Мобильные генетические элементы

Мобильные генетические элементы ( МГЭ ), иногда называемые эгоистичными генетическими элементами , [ 1 ] представляют собой тип генетического материала , который может перемещаться внутри генома или передаваться от одного вида или репликона к другому. МГЭ обнаружены во всех организмах. Считается, что у человека примерно 50% генома составляют MGE. [ 2 ] МГЭ играют особую роль в эволюции. События дупликации генов также могут происходить посредством механизма MGE. MGE также могут вызывать мутации в кодирующих участках белка, что изменяет функции белка. Эти механизмы также могут перестраивать гены в геноме хозяина, создавая вариации. Эти механизмы могут повысить физическую форму за счет приобретения новых или дополнительных функций. Примером MGE в эволюционном контексте является то, что факторы вирулентности и гены устойчивости к антибиотикам MGE могут транспортироваться для совместного использования генетического кода с соседними бактериями. Однако MGE также могут снижать физическую форму, вводя аллели или мутации, вызывающие заболевания. [ 3 ] Совокупность МГЭ в организме называется мобиломом , который состоит из большого числа плазмид , транспозонов и вирусов . [ 4 ]

Типы
[ редактировать ]- Плазмиды : обычно это кольцевые внехромосомные молекулы ДНК , которые реплицируются и передаются независимо от хромосомной ДНК. Эти молекулы присутствуют у прокариот ( бактерий и архей ), а иногда и у эукариотических организмов, таких как дрожжи . Пригодность плазмиды определяется ее подвижностью. Первым фактором приспособленности плазмиды является ее способность реплицировать ДНК. Второй фактор приспособленности — это способность плазмиды к горизонтальному переносу. Плазмиды в течение своего цикла переносят гены от одного организма к другому посредством процесса, называемого конъюгацией . Плазмиды обычно содержат набор генов подвижности, необходимых для конъюгации. Некоторые плазмиды используют мембраносвязанное образование спаривающихся пар (MPF). Плазмида, содержащая собственные гены MPF, считается самопередающейся или конъюгативной. [ 5 ] Плазмиды можно разделить на мобилизуемые и немобилизуемые классы. Плазмиды, которые используют MFP других генетических элементов в клетке, являются мобилизуемыми. Плазмиды, которые не мобилизуются, но распространяются путем трансдукции или трансформации, называются немобилизуемыми. [ 5 ] Плазмиды часто могут вводить гены, которые делают бактерии устойчивыми к антибиотикам . [ 6 ] [ 5 ]
- Векторы клонирования : это типы гибридных плазмид с бактериофагами , используемые для переноса и репликации ДНК. Фрагменты ДНК могут быть вставлены с помощью рекомбинантной ДНК методов . Жизнеспособный вектор должен быть способен реплицироваться вместе с фрагментами ДНК, которые он несет. Эти векторы могут содержать нужные гены для внедрения в геном организма. Примерами являются космиды и фагмиды . [ 7 ]
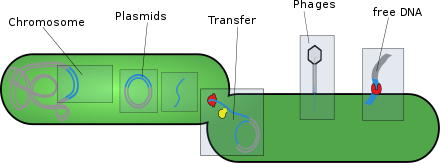
- Транспозоны : это последовательности ДНК клетки , которые могут перемещаться и реплицироваться в разных частях генома . Их также называют «прыгающими генами», они могут передаваться горизонтально между организмами, живущими в симбиозе . Транспозоны присутствуют во всех живых существах и в гигантских вирусах . [ 8 ]
- ДНК-транспозоны : это транспозоны, которые перемещаются непосредственно из одного положения в другое в геноме, используя транспозазу , чтобы разрезать и прикрепляться к другому локусу . [ 9 ] Эти генетические элементы расщепляются транспозазой в четырех одноцепочечных участках ДНК. Чтобы достичь максимальной стабильности промежуточного транспозона, происходит расщепление одной цепи целевой ДНК. Одновременно донорскую цепь лигируют с целевой цепью после расщепления, оставляя одиночный выступ на обоих концах целевой последовательности. Эти сайты обычно содержат выступ из 5–9 пар оснований, который может создать сплоченный конец. [ 10 ] Затем транспозаза удерживает последовательность в перекрестном образовании и связывает донорскую цепь с целевой цепью. Структура, образованная дуплексом ДНК и транспозазы в репликативных транспозонах, известна как промежуточный продукт Шапиро. [ 11 ] Выступающий выступ из 5–9 пар оснований остается по обе стороны от целевой последовательности, что позволяет ей присоединяться к целевой последовательности в любой ориентации. Последовательность этих выступов может определять ориентацию соединения. [ 10 ] Прежде чем может произойти сайт-специфическая рекомбинация, концы олигонуклеотидов должны быть заполнены. Лигирование этих концов образует репликационную вилку на каждом конце мобильного элемента. Смещение одноцепочечной цепи вызывает синтез несвязанной 3'-гидроксильной группы с образованием длинных одноцепочечных участков, прилегающих к 5'-концу. Следовательно, противоположная цепь секвенируется прерывисто по мере того, как обе репликационные вилки приближаются к центру мобильного элемента. В результате образуются два рекомбинантных дуплекса, содержащие полуконсервативный мобильный элемент, фланкированный предыдущим выступающим элементом из 5–9 пар оснований. Между двумя мобильными элементами происходит сайт-специфическая реципрокная рекомбинация, чему способствуют белки. Эта взаимная репликация перекрывается во времени и происходит между дублированными сегментами элемента репликации до завершения репликации. [ 10 ] В результате молекула-мишень содержит вставленный элемент, фланкированный последовательностями из 5-9 пар оснований. Транспозиция этих элементов дублирует элемент транспозиции, оставляя элемент транспозиции в исходном месте и новый транспозон в сайте реципрокной репликации. При этом общее количество пар оснований в геномах организмов увеличивается. Частота транспозиций увеличивается с течением времени и по мере старения организмов.
- Ретротранспозоны : это транспозоны, которые перемещаются по геному и транскрибируются в РНК , а затем в ДНК с помощью обратной транскриптазы . Многие ретротранспозоны также демонстрируют репликативную транспозицию . Ретротранспозоны присутствуют исключительно у эукариот . [ 12 ] Ретротранспозоны состоят из двух основных типов: длинных концевых повторов (LTR) и транспозонов, не относящихся к LTR. Транспозоны, не относящиеся к LTR, можно дополнительно разделить на длинные вкрапленные ядерные элементы (LINE) и короткие вкрапленные ядерные элементы (SINE). [ 13 ] Эти ретротранспозоны регулируются семейством коротких некодирующих РНК, называемых PIWI [P-элементом, индуцирующим слабые семенники]-взаимодействующими РНК (piRNA). [ 14 ] piRNA — это недавно открытый класс нкРНК, длина которых составляет ~24–32 нуклеотида. Первоначально piRNA были описаны как siRNA, ассоциированные с повторами (rasiRNA), поскольку они происходят из повторяющихся элементов, таких как мобильные последовательности генома. Однако позже выяснилось, что они действовали через PIWI-белок. Помимо роли в подавлении геномных транспозонов, недавно сообщалось о различных ролях piRNA, таких как регуляция 3'-UTR генов, кодирующих белок, через РНКи, трансгенерационное эпигенетическое наследование для передачи памяти о прошлой активности транспозонов и РНК-кодирование. индуцированное эпигенетическое молчание. [ 14 ]
- Интегроны : это генные кассеты , которые обычно несут гены устойчивости к антибиотикам бактериальных плазмид и транспозонов. [ 15 ]
- Интроны : групп I и Интроны II представляют собой нуклеотидные последовательности с каталитической активностью, которые являются частью транскриптов хозяина и действуют как рибозимы, которые могут вторгаться в гены, кодирующие тРНК , рРНК и белки . Они присутствуют во всех клеточных организмах и вирусах. [ 16 ]
- Интронеры: последовательности, подобные транспозонам, которые могут прыгать в геноме, оставляя новые интроны там, где они были. Они были указаны как возможный механизм получения интронов в эволюции эукариот, где они присутствуют по крайней мере у 5% всех видов, особенно у водных таксонов, возможно, из-за горизонтального переноса генов , который чаще происходит у этих животных. [ 17 ] [ 18 ] Впервые они были описаны в 2009 году у одноклеточных зеленых водорослей micromonas . [ 19 ]
- Вирусные агенты : в основном это инфекционные бесклеточные агенты, которые реплицируются в клетках-хозяевах. Во время инфекционного цикла они могут переносить гены от одного хозяина к другому. Они также могут переносить гены от одного организма к другому в случае, если вирусный агент заражает более двух разных видов. Традиционно их считают отдельными объектами, но правда в том, что многие исследователи, изучающие их характеристики и эволюцию, называют их мобильными генетическими элементами. Это основано на том факте, что вирусные агенты представляют собой простые частицы или молекулы, которые реплицируются и передаются между различными хозяевами, как и остальные невирусные мобильные генетические элементы. Согласно этой точке зрения, вирусы и другие вирусные агенты не следует считать живыми существами , их лучше рассматривать как мобильные генетические элементы. Вирусные агенты эволюционно связаны с различными мобильными генетическими элементами. [ 20 ] Считается, что эти вирусные агенты возникли из секретируемых или выделяемых плазмид других организмов. Транспозоны также дают представление о том, как эти элементы могли возникнуть изначально. Эта теория известна как гипотеза бродяжничества, предложенная Барбарой МакКлинток в 1950 году. [ 21 ] [ 1 ] [ 22 ] [ 4 ] [ 23 ]
- Вирусы : это вирусные агенты, состоящие из молекул генетического материала (ДНК или РНК) и способные образовывать сложные частицы, называемые вирионами , чтобы легко перемещаться между хозяевами. Вирусы присутствуют во всех живых существах. Вирусные частицы производятся репликативным механизмом хозяина для горизонтального переноса. [ 20 ] [ 21 ] [ 24 ]
- Сателлитные нуклеиновые кислоты : это молекулы ДНК или РНК, которые инкапсулированы в качестве «зайца» в вирионы определенных вирусов-помощников и возможность репликации которых зависит от них. Хотя их иногда считают генетическими элементами вирусов-помощников, они не всегда обнаруживаются в их вирусах-помощниках. [ 20 ] [ 21 ] [ 25 ]
- Вироиды : это вирусные агенты, состоящие из небольших кольцевых молекул РНК, которые заражают и размножаются в растениях . Эти мобильные генетические элементы не имеют защитной белковой оболочки. В частности, эти мобильные генетические элементы обнаружены у покрытосеменных растений . [ 20 ] [ 21 ] [ 26 ]
- Эндогенный вирусный элемент : это вирусные нуклеиновые кислоты, интегрированные в геном клетки. Они могут перемещаться и размножаться несколько раз в клетке-хозяине, не вызывая заболеваний или мутаций. Их считают автономными формами транспозонов. Примерами являются провирусы и эндогенные ретровирусы . [ 27 ]
Примеры исследований
[ редактировать ]Системы CRISPR-Cas у бактерий и архей представляют собой адаптивные иммунные системы для защиты от смертельных последствий МГЭ. Используя сравнительный геномный и филогенетический анализ, исследователи обнаружили, что варианты CRISPR-Cas связаны с разными типами MGE, такими как мобильные элементы. В транспозонах, связанных с CRISPR , CRISPR-Cas контролирует распространение мобильных элементов. [ 28 ]
МГЭ, такие как плазмиды, посредством горизонтальной передачи обычно полезны для организма. Способность передавать плазмиды (совместное использование) важна с эволюционной точки зрения. Таззиман и Бонхеффер обнаружили, что фиксация (получение) перенесенных плазмид в новом организме так же важна, как и способность их переносить. [ 29 ] Полезные редкие и переносимые плазмиды имеют более высокую вероятность фиксации, тогда как вредные переносимые генетические элементы имеют более низкую вероятность фиксации, поскольку они смертельны для организмов-хозяев.
Один тип MGE, а именно интегративные конъюгативные элементы (ICE), играет центральную роль в горизонтальном переносе генов, формируя геномы прокариот, обеспечивая быстрое приобретение новых адаптивных признаков. [ 30 ] [ 31 ]
В качестве репрезентативного примера ICEs, ICE Bs1 хорошо охарактеризован своей ролью в SOS-ответе Bacillus subtilis на глобальное повреждение ДНК. [ 32 ] к радиации и высыханию , а также его потенциальная связь с устойчивостью спор Bacillus pumilus SAFR-032 [ 33 ] изолированы от помещений чистых помещений космического корабля. [ 34 ] [ 35 ] [ 36 ]
Транспозиция мобильными элементами является мутагенной. Таким образом, организмы научились подавлять события транспозиции, а неспособность подавить эти события вызывает рак в соматических клетках. Чекко и др. обнаружили, что в раннем возрасте транскрипция ретротранспозируемых элементов у мышей минимальна, но в пожилом возрасте уровень транскрипции увеличивается. [ 37 ] Этот возрастной уровень экспрессии мобильных элементов снижается с помощью диеты с ограничением калорий. Репликация мобильных элементов часто приводит к добавлению в геном повторяющихся последовательностей. Эти последовательности часто некодируют, но могут мешать кодирующим последовательностям ДНК. Хотя транспозоны обладают мутагенетической природой, они увеличивают геном организма, в который они переносятся. Необходимо провести дополнительные исследования того, как эти элементы могут служить инструментом быстрой адаптации, используемым организмами для создания изменчивости. Многие транспозиционные элементы находятся в состоянии покоя или требуют активации. Следует также отметить, что текущие значения для кодирующих последовательностей ДНК были бы выше, если бы в качестве кодирующих последовательностей рассматривались элементы транспозиции, которые кодируют свой собственный транспозиционный аппарат.
Некоторые другие исследованные примеры включают Mavericks, [ 38 ] [ 39 ] [ 40 ] Звездолеты [ 41 ] [ 40 ] и Космические захватчики (или СПИНЫ) [ 42 ] [ 43 ]
Болезни
[ редактировать ]Последствия мобильных генетических элементов могут изменить паттерны транскрипции, что часто приводит к генетическим нарушениям, таким как иммунные нарушения, рак молочной железы, рассеянный склероз и боковой амиотрофический склероз. У людей стресс может привести к транзакционной активации MGE, таких как эндогенные ретровирусы , и эта активация связана с нейродегенерацией . [ 44 ]
Другие примечания
[ редактировать ]Совокупность всех мобильных генетических элементов в геноме можно назвать мобиломом .
Барбара МакКлинток была удостоена 1983 года Нобелевской премии по физиологии и медицине «за открытие мобильных генетических элементов» ( transposable elements ). [ 45 ]
Мобильные генетические элементы играют решающую роль в распространении факторов вирулентности, таких как экзотоксины и экзоферменты , среди бактерий. Были предложены стратегии борьбы с некоторыми бактериальными инфекциями путем воздействия на эти специфические факторы вирулентности и мобильные генетические элементы. [ 46 ]
См. также
[ редактировать ]- База данных ACLAME (КЛАССификация мобильных генетических элементов)
- Опять генное рождение
- Перетасовка экзонов
- Слияние генов
- Дупликация генов
- Горизонтальный перенос генов
- Факторы вирулентности
- Миниатюрные мобильные элементы с инвертированным повтором (MITE)
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Морейра Д., Лопес-Гарсиа П. (апрель 2009 г.). «Десять причин исключить вирусы из древа жизни». Обзоры природы. Микробиология . 7 (4): 306–311. дои : 10.1038/nrmicro2108 . ПМИД 19270719 . S2CID 3907750 .
- ^ Му Х, Ахмад С, Хур С (2016). Эндогенные ретроэлементы и сенсоры врожденного иммунитета хозяина . Достижения иммунологии. Том. 132. стр. 47–69. дои : 10.1016/bs.ai.2016.07.001 . ISBN 9780128047972 . ПМК 5135014 . ПМИД 27769507 .
- ^ Сингх П.К., Бурк Дж., Крейг Н.Л., Дубнау Дж.Т., Фешотт С., Флаш Д.А. и др. (18 ноября 2014 г.). «Мобильные генетические элементы и эволюция генома 2014» . Мобильная ДНК . 5:26 . дои : 10.1186/1759-8753-5-26 . ПМЦ 4363357 . ПМИД 30117500 .
- ^ Перейти обратно: а б Кунин Е.В., Вольф Ю.И. (декабрь 2008 г.). «Геномика бактерий и архей: новый динамический взгляд на мир прокариот» . Исследования нуклеиновых кислот . 36 (21): 6688–6719. дои : 10.1093/нар/gkn668 . ПМЦ 2588523 . ПМИД 18948295 .
- ^ Перейти обратно: а б с Смилли С., Гарсильян-Барсия, депутат парламента, Франсия М.В., Роча Е.П., де ла Крус Ф. (сентябрь 2010 г.). «Мобильность плазмид» . Обзоры микробиологии и молекулярной биологии . 74 (3): 434–452. дои : 10.1128/MMBR.00020-10 . ПМЦ 2937521 . ПМИД 20805406 .
- ^ Саммерс Д. (1996). «Глава 1 – Функции и организация плазмид» . Биология плазмид (первое изд.). Уайли-Блэквелл. стр. 21–22. ISBN 978-0632034369 .
- ^ Глик Б.Р., Пастернак Дж.Дж. (2005). Принципы молекулярной биотехнологии и применение рекомбинантной ДНК (3-е изд.). АСМ Пресс. ISBN 9781555816124 .
- ^ Макаловский В., Готеа В., Панде А., Макаловска I (2019). «Мобильные элементы: классификация, идентификация и их использование как инструмент сравнительной геномики». В Анисимовой М (ред.). Эволюционная геномика . Методы молекулярной биологии. Том. 1910. Нью-Йорк, штат Нью-Йорк: Humana. стр. 185–186. дои : 10.1007/978-1-4939-9074-0_6 . ISBN 978-1-4939-9074-0 . ПМИД 31278665 . S2CID 195814061 .
- ^ Муньос-Лопес М., Гарсия-Перес Х.Л. (апрель 2010 г.). «ДНК-транспозоны: природа и применение в геномике» . Современная геномика . 11 (2): 115–128. дои : 10.2174/138920210790886871 . ПМЦ 2874221 . ПМИД 20885819 .
- ^ Перейти обратно: а б с Шапиро Дж. А. (апрель 1979 г.). «Молекулярная модель транспозиции и репликации бактериофага Мю и других мобильных элементов» . Труды Национальной академии наук Соединенных Штатов Америки . 76 (4): 1933–1937. Бибкод : 1979ПНАС...76.1933С . дои : 10.1073/pnas.76.4.1933 . ПМЦ 383507 . ПМИД 287033 .
- ^ Бушман Ф (2002). Латеральный перенос ДНК: механизмы и последствия . Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор. ISBN 0-87969-603-6 . OCLC 47283049 .
- ^ Ричардсон С.Р., Дусе А.Дж., Копера ХК, Молдован Ж.Б., Гарсиа-Перес Х.Л., Моран Дж.В. (апрель 2015 г.). «Влияние ретротранспозонов LINE-1 и SINE на геномы млекопитающих» . Микробиологический спектр . 3 (2): 1165–1208. doi : 10.1128/microbiolspec.mdna3-0061-2014 . ISBN 9781555819200 . ПМЦ 4498412 . ПМИД 26104698 .
- ^ Макаловский В., Готеа В., Панде А., Макаловска I (2019). «Мобильные элементы: классификация, идентификация и их использование как инструмент сравнительной геномики». В Анисимовой М (ред.). Эволюционная геномика . Методы молекулярной биологии. Том. 1910. Нью-Йорк, штат Нью-Йорк: Humana. стр. 177–207. дои : 10.1007/978-1-4939-9074-0_6 . ISBN 978-1-4939-9074-0 . ПМИД 31278665 . S2CID 195814061 .
- ^ Перейти обратно: а б Монга I, Банерджи I (ноябрь 2019 г.). «Вычислительная идентификация piRNA с использованием признаков, основанных на последовательности, структуре, термодинамических и физико-химических свойствах РНК» . Современная геномика . 20 (7): 508–518. дои : 10.2174/1389202920666191129112705 . ПМЦ 7327968 . PMID 32655289 .
- ^ Ковалевская Н.П. (2002). «Мобильные генные кассеты и интеграны». Молекулярная биология . 36 (2): 196–201. дои : 10.1023/А:1015361704475 . S2CID 2078235 .
- ^ Хауснер Г., Хафез М., Эджелл Д.Р. (март 2014 г.). «Интроны бактериальной группы I: мобильные РНК-катализаторы» . Мобильная ДНК . 5 (1): 8. дои : 10.1186/1759-8753-5-8 . ПМЦ 3984707 . ПМИД 24612670 .
- ^ Гозашти Л., Рой С.В., Торнлоу Б., Крамер А., Арес М., Корбетт-Детиг Р. (ноябрь 2022 г.). «Мобильные элементы способствуют увеличению количества интронов у различных эукариот» . Труды Национальной академии наук Соединенных Штатов Америки . 119 (48): e2209766119. Бибкод : 2022PNAS..11909766G . дои : 10.1073/pnas.2209766119 . ПМЦ 9860276 . ПМИД 36417430 .
- ^ Бюлер Дж. (30 марта 2023 г.). «Как ДНК-паразит мог фрагментировать наши гены» . Журнал Кванта . Проверено 31 марта 2023 г.
- ^ Уорден А.З., Ли Дж.Х., Мок Т., Рузе П., Симмонс М.П., Аэртс А.Л. и др. (апрель 2009 г.). «Зеленая эволюция и динамические адаптации, выявленные геномами морских пикоэукариот Micromonas» . Наука . 324 (5924): 268–272. Бибкод : 2009Sci...324..268W . дои : 10.1126/science.1167222 . ПМИД 19359590 . S2CID 206516961 .
- ^ Перейти обратно: а б с д Кун Дж.Х., Доля В.В., Крупович М., Адриансенс Э.М., Ди Серио Ф., Дутиль Б.Е. и др. (2020). Расширять, дополнять и дополнять Международный кодекс классификации и номенклатуры вирусов (ICVCN; «Кодекс») и Устав, чтобы четко определить сферу компетенции ICTV (Отчет). дои : 10.13140/RG.2.2.26202.26565 .
- ^ Перейти обратно: а б с д Кунин Е.В., Доля В.В., Крупович М., Кун Дж.Х. (декабрь 2021 г.). «Вирусы, определяемые положением виросферы в пространстве репликатора» . Обзоры микробиологии и молекулярной биологии . 85 (4): e0019320. дои : 10.1128/MMBR.00193-20 . ПМЦ 8483706 . ПМИД 34468181 .
- ^ Кунин Е.В., Доля В.В. (июнь 2014 г.). «Вирусный мир как эволюционная сеть вирусов и бескапсульных эгоистичных элементов» . Обзоры микробиологии и молекулярной биологии . 78 (2): 278–303. дои : 10.1128/MMBR.00049-13 . ПМК 4054253 . ПМИД 24847023 .
- ^ Рэнкин DJ, Rocha EP, Brown SP (январь 2011 г.). «Какие черты передаются мобильным генетическим элементам и почему?» . Наследственность . 106 (1): 1–10. дои : 10.1038/hdy.2010.24 . ПМК 3183850 . ПМИД 20332804 .
- ^ Кроуфорд Д. (2011). Вирусы: очень краткое введение . Нью-Йорк: Издательство Оксфордского университета. п. 4 . ISBN 978-0199574858 .
- ^ Бриддон Р.В., Габриал С., Лин Н.С., Палукайтис П., Шольтхоф КБ, Веттен Х.Дж. «3 – Сателлиты и другие вирусозависимые нуклеиновые кислоты – Субвирусные агенты – Субвирусные агенты (2011)» . Международный комитет по таксономии вирусов (ICTV) . Архивировано из оригинала 13 января 2019 года.
- ^ Ди Серио Ф., Оуэнс Р.А., Ли С.Ф., Матушек Дж., Паллас В., Рэндлс Дж.В. и др. (ноябрь 2020 г.). «Вироиды» . Международный комитет по таксономии вирусов (ICTV). Архивировано из оригинала 2 декабря 2020 года.
- ^ Фешотт С., Гилберт С. (март 2012 г.). «Эндогенные вирусы: понимание вирусной эволюции и влияние на биологию хозяина» (PDF) . Обзоры природы. Генетика . 13 (4): 283–296. дои : 10.1038/nrg3199 . ПМИД 22421730 . S2CID 205485232 .
- ^ Петерс Ю.Э., Макарова К.С., Шмаков С., Кунин Е.В. (август 2017 г.). «Привлечение систем CRISPR-Cas с помощью Tn7-подобных транспозонов» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (35): Е7358–Е7366. Бибкод : 2017PNAS..114E7358P . дои : 10.1073/pnas.1709035114 . ПМЦ 5584455 . ПМИД 28811374 .
- ^ Таззиман С.Дж., Бонхеффер С. (декабрь 2013 г.). «Вероятность фиксации мобильных генетических элементов, таких как плазмиды» . Теоретическая популяционная биология . 90 : 49–55. дои : 10.1016/j.tpb.2013.09.012 . ПМИД 24080312 .
- ^ Гульельмини Дж., Кинтайс Л., Гарсильян-Барсиа, член парламента, де ла Крус Ф., Роча Е.П. (август 2011 г.). «Репертуар ICE у прокариот подчеркивает единство, разнообразие и повсеместность конъюгации» . ПЛОС Генетика . 7 (8): e1002222. дои : 10.1371/journal.pgen.1002222 . ПМК 3158045 . ПМИД 21876676 .
- ^ Возняк Р.А., Вальдор М.К. (август 2010 г.). «Интегративные и конъюгативные элементы: мозаичные мобильные генетические элементы, обеспечивающие динамический боковой поток генов». Обзоры природы. Микробиология . 8 (8): 552–563. дои : 10.1038/nrmicro2382 . ПМИД 20601965 . S2CID 21460836 .
- ^ Аухтунг Дж. М., Ли К. А., Гаррисон К. Л., Гроссман А. Д. (июнь 2007 г.). «Идентификация и характеристика репрессора иммунитета (ImmR), который контролирует мобильный генетический элемент ICEBs1 Bacillus subtilis» . Молекулярная микробиология . 64 (6): 1515–1528. дои : 10.1111/j.1365-2958.2007.05748.x . ПМЦ 3320793 . ПМИД 17511812 .
- ^ Тирумалай М.Р., Fox GE (сентябрь 2013 г.). «Элемент, подобный ICEBs1, может быть связан с чрезвычайной устойчивостью спор Bacillus pumilus SAFR-032 к радиации и высыханию». Экстремофилы . 17 (5): 767–774. дои : 10.1007/s00792-013-0559-z . ПМИД 23812891 . S2CID 8675124 .
- ^ Линк Л., Сойер Дж., Венкатешваран К., Николсон В. (февраль 2004 г.). «Чрезвычайная устойчивость спор к УФ-излучению изолятов Bacillus pumilus, полученных на сверхчистом объекте сборки космических кораблей». Микробная экология . 47 (2): 159–163. Бибкод : 2004MicEc..47..159L . дои : 10.1007/s00248-003-1029-4 . ПМИД 14502417 . S2CID 13416635 .
- ^ Ньюкомб Д.А., Шуергер А.С., Бенардини Дж.Н., Дикинсон Д., Таннер Р., Венкатешваран К. (декабрь 2005 г.). «Выживание микроорганизмов, связанных с космическим кораблем, в условиях искусственного марсианского УФ-облучения» . Прикладная и экологическая микробиология . 71 (12): 8147–8156. Бибкод : 2005ApEnM..71.8147N . дои : 10.1128/АЕМ.71.12.8147-8156.2005 . ПМЦ 1317311 . ПМИД 16332797 .
- ^ Кемпф М.Дж., Чен Ф., Керн Р., Венкатешваран К. (июнь 2005 г.). «Периодическое выделение устойчивых к перекиси водорода спор Bacillus pumilus на сборочном предприятии космического корабля». Астробиология . 5 (3): 391–405. Бибкод : 2005AsBio...5..391K . дои : 10.1089/ast.2005.5.391 . ПМИД 15941382 .
- ^ Де Чекко М., Крисчионе С.В., Петерсон А.Л., Неретти Н., Седиви Дж.М., Крейлинг Дж.А. (декабрь 2013 г.). «Мобильные элементы становятся активными и мобильными в геномах стареющих соматических тканей млекопитающих» . Старение . 5 (12): 867–883. дои : 10.18632/aging.100621 . ПМЦ 3883704 . ПМИД 24323947 .
- ^ Барреат Дж.Г., Кацуракис А. (май 2021 г.). Баттистуцци Ф.У. (ред.). «Филогеномика вирусоподобных мобильных генетических элементов позвоночных животных» . Молекулярная биология и эволюция . 38 (5): 1731–1743. дои : 10.1093/molbev/msa291 . ПМЦ 8097293 . ПМИД 33481003 .
- ^ Виден С.А., Бес И.С., Корешова А., Плиота П., Крогулл Д., Бурга А. (июнь 2023 г.). «Вирусоподобные транспозоны преодолевают видовой барьер и способствуют развитию генетической несовместимости». Наука . 380 (6652): eade0705. дои : 10.1126/science.ade0705 . ПМИД 37384706 . S2CID 250645873 .
- ^ Перейти обратно: а б Болахе С (03 августа 2023 г.). «Эгоистичная вирусоподобная ДНК может переносить гены между видами» . Журнал Кванта . Проверено 6 августа 2023 г.
- ^ Глюк-Талер Э., Ралстон Т., Конкель З., Окампос К.Г., Ганешан В.Д., Дорранс А.Е. и др. (май 2022 г.). Ларракуэнте А. (ред.). «Гигантские элементы звездолета мобилизуют вспомогательные гены в геномах грибов» . Молекулярная биология и эволюция . 39 (5). дои : 10.1093/molbev/msac109 . ПМЦ 9156397 . ПМИД 35588244 .
- ^ Пейс Дж.К., Гилберт С., Кларк М.С., Фешотт С. (ноябрь 2008 г.). «Повторный горизонтальный перенос ДНК-транспозона у млекопитающих и других четвероногих» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (44): 17023–17028. дои : 10.1073/pnas.0806548105 . ПМЦ 2579371 . ПМИД 18936483 .
- ^ «ДНК космического захватчика проникла в геномы млекопитающих» . Наука . 03.11.2008. Архивировано из оригинала 4 мая 2021 года . Проверено 6 августа 2023 г.
- ^ Энтони Дж.М., ван Марл Г., Опи В., Баттерфилд Д.А., Маллет Ф., Йонг В.В. и др. (октябрь 2004 г.). «Человеческий эндогенный ретровирусный гликопротеин, опосредованный индукцией окислительно-восстановительных реагентов, вызывает гибель олигодендроцитов и демиелинизацию». Природная неврология . 7 (10): 1088–1095. дои : 10.1038/nn1319 . ПМИД 15452578 . S2CID 9882712 .
- ^ «Нобелевская премия по физиологии и медицине 1983 года» . nobelprize.org . Проверено 14 июля 2010 г.
- ^ Кин EC (декабрь 2012 г.). «Парадигмы патогенеза: воздействие на мобильные генетические элементы болезней» . Границы клеточной и инфекционной микробиологии . 2 : 161. дои : 10.3389/fcimb.2012.00161 . ПМЦ 3522046 . ПМИД 23248780 .
Библиография
[ редактировать ]- Миллер У.Дж., Кэпи П., ред. (2004). Мобильные генетические элементы: протоколы и геномные приложения . Хумана Пресс. ISBN 978-1-58829-007-6 .
- Шапиро Дж.А. , изд. (1983). Мобильные генетические элементы . Академическая пресса. ISBN 978-0-12-638680-6 .