Фаркас И.Дж., Вичек Т. (2000). «Замораживание при нагреве в приводной мезоскопической системе». Письма о физических отзывах . 84 (6): 1240–1243. arXiv : cond-mat/9904326 . Бибкод : 2000PhRvL..84.1240H . дои : 10.1103/PhysRevLett.84.1240 . ПМИД 11017488 . S2CID 18649078 .
Робототехника
[ редактировать ]
Применение принципов роя к роботам называется роевой робототехникой , а роевой интеллект относится к более общему набору алгоритмов.
| Внешние видео | |
|---|---|
Частично вдохновленные колониями насекомых, таких как муравьи и пчелы, исследователи моделируют поведение стаи тысяч крошечных роботов, которые вместе выполняют полезную задачу, например, находят что-то спрятанное, убирают или шпионят. Каждый робот довольно прост, но возникающее поведение роя более сложное. [1] Всю совокупность роботов можно рассматривать как одну распределенную систему, точно так же, как колонию муравьев можно считать суперорганизмом , обладающим роевым интеллектом . Самым большим роем, созданным на данный момент, является рой роботов-килоботов из 1024 человек. [148] Другие крупные стаи включают рой iRobot , проект SRI International /ActivMedia Robotics Centibots , [149] и рой проекта микророботов с открытым исходным кодом, который используется для исследования коллективного поведения. [150] [151] Рои также более устойчивы к сбоям. В то время как один большой робот может потерпеть неудачу и разрушить миссию, рой может продолжить работу, даже если несколько роботов потерпят неудачу. Это может сделать их привлекательными для миссий по исследованию космоса, где неудача обычно обходится чрезвычайно дорого. [152] Помимо наземной техники, роевая робототехника включает в себя также исследование стаев воздушных роботов. [147] [153] и разнородные группы наземных и воздушных транспортных средств. [154] [155]
В отличие от макроскопических роботов, коллоидные частицы на микроуровне также могут быть использованы в качестве агентов для коллективного поведения при выполнении задач с использованием механических и физических подходов, таких как реконфигурируемый микророй, похожий на торнадо. [156] имитируя стайную рыбу, [157] иерархические виды частиц [158] имитация хищнического поведения млекопитающих, манипулирование микрообъектами с помощью трансформируемого микророя. [159] Изготовление таких коллоидных частиц обычно основано на химическом синтезе.
Военный
[ редактировать ]
Военное роение — это поведение, при котором автономные или частично автономные боевые единицы атакуют врага с нескольких разных направлений, а затем перегруппировываются. Пульсация , при которой отряды смещают точку атаки, также является частью военного роения. Военное роение предполагает использование децентрализованных сил против противника таким образом, чтобы упор делался на мобильность, связь, автономию подразделений, а также координацию или синхронизацию. [160] Исторически вооруженные силы использовали принципы роения, не исследуя их подробно, но сейчас активные исследования сознательно изучают военные доктрины, которые черпают идеи из роения.
Просто потому, что несколько юнитов сходятся к цели, они не обязательно сбиваются в кучу. Осадные операции не включают в себя роение, поскольку нет маневра; схождение есть но на осажденном укреплении. также не Засады партизан представляют собой стаи, поскольку они представляют собой «наезды и бегства». Несмотря на то, что засада может иметь несколько точек атаки на противника, партизаны отступают, когда они либо нанесли достаточный урон, либо когда им угрожает опасность.
США В 2014 году Управление военно-морских исследований опубликовало видео, демонстрирующее испытания группы небольших автономных ударных катеров-дронов, которые могут управлять и предпринимать скоординированные наступательные действия в группе. [161]
Галерея
[ редактировать ] Рой мигрирующих сельдей
Рой мигрирующих сельдей Рой пчел
Рой пчел![Сальпы, выстроенные цепочками, образуют огромные стаи.[162]](//upload.wikimedia.org/wikipedia/commons/thumb/6/63/Salp.jpg/120px-Salp.jpg)
![Люди, толпящиеся через выход, не всегда ведут себя как жидкость.[163][164]](//upload.wikimedia.org/wikipedia/commons/thumb/5/54/Crowd_04378.JPG/120px-Crowd_04378.JPG)
 Рой божьих коровок
Рой божьих коровок Рой роботов
Рой роботов Рой землетрясений
Рой землетрясений Рой древних звезд
Рой древних звезд


![Сальпы, выстроенные цепочками, образуют огромные стаи.[162]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/63/Salp.jpg/120px-Salp.jpg)
![Люди, толпящиеся через выход, не всегда ведут себя как жидкость.[163][164]](http://upload.wikimedia.org/wikipedia/commons/thumb/5/54/Crowd_04378.JPG/120px-Crowd_04378.JPG)




Мифы
[ редактировать ]- Существует популярный миф о том, что лемминги совершают массовое самоубийство , сбегая со скал во время миграции. Движимые сильными биологическими побуждениями, некоторые виды леммингов могут мигрировать большими группами, когда плотность популяции становится слишком большой. Лемминги умеют плавать и могут пересечь водоем в поисках новой среды обитания. В таких случаях многие могут утонуть, если водоем настолько широк, что их физические возможности исчерпаны. Этот факт в сочетании с некоторыми необъяснимыми колебаниями численности норвежских леммингов породил миф. [165]
- Пираньи имеют репутацию бесстрашных рыб, которые сбиваются в свирепые и хищные стаи. Однако недавнее исследование, которое началось «с предпосылки, что они стайиваются как средство совместной охоты», обнаружило, что на самом деле они были довольно пугливыми рыбами, как и другие рыбы, которые обучались для защиты от своих хищников, таких как бакланы, кайманы и дельфины. Исследователь описал их как «по сути обычных рыб с большими зубами». [166]
См. также
[ редактировать ]- Активная материя - поведение материи в масштабе системы
- Рой Дайсона – гипотетическая мегаструктура вокруг звезды.
- Список собирательных существительных в английском языке
- Юбилей Мобил-Бэй - природное явление, происходящее в Мобил-Бэй, штат Алабама, США.
- Популяционный цикл
- Swarm (симуляция) – набор инструментов для моделирования на основе агентов с открытым исходным кодом.
- Свирлонное состояние - недавно (2020 г.) обнаружено новое состояние самодвижущихся частиц.
- Волна трафика - Тип загруженности шоссе.
- Свармалаторы — агенты, которые одновременно выполняют роение и синхронизацию .
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Буффанае, Роланд (2016). Проектирование и управление динамикой роя . SpringerBriefs in Complexity (первое изд.). Спрингер. дои : 10.1007/978-981-287-751-2 . ISBN 978-981-287-750-5 .
- ^ О'Лоан; Эванс (1998). «Переменное устойчивое состояние при одномерном флокинге». Журнал физики A: Математический и общий . 32 (8): Л99–Л105. arXiv : cond-mat/9811336 . Бибкод : 1999JPhA...32L..99O . дои : 10.1088/0305-4470/32/8/002 . S2CID 7642063 .
- ^ Аттанаси, А.; Каванья, А.; Дель Кастелло, Л.; Джардина, И.; Григера, Т.С.; Джелич, А.; Мелилло, С.; Паризи, Л.; Пол, О.; Шен, Э.; Виале, М. (сентябрь 2014 г.). «Передача информации и поведенческая инерция в стаях скворцов» . Физика природы . 10 (9): 691–696. arXiv : 1303.7097 . Бибкод : 2014NatPh..10..691A . дои : 10.1038/nphys3035 . ПМЦ 4173114 . ПМИД 25264452 .
- ^ Перейти обратно: а б с Рейнольдс CW (1987). «Стада, стада и школы: распределенная поведенческая модель». Материалы 14-й ежегодной конференции «Компьютерная графика и интерактивные технологии» . Том. 21. С. 25–34. CiteSeerX 10.1.1.103.7187 . дои : 10.1145/37401.37406 . ISBN 978-0-89791-227-3 . S2CID 546350 .
- ^ Перейти обратно: а б Баллерини М, Кабиббо Н, Кандельер Р, Каванья А, Сисбани Е, Джардина И, Лекомт В, Орланди А, Паризи Г, Прокаччини А, Виале М, Здравкович В (2008). «Взаимодействие, управляющее коллективным поведением животных, зависит от топологического, а не метрического расстояния: данные полевого исследования» . Учеб. Натл. акад. наук. США . 105 (4): 1232–7. arXiv : 0709.1916 . Бибкод : 2008PNAS..105.1232B . дои : 10.1073/pnas.0711437105 . ПМК 2234121 . ПМИД 18227508 .
- ^ Хильденбрандт Х., Карере С., Хемельрейк К.К. (2010). «Самоорганизованные воздушные демонстрации тысяч скворцов: модель» . Поведенческая экология . 21 (6): 1349–1359. arXiv : 0908.2677 . дои : 10.1093/beheco/arq149 .
- ^ Хемельрейк К.К., Хильденбрандт Х (2011). «Некоторые причины изменчивой формы стай птиц» . ПЛОС ОДИН . 6 (8): e22479. Бибкод : 2011PLoSO...622479H . дои : 10.1371/journal.pone.0022479 . ПМК 3150374 . ПМИД 21829627 .
- ^ «Стаи и школы - Роение - Постоянная экспозиция - Посетите наши выставки и семинары! - Наука LinX - Университет Гронингена» . 10 ноября 2007 г.
- ^ «Исследования на факультете математики и естественных наук - Факультет математики и естественных наук - О нас - Университет Гронингена» . 25 октября 2012 г.
- ^ Ян, В.; Шмикл, Т. (2019). «Коллективное движение как конечный эффект в переполненных эгоистичных стадах» . Научные отчеты . 9 (1): 6618. Бибкод : 2019NatSR...9.6618Y . дои : 10.1038/s41598-019-43179-6 . ПМЦ 6488663 . ПМИД 31036873 .
- ^ Олсон Р.С., Кнестер Д.Б., Адами С. (2013). «Критическое взаимодействие между хищничеством, зависящим от плотности, и эволюцией эгоистичного стада» . Материалы 15-й ежегодной конференции по генетическим и эволюционным вычислениям . Гекко '13. стр. 247–254. дои : 10.1145/2463372.2463394 . ISBN 9781450319638 . S2CID 14414033 .
- ^ Уорд Ч.Р., Гобет Ф., Кендалл Дж. (2001). «Эволюционирование коллективного поведения в искусственной экологии» . Искусственная жизнь . 7 (2): 191–209. CiteSeerX 10.1.1.108.3956 . дои : 10.1162/106454601753139005 . ПМИД 11580880 . S2CID 12133884 .
- ^ Релуга ТК, Вискидо С (2005). «Имитация эволюции эгоистичного стадного поведения». Журнал теоретической биологии . 234 (2): 213–225. Бибкод : 2005JThBi.234..213R . дои : 10.1016/j.jtbi.2004.11.035 . ПМИД 15757680 .
- ^ Вуд А.Дж., Экленд Г.Дж. (2007). «Эволюция эгоистичного стада: появление различных стратегий агрегирования в индивидуальной модели» . Учебник по биологическим наукам . 274 (1618): 1637–1642. дои : 10.1098/rspb.2007.0306 . ПМК 2169279 . ПМИД 17472913 .
- ^ Олсон Р.С., Хинтце А., Дайер ФК, Кнестер Д.Б., Адами С. (2013). «Замешательства хищников достаточно, чтобы развить роевое поведение» . JR Soc. Интерфейс . 10 (85): 20130305. doi : 10.1098/rsif.2013.0305 . ПМК 4043163 . ПМИД 23740485 .
- ^ Демсар Дж., Хемельрейк К.К., Хильденбрандт Х., Баец И.Л. (2015). «Имитация нападения хищников на школы: развитие комплексной тактики» (PDF) . Экологическое моделирование . 304 : 22–33. Бибкод : 2015EcMod.304...22D . doi : 10.1016/j.ecolmodel.2015.02.018 . hdl : 11370/0bfcbb69-a101-4ec1-833a-df301e49d8ef . S2CID 46988508 .
- ^ Тош ЧР (2011). «Какие условия способствуют отбору, зависящему от отрицательной плотности скоплений добычи?» (PDF) . Журнал теоретической биологии . 281 (1): 24–30. Бибкод : 2011JThBi.281...24T . дои : 10.1016/j.jtbi.2011.04.014 . ПМИД 21540037 .
- ^ Иоанну К.С., Гуттал В., Кузин И.Д. (2012). «Выбор хищных рыб для скоординированного коллективного движения виртуальной добычи» . Наука . 337 (6099): 1212–1215. Бибкод : 2012Sci...337.1212I . дои : 10.1126/science.1218919 . ПМИД 22903520 . S2CID 10203872 .
- ^ Олсон Р.С., Хейли П.Б., Дайер ФК, Адами С. (2015). «Изучение эволюции компромисса между бдительностью и поиском пищи у групповых организмов» . Королевское общество открытой науки . 2 (9): 150135. arXiv : 1408.1906 . Бибкод : 2015RSOS....250135O . дои : 10.1098/rsos.150135 . ПМЦ 4593673 . ПМИД 26473039 .
- ^ «Иерархия жизни» . 14 сентября 2008 г. Архивировано из оригинала 3 июля 2016 г. . Проверено 6 октября 2009 г.
- ^ Парунак, Х. против Д. (2003). «Как создать роение» В: Материалы конференции по роению и сетевому командованию, контролю, связи, компьютерам, разведке, наблюдению и разведке (C4ISR), Маклин, Вирджиния, США, 3 января 2003 г.
- ^ Перейти обратно: а б Марш Л.; Оноф К. (2008). «Стигмергическая эпистемология, стигмергическое познание» (PDF) . Исследование когнитивных систем . 9 (1): 136–149. дои : 10.1016/j.cogsys.2007.06.009 . S2CID 23140721 .
- ^ Бени, Г., Ван, Дж. Роевой интеллект в клеточных робототехнических системах, Продолжайте. Расширенный семинар НАТО по роботам и биологическим системам, Тоскана, Италия, 26–30 июня (1989 г.)
- ^ Дориго, М; Бираттари, М (2007). «Ройовой интеллект» . Схоларпедия . 2 (9): 1462. Бибкод : 2007SchpJ ...2.1462D . дои : 10.4249/scholarpedia.1462 .
- ^ Перейти обратно: а б Ли, YX; Люкман, Р; Эдельштейн-Кешет, Л. (2007). «Минимальные механизмы формирования школ самодвижущихся частиц» (PDF) . Физика D: Нелинейные явления . 237 (5): 699–720. Бибкод : 2008PhyD..237..699L . дои : 10.1016/j.physd.2007.10.009 .
- ^ Тонер Дж и Ту Ю (1995) «Дальний порядок в двумерной модели xy: как птицы летают вместе» Physical Revue Letters, 75 (23) (1995), 4326–4329.
- ^ Топаз С, Бертоцци А (2004). «Схемы роения в двумерной кинематической модели биологических групп». SIAM J Appl Math . 65 (1): 152–174. Бибкод : 2004APS..MAR.t9004T . CiteSeerX 10.1.1.88.3071 . дои : 10.1137/S0036139903437424 . S2CID 18468679 .
- ^ Топаз С., Бертоцци А., Льюис М. (2006). «Нелокальная континуальная модель биологической агрегации». Бычья Математика Биол . 68 (7): 1601–1623. arXiv : q-bio/0504001 . дои : 10.1007/s11538-006-9088-6 . ПМИД 16858662 . S2CID 14750061 .
- ^ Каррильо, Дж; Форназье, М; Тоскани, Дж. (2010). «Частичные, кинетические и гидродинамические модели роения» (PDF) . Математическое моделирование коллективного поведения в социально-экономических науках и науках о жизни . Моделирование и симуляция в науке, технике и технологиях. Том. 3. С. 297–336. CiteSeerX 10.1.1.193.5047 . дои : 10.1007/978-0-8176-4946-3_12 . ISBN 978-0-8176-4945-6 .
- ^ «Проект Сварманоид» . [ постоянная мертвая ссылка ]
- ^ Оптимизация колонии муравьев . Проверено 15 декабря 2010 г.
- ^ А. Колорни, М. Дориго и В. Маньеццо, Распределенная оптимизация муравьиных колоний , материалы первой европейской конференции по искусственной жизни, Париж, Elsevier Publishing, 134–142, 1991.
- ^ М. Дориго, Оптимизация, обучение и естественные алгоритмы , докторская диссертация, Миланский политехнический университет, Италия, 1992.
- ^ Хёльдоблер и Уилсон (1990), стр. 143–179.
- ^ ДОРИГО, М.; ДИ КАРО, Г.; ГАМБЕРЕЛЛА, LM (1999). Муравьиные алгоритмы дискретной оптимизации искусственной жизни . МТИ Пресс.
- ^ Модель самоуправляемых частиц. Архивировано 14 октября 2012 г. в интерактивном моделировании Wayback Machine , 2005 г., Университет Колорадо. Проверено 10 апреля 2011 г.
- ^ Перейти обратно: а б Вичек Т. , Чирок А., Бен-Джейкоб Э., Коэн И., Шошет О. (1995). «Новый тип фазового перехода в системе самодвижущихся частиц». Письма о физических отзывах . 75 (6): 1226–1229. arXiv : cond-mat/0611743 . Бибкод : 1995PhRvL..75.1226V . дои : 10.1103/PhysRevLett.75.1226 . ПМИД 10060237 . S2CID 15918052 .
- ^ Чирок А, Вичек Т (2006). «Коллективное поведение взаимодействующих самодвижущихся частиц». Физика А. 281 (1–4): 17–29. arXiv : cond-mat/0611742 . Бибкод : 2000PhyA..281...17C . дои : 10.1016/S0378-4371(00)00013-3 . S2CID 14211016 .
- ^ Буль Дж., Самптер Д.Д.Т., Кузен Д., Хейл Дж.Дж., Деспланд Э., Миллер Э.Р., Симпсон С.Дж. и др. (2006). «От беспорядка к порядку в марширующей саранче» (PDF) . Наука . 312 (5778): 1402–1406. Бибкод : 2006Sci...312.1402B . дои : 10.1126/science.1125142 . ПМИД 16741126 . S2CID 359329 . Архивировано из оригинала (PDF) 29 сентября 2011 года . Проверено 13 апреля 2011 г.
- ^ Тонер Дж., Ту Ю., Рамасвами С. (2005). «Гидродинамика и фазы скоплений» (PDF) . Анналы физики . 318 (1): 170–244. Бибкод : 2005АнФиз.318..170Т . дои : 10.1016/j.aop.2005.04.011 . Архивировано из оригинала (PDF) 18 июля 2011 года . Проверено 13 апреля 2011 г.
- ^ Бертен, Э; Дроз; Грегуар, Дж. (2009). «Гидродинамические уравнения для самодвижущихся частиц: микроскопический вывод и анализ устойчивости». Дж. Физ. А. 42 (44): 445001. arXiv : 0907.4688 . Бибкод : 2009JPhA...42R5001B . дои : 10.1088/1751-8113/42/44/445001 . S2CID 17686543 .
- ^ Кеннеди, Дж.; Эберхарт, Р. (1995). «Оптимизация роя частиц». Материалы Международной конференции IEEE по нейронным сетям . Том. IV. стр. 1942–1948.
- ^ Кеннеди, Дж. (1997). «Рой частиц: социальная адаптация знаний». Материалы Международной конференции IEEE по эволюционным вычислениям . стр. 303–308.
- ^ Hu X Оптимизация роя частиц: Учебное пособие . Проверено 15 декабря 2010 г.
- ^ Кеннеди, Дж.; Эберхарт, Р.К. (2001). Роевой интеллект . Морган Кауфманн. ISBN 978-1-55860-595-4 .
- ^ Поли, Р. (2007). «Анализ публикаций по приложениям оптимизации роя частиц» (PDF) . Технический отчет CSM-469 . Архивировано из оригинала (PDF) 16 июля 2011 года . Проверено 15 декабря 2010 г.
- ^ Поли, Р. (2008). «Анализ публикаций по применению оптимизации роя частиц» (PDF) . Журнал искусственной эволюции и приложений . 2008 : 1–10. дои : 10.1155/2008/685175 .
- ↑ Альтруизм помогает роящимся роботам летать лучше. Архивировано 15 сентября 2012 г. на сайте Wayback Machine Genevalunch.com , 4 мая 2011 г.
- ^ Вайбель, М; Флореано, Д; Келлер, Л. (2011). «Количественная проверка правила Гамильтона для эволюции альтруизма» . ПЛОС Биология . 9 (5): 1000615. doi : 10.1371/journal.pbio.1000615 . ПМК 3086867 . ПМИД 21559320 .
- ^ Хернкинд, W (1969). «Очередное поведение лангустов». Наука . 164 (3886): 1425–1427. Бибкод : 1969Sci...164.1425H . дои : 10.1126/science.164.3886.1425 . ПМИД 5783720 . S2CID 10324354 .
- ↑ Ископаемые «линии конги» раскрывают происхождение стаи животных National Geographic , 17 октября 2019 г.
- ^ Ваннье, Дж; Видаль, М; Маршан, Р; Эль Харири, К.; Курайс, К; Питтет, Б; Эль Альбани, А; Мазурье, А; Мартин, Э (2019). «Коллективное поведение членистоногих-трилобитов возрастом 480 миллионов лет из Марокко» . Научные отчеты . 9 (1): 14941. Бибкод : 2019НатСР...914941В . дои : 10.1038/s41598-019-51012-3 . ПМК 6797724 . ПМИД 31624280 .
- ^ Fear C (1984) Скворец , Издательство Оксфордского университета. ISBN 978-0-19-217705-6 .
- ^ Куропатка Б.Л. (1982). «Структура и функции рыбных косяков» (PDF) . Научный американец . Том. 246, нет. 6. С. 114–123. Бибкод : 1982SciAm.246f.114P . doi : 10.1038/scientificamerican0682-114 . ПМИД 7201674 . Архивировано из оригинала (PDF) 3 июля 2011 года.
- ^ Хаббард С., Бабак П., Сигурдссон С., Магнуссон К. (2004). «Модель формирования рыбных косяков и миграций рыб». Экол. Модель . 174 (4): 359–374. Бибкод : 2004EcMod.174..359H . doi : 10.1016/j.ecolmodel.2003.06.006 .
- ^ Раух Э., Миллонас М., Чиалво Д. (1995). «Формирование шаблонов и функциональность в роевых моделях». Буквы по физике А. 207 (3–4): 185–193. arXiv : adap-org/9507003 . Бибкод : 1995PhLA..207..185R . дои : 10.1016/0375-9601(95)00624-C . S2CID 120567147 .
- ^ Перейти обратно: а б Эллисон С., Хьюз С. (1991). «Бактериальное роение: пример прокариотической дифференциации и многоклеточного поведения». Научный прогресс . 75 (298, ч. 3–4): 403–422. ПМИД 1842857 .
- ^ Бен-Джейкоб Э., Коэн И., Шошет О., Чирок А., Вичек Т. (1995). «Кооперативное формирование хиральных паттернов при росте бактериальных колоний». Письма о физических отзывах . 75 (15): 2899–2902. Бибкод : 1995PhRvL..75.2899B . doi : 10.1103/PhysRevLett.75.2899 . ПМИД 10059433 .
- ^ Раппель В.Дж., Никол А., Саркисян А., Левин Х., Лумис В.Ф. (1999). «Самоорганизованное вихревое состояние в двумерной динамике Dictyostelium». Письма о физических отзывах . 83 (6): 1247–1250. arXiv : patt-sol/9811001 . Бибкод : 1999PhRvL..83.1247R . дои : 10.1103/PhysRevLett.83.1247 . S2CID 1590827 .
- ^ Чоудхури, Д. (2006). «Коллективные эффекты во внутриклеточном молекулярном моторном транспорте: координация, сотрудничество и конкуренция». Физика А. 372 (1): 84–95. arXiv : физика/0605053 . Бибкод : 2006PhyA..372...84C . дои : 10.1016/j.physa.2006.05.005 . S2CID 14822256 .
- ^ Пэрриш Дж. К. и Хамнер В. М. (редакторы) (1997) Группы животных в трех измерениях Издательство Кембриджского университета. ISBN 978-0-521-46024-8 .
- ^ Хелбинг Д., Кельч Дж., Молнар П. (1997). «Моделирование эволюции систем человеческих следов». Природа . 388 (6637): 47–50. arXiv : cond-mat/9805158 . Бибкод : 1997Natur.388...47H . дои : 10.1038/40353 . ПМИД 9214501 . S2CID 4364517 .
- ^ Хелбинг Д., Фаркас И., Вичек Т. (2000). «Моделирование динамических особенностей побега паники». Природа . 407 (6803): 487–490. arXiv : cond-mat/0009448 . Бибкод : 2000Natur.407..487H . дои : 10.1038/35035023 . ПМИД 11028994 . S2CID 310346 .
- ^ Хелбинг Д., Фаркас И.Дж., Вичек Т. (2000). «Замораживание при нагреве в приводной мезоскопической системе». Письма о физических отзывах . 84 (6): 1240–1243. arXiv : cond-mat/9904326 . Бибкод : 2000PhRvL..84.1240H . дои : 10.1103/PhysRevLett.84.1240 . ПМИД 11017488 . S2CID 18649078 .
- ^
- Теория роя National Geographic . Тематическая статья, июль 2007 г.
- Бикман М., Сворд Г.А. и Симпсон С.К. (2008) Биологические основы роевого интеллекта . В книге «Ройовый интеллект: введение и применение» , Эдс Блюм С. и Меркл Д. Спрингер, Япония, К.К. ISBN 978-3-540-74088-9
- Пэрриш Дж. К., Эдельштейн-Кешет Л. (1999). «Сложность, закономерности и эволюционные компромиссы в агрегации животных» (PDF) . Наука . 284 (5411): 99–101. Бибкод : 1999Sci...284...99P . CiteSeerX 10.1.1.560.5229 . дои : 10.1126/science.284.5411.99 . ПМИД 10102827 . Архивировано из оригинала (PDF) 20 июля 2011 года.
- ^ Перейти обратно: а б Бонабо Э. и Тераулаз Дж. (2008) «Рой ум». В вашем будущем с роботами . Специальные издания Scientific American.
- ^ Чачкес, Ти Джей; Грютер, К.; Ратниекс, FLW (2015). «Следовые феромоны: интегративный взгляд на их роль в организации колонии» . Ежегодный обзор энтомологии . 60 : 581–599. doi : 10.1146/annurev-ento-010814-020627 . ПМИД 25386724 . S2CID 37972066 .
- ^ Беккерс, Р.; Денебур, JL; Госс, С. (1993). «Модуляция прокладки следов у муравья Lasius niger (Hymenoptera: Formicidae) и его роль в коллективном выборе источника пищи». Журнал поведения насекомых . 6 (6): 751–759. Бибкод : 1993JIBeh...6..751B . CiteSeerX 10.1.1.161.9541 . дои : 10.1007/BF01201674 . S2CID 14670150 .
- ^ Госс, С.; Арон, С.; Денебур, JL; Пастилс, Дж. М. (1989). «Самоорганизованные ярлыки у аргентинского муравья» (PDF) . Naturwissenschaften . 76 (12): 579–581. Бибкод : 1989NW.....76..579G . дои : 10.1007/BF00462870 . S2CID 18506807 .
- ^ Брэди, Шон Г. (2003). «Эволюция синдрома армейского муравья: происхождение и долгосрочный эволюционный застой комплекса поведенческих и репродуктивных адаптаций» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (11): 6575–9. Бибкод : 2003PNAS..100.6575B . дои : 10.1073/pnas.1137809100 . ПМК 164488 . ПМИД 12750466 .
- ^ Дикке Э., Байд А., Клифф Д., Лэйзелл П. (2004). «Муравьиный метод проектирования сетей хранения данных». В AJ Ispeert, М. Мурата, Н. Вакамия (ред.). Труды биологических подходов к передовым информационным технологиям: первый международный семинар, BioADIT 2004 LNCS 3141 . Спрингер. стр. 364–379 . ISBN 9783540233398 .
- ^ Олдройд, БП (1998). «Сродство колоний в скоплениях Apis dorsata Fabricius (Hymenoptera, Apidae)». Общество насекомых . 47 : 94–95. дои : 10.1007/s000400050015 . S2CID 40346464 .
- ^ Милиус, Сьюзен Swarm Savvy. Архивировано 27 сентября 2012 г. в Wayback Machine . Как пчелы, муравьи и другие животные избегают глупых коллективных решений; Новости науки, 9 мая 2009 г.; Том 175 #10 (с. 16)
- ^ Стаи пчел следуют за высокоскоростными пчелами-стрикерами, чтобы найти новое гнездо ; ScienceDaily (24 ноября 2008 г.)
- ^ Сили, Томас Д.; Вишер, П. Кирк (2003). «Выбор дома: как разведчики в рое медоносных пчел воспринимают завершение группового принятия решения» (PDF) . Поведенческая экология и социобиология . 54 (5): 511–520. дои : 10.1007/s00265-003-0664-6 . S2CID 16948152 . Архивировано из оригинала (PDF) 31 января 2009 года . Проверено 14 декабря 2010 г.
- ^ Морс, РА (1963). «Ориентация роя у медоносных пчел». Наука . 141 (3578): 357–358. Бибкод : 1963Sci...141..357M . дои : 10.1126/science.141.3578.357 . ПМИД 17815993 . S2CID 46516976 .
- ^ Сили, Томас (2003). «Достижение консенсуса при выборе места для гнезда в стаях медоносных пчел: прекращение разногласий». Поведенческая экология и социобиология . 53 (6): 417–424. дои : 10.1007/s00265-003-0598-z . S2CID 34798300 .
- ^ Степьен, ТЛ; Змурчок, С.; Хенгениус, Дж.Б.; Каха Ривера, РМ; Д'Орсонья, MR; Линдси, А.Е. (2000). «Спаривание бабочек: моделирование призыва женских феромонов и стратегий навигации самцов для оптимизации репродуктивного успеха» . Прикладные науки . 10 (18): 6543. doi : 10.3390/app10186543 .
- ^ Бадеке, Б.; Хаверкамп, А.; Саксе, ЮАР (2016). «Задача для самца совки? Различение женского полового феромона на фоне летучих веществ растений» . Границы в физиологии . 7 : 143. doi : 10.3389/fphys.2016.00143 . ПМЦ 4843018 . ПМИД 27199761 .
- ^ Гринфилд, доктор медицины (1981). «Половые феромоны моли: эволюционная перспектива». Энтомолог Флориды . 64 (1): 4–17. дои : 10.2307/3494597 . JSTOR 3494597 .
- ^ Умберс, КДЛ; Саймондс, MRE; Кокко, Х. (2015). «Мотематика передачи сигналов женских феромонов: стратегии старения девственниц». Американский натуралист . 185 (3): 417–432. дои : 10.1086/679614 . hdl : 1885/13166 . ПМИД 25674695 . S2CID 13846468 .
- ^ Мейсон, Д.С.; Баруцци, К. (2019). «Любовь в странных местах» . Границы в экологии и окружающей среде . 17 (3): 184. Бибкод : 2019FrEE...17..184M . дои : 10.1002/плата.2027 .
- ^ «Мошки» . MDC Откройте для себя природу . Проверено 19 ноября 2019 г.
- ^ Киркеби, Карстен (30 июня 2018 г.). «Наблюдения за роевым поведением самок и смешанного пола у Culicoides LATREILLE, 1809 (Diptera: Ceratopogonidae)» . Польский журнал энтомологии . 87 (2): 191–197. дои : 10.2478/pjen-2018-0014 .
- ^ Дженнифер Вьегас. «Тараканы принимают групповые решения» . Канал Дискавери . Архивировано из оригинала 4 июля 2008 года . Проверено 10 июня 2006 г.
- ^ Лемоник, Майкл Д. (15 ноября 2007 г.). «Роботы-тараканы делают свое дело» . Время . Архивировано из оригинала 16 ноября 2007 года.
- ^ Саранча Locustidae National Geographic . Проверено 12 декабря 2010 г.
- ^ «Саранча полна серотонина» . 29 января 2009 г. – через BBC.
- ^ Роджерс С.М., Мэтисон Т., Деспланд Э., Доджсон Т., Берроуз М., Симпсон С.Дж. (ноябрь 2003 г.). «Механосенсорно-индуцированная поведенческая агрегация у пустынной саранчи Schistocerca gregaria » . Дж. Эксп. Биол . 206 (Часть 22): 3991–4002. дои : 10.1242/jeb.00648 . ПМИД 14555739 .
- ^ Стивенсон, Пенсильвания (2009). «Ключ от ящика Пандоры». Наука . 323 (5914): 594–5. дои : 10.1126/science.1169280 . ПМИД 19179520 . S2CID 39306643 .
- ↑ Блокирующее химическое вещество «счастье» может предотвратить нашествие саранчи , New Scientist , 29 января 2009 г., по состоянию на 31 января 2009 г.
- ^ Моше Гершон; Амир Аяли (май 2012 г.). «Врожденное фазовое поведение пустынной саранчи Schistocerca gregaria » . Наука о насекомых . 19 (6): 649–656. Бибкод : 2012InsSc..19..649G . дои : 10.1111/j.1744-7917.2012.01518.x . S2CID 83412818 .
- ^ Йейтс, Калифорния; Эрбан, Р; Эскудеро, К; Кузен, ID; Буль, Дж; Кеврекидис, ИГ; Майни, ПК; Самптер, диджей (2009). «Свойственный шум может способствовать слаженности коллективного движения стаи» . Учеб. Натл. акад. наук. США . 106 (14): 5464–9. Бибкод : 2009PNAS..106.5464Y . дои : 10.1073/pnas.0811195106 . ПМК 2667078 . ПМИД 19336580 .
- ^ Пайл, Роберт Майкл, «Полевой справочник Национального общества Одюбона по бабочкам Северной Америки», стр. 712-713, Альфред А. Кнопф, Нью-Йорк, ISBN 0-394-51914-0
- ^ «Монарх, Данай Плексипп» . Архивировано из оригинала 15 декабря 2012 года . Проверено 27 августа 2008 г.
- ^ Гульотта, Гай (2003): Бабочки, управляемые биологическими часами, ученые Солнца проливают свет на паломничество монархов. Архивировано 5 марта 2006 г. в Wayback Machine . Washington Post , 23 мая 2003 г., стр. A03. Проверено 7 января 2006 г.
- ^ Мерлин С., Гегир Р.Дж., Репперт С.М. (2009). «Усиковые циркадные часы координируют ориентацию по солнечному компасу у мигрирующих бабочек-монархов» . Наука . 325 (5948): 1700–1704. Бибкод : 2009Sci...325.1700M . дои : 10.1126/science.1176221 . ПМЦ 2754321 . ПМИД 19779201 .
- ^ Кириаку КП (2009). «Разоблачающее путешествие». Наука . 325 (5948): 1629–1630. дои : 10.1126/science.1178935 . ПМИД 19779177 . S2CID 206522416 .
- ^ Надь, М; Акос Зс, Биро Д; Вичек, Т (2010). «Иерархическая групповая динамика в голубиных стаях» (PDF) . Природа . 464 (7290): 890–893. arXiv : 1010.5394 . Бибкод : 2010Natur.464..890N . дои : 10.1038/nature08891 . ПМИД 20376149 . S2CID 4430488 . Архивировано из оригинала (PDF) 6 июля 2010 г. Дополнительный PDF-файл.
- ^ Секерджиоглу, Швейцария (2007). «Экология сохранения: территория превосходит мобильность в результате фрагментарного вымирания птиц» . Современная биология . 17 (8): Р283–Р286. Бибкод : 2007CBio...17.R283S . дои : 10.1016/j.cub.2007.02.019 . ПМИД 17437705 . S2CID 744140 .
- ^ Уменьшение сопротивления при полете строем. Летающие самолеты в птичьих формированиях могут значительно увеличить дальность полета ; Центр оборонной технической информации; апрель 2002 г.; Проверено 27 февраля 2008 г.
- ^ Хаммель Д.; Бойкенберг М. (1989). «Аэродинамические интерференционные эффекты в групповом полете птиц». Дж. Орнитол . 130 (1): 15–24. дои : 10.1007/BF01647158 . S2CID 823269 .
- ^ Каттс, CJ и JR Speakman (1994). «Энергосбережение при групповом полете розовоногих гусей» (PDF) . Дж. Эксп. Биол . 189 (1): 251–261. дои : 10.1242/jeb.189.1.251 . ПМИД 9317742 .
- ^ Ньютон, И. (2008). Миграционная экология птиц . Эльсельвир. ISBN 978-0-12-517367-4 .
- ^ Питчер и др. 1982.
- ^ Питчер Т.Дж. и Пэриш Дж.К. (1993) «Функции стайного поведения костистых рыб» В: Питчер Т.Дж. (ред.) Поведение костистых рыб . Чепмен и Холл, Нью-Йорк, стр. 363–440.
- ^ Hoare DJ, Krause J, Peuhkuri N и Godin JGJ (2000) Размер тела и стайность рыб в Journal of Fish Biology, 57 (6) 1351-1366.
- ^ Снексер Дж.Л., Рул Н., Бауэр К., МакРоберт С.П. (2010). «Влияние пола и фенотипа на решения об обмелении рыбок данио» (PDF) . Международный журнал сравнительной психологии . 23 : 70–81. дои : 10.46867/IJCP.2010.23.01.04 . Архивировано из оригинала (PDF) 25 июля 2011 года.
- ^ Энгесзер Р.Э., Райан М.Дж., Паричи Д.М. (2004). «Обучение социальным предпочтениям у рыбок данио» (PDF) . Современная биология . 14 (10): 881–884. Бибкод : 2004CBio...14..881E . дои : 10.1016/j.cub.2004.04.042 . ПМИД 15186744 . S2CID 18741014 .
- ^ Рибс, С.Г. (2000). «Может ли меньшинство информированных лидеров определить перемещения косяка рыб в поисках пищи?». Поведение животных . 59 (2): 403–9. дои : 10.1006/anbe.1999.1314 . ПМИД 10675263 . S2CID 4945309 .
- ^ Радаков Д.В. (1973) Обучение экологии рыб. Израильская программа научного перевода, перевод Mill H. Halsted Press, Нью-Йорк. ISBN 978-0-7065-1351-6
- ^ Фотограф: Марк ван Коллер
- ^ Хамнер, В.М.; Хамнер, ПП; Стрэнд, Юго-Запад; Гилмер, Р.В. (1983). «Поведение антарктического криля Euphausia superba : хеморецепция, питание, стайное обучение и линька ». Наука . 220 (4595): 433–5. Бибкод : 1983Sci...220..433H . дои : 10.1126/science.220.4595.433 . ПМИД 17831417 . S2CID 22161686 .
- ^ У. Килс; П. Маршалл (1995). «Криль, как он плавает и питается – новые знания с помощью новых методов (« Антарктический криль – как он плавает и питается – новые знания с помощью новых методов »)». У И. Гемпеля; Г. Хемпель (ред.). биологии полярных океанов Опыт и . результаты Издательство Фишер . стр. 201–210. ISBN 978-3-334-60950-7 .
- ^ Р. Пайпер (2007). Необыкновенные животные: Энциклопедия любопытных и необычных животных . Гринвуд Пресс. ISBN 978-0-313-33922-6 .
- ^ Хоар, Бен (2009). Миграция животных. Лондон: Музей естественной истории. п. 107. ISBN 978-0-565-09243-6 .
- ^ Перейти обратно: а б с Хоар, Бен (2009). Миграция животных. Лондон: Музей естественной истории. п. 107. ISBN 978-0-565-09243-6
- ^ Дж. С. Яффе; доктор медицинских наук Оманн; А. де Робертис (1999). «Сонарные оценки уровней дневной активности Euphausia pacifica в заливе Саанич» (PDF) . Канадский журнал рыболовства и водных наук . 56 (11): 2000–10. doi : 10.1139/cjfas-56-11-2000 . S2CID 228567512 . Архивировано из оригинала (PDF) 20 июля 2011 года.
- ^ Герайнт А. Тарлинг и Магнус Л. Джонсон (2006). «Насыщение вызывает у криля чувство погружения» . Современная биология . 16 (3): 83–4. Бибкод : 2006CBio...16..R83T . дои : 10.1016/j.cub.2006.01.044 . ПМИД 16461267 .
- ^ Ховард, Д.: « Криль », стр. 133–140 в книге Karl, HA et al. (ред.): За Золотыми воротами – океанография, геология, биология и проблемы окружающей среды в заливе Фараллон , циркуляр Геологической службы США 1198, 2001 г. Последний раз доступ к URL-адресам осуществлялся 4 июня 2010 г.
- ^ Д. Ховард. «Криль в Национальном морском заповеднике Корделл-Бэнк» . НОАА . Проверено 15 июня 2005 г.
- ^ Гандоми, АХ; Алави, АХ (2012). «Алгоритм стада криля: новый алгоритм оптимизации, основанный на биотехнологиях». Коммуникации в нелинейной науке и численном моделировании . 17 (12): 4831–4845. Бибкод : 2012CNSNS..17.4831G . дои : 10.1016/j.cnsns.2012.05.010 .
- ^ Хамнер, В.М.; Карлтон, Дж. Х. (1979). «Рой веслоногих ракообразных: характеристики и роль в экосистемах коралловых рифов» . Лимнол. Океаногр . 24 (1): 1–14. Бибкод : 1979LimOc..24....1H . дои : 10.4319/lo.1979.24.1.0001 .
- ^ Йоханнес Дюрбаум и Торстен Кюннеманн (5 ноября 1997 г.). «Биология копепод: Введение» . Ольденбургский университет Карла фон Осецкого . Архивировано из оригинала 26 мая 2010 года . Проверено 8 декабря 2009 г.
- ^ Линдси Р. и Скотт М. (2010) Что такое фитопланктон, Земная обсерватория НАСА .
- ^ Вредное цветение водорослей в Великих озерах. Архивировано 16 июня 2010 г. в Wayback Machine , 2009 г., NOAA , Центр передового опыта в области Великих озер и здоровья человека.
- ^ Дарвин, Эразм (1 января 1800 г.). Фитология: или Философия сельского хозяйства и садоводства. С теорией осушения болот и с усовершенствованной конструкцией бурового плуга . П. Бирн.
- ^ Чишак, Маржена; Компарини, Диего; Маццолаи, Барбара; Балушка, Франтишек; Арекки, Ф. Тито; Вичек, Тамаш; Манкузо, Стефано (17 января 2012 г.). «Роение в корнях растений» . ПЛОС ОДИН . 7 (1): e29759. Бибкод : 2012PLoSO...729759C . дои : 10.1371/journal.pone.0029759 . ISSN 1932-6203 . ПМК 3260168 . ПМИД 22272246 .
- ^ Балушка, Франтишек; Манкузо, Стефано; Фолькманн, Дитер; Барлоу, Питер В. (1 июля 2010 г.). «Переходная зона верхушки корня: связь сигнал-реакция в корне». Тенденции в науке о растениях . 15 (7): 402–408. Бибкод : 2010TPS....15..402B . doi : 10.1016/j.tplants.2010.04.007 . ПМИД 20621671 .
- ^ Дж., Тревавас А. (2014). Поведение и интеллект растений . Издательство Оксфордского университета. ISBN 9780199539543 . OCLC 961862730 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Райхенбах Х (2001). «Миксобактерии – продуценты новых биологически активных веществ» . J Ind Microbiol Biotechnol . 27 (3): 149–56. дои : 10.1038/sj.jim.7000025 . ПМИД 11780785 . S2CID 34964313 .
- ^ Фаркас И., Хелбинг Д., Вичек Т. (2002). «Мексиканские волны в возбудимой среде» (PDF) . Природа . 419 (6903): 131–132. arXiv : cond-mat/0210073 . Бибкод : 2002Natur.419..131F . дои : 10.1038/419131a . ПМИД 12226653 . S2CID 4309609 . Архивировано из оригинала (PDF) 10 июля 2007 года.
- ^ Неда З., Равас Э., Бреше Ю., Вичек Т., Барабаси А.Л. (2002). «Физика ритмических аплодисментов» (PDF) . Физический обзор E . 61 (6): 6987–6992. arXiv : cond-mat/0006423 . Бибкод : 2000PhRvE..61.6987N . дои : 10.1103/physreve.61.6987 . ПМИД 11088392 . S2CID 14135891 . Архивировано из оригинала (PDF) 11 июня 2011 года.
- ^ Хелбинг, Д; Кельч, Дж; Мольнар, П. (1997). «Моделирование эволюции систем человеческих следов». Природа . 388 (6637): 47–50. arXiv : cond-mat/9805158 . Бибкод : 1997Natur.388...47H . дои : 10.1038/40353 . ПМИД 9214501 . S2CID 4364517 .
- ^ " http://psychcentral.com/news/2008/02/15/herd-mentality-explained/1922.html Архивировано 29 ноября 2014 г. в Wayback Machine ". Проверено 31 октября 2008 года.
- ^ «Численная опасность во время хаджа» . Национальный. 6 сентября 2009 г.
- ^ Кузен И.Д., Краузе Дж. (2003). Самоорганизация и коллективное поведение у позвоночных (PDF) . Достижения в изучении поведения. Том. 32. стр. 1–75. дои : 10.1016/S0065-3454(03)01001-5 . ISBN 978-0-12-004532-7 . Архивировано из оригинала (PDF) 13 марта 2012 года . Проверено 14 апреля 2011 г.
- ^ Габбай, JME (2005). Сложность и аэрокосмическая промышленность: понимание возникновения путем связи структуры с производительностью с использованием мультиагентных систем (Диссертация). Манчестер: Докторская диссертация Манчестерского университета. Архивировано из оригинала 19 декабря 2014 года . Проверено 11 июля 2009 г.
- ^ Ливермор Р. (2008) «Мультиагентный системный подход к симуляционному исследованию, сравнивающему эффективность посадки в самолет с использованием заранее назначенных мест и стратегий свободного доступа для всех». Архивировано 13 августа 2011 г. в Wayback Machine Открытом университете , Технический отчет Нет. 2008/25.
- ^ «Самолеты, поезда и муравейники: ученые-компьютерщики моделируют деятельность муравьев, чтобы уменьшить задержки рейсов». Архивировано 24 ноября 2010 г. в Wayback Machine Science Daily , 1 апреля 2008 г.
- ^ Берд, Мартин; Н. Аранвела (февраль 2003 г.). «Частота встреч в лоб и скорость ходьбы собирателей в движении муравьев-листорезов». Общество насекомых . 50 (1): 3–8. дои : 10.1007/s000400300001 . S2CID 23876486 .
- ^ Рибейро, Педро; Андре Фрасао Элен; Жилберто Ксавьер; Карлос Навас; Фернандо Лейте Рибейро (1 апреля 2009 г.). Дорнхаус, Анна (ред.). «Муравьи могут научиться добывать пищу на тропах с односторонним движением» . ПЛОС ОДИН . 4 (4): е5024. Бибкод : 2009PLoSO...4.5024R . дои : 10.1371/journal.pone.0005024 . ПМЦ 2659768 . ПМИД 19337369 .
- ^ Джон, Александр; Андреас Шадшнайдер; Дебашиш Чоудхури; Кацухиро Нишинари (март 2008 г.). «Характеристики муравьиного транспортного потока». Роевой интеллект . 2 (1): 25–41. arXiv : 0903.1434 . дои : 10.1007/s11721-008-0010-8 . S2CID 18350336 .
- ^ Мы уже почти у цели? Автомобилистам есть чему поучиться у муравьев The Economist , 10 июля 2009 г.
- ^ Хелбинг, Дирк; Фаркас, Элайджа; Вичек, Тамаш (2000). «Моделирование динамических особенностей побега паники». Природа . 407 (6803): 487–490. arXiv : cond-mat/0009448 . Бибкод : 2000Natur.407..487H . дои : 10.1038/35035023 . ПМИД 11028994 . S2CID 310346 .
- ^ «Роение на полках: как магазины могут использовать стадный менталитет людей для увеличения продаж?». Экономист . 11 ноября 2006 г. с. 90.
- ^ Перейти обратно: а б Кушлеев, Алекс; Меллингер, Дэниел; Пауэрс, Кейтлин; Кумар, Виджай (2013). «К рою маневренных микроквадроторов». Автономные роботы . 35 (4): 287–300. дои : 10.1007/s10514-013-9349-9 . S2CID 18340816 .
- ^ «Группа исследования самоорганизующихся систем» . Архивировано из оригинала 26 октября 2014 года.
- ^ «Совместный разведывательный проект Centibots 100-роботов» . АктивМедиа Робототехника. Архивировано из оригинала 14 июля 2011 года.
- ^ «Проект микророботов с открытым исходным кодом» . Проверено 28 октября 2007 г.
- ^ «Рой» . Корпорация АйРобот. Архивировано из оригинала 27 сентября 2007 года . Проверено 28 октября 2007 г.
- ^ Кнапп, Луиза (21 декабря 2000 г.). «Смотри, в небе: Робофлай» . Проводной . Проверено 25 сентября 2008 г.
- ^ Саска, Мартин; Ян, Вакула; Либор, Преусил (2014). Стаи микролетательных аппаратов стабилизировались при визуальной относительной локализации . Международная конференция IEEE по робототехнике и автоматизации (ICRA).
- ^ Саска, Мартин; Вонасек, Войтех; Крайник, Томас; Преусил, Либор (2014). «Координация и навигация гетерогенных образований MAV–UGV, локализованных с помощью подхода «ястребий глаз», в рамках модельной схемы прогнозирующего управления» (PDF) . Международный журнал исследований робототехники . 33 (10): 1393–1412. дои : 10.1177/0278364914530482 . S2CID 1195374 .
- ^ Саска, Мартин; Вонасек, Войтех; Крайник, Томас; Преусил, Либор (2012). Координация и навигация разнородных групп БПЛА-УГВ, локализованных методом «ястребиного глаза» . Международная конференция IEEE/RSJ по интеллектуальным роботам и системам (IROS).
- ^ Цзи, Фэнтун; Джин, Дондон; Ван, Бен; Чжан, Ли (23 июня 2020 г.). «Световое зависание магнитного микророя в жидкости» . АСУ Нано . 14 (6): 6990–6998. дои : 10.1021/acsnano.0c01464 . ISSN 1936-0851 . ПМИД 32463226 . S2CID 218976382 .
- ^ Хьюз, Робин (22 февраля 2007 г.), Barracuda Tornado , получено 7 февраля 2022 г.
- ^ Моу, Фанчжи; Ли, Сяофэн; Се, Ци; Чжан, Цзяньхуа; Сюн, Канг; Сюй, Лейлей; Гуань, Цзяньго (20 декабря 2019 г.). «Активные микромоторные системы, построенные из пассивных частиц с биомиметическими взаимодействиями хищник-жертва» . АСУ Нано . 14 (1): 406–414. дои : 10.1021/acsnano.9b05996 . ISSN 1936-0851 . ПМИД 31860277 . S2CID 209435036 .
- ^ Ю, Цзянфань; Ван, Бен; Ду, Синчжоу; Ван, Цяньцянь; Чжан, Ли (21 августа 2018 г.). «Сверхрасширяемый лентообразный магнитный микророй» . Природные коммуникации . 9 (1): 3260. Бибкод : 2018NatCo...9.3260Y . дои : 10.1038/s41467-018-05749-6 . ISSN 2041-1723 . ПМК 6104072 . ПМИД 30131487 .
- ^ Эдвардс, Шон Дж. А. (2000). Роение на поле боя: прошлое, настоящее и будущее . Монография Рэнда MR-1100. Корпорация Рэнд. ISBN 978-0-8330-2779-5 .
- ↑ ВМС США могут «наводнить» противников лодками-роботами , CNN , 13 октября 2014 г.
- ^ «Ныряй и открывай: Научная экспедиция 10: Антарктида» . Проверено 3 сентября 2008 г.
- ↑ Моделирование толпы: моделирование поведения толпы людей или стаи животных имеет как легкомысленное, так и важное применение The Economist , 5 марта 2009 г.
- ^ Фишер, Лен (2009) Идеальный рой: наука о сложности в повседневной жизни. Страница 57. Основные книги. ISBN 978-0-465-01884-0
- ^ Вудфорд, Райли. «Миф о самоубийстве леммингов: фальшивое поведение диснеевского фильма» . Архивировано из оригинала 3 января 2010 года.
{{cite web}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Краснобрюхая пиранья действительно желтая New York Times , 24 мая 2005 г.
Источники
[ редактировать ]- Блюм С. и Меркл Д. (2008) Ройовый интеллект: введение и применение Springer. ISBN 978-3-540-74088-9 .
- Камазин С., Денебур Дж.Л., Фрэнкс Н.Р. , Снейд Дж., Тераулаз Г. и Бонабо Э. (2003) Самоорганизация в биологических системах Издательство Принстонского университета. ISBN 978-0-691-11624-2 .
- Фишер Л. (2009) Идеальный рой: наука о сложности в повседневной жизни . Основные книги. ISBN 978-0-465-01884-0 .
- Кеннеди Дж. Ф., Кеннеди Дж., Эберхарт Р. К. и Ши Ю (2001) Роевой интеллект Морган Кауфманн. ISBN 978-1-55860-595-4 .
- Краузе, Дж. (2005) Жизнь в группах Oxford University Press. ISBN 978-0-19-850818-2
- Лим С.П., Джайн Л.С. и Дехури С. (2009) Инновации в роевом интеллекте Springer. ISBN 978-3-642-04224-9 .
- Миллер, Питер (2010) Умный рой: как понимание стад, школ и колоний может помочь нам лучше общаться, принимать решения и добиваться результатов. Пингвин, ISBN 978-1-58333-390-7
- Неджа Н. и Мурель ЛдМ (2006) Интеллектуальные системы Swarm Springer. ISBN 978-3-540-33868-0 .
- Самптер, Дэвид Дж. Т. (2010) Коллективное поведение животных, издательство Принстонского университета. ISBN 978-0-691-14843-4 .
- Вичек А., Зафейрис А. (2012). «Коллективное движение». Отчеты по физике . 517 (3–4): 71–140. arXiv : 1010.5017 . Бибкод : 2012ФР...517...71В . дои : 10.1016/j.physrep.2012.03.004 . S2CID 119109873 .
Внешние ссылки
[ редактировать ]- Статья New York Times о расследовании роения
- Из демонстрационного проекта Wolfram – требуется проигрыватель CDF (бесплатно) :
| Биологическое роение |  | |
|---|---|---|
| Миграция животных | ||
| Роевые алгоритмы | ||
| Коллективное движение | ||
| Роевая робототехника | ||
| Связанные темы | ||