сетчатка

| |

| |
| Имена | |
|---|---|
| Название ИЮПАК
сетчатка
| |
| Систематическое название ИЮПАК
(2 E ,4 E ,6 E ,8 E )-3,7-Диметил-9-(2,6,6-триметилциклогекс-1-ен-1-ил)нона-2,4,6,8-тетраеналь | |
Другие имена
| |
| Идентификаторы | |
3D model ( JSmol )
|
|
| КЭБ | |
| ХимическийПаук | |
| Информационная карта ECHA | 100.003.760 |
ПабХим CID
|
|
| НЕКОТОРЫЙ | |
Панель управления CompTox ( EPA )
|
|
| Характеристики | |
| С 20 Н 28 О | |
| Молярная масса | 284.443 g·mol −1 |
| Появление | Оранжевые кристаллы петролейного эфира [ 1 ] |
| Температура плавления | От 61 до 64 ° C (от 142 до 147 ° F; от 334 до 337 К) [ 1 ] |
| Почти нерастворим | |
| Растворимость в жире | Растворимый |
| Родственные соединения | |
Родственные соединения
|
ретинол ; ретиноевая кислота ; бета-каротин ; дегидроретиналь ; 3-гидроксиретиналь; 4-гидроксиретиналь |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа).
| |
Ретиналь (также известный как ретинальдегид ) представляет собой полиеновый хромофор . Ретиналь, связанный с белками, называемыми опсинами , является химической основой зрительной фототрансдукции , стадии обнаружения света зрительного восприятия (зрения).
Некоторые микроорганизмы используют ретиналь для преобразования света в метаболическую энергию. Фактически, недавнее исследование предполагает, что большинство живых организмов на нашей планете около 3 миллиардов лет назад использовали ретиналь для преобразования солнечного света в энергию, а не хлорофилл . Поскольку сетчатка поглощает преимущественно зеленый свет и пропускает фиолетовый, это привело к появлению гипотезы пурпурной Земли . [ 2 ]
Ретиналь сам по себе считается формой витамина А , когда его едят животные. Существует множество форм витамина А, все из которых преобразуются в ретиналь, который без них невозможен. Количество различных молекул, которые могут быть преобразованы в ретиналь, варьируется от вида к виду. Ретиналь первоначально назывался ретиненом . [ 3 ] и был переименован [ 4 ] после того, как было обнаружено, что это витамина альдегид А. [ 5 ] [ 6 ]
Позвоночные животные потребляют ретиналь непосредственно из мяса или производят ретиналь из каротиноидов — либо из α-каротина , либо из β-каротина — оба из которых являются каротинами . Они также производят его из β-криптоксантина , разновидности ксантофилла . Эти каротиноиды должны быть получены из растений или других фотосинтезирующих организмов. Никакие другие каротиноиды не могут быть преобразованы животными в сетчатку. Некоторые плотоядные животные вообще не могут перерабатывать каротиноиды. Другие основные формы витамина А — ретинол и частично активная форма, ретиноевая кислота — могут вырабатываться из ретиналя.
Беспозвоночные, такие как насекомые и кальмары, используют в своей зрительной системе гидроксилированные формы ретиналя, которые образуются в результате преобразования других ксантофиллов .
Метаболизм витамина А
[ редактировать ]Живые организмы производят ретиналь путем необратимого окислительного расщепления каротиноидов. [ 7 ]
Например:
- бета-каротин + О 2 → 2 ретиналь,
катализируется бета-каротин-15,15'-монооксигеназой [ 8 ] или бета-каротин-15,15'-диоксигеназа. [ 9 ]
Так же, как каротиноиды являются предшественниками ретиналя, ретиналь является предшественником других форм витамина А. Ретиналь взаимопревращается с ретинолом , формой транспорта и хранения витамина А:
катализируется ретинолдегидрогеназами (RDH) [ 10 ] и алкогольдегидрогеназы (АДГ). [ 11 ]
Ретинол называют спиртом витамина А или, чаще, просто витамином А. Ретиналь также может окисляться до ретиноевой кислоты :
- сетчатка+НАД + + H 2 O → ретиноевая кислота + НАДН + Н + (катализируемый RALDH)
- ретиналь + O 2 + H 2 O → ретиноевая кислота + H 2 O 2 (катализируется ретинальоксидазой),
катализируется дегидрогеназами сетчатки [ 12 ] также известные как ретинальдегиддегидрогеназы (RALDH) [ 11 ] а также оксидазы сетчатки . [ 13 ]
Ретиноевая кислота, иногда называемая кислотой витамина А , является важной сигнальной молекулой и гормоном у позвоночных животных.
Зрение
[ редактировать ]Ретиналь представляет собой конъюгированный хромофор . В глазах млекопитающих сетчатка начинается с 11- цис -ретинальной конфигурации, которая — после захвата фотона правильной длины волны — выпрямляется в полностью транс -ретинальную конфигурацию. Это изменение конфигурации воздействует на белок опсин в сетчатке , который запускает химический сигнальный каскад, который приводит к восприятию света или изображений мозгом. Спектр поглощения хромофора зависит от его взаимодействия с белком опсина, с которым он связан, так что разные комплексы ретиналь-опсин будут поглощать фотоны разных длин волн (т. е. разных цветов света).
Опсины
[ редактировать ]
Ретиналь связан с опсинами , которые представляют собой рецепторы, связанные с G-белком (GPCR). [ 14 ] [ 15 ] Опсины, как и другие GPCR, имеют семь трансмембранных альфа-спиралей, соединенных шестью петлями. Они обнаружены в фоторецепторных клетках сетчатки глаза . Опсин в палочковидных клетках позвоночных — родопсин . Палочки образуют диски, в мембранах которых содержатся молекулы родопсина и которые полностью находятся внутри клетки. N -концевая головка молекулы простирается внутрь диска, а С-концевой хвост — в цитоплазму клетки. Опсины в конусных клетках — OPN1SW , OPN1MW и OPN1LW . Колбочки образуют неполные диски, являющиеся частью плазматической мембраны , поэтому N-концевая головка выходит за пределы клетки. В опсинах ретиналь ковалентно связывается с лизином. [ 16 ] в седьмой трансмембранной спирали [ 17 ] [ 18 ] [ 19 ] через базу Шиффа . [ 20 ] [ 21 ] Образование связи основания Шиффа включает удаление атома кислорода из ретиналя и двух атомов водорода из свободной аминогруппы лизина с образованием H 2 O. Ретинилиден представляет собой двухвалентную группу, образующуюся при удалении атома кислорода из ретиналя, поэтому опсины были названы ретинилиденами. белки .
Опсины представляют собой прототипы рецепторов, связанных с G-белком (GPCR). [ 22 ] Родопсин крупного рогатого скота, опсин палочковидных клеток, был первым GPCR, аминокислотная последовательность которого имела свою аминокислотную последовательность. [ 23 ] и 3D-структура (с помощью рентгеновской кристаллографии ). определена [ 18 ] Родопсин крупного рогатого скота содержит 348 аминокислотных остатков. Ретиналь связывается как хромофор с Lys. 296 . [ 18 ] [ 23 ] Этот лизин консервативен почти во всех опсинах, лишь немногие опсины утратили его в ходе эволюции . [ 24 ] Опсины без лизина, связывающего сетчатку, не светочувствительны. [ 25 ] [ 26 ] [ 27 ] Такие опсины могут иметь и другие функции. [ 26 ] [ 24 ]
Хотя млекопитающие используют ретиналь исключительно в качестве хромофора опсина, другие группы животных дополнительно используют четыре хромофора, тесно связанных с ретиналем: 3,4-дидегидроретиналь (витамин А 2 ), (3 R )-3-гидроксиретиналь, (3 S )-3- гидроксиретиналь (оба витамина А 3 ) и (4 R )-4-гидроксиретиналь (витамин А 4 ). Многие рыбы и земноводные используют 3,4-дидегидроретиналь, также называемый дегидроретиналем . За исключением подотряда двукрылых Cyclorrhapha ( так называемых высших мух), все исследованные насекомые используют ( R ) -энантиомер 3-гидроксиретиналя. ( R )-энантиомер следует ожидать, если 3-гидроксиретиналь производится непосредственно из каротиноидов ксантофилла . Циклорафаны, включая Drosophila , используют ( 3S )-3-гидроксиретиналь. [ 28 ] [ 29 ] кальмары-светлячки Было обнаружено, что используют ( 4R )-4-гидроксиретиналь.
Визуальный цикл
[ редактировать ]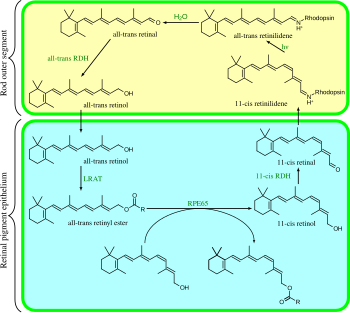
Зрительный цикл представляет собой круговой ферментативный путь , который является началом фототрансдукции. Он регенерирует 11- цис -ретиналь. Например, зрительный цикл палочек млекопитающих выглядит следующим образом:
- полностью транс- ретиниловый эфир + H 2 O → 11 -цис -ретинол + жирная кислота ; RPE65 ; изомерогидролазы [ 30 ]
- 11- цис -ретинол + НАД + → 11- цис -ретиналь + НАДН + Н + ; 11- цис -ретинолдегидрогеназы;
- 11- цис -ретиналь + апорходопсин → родопсин + H 2 O; образует связь основания Шиффа с лизином , -CH=N + ЧАС-;
- родопсин + hν → метародопсин II (т.е. 11- цис- фотоизомеризуется до полностью- транс ):
- (родопсин + hν → фотородопсин → батородопсин → люмирродопсин → метародопсин I → метародопсин II);
- метародопсин II + H 2 O → апорходопсин + all- транс -ретиналь;
- полностью транс -ретиналь + НАДФН + H + → полностью транс -ретинол + НАДФ + ; полностью транс ретинолдегидрогеназы ; -
- полностью транс -ретинол + жирная кислота → полностью транс- ретиниловый эфир + H 2 O; лецитин-ретинолацилтрансферазы (LRAT). [ 31 ]
Шаги 3, 4, 5 и 6 происходят во внешних сегментах палочек ; Этапы 1, 2 и 7 происходят в клетках пигментного эпителия сетчатки (RPE).
Изомерогидролазы RPE65 гомологичны бета-каротинмонооксигеназам; [ 7 ] гомологичный фермент ninaB у дрозофилы обладает как каротиноид-оксигеназной активностью, образующей сетчатку, так и активностью all- транс -11 -цис -изомеразы. [ 32 ]
Микробные родопсины
[ редактировать ]Полностью транс -ретиналь также является важным компонентом микробных опсинов, таких как бактериородопсин , каналородопсин и галородопсин , которые играют важную роль в бактерий и архей аноксигенном фотосинтезе . В этих молекулах свет заставляет полностью транс -ретиналь превращаться в 13 -цис -ретиналь, который затем снова превращается в полностью транс в темном состоянии -ретиналь. Эти белки эволюционно не связаны с опсинами животных и не являются GPCR; тот факт, что они оба используют ретиналь, является результатом конвергентной эволюции . [ 33 ]
История
[ редактировать ]Американский биохимик Джордж Уолд и другие наметили визуальный цикл к 1958 году. За свою работу Уолд получил долю Нобелевской премии по физиологии и медицине 1967 года вместе с Халданом Кеффером Хартлайном и Рагнаром Гранитом . [ 34 ]
См. также
[ редактировать ]- Гипотеза Фиолетовой Земли
- Сенсорная нервная система
- Визуальное восприятие
- Визуальная фототрансдукция
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Индекс Merck , 13-е издание, 8249
- ^ ДасСарма, Шиладитья; Швитерман, Эдвард В. (2018). «Ранняя эволюция фиолетовых пигментов сетчатки на Земле и значение биосигнатур экзопланет» . Международный журнал астробиологии . 20 (3) (опубликовано 11 октября 2018 г.): 241–250. arXiv : 1810.05150 . дои : 10.1017/S1473550418000423 . ISSN 1473-5504 . S2CID 119341330 .
- ^ Уолд, Джордж (14 июля 1934 г.). «Каротиноиды и цикл витамина А в зрении» . Природа . 134 (3376): 65. Бибкод : 1934Natur.134...65W . дои : 10.1038/134065a0 . S2CID 4022911 .
- ^ Вальд, Г. (11 октября 1968 г.). «Молекулярные основы зрительного возбуждения». Наука . 162 (3850): 230–9. Бибкод : 1968Sci...162..230W . дои : 10.1126/science.162.3850.230 . ПМИД 4877437 .
- ^ МОРТОН, РА; ГУДВИН, Т.В. (1 апреля 1944 г.). «Приготовление ретинена in vitro». Природа . 153 (3883): 405–406. Бибкод : 1944Natur.153..405M . дои : 10.1038/153405a0 . S2CID 4111460 .
- ^ ШАР, С; ГУДВИН, ТВ; МОРТОН, РА (1946). «Ретинен-1-витамин А альдегид». Биохимический журнал . 40 (5–6): ликс. ПМИД 20341217 .
- ^ Перейти обратно: а б фон Линтиг, Йоханнес; Фогт, Клаус (2000). «Заполнение пробелов в исследованиях витамина А: молекулярная идентификация фермента, расщепляющего бета-каротин до сетчатки» . Журнал биологической химии . 275 (16): 11915–11920. дои : 10.1074/jbc.275.16.11915 . ПМИД 10766819 .
- ^ Воггон, Вольф-Д. (2002). «Окислительное расщепление каротиноидов, катализируемое моделями ферментов и бета-каротин-15,15'-монооксигеназой» . Чистая и прикладная химия . 74 (8): 1397–1408. дои : 10.1351/pac200274081397 .
- ^ Ким, Ён Су; Ким, Нам Хи; Ём, Су-Джин; Ким, Сон-Вон; О, Док-Кун (2009). «Характеристика in vitro рекомбинантного белка Blh из некультивируемой морской бактерии как β-каротин-15,15'-диоксигеназы» . Журнал биологической химии . 284 (23): 15781–93. дои : 10.1074/jbc.M109.002618 . ПМК 2708875 . ПМИД 19366683 .
- ^ Лиден, М; Эрикссон, У (2006). «Понимание метаболизма ретинола: структура и функция ретинолдегидрогеназ» . Журнал биологической химии . 281 (19): 13001–04. дои : 10.1074/jbc.R500027200 . ПМИД 16428379 .
- ^ Перейти обратно: а б Дустер, Дж. (сентябрь 2008 г.). «Синтез ретиноевой кислоты и передача сигналов во время раннего органогенеза» . Клетка . 134 (6): 921–31. дои : 10.1016/j.cell.2008.09.002 . ПМК 2632951 . ПМИД 18805086 .
- ^ Лин, Мин; Чжан, Мин; Авраам, Майкл; Смит, Сьюзен М.; Наполи, Джозеф Л. (2003). «Мышиная дегидрогеназа сетчатки 4 (RALDH4), молекулярное клонирование, клеточная экспрессия и активность в биосинтезе 9-цис-ретиноевой кислоты в интактных клетках» . Журнал биологической химии . 278 (11): 9856–9861. дои : 10.1074/jbc.M211417200 . ПМИД 12519776 .
- ^ «ФЕРМЕНТ КЕГГ: 1.2.3.11 оксидаза сетчатки» . Проверено 10 марта 2009 г.
- ^ Кейси, Пи Джей; Гилман, А.Г. (февраль 1988 г.). «Участие G-белка в соединении рецептор-эффектор» . Журнал биологической химии . 263 (6): 2577–2580. дои : 10.1016/s0021-9258(18)69103-3 . ПМИД 2830256 . S2CID 38970721 .
- ^ Эттвуд, ТК; Финдли, JBC (1994). «Отпечатки пальцев, связанные с G-белком рецепторы». Белковая инженерия, проектирование и отбор . 7 (2): 195–203. дои : 10.1093/белок/7.2.195 . ПМИД 8170923 .
- ^ Баундс, Дерик (декабрь 1967 г.). «Место прикрепления сетчатки в родопсине». Природа . 216 (5121): 1178–1181. Бибкод : 1967Natur.216.1178B . дои : 10.1038/2161178a0 . ПМИД 4294735 . S2CID 1657759 .
- ^ Харгрейв, Пенсильвания; Макдауэлл, Дж. Х.; Кертис, Донна Р.; Ван, Джанет К.; Ющак, Элизабет; Фонг, Шао-Лин; Мохана Рао, JK; Аргос, П. (1983). «Структура бычьего родопсина». Биофизика структуры и механизма . 9 (4): 235–244. дои : 10.1007/BF00535659 . ПМИД 6342691 . S2CID 20407577 .
- ^ Перейти обратно: а б с Пальчевски К., Кумасака Т., Хори Т., Бенке К.А., Мотошима Х., Фокс Б.А. и др. (август 2000 г.). «Кристаллическая структура родопсина: рецептор, связанный с белком AG». Наука . 289 (5480): 739–45. Бибкод : 2000Sci...289..739P . CiteSeerX 10.1.1.1012.2275 . дои : 10.1126/science.289.5480.739 . ПМИД 10926528 .
- ^ Мураками М., Кояма Т. (май 2008 г.). «Кристаллическая структура родопсина кальмара». Природа . 453 (7193): 363–7. Бибкод : 2008Natur.453..363M . дои : 10.1038/nature06925 . ПМИД 18480818 . S2CID 4339970 .
- ^ Коллинз, Флорида (март 1953 г.). «Родопсин и индикаторный желтый». Природа . 171 (4350): 469–471. Бибкод : 1953Natur.171..469C . дои : 10.1038/171469a0 . ПМИД 13046517 . S2CID 4152360 .
- ^ Питт, ГАДЖ; Коллинз, Флорида; Мортон, РА; Сток, Полина (1 января 1955 г.). «Исследования родопсина. 8. Ретинилиденметиламин, аналог индикаторного желтого» . Биохимический журнал . 59 (1): 122–128. дои : 10.1042/bj0590122 . ПМК 1216098 . ПМИД 14351151 .
- ^ Лэмб, Т.Д. (1996). «Усиление и кинетика активации G-белкового каскада фототрансдукции» . Труды Национальной академии наук . 93 (2): 566–570. Бибкод : 1996PNAS...93..566L . дои : 10.1073/pnas.93.2.566 . ПМК 40092 . ПМИД 8570596 .
- ^ Перейти обратно: а б Овчинников, Ю.А. (8 ноября 1982 г.). «Родопсин и бактериородопсин: структурно-функциональные взаимоотношения» . Письма ФЭБС . 148 (2): 179–191. Бибкод : 1982FEBSL.148..179O . дои : 10.1016/0014-5793(82)80805-3 . ПМИД 6759163 . S2CID 85819100 .
- ^ Перейти обратно: а б Гюманн М., Портер М.Л., Бок М.Дж. (август 2022 г.). «Глюопсины: опсины без лизина, связывающего сетчатку» . Клетки . 11 (15): 2441. doi : 10.3390/cells11152441 . ПМЦ 9368030 . ПМИД 35954284 .
- ^ Катана, Радослав; Гуань, Чонглинь; Занини, Дамиано; Ларсен, Мэтью Э.; Хиральдо, Диего; Гертен, Барт Р.Х.; Шмидт, Кристофер Ф.; Бритт, Стивен Г.; Гёпферт, Мартин К. (сентябрь 2019 г.). «Независимая от хромофора роль опсин-апопротеинов в механорецепторах дрозофилы» . Текущая биология 29 (17): 2961–2969.e4. Бибкод : 2019CBio... 29E2961K дои : 10.1016/j.cub.2019.07.036 . ПМИД 31447373 . S2CID 201420079 .
- ^ Перейти обратно: а б Люнг, Николь Ю.; Тхакур, Дхананджай П.; Гурав, Адишти С.; Ким, Сан Хун; Ди Пицио, Антонелла; Нив, Маша Ю.; Монтелл, Крейг (апрель 2020 г.). «Функции опсинов во вкусе дрозофилы» . Современная биология . 30 (8): 1367–1379.е6. Бибкод : 2020CBio...30E1367L . дои : 10.1016/j.cub.2020.01.068 . ПМЦ 7252503 . ПМИД 32243853 .
- ^ Кумбаласири Т., Роллаг, доктор медицинских наук, Изольди М.К., Каструччи А.М., Провенсио I (март 2007 г.). «Меланопсин запускает высвобождение внутренних запасов кальция в ответ на свет». Фотохимия и фотобиология . 83 (2): 273–279. дои : 10.1562/2006-07-11-RA-964 . ПМИД 16961436 . S2CID 23060331 .
- ^ Секи, Такахару; Исоно, Кунио; Ито, Масаеши; Кацута, Юко (1994). «Мухи группы Cyclorrhapha используют (3S)-3-гидроксиретиналь в качестве уникального зрительного пигмента-хромофора». Европейский журнал биохимии . 226 (2): 691–696. дои : 10.1111/j.1432-1033.1994.tb20097.x . ПМИД 8001586 .
- ^ Секи, Такахару; Исоно, Кунио; Одзаки, Каору; Цукахара, Ясуо; Сибата-Кацута, Юко; Ито, Масаеши; Ириэ, Тошиаки; Катагири, Масанао (1998). «Метаболический путь образования хромофора зрительного пигмента у Drosophila melanogaster: All-транс (3S)-3-гидроксиретиналь образуется из полностью транс-ретиналя через (3R)-3-гидроксиретиналь в темноте» . Европейский журнал биохимии . 257 (2): 522–527. дои : 10.1046/j.1432-1327.1998.2570522.x . ПМИД 9826202 .
- ^ Моисеев Геннадий; Чен, Ин; Такахаси, Юсуке; Ву, Билл X.; Ма, Цзянь-син (2005). «RPE65 представляет собой изомерогидролазу ретиноидного зрительного цикла» . Труды Национальной академии наук . 102 (35): 12413–12418. Бибкод : 2005PNAS..10212413M . дои : 10.1073/pnas.0503460102 . ПМЦ 1194921 . ПМИД 16116091 .
- ^ Цзинь, Минхао; Юань, Цюань; Ли, Сунгари; Трэвис, Габриэль Х. (2007). «Роль LRAT в активности ретиноид-изомеразы и мембранной ассоциации Rpe65» . Журнал биологической химии . 282 (29): 20915–20924. дои : 10.1074/jbc.M701432200 . ПМЦ 2747659 . ПМИД 17504753 .
- ^ Оберхаузер, Витус; Вулстра, Олаф; Бангерт, Аннетт; фон Линтиг, Йоханнес; Фогт, Клаус (2008). «NinaB сочетает в себе активность каротиноидоксигеназы и ретиноидизомеразы в одном полипептиде» . Труды Национальной академии наук . 105 (48): 19000–5. Бибкод : 2008PNAS..10519000O . дои : 10.1073/pnas.0807805105 . ПМК 2596218 . ПМИД 19020100 .
- ^ Чен, Де-Лян; Ван, Гуан-юй; Сюй, Бин; Ху, Кунь-Шэн (2002). «Изомеризация полностью транс-13-цис сетчатки в светоадаптированном бактериородопсине при кислом pH». Журнал фотохимии и фотобиологии B: Биология . 66 (3): 188–194. Бибкод : 2002JPPB...66..188C . дои : 10.1016/S1011-1344(02)00245-2 . ПМИД 11960728 .
- ^ Нобелевская премия по физиологии и медицине 1967 г.
Дальнейшее чтение
[ редактировать ]- Фернальд, Рассел Д. (2006). «Проливая генетический свет на эволюцию глаз». Наука . 313 (5795): 1914–1918. Бибкод : 2006Sci...313.1914F . дои : 10.1126/science.1127889 . ПМИД 17008522 . S2CID 84439732 .
- Амора, Табита Л.; Рамос, Лавуазье С.; Галан, Дженни Ф.; Бирдж, Роберт Р. (2008). «Спектральная настройка темно-красных конусных пигментов» . Биохимия . 47 (16): 4614–20. дои : 10.1021/bi702069d . ПМЦ 2492582 . ПМИД 18370404 .
- Барлоу, HB; Левик, WR; Юн, М. (1971). «Реакция на одиночные кванты света в ганглиозных клетках сетчатки кошки». Исследование зрения . 11 (Приложение 3): 87–101. дои : 10.1016/0042-6989(71)90033-2 . ПМИД 5293890 .
- Бэйлор, округ Колумбия; Лэмб, ТД; Яу, К.В. (1979). «Реакция палочек сетчатки на одиночные фотоны» . Журнал физиологии . 288 : 613–634. дои : 10.1113/jphysicalol.1979.sp012716 . ПМЦ 1281447 . ПМИД 112243 .
- Фан, Цзе; Вудрафф, Майкл Л.; Силлуффо, Марианна С; Крауч, Розали К; Фейн, Гордон Л. (2005). «Активация трансдукции опсином в палочках мышей с нокаутом Rpe65, выращенных в темноте» . Журнал физиологии . 568 (1): 83–95. дои : 10.1113/jphysicalol.2005.091942 . ПМЦ 1474752 . ПМИД 15994181 .
- Хехт, Селиг; Шлаер, Саймон; Пиренн, Морис Анри (1942). «Энергия, кванты и видение» . Журнал общей физиологии . 25 (6): 819–840. дои : 10.1085/jgp.25.6.819 . ПМК 2142545 . ПМИД 19873316 .
- Кавагути, Рики; Ю, Цзямэй; Хонда, Джейн; Ху, Джейн; Уайтлегг, Джулиан; Пинг, Пейбэй; Виита, Патрик; Бок, Дин; Сунь, Хуэй (2007). «Мембранный рецептор ретинол-связывающего белка опосредует клеточное поглощение витамина А» . Наука . 315 (5813): 820–825. Бибкод : 2007Sci...315..820K . дои : 10.1126/science.1136244 . ПМИД 17255476 . S2CID 25258551 .
- Клоер, Дэниел П.; Рух, Сандра; Аль-Бабили, Салим; Бейер, Питер; Шульц, Георг Э. (2005). «Структура ретинальобразующей каротиноидной оксигеназы». Наука . 308 (5719): 267–269. Бибкод : 2005Sci...308..267K . дои : 10.1126/science.1108965 . ПМИД 15821095 . S2CID 6318853 .
- Ло, Донг-Ген; Сюэ, Тянь; Яу, Кинг-Вай (2008). «Как начинается видение: Одиссея» . Труды Национальной академии наук . 105 (29): 9855–9862. Бибкод : 2008PNAS..105.9855L . дои : 10.1073/pnas.0708405105 . ПМЦ 2481352 . ПМИД 18632568 . Хороший исторический обзор.
- Прадо-Кабреро, Альфонсо; Шерзингер, Дэниел; Авалос, Хавьер; Аль-Бабили, Салим (2007). «Биосинтез сетчатки у грибов: характеристика каротиноидоксигеназы CarX из Fusarium fujikuroi» . Эукариотическая клетка . 6 (4): 650–657. дои : 10.1128/EC.00392-06 . ПМК 1865656 . ПМИД 17293483 .
- Рэкер, Эфраим; Стокениус, Вальтер (1974). «Восстановление пурпурных мембранных везикул, катализирующих светоуправляемое поглощение протонов и образование аденозинтрифосфата» . Журнал биологической химии . 249 (2): 662–663. дои : 10.1016/S0021-9258(19)43080-9 . ПМИД 4272126 .
- Садекар, Сумедха; Раймонд, Джейсон; Бланкеншип, Роберт Э. (2006). «Сохранение отдаленно связанных мембранных белков: фотосинтетические реакционные центры имеют общее структурное ядро». Молекулярная биология и эволюция . 23 (11): 2001–2007. дои : 10.1093/molbev/msl079 . ПМИД 16887904 .
- Салом, Дэвид; Лодовски, Дэвид Т.; Стенкамп, Рональд Э.; Ле Тронг, Изольда; Гольчак, Марцин; Ястржебска, Беата; Харрис, Тим; Баллестерос, Хуан А.; Пальчевский, Кшиштоф (2006). «Кристаллическая структура фотоактивируемого депротонированного промежуточного продукта родопсина» . Труды Национальной академии наук . 103 (44): 16123–16128. Бибкод : 2006PNAS..10316123S . дои : 10.1073/pnas.0608022103 . ПМЦ 1637547 . ПМИД 17060607 .
- Шефер, Гюнтер; Энгельхард, Мартин; Мюллер, Волкер (1999). «Биоэнергетика архей» . Обзоры микробиологии и молекулярной биологии . 63 (3): 570–620. дои : 10.1128/MMBR.63.3.570-620.1999 . ПМЦ 103747 . ПМИД 10477309 .
- Шмидт, Хольгер; Курцер, Роберт; Эйзенрайх, Вольфганг; Шваб, Вильфрид (2006). «Каротиназа AtCCD1 Arabidopsis thaliana представляет собой диоксигеназу» . Журнал биологической химии . 281 (15): 9845–9851. дои : 10.1074/jbc.M511668200 . ПМИД 16459333 .
- Отправь, Роберт; Сундхольм, Даге (2007). «Лестница к коническому пересечению: компьютерное исследование изомеризации сетчатки». Журнал физической химии А. 111 (36): 8766–8773. Бибкод : 2007JPCA..111.8766S . дои : 10.1021/jp073908l . ПМИД 17713894 .
- Су, Чжи-Ин; Ло, Донг-Ген; Теракита, Акихиса; Ситида, Ёсинори; Ляо, Си-Вэнь; Казми, Мания А.; Сакмар, Томас П.; Яу, Кинг-Вай (2006). «Компоненты фототрансдукции теменного глаза и их потенциальное эволюционное значение». Наука . 311 (5767): 1617–1621. Бибкод : 2006Sci...311.1617S . дои : 10.1126/science.1123802 . ПМИД 16543463 . S2CID 28604455 .
- Вентер, Дж. Крейг ; Ремингтон, К; Гейдельберг, Дж. Ф.; Халперн, Алабама; Раш, Д; Эйзен, Дж. А.; Ву, Д; Полсен, я; и др. (2004). «Секвенирование экологического генома Саргассова моря». Наука . 304 (5667): 66–74. Бибкод : 2004Sci...304...66V . CiteSeerX 10.1.1.124.1840 . дои : 10.1126/science.1093857 . ПМИД 15001713 . S2CID 1454587 . Океаны полны родопсина 1 типа.
- Ван, Тао; Цзяо, Юйчэнь; Монтелл, Крейг (2007). «Вскрытие пути, необходимого для выработки витамина А и фототрансдукции дрозофилы» . Журнал клеточной биологии . 177 (2): 305–316. дои : 10.1083/jcb.200610081 . ПМК 2064138 . ПМИД 17452532 .
- Ващук, Стивен А.; Безерра, Аранди Г.; Ши, Личи; Браун, Леонид С. (2005). «Leptosphaeria rhodopsin: Бактериородопсин-подобный протонный насос эукариот» . Труды Национальной академии наук . 102 (19): 6879–6883. Бибкод : 2005PNAS..102.6879W . дои : 10.1073/pnas.0409659102 . ПМК 1100770 . ПМИД 15860584 .
- Ёкояма, Сёдзо; Радлвиммер, Ф. Бернхард (2001). «Молекулярная генетика и эволюция красного и зеленого цветового зрения у позвоночных» . Генетика . 158 (4): 1697–1710. дои : 10.1093/генетика/158.4.1697 . ПМЦ 1461741 . ПМИД 11545071 .
- Бриггс, Уинслоу Р.; Спудич, Джон Л., ред. (2005). Справочник фотосенсорных рецепторов . Уайли. ISBN 978-3-527-31019-7 .
- Уолд, Джордж (1967). «Нобелевская лекция: Молекулярная основа зрительного возбуждения» (PDF) . Проверено 23 февраля 2009 г.
Внешние ссылки
[ редактировать ]- Первые шаги видения - Национальный музей здравоохранения
- Зрение и молекулярные изменения, вызванные светом
- Анатомия сетчатки и зрительные возможности
- Retinal , Imperial College v-chemlib
| Базы данных органов управления : Национальные |
|---|