Синапс
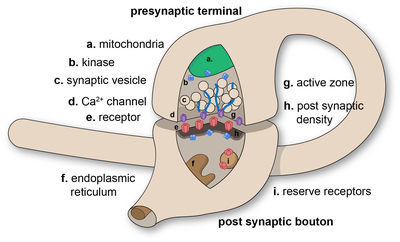
В системе синапс нервной [ 1 ] представляет собой структуру, которая позволяет нейрону (или нервной клетке) передавать электрический или химический сигнал другому нейрону или целевой эффекторной клетке.
Синапсы необходимы для передачи нервных импульсов от одного нейрона к другому. [ 2 ] играют ключевую роль в обеспечении быстрого и прямого общения путем создания цепей. Кроме того, синапс служит местом соединения, где происходит передача и обработка информации, что делает его жизненно важным средством связи между нейронами. [ 3 ] Нейроны специализируются на передаче сигналов отдельным клеткам-мишеням, а синапсы являются средством, с помощью которого они это делают.
В синапсе плазматическая мембрана передающего сигнал нейрона ( пресинаптического нейрона) приходит в тесное прилегание к мембране целевой ( постсинаптической ) клетки. Как пресинаптические, так и постсинаптические участки содержат обширный массив молекулярных механизмов , которые связывают две мембраны вместе и осуществляют процесс передачи сигналов. Во многих синапсах пресинаптическая часть расположена на аксоне , а постсинаптическая часть — на дендрите или соме . Астроциты также обмениваются информацией с синаптическими нейронами, реагируя на синаптическую активность и, в свою очередь, регулируя нейротрансмиссию . [ 2 ] Синапсы (по крайней мере, химические синапсы) стабилизируются в положении с помощью молекул синаптической адгезии (SAM), выступающих как из пре-, так и из постсинаптического нейрона и слипающихся вместе в местах перекрытия; SAM также могут способствовать образованию и функционированию синапсов. [ 4 ] Более того, SAM координируют формирование синапсов, при этом различные типы работают вместе для достижения замечательной специфичности синапсов. [ 3 ] [ 5 ] По сути, SAM функционируют как в возбуждающих, так и в тормозных синапсах, вероятно, служа устройствами для передачи сигналов. [ 3 ]
История
[ редактировать ]Сантьяго Рамон-и-Кахаль предположил, что нейроны не являются непрерывными по всему телу, но все же общаются друг с другом. Эта идея известна как доктрина нейронов . [ 6 ] Слово «синапс» было введено в 1897 году английским нейрофизиологом Чарльзом Шеррингтоном в Майкла Фостера «Учебнике физиологии» . [ 1 ] Шеррингтон изо всех сил пытался найти хороший термин, который подчеркивал бы союз между двумя отдельными элементами, а сам термин «синапс» был предложен английским ученым-классиком Артуром Вуллгаром Верраллом , другом Фостера. [ 7 ] [ 8 ] Слово произошло от греческого синапсиса ( σύναψις ), что означает «соединение», которое, в свою очередь, происходит от синаптеина ( συνάπτειν ), от син ( σύν ) «вместе» и гаптеина ( ἅπτειν ) «скреплять». [ 7 ] [ 9 ]
Однако, хотя синаптическая щель оставалась теоретической конструкцией и иногда о ней сообщалось как о разрыве между смежными окончаниями аксонов и дендритами или телами клеток, гистологические методы с использованием лучших световых микроскопов того времени не могли визуально разрешить их разделение, которое, как теперь известно, около 20 нм. В 1950-х годах понадобился электронный микроскоп, чтобы показать более тонкую структуру синапса с его отдельными параллельными пре- и постсинаптическим мембранами и отростками, а также щель между ними. [ 10 ] [ 11 ] [ 12 ]
Типы
[ редактировать ]
Химические и электрические синапсы представляют собой два способа синаптической передачи.
- В химическом синапсе электрическая активность пресинаптического нейрона преобразуется (посредством активации потенциалзависимых кальциевых каналов ) в высвобождение химического вещества, называемого нейротрансмиттером , которое связывается с рецепторами, расположенными на плазматической мембране постсинаптической клетки. Нейромедиатор может инициировать электрический ответ или вторичный мессенджерный путь, который может либо возбуждать, либо ингибировать постсинаптический нейрон. Химические синапсы можно классифицировать в зависимости от высвобождаемого нейромедиатора: глутаматергические (часто возбуждающие), ГАМКергические (часто тормозящие), холинергические (например, нервно-мышечные соединения позвоночных ) и адренергические (высвобождающие норадреналин ). Из-за сложности передачи сигнала рецептором химические синапсы могут оказывать сложное воздействие на постсинаптическую клетку.
- В электрическом синапсе пресинаптическая и постсинаптическая клеточные мембраны соединены специальными каналами, называемыми щелевыми контактами , которые способны пропускать электрический ток, вызывая изменения напряжения в пресинаптической клетке, вызывающие изменения напряжения в постсинаптической клетке. [ 13 ] [ 14 ] Фактически, щелевые контакты способствуют прямому прохождению электрического тока без необходимости использования нейротрансмиттеров, а также небольших молекул, таких как кальций. [ 15 ] Таким образом, главным преимуществом электрического синапса является быстрая передача сигналов от одной клетки к другой. [ 13 ]
- Смешанные химические электрические синапсы представляют собой синаптические участки, в которых имеется как щелевой контакт, так и высвобождение нейромедиатора. [ 16 ] [ 17 ] Эта комбинация позволяет сигналу иметь как быструю (электрическую), так и медленную (химическую) составляющую.
Формирование нейронных цепей в нервной системе, по-видимому, во многом зависит от важнейших взаимодействий между химическими и электрическими синапсами. Таким образом, эти взаимодействия управляют генерацией синаптической передачи. [ 14 ] Синаптическая коммуникация отличается от эфаптической связи , при которой связь между нейронами происходит посредством непрямых электрических полей. Аутапс — это химический или электрический синапс, который образуется, когда аксон одного нейрона синапсирует с дендритами того же нейрона.
Возбуждающее и тормозящее
[ редактировать ]- Возбуждающий синапс: повышает вероятность деполяризации постсинаптических нейронов и возникновения потенциала действия.
- Тормозной синапс: уменьшает вероятность деполяризации постсинаптических нейронов и возникновения потенциала действия.
Приток Na+, вызываемый возбуждающими нейротрансмиттерами, открывает катионные каналы, деполяризуя постсинаптическую мембрану в сторону порога потенциала действия. Напротив, тормозные нейротрансмиттеры заставляют постсинаптическую мембрану становиться менее деполяризованной, открывая либо Cl-, либо K+-каналы, уменьшая возбуждение. В зависимости от места высвобождения, рецепторов, с которыми они связываются, и ионных обстоятельств, с которыми они сталкиваются, различные передатчики могут быть либо возбуждающими, либо тормозящими. Например, ацетилхолин может либо возбуждать, либо ингибировать в зависимости от типа рецепторов, с которыми он связывается. [ 18 ] Например, глутамат служит возбуждающим нейромедиатором, в отличие от ГАМК, которая действует как тормозной нейромедиатор. Кроме того, дофамин является нейромедиатором, который оказывает двойное действие, оказывая как возбуждающее, так и тормозящее воздействие посредством связывания с отдельными рецепторами. [ 19 ]
Мембранный потенциал предотвращает попадание Cl- в клетку, даже если его концентрация снаружи значительно выше, чем внутри. Потенциал обращения Cl- во многих нейронах весьма отрицателен и почти равен потенциалу покоя . Открытие каналов Cl- имеет тенденцию буферизировать мембранный потенциал, но этому эффекту противодействуют, когда мембрана начинает деполяризоваться, позволяя большему количеству отрицательно заряженных ионов Cl- проникать в клетку. Следовательно, становится труднее деполяризовать мембрану и возбудить клетку, когда Cl-каналы открыты. Аналогичные эффекты возникают в результате открытия K+-каналов. Значение тормозных нейротрансмиттеров очевидно из воздействия токсинов, затрудняющих их активность. Например, стрихнин связывается с глициновыми рецепторами, блокируя действие глицина и приводя к мышечным спазмам, судорогам и смерти. [ 18 ]
Интерфейсы
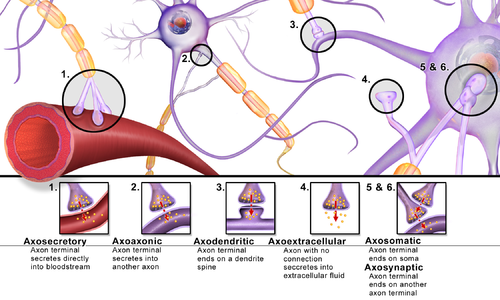
[ редактировать ]Синапсы можно классифицировать по типу клеточных структур, выступающих в качестве пре- и постсинаптических компонентов. Подавляющее большинство синапсов в нервной системе млекопитающих представляют собой классические аксо-дендритные синапсы (синапсы аксона на дендрите), однако существует множество других механизмов. К ним относятся, помимо прочего, [ нужны разъяснения ] аксо-аксонный , дендро-дендритный , аксо-секреторный, аксо-цилиарный, [ 20 ] сомато-дендритные, дендро-соматические и сомато-соматические синапсы. [ нужна ссылка ]
Фактически, аксон может образовывать синапсы с дендритом, с телом клетки или с другим аксоном или окончанием аксона, а также попадать в кровоток или диффузно в прилежащую нервную ткань.
Преобразование химических веществ в электрические сигналы
[ редактировать ]Нейротрансмиттеры — это крошечные сигнальные молекулы, хранящиеся в синаптических везикулах, закрытых мембраной, и высвобождаемые посредством экзоцитоза. Действительно, изменение электрического потенциала в пресинаптической клетке запускает высвобождение этих молекул. Прикрепляясь к ионным каналам, управляемым трансмиттером, нейромедиатор вызывает электрические изменения в постсинаптической клетке и быстро диффундирует через синаптическую щель. После высвобождения нейромедиатор быстро выводится, либо поглощаясь нервным окончанием, которое его продуцирует, либо поглощается близлежащими глиальными клетками, либо расщепляется специфическими ферментами в синаптической щели. Многочисленные Na+-зависимые белки-переносчики нейротрансмиттеров перерабатывают нейротрансмиттеры и позволяют клеткам поддерживать высокую скорость высвобождения.
В химических синапсах ионные каналы, управляемые трансмиттером, играют жизненно важную роль в быстром преобразовании внеклеточных химических импульсов в электрические сигналы. Эти каналы расположены в плазматической мембране постсинаптической клетки в области синапса и временно открываются в ответ на связывание молекулы нейромедиатора, вызывая мгновенное изменение проницаемости мембраны. Кроме того, трансмиттер-управляемые каналы сравнительно менее чувствительны к мембранному потенциалу, чем потенциал-управляемые каналы, поэтому они не способны самостоятельно генерировать самоусиливающееся возбуждение. Однако они приводят к ступенчатым изменениям мембранного потенциала из-за местной проницаемости, на которую влияет количество и продолжительность высвобождения нейромедиатора в синапсе. [ 18 ]
Недавно было обнаружено, что механическое напряжение, явление, которое никогда не считалось связанным с функцией синапсов, необходимо для возбуждения нейронов гиппокампа. [ 21 ]
Высвобождение нейромедиаторов
[ редактировать ]Нейротрансмиттеры связываются с ионотропными рецепторами постсинаптических нейронов, вызывая их открытие или закрытие. [ 19 ] Вариации количества нейромедиаторов, высвобождаемых пресинаптическим нейроном, могут играть роль в регуляции эффективности синаптической передачи. Фактически, концентрация цитоплазматического кальция участвует в регуляции высвобождения нейромедиаторов из пресинаптических нейронов. [ 22 ]
Химическая передача включает в себя несколько последовательных процессов:
- Синтез нейромедиаторов в пресинаптическом нейроне.
- Загрузка нейротрансмиттеров в секреторные пузырьки.
- Контроль высвобождения нейромедиаторов в синаптическую щель.
- Связывание нейромедиаторов с постсинаптическими рецепторами.
- Прекращение активности высвободившихся нейромедиаторов. [ 23 ]
Синаптическая поляризация
[ редактировать ]Функция нейронов зависит от полярности клеток . Отличительная структура нервных клеток позволяет потенциалам действия перемещаться направленно (от дендритов к телу клетки вниз по аксону), а затем эти сигналы принимаются и передаются постсинаптическим нейронам или эффекторным клеткам. Нервные клетки уже давно используются в качестве моделей клеточной поляризации, и особый интерес представляют механизмы, лежащие в основе поляризованной локализации синаптических молекул. Передача сигналов PIP2 , регулируемая IMPase, играет важную роль в синаптической полярности.
Фосфоинозитиды ( PIP , PIP2 и PIP3 ) — это молекулы, которые, как было показано, влияют на полярность нейронов. [ 24 ] ген ( ttx-7 был идентифицирован У Caenorhabditis elegans ) , который кодирует мио -инозитолмонофосфатазу (ИМФазу), фермент, который производит инозитол путем дефосфорилирования инозитолфосфата . Организмы с мутантными генами ttx-7 демонстрировали дефекты поведения и локализации, которые устранялись экспрессией IMPase. Это привело к выводу, что ИМФаза необходима для правильной локализации синаптических белковых компонентов. [ 25 ] [ 26 ] Ген egl-8 кодирует гомолог фосфолипазы Cβ (PLCβ), фермента, расщепляющего PIP2. Когда мутанты ttx-7 также имели мутантный ген egl-8 , дефекты, вызванные дефектным геном ttx-7, в значительной степени обращались вспять. Эти результаты позволяют предположить, что передача сигналов PIP2 устанавливает поляризованную локализацию синаптических компонентов в живых нейронах. [ 25 ]
Пресинаптическая модуляция
[ редактировать ]Модуляция нейромедиаторов высвобождения с помощью рецепторов, связанных с G-белком (GPCR), является важным пресинаптическим механизмом регуляции синаптической передачи . Активация GPCR, расположенных на пресинаптическом терминале, может снизить вероятность высвобождения нейромедиатора. Эта пресинаптическая депрессия включает активацию Gi/o -типа G-белков , которые опосредуют различные тормозные механизмы, включая ингибирование потенциалзависимых кальциевых каналов , активацию калиевых каналов и прямое ингибирование процесса слияния пузырьков .
Эндоканнабиноиды , синтезируемые и высвобождаемые из постсинаптических нейрональных элементов и родственных им рецепторов , включая рецептор (GPCR) CB1, расположенный на пресинаптическом конце, участвуют в этой модуляции посредством ретроградного сигнального процесса, при котором эти соединения синтезируются и высвобождаются из постсинаптических клеток. нейрональные элементы и возвращаются к пресинаптической терминали, чтобы воздействовать на рецептор CB1, вызывая кратковременную или долговременную синаптическую депрессию, что вызывает кратковременное или длительное снижение высвобождения нейромедиаторов. [ 27 ]
Влияние лекарств на лиганд-управляемые ионные каналы
[ редактировать ]Наркотики уже давно считаются важнейшими мишенями для ионных каналов, управляемых трансмиттером. Большинство лекарств, используемых для лечения шизофрении, тревоги, депрессии и бессонницы, воздействуют на химические синапсы, и многие из этих фармацевтических препаратов действуют путем связывания с трансмиттер-управляемыми каналами. Например, некоторые лекарства, такие как барбитураты и транквилизаторы, связываются с рецепторами ГАМК и усиливают ингибирующий эффект нейромедиатора ГАМК. Таким образом, пониженная концентрация ГАМК способствует открытию Cl-каналов.
Более того, психоактивные препараты потенциально могут воздействовать на многие другие компоненты синаптического сигнального механизма. Фактически, многочисленные нейротрансмиттеры высвобождаются Na+-переносчиками и впоследствии удаляются из синаптической щели. За счет ингибирования таких носителей синаптическая передача усиливается по мере продления действия передатчика. Например, Прозак — это антидепрессант, который предотвращает всасывание нейротрансмиттера серотонина. Кроме того, другие антидепрессанты действуют путем ингибирования реабсорбции серотонина и норадреналина. [ 18 ]
Биогенез
[ редактировать ]В нервных окончаниях быстро образуются синаптические везикулы, чтобы компенсировать их быстрое истощение во время высвобождения нейротрансмиттеров. Их биогенез включает отделение мембранных белков синаптических везикул от других клеточных белков и упаковку этих отдельных белков в везикулы соответствующего размера. Кроме того, это влечет за собой эндоцитоз мембранных белков синаптических везикул из плазматической мембраны. [ 28 ]
Синаптобластические и синаптокластические относятся к деятельности по производству и удалению синапсов в биохимической сигнальной цепи. Эта терминология связана с Протоколом Бредесена по лечению болезни Альцгеймера , который концептуализирует болезнь Альцгеймера как дисбаланс между этими процессами. По состоянию на октябрь 2023 года исследования, касающиеся этого протокола, остаются небольшими, и в рамках стандартизированной системы контроля было получено мало результатов.
Роль в памяти
[ редактировать ]Потенцирование и депрессия
[ редактировать ]Широко признано, что синапс играет ключевую роль в формировании памяти . [ 29 ] Стабильность долговременной памяти может сохраняться в течение многих лет; тем не менее, синапсы, неврологическая основа памяти, очень динамичны. [ 30 ] Формирование синаптических связей существенно зависит от зависящей от активности синаптической пластичности, наблюдаемой в различных синаптических путях. Действительно, связь между формированием памяти и изменениями синаптической эффективности позволяет усиливать нейронные взаимодействия между нейронами. Поскольку нейротрансмиттеры активируют рецепторы через синаптическую щель, связь между двумя нейронами усиливается, когда оба нейрона активны одновременно, в результате сигнальных механизмов рецептора. Считается, что сила двух связанных нервных путей приводит к хранению информации, что приводит к памяти. Этот процесс усиления синапсов известен как долговременная потенциация (ДП) . [ 29 ]
Изменяя высвобождение нейротрансмиттеров, можно контролировать пластичность синапсов в пресинаптической клетке. Постсинаптическую клетку можно регулировать путем изменения функции и количества ее рецепторов. Изменения в постсинаптической передаче сигналов чаще всего связаны с рецептором N-метил-d-аспарагиновой кислоты (NMDAR)-зависимой ДП и долговременной депрессией (ДЛТ) из-за притока кальция в постсинаптическую клетку, которые являются наиболее частыми. проанализировали формы пластичности возбуждающих синапсов. [ 31 ]
Механизм протеинкиназы
[ редактировать ]Более того, Ca2+/кальмодулин (CaM)-зависимая протеинкиназа II (CaMKII) лучше всего известна благодаря своей роли в головном мозге, особенно в областях неокортекса и гиппокампа, поскольку она служит универсальным медиатором клеточных сигналов Ca2+. CaMKII широко распространен в нервной системе, в основном сконцентрирован в синапсах нервных клеток. Действительно, CaMKII был окончательно идентифицирован как ключевой регулятор когнитивных процессов, таких как обучение и нейронная пластичность. Были продемонстрированы первые конкретные экспериментальные доказательства давно предполагаемой функции CaMKII в хранении памяти.
В то время как связывание Ca2+/CaM стимулирует активность CaMKII, Ca2+-независимая автономная активность CaMKII также может быть вызвана рядом других процессов. CaMKII становится активным путем аутофосфорилирования при связывании Ca2+/кальмодулина. CaMKII все еще активен и фосфорилирует себя даже после отщепления Ca2+; в результате мозг сохраняет долговременные воспоминания, используя этот механизм. Тем не менее, когда фермент CaMKII дефосфорилируется ферментом фосфатазой, он становится неактивным, и воспоминания теряются. Следовательно, CaMKII играет жизненно важную роль как в индукции, так и в поддержании LTP. [ 32 ]
Экспериментальные модели
[ редактировать ]По техническим причинам синаптическая структура и функции исторически изучались на необычно больших синаптических моделях , например:
- Гигантский синапс кальмара
- Нервно-мышечное соединение (НМС), холинергический синапс у позвоночных, глутаматергический у насекомых.
- Ресничная чашечка цилиарного узла цыплят. [ 33 ]
- Чашечка Хелда в стволе мозга
- Ленточный синапс в сетчатке
- Коллатеральные синапсы Шаффера в гиппокампе . Эти синапсы небольшие, но их пре- и постсинаптические нейроны хорошо разделены (СА3 и СА1 соответственно).
Синапсы и болезни
[ редактировать ]Синапсы функционируют как ансамбли внутри определенных сетей мозга, контролируя уровень активности нейронов, который необходим для памяти, обучения и поведения. Следовательно, синаптические нарушения могут иметь негативные последствия. Фактически, изменения во внутренних молекулярных системах клетки или модификации биохимических процессов окружающей среды могут привести к синаптической дисфункции. Синапс является основной единицей передачи информации в нервной системе, и правильное создание синаптических контактов во время развития имеет важное значение для нормальной функции мозга. Кроме того, несколько мутаций связаны с нарушениями развития нервной системы, и нарушение функции различных мест синапсов является отличительной чертой нейродегенеративных заболеваний.
Синаптические дефекты причинно связаны с ранними неврологическими заболеваниями, включая расстройства аутистического спектра (РАС), шизофрению (СКЗ) и биполярное расстройство (БП). С другой стороны, при дегенеративных патологиях с поздним началом, таких как болезни Альцгеймера (БА), Паркинсона (БП) и Гентингтона (БГ), синаптопатия считается неизбежным конечным результатом продолжающегося патофизиологического каскада. Эти заболевания характеризуются постепенной потерей когнитивных и поведенческих функций и устойчивой потерей мозговой ткани. Более того, эти ухудшения в основном связаны с постепенным накоплением белковых агрегатов в нейронах, состав которых может варьироваться в зависимости от патологии; все они оказывают одинаковое вредное воздействие на целостность нейронов. Более того, большое количество мутаций, связанных со структурой и функцией синапсов, а также изменениями дендритных отростков в посмертных тканях, привело к связи между синаптическими дефектами и расстройствами нервного развития, такими как РАС и SCZ, характеризующимися аномальными поведенческими или когнитивными нарушениями. фенотипы.
Тем не менее, из-за ограниченного доступа к тканям человека на поздних стадиях и отсутствия тщательной оценки основных компонентов заболеваний человека на доступных экспериментальных моделях животных было трудно полностью понять происхождение и роль синаптической дисфункции в неврологических расстройствах. [ 34 ]
Дополнительные изображения
[ редактировать ]-
 Схема синапса. Посетите сайт Learnbio.org для интерактивной версии.
Схема синапса. Посетите сайт Learnbio.org для интерактивной версии. -
 Типичный синапс центральной нервной системы
Типичный синапс центральной нервной системы -
 Синапс и цикл синаптических пузырьков
Синапс и цикл синаптических пузырьков -
 Основные элементы химической синаптической передачи
Основные элементы химической синаптической передачи




См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Фостер М., Шеррингтон CS (1897). Учебник физиологии . Том. 3 (7-е изд.). Лондон: Макмиллан. п. 929. ИСБН 978-1-4325-1085-5 .
- ^ Перейти обратно: а б Переа Г., Наваррете М., Араке А. (август 2009 г.). «Трехсторонние синапсы: астроциты обрабатывают и контролируют синаптическую информацию». Тенденции в нейронауках . 32 (8). Cell Press : 421–431. doi : 10.1016/j.tins.2009.05.001 . ПМИД 19615761 . S2CID 16355401 .
- ^ Перейти обратно: а б с Зюдхоф ТЦ (июль 2021 г.). «Клеточная биология образования синапсов» . Журнал клеточной биологии . 220 (7): e202103052. дои : 10.1083/jcb.202103052 . ПМК 8186004 . ПМИД 34086051 .
- ^ Мисслер М, Зюдхоф ТЦ, Бидерер Т (апрель 2012 г.). «Синаптическая клеточная адгезия» . Перспективы Колд-Спринг-Харбор в биологии . 4 (4): а005694. doi : 10.1101/cshperspect.a005694 . ПМК 3312681 . ПМИД 22278667 .
- ^ Хейл В.Д., Зюдхоф ТК, Хуганир Р.Л. (январь 2023 г.). «Сконструированные молекулы адгезии управляют организацией синапсов» . Труды Национальной академии наук Соединенных Штатов Америки . 120 (3): e2215905120. Бибкод : 2023PNAS..12015905H . дои : 10.1073/pnas.2215905120 . ПМЦ 9934208 . ПМИД 36638214 .
- ^ Элиас Л.Дж., Сосье Д.М. (2006). Нейропсихология: клинические и экспериментальные основы . Бостон: Пирсон/Аллин и Бэкон . ISBN 978-0-20534361-4 . LCCN 2005051341 . OCLC 61131869 .
- ^ Перейти обратно: а б Харпер Д. «Синапс» . Интернет-словарь этимологии .
- ^ Тэнси Э.М. (1997). «Не совершать варварства: Шеррингтон и синапс, 1897». Бюллетень исследований мозга . 44 (3). Эльзевир : 211–212. дои : 10.1016/S0361-9230(97)00312-2 . ПМИД 9323432 . S2CID 40333336 .
Слово «синапс» впервые появилось в 1897 году в седьмом издании « Учебника физиологии» Майкла Фостера .
- ^ синапсис , синаптеин , син , гаптеин . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей» .
- ^ Де Робертис ЭД, Беннетт Х.С. (январь 1955 г.). «Некоторые особенности субмикроскопической морфологии синапсов лягушки и дождевого червя» . Журнал биофизической и биохимической цитологии . 1 (1): 47–58. дои : 10.1083/jcb.1.1.47 . ПМЦ 2223594 . ПМИД 14381427 .
- ^ Палай С.Л., Паладе Г.Е. (январь 1955 г.). «Тонкая структура нейронов» . Журнал биофизической и биохимической цитологии . 1 (1): 69–88. дои : 10.1083/jcb.1.1.69 . ПМЦ 2223597 . ПМИД 14381429 .
- ^ Палай С.Л. (июль 1956 г.). «Синапсы в центральной нервной системе» . Журнал биофизической и биохимической цитологии . 2 (4 приложения): 193–202. дои : 10.1083/jcb.2.4.193 . ПМК 2229686 . ПМИД 13357542 .
- ^ Перейти обратно: а б Сильверторн ДУ (2007). Физиология человека: комплексный подход (4-е изд.). Сан-Франциско: Пирсон/Бенджамин Каммингс . п. 271. ИСБН 978-0-8053-6851-2 . LCCN 2005056517 . OCLC 62742632 .
- ^ Перейти обратно: а б Переда А.Е. (апрель 2014 г.). «Электрические синапсы и их функциональные взаимодействия с химическими синапсами» . Обзоры природы. Нейронаука . 15 (4): 250–263. дои : 10.1038/nrn3708 . ПМК 4091911 . ПМИД 24619342 .
- ^ Кэр М.Дж., Редди В., Варакалло М. (2023). «Физиология, Синапс» . СтатПерлс . Остров сокровищ (Флорида): StatPearls Publishing. ПМИД 30252303 . Проверено 1 января 2024 г.
- ^ Сотело С., Палай С.Л. (февраль 1970 г.). «Тонкая структура позднего вестибулярного ядра у крысы. II. Синаптическая организация». Исследования мозга . 18 (1): 93–115. дои : 10.1016/0006-8993(70)90459-2 . ПМИД 4313893 .
- ^ Штраусфельд, штат Нью-Джерси, Бассемир, Великобритания (декабрь 1983 г.). «Кобальт-связанные нейроны гигантской волоконной системы двукрылых». Журнал нейроцитологии . 12 (6): 971–91. дои : 10.1007/BF01153345 . ПМИД 6420522 . S2CID 19764983 .
- ^ Перейти обратно: а б с д Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «Ионные каналы и электрические свойства мембран» . Молекулярная биология клетки (4-е изд.). Гирляндная наука . Проверено 20 января 2024 г.
- ^ Перейти обратно: а б Ласица А., Брюэр С. (2023). «Возбуждающая и тормозная синаптическая передача сигналов» . Научите меня физиологии .
- ^ Шеу С.Х., Упадьяюла С., Дюпуи В., Пан С., Дэн Ф., Ван Дж. и др. (сентябрь 2022 г.). «Серотонинергический аксон-ресничный синапс управляет ядерной передачей сигналов, изменяя доступность хроматина» . Клетка . 185 (18): 3390–3407.e18. дои : 10.1016/j.cell.2022.07.026 . ПМЦ 9789380 . ПМИД 36055200 . S2CID 251958800 .
- Пресс-релиз университета: «Ученые открыли новый вид синапсов в крошечных волосках нейронов» . Медицинский институт Говарда Хьюза через phys.org . Проверено 19 октября 2022 г.
- ^ Джой М.С., Налл Д.Л., Эмон Б., Ли К.Ю., Баришман А., Ахмед М., Рахман С., Селвин П.Р., Саиф М.Т. (26 декабря 2023 г.). «Синапсы без напряжения не срабатывают в сети нейронов гиппокампа in vitro» . Труды Национальной академии наук . 120 (52). дои : 10.1073/pnas.2311995120 . ISSN 0027-8424 . ПМЦ 10756289 . ПМИД 38113266 .
- ^ Питман Р.М. (сентябрь 1984 г.). «Универсальный синапс». Журнал экспериментальной биологии . 112 (1): 199–224. дои : 10.1242/jeb.112.1.199 . ПМИД 6150966 .
- ^ Хольц Р.В., Фишер С.К. (1999). «Синапсическая передача» . В: Сигел Г.Дж., Агранов Б.В., Альберс Р.В., Фишер С.К., Улер, доктор медицинских наук (ред.). Основная нейрохимия: молекулярные, клеточные и медицинские аспекты (6-е изд.). Липпинкотт-Рейвен . Проверено 4 января 2024 г.
- ^ Аримура Н., Кайбучи К. (декабрь 2005 г.). «Ключевые регуляторы нейрональной полярности» . Нейрон . 48 (6). Cell Press: 881–884. дои : 10.1016/j.neuron.2005.11.007 . ПМИД 16364893 .
- ^ Перейти обратно: а б Кимата Т., Танидзава Ю., Джан Ю., Икеда С., Кухара А., Мори И. (июнь 2012 г.). «Синапсическая полярность зависит от передачи сигналов фосфатидилинозитола, регулируемой мио-инозитолмонофосфатазой у Caenorhabditis elegans» . Генетика . 191 (2). Американское генетическое общество : 509–521. дои : 10.1534/genetics.111.137844 . ПМЦ 3374314 . ПМИД 22446320 .
- ^ Танидзава Ю, Кухара А, Инада Х, Кодама Э, Мизуно Т, Мори И (декабрь 2006 г.). «Инозитолмонофосфатаза регулирует локализацию синаптических компонентов и поведение в зрелой нервной системе C. elegans» . Гены и развитие . 20 (23). Лабораторное издательство Колд-Спринг-Харбор : 3296–3310. дои : 10.1101/gad.1497806 . ПМК 1686606 . ПМИД 17158747 .
- ^ Любовник ДМ (2008). «Пресинаптическая модуляция эндоканнабиноидами». В Südhof TC, Старке К. (ред.). Фармакология высвобождения нейромедиаторов . Справочник по экспериментальной фармакологии. Том. 184. Шпрингер Берлин Гейдельберг. стр. 435–477. дои : 10.1007/978-3-540-74805-2_14 . ISBN 9783540748052 . ПМИД 18064422 .
- ^ Деснос С., Клифт-о'Грейди Л., Келли Р.Б. (1 сентября 1995 г.). «Биогенез синаптических везикул in vitro» . Журнал клеточной биологии . 130 (5): 1041–1049. дои : 10.1083/jcb.130.5.1041 . ISSN 0021-9525 . ПМК 2120557 . ПМИД 7544795 .
- ^ Перейти обратно: а б Линч М.А. (январь 2004 г.). «Долговременная потенциация и память» . Физиологические обзоры . 84 (1): 87–136. doi : 10.1152/physrev.00014.2003 . ПМИД 14715912 .
- ^ Ян Ю, Лу Дж, Цзо Ю (декабрь 2018 г.). «Изменения синаптических структур, связанные с обучением, памятью и болезнями» . Достижения науки о мозге . 4 (2): 99–117. дои : 10.26599/BSA.2018.2018.9050012 .
- ^ Крюгерс Х.Дж., Чжоу М., Джоэлс М., Киндт М. (11 октября 2011 г.). «Регуляция возбуждающих синапсов и страшных воспоминаний гормонами стресса» . Границы поведенческой нейронауки . 5 . Frontiers Media SA : 62. doi : 10.3389/fnbeh.2011.00062 . ПМК 3190121 . ПМИД 22013419 .
- ^ Байер КУ, Шульман Х (август 2019 г.). «CaM-киназы: все еще вдохновляет в 40 лет» . Нейрон . 103 (3): 380–394. дои : 10.1016/j.neuron.2019.05.033 . ПМЦ 6688632 . ПМИД 31394063 .
- ^ Стэнли Э.Ф. (1992). «Синапс типа чашечки цилиарного ганглия цыпленка как модель быстрой холинергической передачи». Канадский журнал физиологии и фармакологии . 70 (Дополнение): S73–S77. дои : 10.1139/y92-246 . ПМИД 1338300 .
- ^ Тауфик Э., Курупи Г., Зигоянни О., Матсас Р. (05.09.2018). «Синапсическая дисфункция при нейродегенеративных заболеваниях и заболеваниях, связанных с развитием нервной системы: обзор моделей индуцированных плюрипотентных заболеваний на основе стволовых клеток» . Открытая биология . 8 :180138.doi : (9 ) 10.1098/rsob.180138 . ISSN 2046-2441 . ПМК 6170506 . ПМИД 30185603 .