Ретикулярная формация
| Ретикулярная формация | |
|---|---|
 | |
 Траверс Секция медуллы Илонгата примерно в середине оливы. (Formatio reticularis grisea и формация reticularis alba, помеченная слева.) | |
| Подробности | |
| Расположение | Ствол мозга , гипоталамус и другие регионы |
| Идентификаторы | |
| латинский | формирование ретикулярного |
| Сетка | D012154 |
| Нейроналы | 1223 |
| Neurolex Id | NLX_143558 |
| TA98 | A14.1.00.021 A14.1.05.403 A14.1.06.327 |
| TA2 | 5367 |
| FMA | 77719 |
| Анатомические термины нейроанатомии | |
Ретикулярное образование представляет собой набор взаимосвязанных ядер в стволе мозга , который охватывает от нижнего конца мозгового мозга до верхнего конца среднего мозга . [ 2 ] Нейроны ретикулярной формирования составляют сложный набор нейронных сетей в ядре ствола мозга. [ 3 ] Это не анатомически хорошо определен, потому что включает в себя нейроны, расположенные в разных частях мозга .
Ретикулярное образование функционально делится на восходящие пути к коре в восходящей ретикулярной активационной системе ( ARAS ), а также нисходящие пути к спинному мозгу в нисходящей ретикулярной системе через ретикулоспинальные тракты. [ 4 ] [ 5 ] [ 6 ] [ 7 ] Из -за своей степени вдоль ствола мозга он может быть разделен на различные области, такие как ретикулярное образование среднего мозга, центральная мезоэнцефатическая ретикулярная образования, ретикулярная ретикулярная ретикуляция, ретикулярная ретикулярная ретикуляция парамейна, дорсолатеральная рекордная резация и медуллярная проценкура. [ 8 ]
Нейроны ARAS в основном действуют как включение/выключение в кору головного мозга и, следовательно, играют решающую роль в регулировании бодрствования ; Поведенческое возбуждение и сознание функционально связаны в ретикулярной формировании с использованием ряда систем возбуждения нейротрансмиттера. Общие функции ретикулярного образования являются модулирующими и премоторными, [ А ] включает в себя соматический моторный контроль, сердечно -сосудистый контроль, боли, сон и сознание и привыкание. [ 9 ] Модуляторные функции в основном обнаруживаются в ростральном секторе ретикулярной формирования, а премоторные функции локализованы в нейронах в более каудальных областях.
Ретикулярное образование разделено на три столбца: ядра рафе (медиана), гигантоцеллюлярные ретикулярные ядра (медиальная зона) и парвоцеллюлярные ретикулярные ядра (боковая зона). Ядра рафе являются местом синтеза нейротрансмиттера серотонина , который играет важную роль в регуляции настроения. Гигантоцеллюлярные ядра участвуют в моторной координации. Парвоцеллюлярные ядра регулируют выдох . [ 10 ]
Ретикулярная формация имеет важное значение для управления некоторыми основными функциями более высоких организмов. Он филогенетически старый и обнаружен у нижних позвоночных . [ 2 ]
Структура
[ редактировать ]
Образование гуманитарной ретикуляции состоит из почти 100 ядер и содержит много проекций в переднем мозге , стволе мозга и мозжечка , среди других регионов. [ 5 ] Он включает в себя ретикулярные ядра , ретикулоталамические проекционные волокна, диффузные таламокортикальные проекции , восходящие холинергические проекции , нисходящие нехолинергические проекции и нисходящие рецензионные проекции. [ 6 ] Ретикулярная формация также содержит две основные нейронные подсистемы : восходящая ретикулярная активационная система и нисходящие ретикулоспинальные тракты, которые опосредуют различные когнитивные и физиологические процессы. [ 5 ] [ 6 ] Это было функционально расщеплено как сагиттально , так и коронально .
Традиционно ретикулярные ядра делятся на три столбца: [ Цитация необходима ]
- В срединной колонке - ядра рафе
- В медиальной колонке - гигантоцеллюлярные ядра (из -за большего размера клеток)
- В боковой колонке - парвоцеллюлярные ядра (из -за меньшего размера клеток)
Первоначальная функциональная дифференциация была разделением каудальной и ростральной . Это было основано на наблюдении, что поражение ростральной ретикулярной образования вызывает гиперсомнию в мозге кошки. Напротив, поражение более каудальной части ретикулярной формирования приводит к бессоннице у кошек. Это исследование привело к идее, что каудальная часть ингибирует ростральную часть ретикулярного образования. [ Цитация необходима ]
Сагиттальное разделение выявляет более морфологические различия. Ядра рафе образуют гребень в середине ретикулярного образования, и, непосредственно к его периферии, существует разделение, называемое медиальной ретикулярной образованием. Медиальная RF является большим и имеет давно восходящие и нисходящие волокна, и окружен боковой ретикулярной образованием. Боковой РФ находится близко к моторным ядрам черепных нервов и в основном опосредует их функции. [ Цитация необходима ]
Медиальная и боковая ретикулярная формация
[ редактировать ]Медиальная ретикулярная формация и боковое ретикулярное образование представляют собой две колонны ядер с четко определенными границами, которые посылают проекции через мозговое вещество и в средний мозг . Ядра могут быть дифференцированы по функциям, типам клеток и проекциям эфферентных или афферентных нервов. Двигаясь каудально из рострального среднего мозга , на месте ростральных поне и среднего мозга, медиальный РЧ становится менее заметным, а боковой РФ становится более заметным. [ Цитация необходима ]
На бокам медиальной ретикулярной образования существует его боковой двоюродный брат, который особенно выражен в ростральном мозговом и каудальном поне. Из этой области весны черепные нервы, в том числе очень важный блуждающий нерв . [ нужно разъяснения ] Латеральный RF известен своими ганглионами и областями интернейронов вокруг черепных нервов , которые служат для опосредования их характерных рефлексов и функций.
Основные подсистемы
[ редактировать ]Подсистемы ретикулярного образования являются восходящей ретикулярной активационной системой и нисходящей ретикулярной системой. [ 6 ]
Восходящая ретикулярная активационная система
[ редактировать ]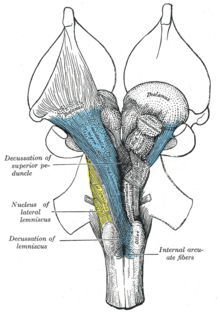
Восходящая ретикулярная активирующая система (ARAS), также известная как модуляторная система управления экстратхаламической контролем или просто ретикулярная активирующая система (RAS), представляет собой набор подключенных ядер в мозге позвоночных, которые отвечают за регулирование переходов бодрствования и бодрствования сна . Арас находится в ретикулярном образовании среднего мозга. [ 11 ] Он в основном состоит из различных ядер в таламусе / гипоталамусе и ряд дофаминергических , норадренергических , серотонинергических , гистаминергических , холинергических и глутаматергических ядер головного мозга. [ 5 ] [ 12 ] [ 13 ] [ 14 ]
Структура
[ редактировать ]ARAS состоит из нескольких нейронных цепей, соединяющих дорсальную часть заднего среднего мозга , и вентральных полов с корой головного мозга через различные пути, которые проецируются через таламус и гипоталамус . [ 5 ] [ 13 ] [ 14 ] ARAS представляет собой набор различных ядер - более 20 с каждой стороны в верхней стволе мозга, поне, мозговой сети и задней гипоталамуса. [ 11 ] Нейротрансмиттеры, которые высвобождают эти нейроны, включают дофамин , норэпинефрин , серотонин , гистамин , ацетилхолин и глутамат . [ 5 ] [ 12 ] [ 13 ] [ 14 ] Они оказывают корковое влияние через прямые аксональные проекции и косвенные проекции через реле таламуса. [ 13 ] [ 14 ] [ 11 ]
Таламический путь состоит в основном из холинергических нейронов в понтине -тегментаме , тогда как гипоталамический путь состоит в основном из нейронов, которые высвобождают моноротрансмиттеры , а именно дофамин, норпинефрин, серотонин и гистамин. [ 5 ] [ 12 ] Глутаматные нейроны в арасах были идентифицированы гораздо недавно по сравнению с моноаминергическими и холинергическими ядрами; [ 15 ] Глутаматергический компонент ARAS включает в себя одно ядро в гипоталамусе и различные ядра ствола мозга. [ 13 ] [ 15 ] [ 16 ] Нейроны орексина бокового гипоталамуса иннервируют каждый компонент восходящей ретикулярной активационной системы и координатной активности во всей системе. [ 14 ] [ 17 ] [ 18 ]
| Тип ядра | Соответствующие ядра, которые опосредуют возбуждение | Источники |
|---|---|---|
| Дофаминергические ядра | [ 5 ] [ 12 ] [ 13 ] [ 14 ] | |
| Норадренергические ядра |
|
[ 5 ] [ 12 ] [ 14 ] |
| Серотонинергические ядра | [ 5 ] [ 12 ] [ 14 ] | |
| Гистаминергические ядра | [ 5 ] [ 12 ] [ 19 ] | |
| Холинергические ядра | [ 5 ] [ 13 ] [ 14 ] [ 15 ] | |
| Глутаматергические ядра |
|
[ 13 ] [ 14 ] [ 15 ] [ 16 ] [ 19 ] [ 20 ] |
| Таламические ядра | [ 5 ] [ 13 ] [ 21 ] |
ARAS состоит из эволюционно древних областей мозга, которые имеют решающее значение для выживаемости животного и защищены в неблагоприятные периоды, например, в течение ингибирующих периодов гипноза животных, также известных как TotstellReflex . [ 22 ] Восходящая ретикулярная активационная система, которая посылает нейромодулирующие проекции в кору - в основном соединяется с префронтальной корой . [ 23 ] Похоже, что существует низкая связь с двигательными областями коры. [ 23 ]
Функция
[ редактировать ]Сознание
[ редактировать ]Восходящая ретикулярная активационная система является важным фактором, способствующим состоянию сознания . [ 11 ] Считается, что восходящая система способствует бодрствованию, характеризующемуся кортикальным и поведенческим возбуждением. [ 7 ]
Регулирование переходов сна бодрствования
[ редактировать ]Основная функция ARA - это модифицировать и потенциальный таламический и кортикальную функцию так, чтобы наступила дезинхронизация электроэнцефалограммы (ЭЭГ). [ B ] [ 25 ] [ 26 ] Существуют четкие различия в электрической активности мозга во время периодов бодрствования и сна: быстрый взрыв мозга с низким напряжением (Desynchronization EEG) связаны с бодрствованием и восстановлением (которые электрофизиологически сходны); Медленные волны высокого напряжения обнаруживаются во время не снового сна. Вообще говоря, когда нейроны реле таламуса находятся в режиме взрыва, ЭЭГ синхронизируется, и когда они находятся в тонизированном режиме, он десинхронизируется. [ 26 ] Стимуляция ARAS вызывает десинхронизацию ЭЭГ путем подавления медленных кортикальных волн дельта , ( Гц ) 0,3–1 Полем [ 17 ]
Физиологические изменения от состояния глубокого сна до бодрствования обратимы и опосредованы арас. [ 27 ] Вентролатеральное преоптическое ядро (VLPO) гипоталамуса ингибирует нейронные цепи, ответственные за состояние бодрствования, а активация VLPO способствует началу сна. [ 28 ] Во время сна нейроны в ARA будут иметь гораздо более низкую скорость стрельбы; И наоборот, у них будет более высокий уровень активности во время состояния бодрствования. [ 29 ] Чтобы мозг мог спать, должно быть снижение восходящей афферентной активности, достигающей коры путем подавления ARA. [ 27 ] Дисфункция паравентрикулярного ядра гипоталамуса может привести к сонливости до 20 часов в день. [ 30 ]
Внимание
[ редактировать ]ARAS также помогает опосредовать переходы от расслабленного бодрствования к периодам высокого внимания . [ 21 ] Существует повышенное региональное кровоток (предположительно, указывает на повышенную меру нейрональной активности) в ретикулярном образовании среднего мозга (MRF) и внутриламин -ядрах таламуса во время задач, требующих повышенной бдительности и внимания. [ Цитация необходима ]
Клиническое значение ARAS
[ редактировать ]Массовые поражения в ядрах Арас могут вызвать потерю сознания. [ 11 ] [ 31 ] Двусторонний ущерб ядрам арас может привести к коме или смерти. [ 32 ]
Прямая электрическая стимуляция ARA производит обезболивающие реакции у кошек и вызывает устные сообщения о боли у людей. [ Цитация необходима ] Восходящая ретикулярная активация у кошек может производить мидриаз , [ 33 ] что может быть результатом длительной боли. Эти результаты свидетельствуют о некоторой взаимосвязи между схемами ARAS и физиологическими боли. [ 33 ]
Некоторые патологии ARA могут быть связаны с старением , так как, по -видимому, существует общее снижение реактивности ARA с прогрессирующими годами. [ 34 ] Изменения в электрической связи [ C ] были предложены для учета некоторых изменений в активности ARAS: если бы связь была пониженной регуляцией , было бы соответствующее снижение синхронизации с более высокой частотой (гамма-полоса). И наоборот, усиление электрической связи увеличит синхронизацию быстрых ритмов, которые могут привести к увеличению возбуждения и быстрого сна. [ 36 ] В частности, нарушение ARA было вовлечено в следующие расстройства:
- Наколепсия : поражения вдоль педункулопонтина (PPN) и в латеодорсальных сегментарных ядрах связаны с нарколепсией. [ 37 ] Существует значительная понижающая регуляция выхода PPN и потери пептидов орексина, способствуя чрезмерной дневной сонливостью, которая характерна для этого расстройства. [ 17 ]
- Прогрессивный наддерный паралич (PSP): дисфункция передачи сигналов оксида азота была вовлечена в развитие PSP. [ 38 ]
- Болезнь Паркинсона : недостатки сна распространены у Паркинсона. В основном это дофаминергическое заболевание, но холинергические ядра также истощены. Дегенерация в ARA начинается на ранней стадии процесса заболевания. [ 37 ]
Влияния развития
[ редактировать ]Существует несколько потенциальных факторов, которые могут отрицательно влиять на развитие восходящей ретикулярной активационной системы:
- Преждевременные роды : [ 39 ] Независимо от веса при рождении или недели беременности, преждевременное рождение вызывает постоянное вредное воздействие на преотъемлемые (возбуждение и аномалии бодрствования сон), внимание (время реакции и сенсорное стробирование) и корковые механизмы на протяжении всего развития.
- Курение во время беременности : [ 40 ] Известно, что пренатальное воздействие сигаретного дыма вызывает длительное возбуждение, внимание и когнитивный дефицит у людей. Это воздействие может индуцировать активизацию никотиновых рецепторов α4β2 на клетках пенонкулопонтинного ядра (PPN), что приводит к повышению тонической активности, мембранному поколению и катионному току, активируемому гиперполяризацией . Эти основные нарушения внутренних мембранных свойств нейронов PPN приводят к повышению уровня возбуждения и сенсорного стробирования , дефициты (продемонстрированные уменьшенным количеством привыкания к повторным слуховым стимулам). Предполагается, что эти физиологические изменения могут усилить дисрегуляцию внимания в более позднем возрасте.
Нисходящая ретикулоспинальная система
[ редактировать ]Ретикулоспинальные тракты , представляют собой экстрапирамидные двигательные тракты, которые спускаются из ретикулярного образования [ 41 ] В двух трактах, чтобы действовать на моторных нейронах, поставляющих сгибатели и разгибатели с туловищем и проксимальные конечности. Ретикулоспинальные тракты участвуют в основном в локомоции и постуральном контроле, хотя у них также есть и другие функции. [ 42 ] Нисходящие ретикулоспинальные тракты являются одним из четырех основных корковых путей к спинному мозгу для опорно -двигательной активности. Ретикулоспинальные тракты работают с другими тремя путями, чтобы дать скоординированный контроль над движением, включая деликатные манипуляции. [ 41 ] Четыре пути могут быть сгруппированы в два основных системных путей - медиальную систему и боковую систему. Медиальная система включает в себя ретикулоспинальный тракт и вестибулоспинальный тракт и обеспечивает контроль над осанкой. Кортикоспинальный тракт и руброспинальный тракт принадлежат к боковой системе, которая обеспечивает тонкий контроль движения. [ 41 ]
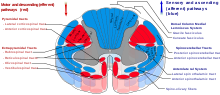
Ретикулоспинальные тракты представляют собой медиальные ретикулоспинальные тракты и латеральный ретикулоспинальный тракт. [ Цитация необходима ]
- Медиальный ретикулоспинальный тракт (понтин) отвечает за захватывающие антигравитации, разгибательские мышцы. Волокна этого тракта возникают из хвостового ретикулярного ядра понтины и перорального понтитинского ретикулярного ядра и проецируют на пластину VII и пластинку VIII спинного мозга. [ Цитация необходима ]
- Боковой ретикулоспинальный тракт (медуллярный) отвечает за ингибирование возбуждающих осевых мышц движения. Это также отвечает за автоматическое дыхание. Волокна этого тракта возникают из медуллярного ретикулярного образования, в основном из гигантоцеллюлярного ядра , и спускаются по длине спинного мозга в передней части латеральной белой колонны (funiculus). Тракт заканчивается в пластинке VII в основном с некоторыми волокнами, заканчивающимися в пластинке IX спинного мозга. [ Цитация необходима ]
Восходящим сенсорным трактом, передавающим информацию в противоположном направлении, является спиноретичным трактом .
Функция
[ редактировать ]- Интегрирует информацию из моторных систем для координации автоматических движений локомоции и осанки
- Облегчает и ингибирует добровольное движение; влияет на мышечный тонус
- Помновляет вегетативные функции
- Модулирует боли
- Влияет на кровоток к боковому геникуляционному ядру таламуса. [ 43 ]
Клиническое значение
[ редактировать ]Ретикулоспинальные тракты обеспечивают путь, по которому гипоталамус может контролировать симпатический отток тораколумба и парасимпатический сакральный отток. [ Цитация необходима ]
Две основные нисходящие системы, несущие сигналы от ствола мозга и мозжечка до спинного мозга, могут вызвать автоматический постуральный отклик для баланса и ориентации: вестибулоспинальные тракты из вестибулярных ядер и ретикулоспинальные тракты от поннов и медулла. Поражения этих трактов приводят к глубокой атаксии и постуральной нестабильности . [ 44 ]
Физическое или сосудистое повреждение ствола ствола мозга, отключающее красное ядро (средний мозг), и вестибулярные ядра (PONS) могут вызывать жесткость децеребрата , что имеет неврологический признак повышенного мышечного тонуса и гиперактивных растяжек . Отвечая на поразительный или болезненный стимул, как руки, так и ноги простираются и поворачиваются внутренне. Причиной является тоническая активность боковых вестибулоспинальных и ретикулоспинальных трактов, стимулирующих мотонейроны разгибателей без ингибирования из руброспинальных трактов . [ 45 ]
Повреждение ствола мозга над уровнем ядра красного цвета может вызвать жесткость декартата . Отвечая на поразительный или болезненный стимул, руки сгибаются и вытягивают ноги. Причиной является красное ядро через руброспинальный тракт, противодействующий возбуждению разгибателя Motorneuron от латеральных вестибулоспинальных и ретикулоспинальных трактов. Поскольку руброспинальный тракт только распространяется на шейный спинной мозг, он в основном действует на руках, захватывая мышцы сгибателей и ингибируя разгибателей, а не ноги. [ 45 ]
Повреждение мозгового мозга ниже вестибулярных ядер может вызвать нексустный паралич , гипотонию , потерю дыхательного привода и квадриплегию . Нет никаких рефлексов, напоминающих ранние стадии спинального шока из -за полной потери активности у моторнеронов, поскольку больше нет тонической активности, возникающей из боковых вестибулоспинальных и ретикулоспинальных трактов. [ 45 ]
История
[ редактировать ]Термин «ретикулярная формация» был придуман в конце 19 -го века Отто Дейтерами , совпадая с Рамона и Каджала доктриной нейронов . Аллан Хобсон в своей книге заявляет, что ретикулярная формация пересмотрела , что название является этимологическим остатком из упавшей эпохи совокупного поля в нейронных науках. Термин « ретикулум » означает «сетевую структуру», что и ретикулярное образование напоминает на первый взгляд. Он был описан как слишком сложный для изучения, либо недифференцированной части мозга без организации вообще. Эрик Кандель описывает ретикулярную формацию как организованную аналогичным образом промежуточного серого вещества спинного мозга. Эта хаотическая, свободная и сложная форма организации - это то, что отключило многих исследователей, чтобы посмотреть дальше в эту конкретную область мозга. [ Цитация необходима ] В клетках отсутствуют четкие ганглионические границы, но имеют четкую функциональную организацию и различные типы клеток. Термин «ретикулярная формация» больше редко используется, кроме как говорить в общих чертах. Современные ученые обычно относятся к отдельным ядрам, которые составляют ретикулярную формацию. [ Цитация необходима ]
Moruzzi и Magoun сначала исследовали нейронные компоненты, регулирующие механизмы сна бодрствования мозга в 1949 году. Физиологи предположили, что некоторая структура глубоко внутри мозга контролировала умственное бодрствование и бдительность. [ 25 ] Считалось, что бодрствование зависело только от прямого приема афферентных (сенсорных) стимулов в коре головного мозга . [ Цитация необходима ]
Поскольку прямая электрическая стимуляция мозга могла моделировать электрокортикальные реле, Магун использовал этот принцип, чтобы продемонстрировать в двух отдельных областях ствола мозга кошки, как производить бодрствование от сна. Сначала он стимулировал восходящие соматические и слуховые пути; Во -вторых, серия «восходящих реле из ретикулярного образования нижнего ствола головного мозга через средний мозг тегментам , подпаламус и гипоталамус во внутреннюю капсулу ». [ 46 ] Последнее представляло особый интерес, так как эта серия реле не соответствовала каким -либо известным анатомическим путям для трансдукции сигнала бодрствования и была придумана восходящая ретикулярная активирующая система (ARAS). [ Цитация необходима ]
Затем значение этой недавно выявленной ретранс -системы оценивалось путем размещения поражений в медиальных и боковых частях передней части среднего мозга . Кошки с мезоэнцефалическими прерываниями в ARA, вошли в глубокий сон и показали соответствующие мозговые волны. В качестве альтернативного способа, кошки с аналогичными перерывами на восходящие слуховые и соматические пути демонстрировали нормальный сон и бодрствование, и могут быть пробуждены физическими стимулами. Поскольку эти внешние стимулы будут заблокированы по пути в кору перерывами, это указывало на то, что восходящая передача должна проходить через недавно обнаруженные арас. [ Цитация необходима ]
Наконец, Magoun зарегистрировал потенциалы в медиальной части ствола мозга и обнаружил, что слуховые стимулы непосредственно стреляют в части ретикулярной активирующей системы. в одиночном шоке Кроме того, стимуляция седалищного нерва также активировала медиальную ретикулярную образование, гипоталамус и таламус . Возбуждение ARAS не зависело от дальнейшего распространения сигналов через мозжечковые схемы, так как те же результаты были получены после дезертибелляции и декартации. Исследователи предположили, что колонна ячеек, окружающих ретикулярную ретикулярную образу среднего мозга, получила вклад от всех восходящих трактов ствола мозга и передала эти афференты в кору и, следовательно, регулируемый бодрствование. [ 46 ] [ 27 ]
Смотрите также
[ редактировать ]Сноски
[ редактировать ]- ^ Предмоторная функция, как в интеграции сенсорных сигналов обратной связи с командами из верхних моторных нейронов и глубоких ядер мозжечка , и организации эфферентной активности нижнего висцерального моторного мотора и некоторых соматических моторных нейронов в стволе мозга и спинного мозга . [ 3 ]
- ^ ЭЭГ Электрод на коже головы измеряет активность очень большого количества пирамидных нейронов в нижней области мозга. Каждый нейрон генерирует небольшое электрическое поле, которое меняется со временем. В состоянии сна, нейроны активируются примерно в одно и то же время, а волна ЭЭГ, представляющая суммирование электрических полей нейронов, имеет тенденцию находиться в фазе и имеет более высокую амплитуду, и, следовательно, это «синхронизировано». В состоянии бодрствования они не активируются одновременно из-за нерегулярных или внефазных входов, волна ЭЭГ, представляющая алгебраическую сумму, будет иметь меньшую амплитуду и, следовательно, «дисинхронизированную». [ 24 ]
- ^ Электрическая связь - это пассивный поток электрического тока из одной ячейки в соседнюю клетку через щелевые соединения , такие как клетки в сердечной мышце или нейроны с электрическими синапсами . Электрически связанные ячейки синхронно стреляют, потому что генерируемые токи в одной ячейке быстро распространяются на другие ячейки. [ 35 ]
Ссылки
[ редактировать ]- ^ Грей, Генри. «Рис. 701: Генри Грей (1825–1861). Анатомия человеческого тела. 1918» . Bartleby.com. Архивировано из оригинала 2018-04-21 . Получено 2019-09-12 .
- ^ Jump up to: а беременный Brodal, per (2010). Центральная нервная система: структура и функция (4 -е изд.). Оксфорд: издательство Оксфордского университета. п. 373. ISBN 9780195381153 .
- ^ Jump up to: а беременный Purves, Dale (2011). Нейробиология (5. изд.). Сандерленд, Массачусетс: Синауэр. С. 390–395. ISBN 978-0878936953 .
- ^ Сингх, Вишрам (2014). Объем анатомии объем III . п. 372. ISBN 9788131237274 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м Iwańczuk W, Guźniczak P (2015). «Нейрофизиологические основы сна, возбуждения, осознания и явлений сознания. Часть 1». интенсивный Анастроил 47 (2): 162–167. doi : 10.5603/ait.2015.0015 . PMID 25940332 .
Восходящая ретикулярная активационная система (ARAS) отвечает за устойчивое состояние бодрствования. Он получает информацию от сенсорных рецепторов различных модальностей, передаваемых через спиноретикулярные пути и черепные нервы (тригемидуальный нерв - полимодальные пути, обонятельный нерв, зрительный нерв и вестибулокохлеарный нерв - мономодальные пути). Эти пути прямо или косвенно достигают таламуса через медиальную колонну ядер ретикулярных образования (магноцеллюлярные ядра и ретикулярные ядра понтине -тегментам). Ретикулярная активирующая система начинается в дорсальной части заднего среднего мозга и передних полов, продолжается в диоэфалоне, а затем делятся на две части, достигающие таламуса и гипоталамуса, которые затем проецируются в кору головного мозга (рис. 1). В проекции таламуса преобладают холинергические нейроны, происходящие из педункулопонтинового сегментарного ядра понса и среднего мозга (PPT) и лакообразного сегментарного ядра посонов и среднего мозга (LDT) ядер [17, 18]. Гипоталамическая проекция включает в себя норадренергические нейроны локуса coeruleus (LC) и серотонинергических нейронов дорсальных и медианных ядер рафе (DR), которые проходят через латеральный гипоталамус и достигают аксонов гистаминергического туберо-мимиллярного ядра (TMN), вместе формируют путь, простирающийся в передний мозг, кору и гиппокамп. Корковое возбуждение также использует дофаминергические нейроны субстанции NIGRA (SN), вентральной области (VTA) и периакредуктальной серой области (PAG). Меньшее количество холинергических нейронов Понс и среднего мозга посылают прогнозы на передний мозг вдоль вентрального пути, обходя таламус [19, 20].
- ^ Jump up to: а беременный в дюймовый Августин младший (2016). «Глава 9: ретикулярная формация» . Нейроанатомия человека (2 -е изд.). Джон Уайли и сыновья. С. 141–153. ISBN 978-1119073994 Полем Архивировано из оригинала 4 мая 2018 года . Получено 4 сентября 2017 года .
- ^ Jump up to: а беременный Джонс, Бэй (2008). «Модуляция кортикальной активации и поведенческого возбуждения холинергическими и орексинергическими системами». Анналы нью -йоркской академии наук . 1129 (1): 26–34. BIBCODE : 2008NASA1129 ... 26J . doi : 10.1196/annals.1417.026 . PMID 18591466 . S2CID 16682827 .
- ^ Ван, н; Перкинс, E; Чжоу, л; Уоррен, с; Май, PJ (2017). «Подключения ретикулярного образования, лежащие в основе горизонтального взгляда: центральная мезоэнцефальная ретикулярная образования (CMRF) в качестве канала для сигнала колликулярного саккада» . Границы в нейроанатомии . 11 : 36. doi : 10.3389/fnana.2017.00036 . PMC 5403835 . PMID 28487639 .
- ^ Саладин, Кеннет С. (2011). Анатомия человека (3 -е изд.). Нью-Йорк: МакГроу-Хилл. п. 410. ISBN 9780071222075 .
- ^ «Мозг сверху донизу» . Thebrain.mcgill.ca. Архивировано с оригинала 2016-04-23 . Получено 2016-04-28 .
- ^ Jump up to: а беременный в дюймовый и Squire L (2013). Фундаментальная нейробиология (4 -е изд.). Амстердам: Elsevier/Academic Press. п. 1095. ISBN 978-0123858702 .
- ^ Jump up to: а беременный в дюймовый и фон глин Malenka RC, Nestler EJ, Hyman SE (2009). «Глава 12: Сон и возбуждение». В Сидоре А, Браун Р.Ю. (ред.). Молекулярная нейрофармакология: основание для клинической нейробиологии (2 -е изд.). Нью-Йорк: McGraw-Hill Medical. п. 295. ISBN 978-0071481274 Полем
RAS представляет собой сложную структуру, состоящую из нескольких различных цепей, включая четыре моноаминергические пути ... путь норэпинефрина происходит из локуса Ceruleus (LC) и родственных ядер ствола мозга; Серотонинергические нейроны происходят из ядер Рафе в стволе мозга; Дофаминергические нейроны происходят в вентральной области сегментарной области (VTA); и гистаминергический путь происходит из нейронов в туберкулетном ядре (TMN) заднего гипоталамуса. Как обсуждалось в главе 6, эти нейроны широко проецируются по всему мозгу от ограниченных коллекций клеточных тел. Норепинефрин, серотонин, дофамин и гистамин имеют сложные модулирующие функции и, как правило, способствуют бодрствованию. PT в стволе мозга также является важным компонентом ARAS. Активность PT-холинергических нейронов (клетки REM-на) способствует восстановлению сна. Во время пробуждения клетки REM-на ингибируются подмножеством нейронов Aras Norepinephrin и серотонина, называемых rem-off-клетками.
- ^ Jump up to: а беременный в дюймовый и фон глин час я Брудзинский С.М. (июль 2014 г.). «Восходящая мезолимбическая холинергическая система - специфическое разделение ретикулярной активирующей системы, вовлеченного в начало негативных эмоциональных состояний» . Журнал молекулярной нейробиологии . 53 (3): 436–445. doi : 10.1007/s12031-013-0179-1 . PMID 24272957 . S2CID 14615039 .
Понимание возбуждения и поддержки бодрствования ARAS было еще более осложнялось нейрохимическими открытиями многочисленных групп нейронов с восходящими путями, происходящими в ретикулярном ядра ствола ствола, включая пантомэндэценфальные ядра, которые синтезируют различные передатчики и высвобождают их в общих областях мозг и во всем неокортекс (для обзора см. Jones 2003; Lin et al. 2011). Они включали глутаматергический, холинергический, норадренергический, дофаминергический, серотонинергический, гистаминергический и орексинергические системы (для обзора см. Lin et al. 2011). ... ARAS представлял собой диффузные, неспецифические пути, которые, работая через среднюю линию и внутриламинные ядра таламуса, могли бы изменить активность всего неокортекса, и, таким образом, эта система была первоначально предложена в качестве общей системы возбуждения для естественных стимулов и критической системы, лежащей в основе системы, лежащей в основе системы, лежащей в основе критической системы бодрствование (Moruzzi and Magoun 1949; Lindsley et al. 1949; Starzl et al. 1951, см. Область на рис. 1). ... в недавнем исследовании на крысе было обнаружено, что состояние бодрствования в основном поддерживается восходящей глутаматергической проекцией из паработ -ядра и областей Precoeruleus к базальному переднему мозгу, а затем передается в кору церебрального (Fuller et al. 2011. ) ... Анатомические исследования показали два основных пути, участвующих в возбуждении, и происходящие из областей с группами холинергических клеток, один через таламус, а другой - вентрально через гипоталамус и преоптическую область и взаимно связаны с лимбической системой (Наута и Kuypers 1958; ... как считается в холинергических связях с таламическим ретикулярным ядром ...
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Schwartz MD, Kilduff TS (декабрь 2015 г.). «Нейробиология сна и бодрствования» . Психиатрические клиники Северной Америки . 38 (4): 615–644. doi : 10.1016/j.psc.2015.07.002 . PMC 4660253 . PMID 26600100 .
Эта восходящая ретикулярная активирующая система (ARAS) состоит из холинергической латереодорсальной и педункулопонтинной сегмента (LDT/PPT), норадренергического локуса Coeruleus (LC), серотонергических (5-HT) ядер Raphe и допаминергическая вентральная область (VTA), субъект NIGRA (SN SN (SN SN (SN SN (SN SN (SN SN и допаминергическая вентральная область (VTA), субъекта (SN SN (SN SN (SN ) и периакедуктальные серого проекции, которые прямо и косвенно стимулируют кору через таламус, гипоталамус и BF. 6, 12-18 Эти аминергические и катехоламинергические популяции имеют многочисленные взаимосвязи и параллельные проекции, которые, вероятно, придают функциональную избыточность и устойчивость к системе. 6, 13, 19 ... Совсем недавно медуллярная парафициальная зона (PZ), прилегающая к лицевому нерву, была идентифицирована как центр, способствующий сон, на основе анатомических, электрофизиологических и химио- и оптогенетических исследований. 23, 24 ГАМКЕРГИЧЕСКИЕ ПЗ Нейроны ингибируют глутаматергические паракрахиальные (ПБ) нейроны, которые проецируют на BF, 25 тем самым способствуя сону NREM за счет бодрствования и быстрого сна. ... Нейроны HCRT широко проецируются на протяжении всего мозга и спинного мозга 92, 96, 99, 100 включая основные прогнозы к группам, способствующим пробуждению клеток, таких как клетки HA TM, 101 5-HT-клетки дорсальных ядер рафе (DRN), 101 норадренергические клетки LC, 102 и холинергические клетки в LDT, PPT и BF. 101, 103 ... HCRT непосредственно возбуждает клеточные системы, участвующие в бодрствовании и возбуждении, включая LC, 102, 106, 107 Drn, 108, 109 ТМ, 110-112 LDT, 113, 114 холинергический BF, 115 и как дофамин (DA), так и нейроны не-DA в VTA. 116, 117
- ^ Jump up to: а беременный в дюймовый Saper CB, Fuller PM (июнь 2017 г.). «Схема бодрствования: обзор» . Современное мнение о нейробиологии . 44 : 186–192. doi : 10.1016/j.conb.2017.03.021 . PMC 5531075 . PMID 28577468 .
Парабрахиальная и педункулопонтинная глутаматергическая система возбуждения
Ретроградные трассеры из BF постоянно идентифицировали один сайт ввода ствола мозга, который не является частью классической моноаминергической системы восходящего возбуждения: глутаматергические нейроны в парабрахиальном и педункулопонтинном ядре ... Соклеточные записи из педункновых нейронов обнаружили, что почти все холинергические нейроны в этом в этом. Регион, а также многие глутаматергические и ГАМКергические нейроны, наиболее активны во время бодрствования и восстановления [25], хотя некоторые из последних нейронов были максимально активны во время Wake или REM, но не оба. ... [Парабрахиальные и педункулопонтинные глутаматергические нейроны] обеспечивают тяжелую иннервацию к боковому гипоталамусу, центральному ядру миндалины и BF - ^ Jump up to: а беременный Pedersen NP, Ferrari L, Venner A, Wang JL, Abbott SG, Vujovic N, Arrigoni E, Saper CB, Fuller PM (ноябрь 2017). «Нейроны супрамаммиллярного глутамата являются ключевым узлом системы возбуждения» . Природная связь . 8 (1): 1405. Bibcode : 2017natco ... 8.1405p . doi : 10.1038/s41467-017-01004-6 . PMC 5680228 . PMID 29123082 .
Основные и клинические наблюдения предполагают, что каудальный гипоталамус содержит ключевой узел системы восходящего возбуждения, но типы клеток, лежащие в основе этого, не до конца понятны. Здесь мы сообщаем, что глютаматные, выделяющие нейроны супрамаммиллической области (SUMVGLUT2) продуцируют устойчивое поведенческое и возбуждение ЭЭГ при химиогенетически активировании.
- ^ Jump up to: а беременный в Burlet S, Tyler CJ, Leonard CS (апрель 2002 г.). «Прямое и косвенное возбуждение латеодорсальных сегментарных нейронов гипокретином/орексином пептидами: значение для бодрствования и нарколепсии» . J. Neurosci . 22 (7): 2862–2872. doi : 10.1523/jneurosci.22-07-02862.2002 . PMC 6758338 . PMID 11923451 .
- ^ Malenka RC, Nestler EJ, Hyman SE (2009). «Глава 12: Сон и возбуждение». В Сидоре А, Браун Р.Ю. (ред.). Молекулярная нейрофармакология: основание для клинической нейробиологии (2 -е изд.). Нью-Йорк: McGraw-Hill Medical. п. 295. ISBN 978-0071481274 Полем
Нейроны орексина расположены в боковом гипоталамусе. Они организованы широко выступающим образом, так же, как моноамины (глава 6), и иннервируют все компоненты ARA. Они возбуждают моноаминергические нейроны из-за удаления во время бодрствования и холинергические нейроны PT во время быстрого сна. Они ингибируются нейронами VLPO во время сна NREM.
- ^ Jump up to: а беременный Cherasse Y, урада Y (ноябрь 2017 г.). «Диетический цинк действует как модулятор сна» . Международный журнал молекулярных наук . 18 (11): 2334. doi : 10.3390/ijms18112334 . PMC 5713303 . PMID 29113075 .
Регуляция сна и бодрствования включает в себя множество областей и клеточных подтипов в мозге. Действительно, система восходящего возбуждения способствует бодрствованию через сеть, состоящую из монаминергических нейронов в локусе Coeruleus (LC), гистаминергических нейронов в туберомамилийском ядре (TMN), глутаматергические нейроны в парабара -ядрах (PB) ...
- ^ Fuller PM, Fuller P, Sherman D, Pedersen NP, Saper CB, Lu J (апрель 2011 г.). «Переоценка структурной основы восходящей системы возбуждения» . Журнал сравнительной неврологии . 519 (5): 933–956. doi : 10.1002/cne.225559 . PMC 3119596 . PMID 21280045 .
- ^ Jump up to: а беременный Kinomura S, Larsson J, Gulyás B, Roland PE (январь 1996 г.). «Активация при внимании человеческой ретикулярной образования и таламических внутриламиновных ядер». Наука . 271 (5248): 512–515. Bibcode : 1996sci ... 271..512K . doi : 10.1126/science.271.5248.512 . PMID 8560267 . S2CID 43015539 .
Это соответствует центро-медианы и ядрам централиса внутриламинарной группы
- ^ Сворад Д. (январь 1957 г.). «Ретикулярная активационная система ствола мозга и гипноза животных». Наука . 125 (3239): 156. Bibcode : 1957sci ... 125..156s . doi : 10.1126/science.125.3239.156 . PMID 13390978 .
- ^ Jump up to: а беременный Jang SH, Kwon HG (октябрь 2015 г.). «Прямой путь от ретикулярной образования ствола к стволу мозга к коре головного мозга в восходящей ретикулярной активационной системе: исследование диффузионной тензорной визуализации». Нейроски. Летал 606 : 200–203. doi : 10.1016/j.neulet.2015.09.004 . PMID 26363340 . S2CID 37083435 .
- ^ Purves et al (2018b) , Box 28a - Электроэнцефалография, с. 647–649
- ^ Jump up to: а беременный Steriade, M. (1996). «Пробуждение: пересмотр системы активации ретикуляции». Наука . 272 (5259): 225–226. Bibcode : 1996sci ... 272..225s . doi : 10.1126/science.272.5259.225 . PMID 8602506 . S2CID 39331177 .
- ^ Jump up to: а беременный Рейнер, П.Б. (1995). «Месопонтин -холинергические нейроны либо необходимыми, либо достаточными компонентами восходящей ретикулярной активационной системы?». Семинары в нейробиологии . 7 (5): 355–359. doi : 10.1006/smns.1995.0038 . S2CID 5575547 .
- ^ Jump up to: а беременный в Эванс, Б.М. (2003). «Сон, сознание и спонтанная и вызванная электрическая активность мозга. Существует ли корковый интегрирующий механизм?». Neurophysiologie Clinique . 33 (1): 1–10. doi : 10.1016/s0987-7053 (03) 00002-9 . PMID 12711127 . S2CID 26159370 .
- ^ Purves et al (2018b) , Нейронные схемы, регулирующие сон, с. 655–656
- ^ Мохан Кумар В., Маллик Б.Н., Чина Г.С., Сингх Б. (октябрь 1984 г.). «Влияние восходящей ретикулярной активационной системы на преоптическую активность нейронов». Эксплуат Неврол . 86 (1): 40–52. doi : 10.1016/0014-4886 (84) 90065-7 . PMID 6479280 . S2CID 28688574 .
- ^ Ван, Зан; Чжун, Ю-Хенг; Цзян, Шан; Qu, Wei-Min; Хуан, Чжи-Ли; Чен, Чанг-Руи (2022-03-14). «Случайный отчет: дисфункция площади паравентрикулярного ядра гипоталамуса вызывает гиперсомнию у пациентов» . Границы в нейробиологии . 16 doi : 10.3389/fnins.2022.830474 . ISSN 1662-453x . PMC 8964012 . PMID 35360167 .
- ^ Tindall SC (1990). «Глава 57: Уровень сознания» . В Walker HK, Hall WD, Hurst JW (ред.). Клинические методы: история, физические и лабораторные обследования . Butterworth Publishers. ISBN 978-0409900774 Полем Архивировано из оригинала на 2009-01-29 . Получено 2008-07-04 .
- ^ Nolte, J (ed.). "CHPT 11". Человеческий мозг: введение в его функциональную анатомию (5 -е изд.). С. 262–290.
- ^ Jump up to: а беременный Рут Р. Р., Розенфельд Дж.П. (октябрь 1977 г.). «Тоническая ретикулярная активационная система: связь с неприятными эффектами стимуляции мозга». Эксплуат Неврол . 57 (1): 41–56. doi : 10.1016/0014-4886 (77) 90043-7 . PMID 196879 . S2CID 45019057 .
- ^ Робинсон Д. (1999). «Техническая, неврологическая и психологическая значимость« альфа »,« Дельта »и« Тета », смешанных в ЭЭГ, вызванных потенциалами: исследование пиковых задержек». Клиническая нейрофизиология . 110 (8): 1427–1434. doi : 10.1016/s1388-2457 (99) 00078-4 . PMID 10454278 . S2CID 38882496 .
- ^ Лоуренс, Элеонора, изд. (2005). Электрическая связь (13 -е изд.). Pearson Education Limited. С. 195 . ISBN 978-0131273849 .
{{cite book}}:|work=игнорируется ( помощь ) - ^ Гарсия-Рил Э., Хейстер Д.С., Е. М., Чарльзворт А., Хаяр А. (2007). «Электрическая связь: новый механизм для бодрствования сна» . Спать . 30 (11): 1405–1414. doi : 10.1093/sleep/30.11.1405 . PMC 2082101 . PMID 18041475 .
- ^ Jump up to: а беременный Schwartz Jr, Roth T (декабрь 2008 г.). «Нейрофизиология сна и бодрствования: основные науки и клинические последствия» . Curr Neuropharmacol . 6 (4): 367–378. doi : 10.2174/157015908787386050 . PMC 2701283 . PMID 19587857 .
- ^ Винсент, SR (2000). «Восходящая ретикулярная активирующая система - от аминергических нейронов до оксида азота». Журнал химической нейроанатомии . 18 (1–2): 23–30. doi : 10.1016/s0891-0618 (99) 00048-4 . PMID 10708916 . S2CID 36236217 .
- ^ Hall RW, Huitt TW, Thapa R, Williams DK, Anand KJ, Garcia-Rill E (июнь 2008 г.). «Долгосрочный дефицит преждевременных родов: доказательства возбуждения и нарушений внимания» . Clin Neurophysiol . 119 (6): 1281–1291. doi : 10.1016/j.clinph.2007.12.021 . PMC 2670248 . PMID 18372212 .
- ^ Гарсия-Рилл Е., Бьюкенен Р., МакКеон К., Скиннер Р.Д., Уоллес Т (сентябрь 2007 г.). «Курение во время беременности: постнатальное воздействие на системы возбуждения и мозга» . Нейротоксикология . 28 (5): 915–923. Bibcode : 2007neutx..28..915g . doi : 10.1016/j.neuro.2007.01.007 . PMC 3320145 . PMID 17368773 .
- ^ Jump up to: а беременный в Squire L (2013). Фундаментальная нейробиология (4 -е изд.). Амстердам: Elsevier/Academic Press. С. 631–632. ISBN 978-0123858702 .
- ^ Fitzgerald MT, Gruener G, Mtui E (2012). Клиническая нейроанатомия и нейробиология . Филадельфия: Saunders Elsevier. п. 192. ISBN 978-0702037382 .
- ^ Браунстоун, Роберт М.; Чопек, Джереми В. (2018). «Ретикулоспинальные системы для настройки моторных команд» . Границы в нейронных цепях . 12:30 . DOI : 10.3389/fncir.2018.00030 . ISSN 1662-5110 . PMC 5915564 . PMID 29720934 .
- ^ Пирсон, Кейр Г; Гордон, Джеймс Э. (2013). «Глава 41 / осанка». В Канделе, Эрик Р; Шварц, Джеймс Х; Джесселл, Томас М; Зигельбаум, Стивен А; Hudspeth, AJ (Eds.). Принципы нейронной науки (5 -е изд.). Соединенные Штаты: МакГроу-Хилл. Ствол мозга и мозжечок интегрируют сенсорные сигналы для осанки, с. 954. ISBN 978-0071390118 .
- ^ Jump up to: а беременный в Майкл-Титус и др. (2010b) , вставка 9.5 Декортеката и обосторена, с. 172
- ^ Jump up to: а беременный Magoun HW (февраль 1952 г.). «Восходящая ретикулярная активационная система в стволе головного мозга». AMA Arch Neurol Psychiatry . 67 (2): 145–154, обсуждение 167–171. doi : 10.1001/archneurpsyc.1952.02320140013002 . PMID 14893989 .
Другие ссылки
[ редактировать ]- Системы тела (2010)
- Майкл-Титус, Адина Т; Revest, Патриция; Шортленд, Питер, ред. (2010a). «Глава 6 - черепные нервы и ствол мозга». Системы тела: нервная система - базовая наука и клинические состояния (2 -е изд.). Черчилль Ливингстон. ISBN 978-0702033735 .
- Майкл-Титус, Адина Т; Revest, Патриция; Шортленд, Питер, ред. (2010b). «Глава 9 - нисходящие пути и мозжечок». Системы тела: нервная система - базовая наука и клинические состояния (2 -е изд.). Черчилль Ливингстон. ISBN 978-0702033735 .
- Нейробиология (2018)
- Purves, Dale; Августин, Джордж Дж; Фицпатрик, Дэвид; Холл, Уильям С; Ламантия, Энтони Самуэль; Муни, Ричард Д; Платт, Майкл Л; Белый, Леонард Э, ред. (2018b). «Глава 28 - Государство коры». Нейробиология (6 -е изд.). Sinauer Associates. ISBN 978-1605353807 .
- Анатомия и физиология (2018)
- Saladin, KS (2018a). «Глава 13 - спинной мозг, спинномозговые нервы и соматические рефлексы». Анатомия и физиология: единство формы и функции (8 -е изд.). Нью-Йорк: МакГроу-Хилл. ISBN 978-1259277726 .
- Saladin, KS (2018b). «Глава 14 - мозг и черепные нервы». Анатомия и физиология: единство формы и функции (8 -е изд.). Нью-Йорк: МакГроу-Хилл. Ретикулярная формация, стр. 518–519. ISBN 978-1259277726 .
Внешние ссылки
[ редактировать ] Словажное определение ретикулярного образования при вики
Словажное определение ретикулярного образования при вики