Плакозоа
| Плакозоя Временной диапазон: [1] | |
|---|---|
 | |
| Прикрепленный трихоплакс | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | Чоанозоа |
| Королевство: | животное |
| Тип: | Плакозоа Грелль, 1971 год. |
| Типовой вид | |
| Прикрепленный трихоплакс | |
| Классы [2] | |
| |
Placozoa ( / p l æ k ə ˈ z oʊ ə / PLAK -ə- ZOH -ə , «плоские животные») [3] — тип морских и свободноживущих (непаразитических) животных. [4] [5] Это похожие на капли животные, состоящие из скоплений клеток. Передвигаясь в воде посредством ресничек , поедая пищу путем поглощения , размножаясь делением или почкованием , плакозоа описываются как «простейшие животные на Земле». [6] Структурный и молекулярный анализ подтвердил, что они относятся к числу наиболее примитивных животных. [7] [8] таким образом, составляя примитивный тип многоклеточных животных . [9]
Первый известный плакозой, Trichoplax adhaerens , был открыт в 1883 году немецким зоологом Францем Эйльхардом Шульце (1840–1921). [10] [11] Описывая уникальность, другой немец, Карл Готлиб Грелль (1912–1994), в 1971 году выделил для него новый тип Placozoa. Оставаясь монотипическим типом более века, [12] [13] новые виды начали добавляться с 2018 года. На данный момент описаны еще три вида в двух отдельных классах: Uniplacotomia ( Hoilungia hongkongensis в 2018 году и Cladtertia collaboinventa в 2022 году). [14] ) и Полиплакотомия ( Polyplacotoma mediterranea , самая базальная, в 2019 г. [15] ).
История
[ редактировать ]Трихоплакс был открыт в 1883 году немецким зоологом Францем Эйльхардом Шульце в аквариуме с морской водой Зоологического института в Граце, Австрия . [10] [16] Родовое название происходит от классического греческого θρίξ ( thrix ), что означает «волосы», и πλάξ ( plax ), «пластина». Специфический эпитет adhaerens на латыни означает «прикрепленный», что отражает его склонность прилипать к предметным стеклам и пипеткам, используемым при его исследовании. [17] Шульце понял, что это животное не может быть членом какого-либо существующего типа, и, основываясь на простом строении и поведении, в 1891 году пришел к выводу, что это должно быть раннее многоклеточное животное. Он также наблюдал размножение путем деления клеток, слоев клеток и передвижения. [18]
В 1893 году итальянский зоолог Франческо Саверио Монтичелли описал другое животное, названное им Трептоплакс , экземпляры которого он собрал в Неаполе. В 1896 году он дал видовое название T. Reptans . [19] Монтичелли их не сохранил, и больше никаких экземпляров обнаружено не было, в результате чего идентификация была признана сомнительной, а вид отвергнут. [20] [21]
против описания Шульце выступили другие зоологи. Например, в 1890 году Ф. К. Нолл утверждал, что это животное было плоским червем (Turbellaria). [22] В 1907 году Тило Крумбах опубликовал гипотезу о том, что не является отдельным животным, а представляет собой форму личинки планулы анемоноподобного гидрозойного трихоплакс растения Eleutheria krohni . Хотя это было опровергнуто в печати Шульце и другими, анализ Крумбаха стал стандартным объяснением в учебниках, и в зоологических журналах ничего не печаталось о трихоплаксе . до 1960-х годов [17]
Развитие электронной микроскопии в середине 20 века позволило провести углубленное наблюдение клеточных компонентов организма, после чего с 1966 года возобновился интерес к трихоплаксу . [23] Наиболее важные описания были сделаны Карлом Готлибом Греллем в Тюбингенском университете с 1971 года. [24] [25] В том же году Грелль возродил интерпретацию Шульца об уникальности животных и создал новый тип Placozoa. [26] [17] Грелль получил это название от гипотезы плакулы, Отто Бючли идеи о происхождении многоклеточных животных . [27]
Биология
[ редактировать ]
1 — липидная капля, 2 — ресничка, 3 — дорсальный слой клеток, 4 — вакуоль,
5 — фиброзный синцитий, 6 — железистая клетка, 7 — вакуоль,
8 — вентральный слой клеток, 9 — зоны межклеточных контактов
Плакозои не имеют четко определенного строения тела, как и амебы , одноклеточные эукариоты. Как сообщил Эндрю Мастерсон: «Они настолько близки, насколько это возможно, к тому, чтобы быть просто маленькой живой каплей». [28] Отдельное тело имеет диаметр около 0,55 мм. [29] Частей тела нет; как описал один из исследователей Майкл Эйтель: «Нет ни рта, ни спины, ни нервных клеток, ничего». [30] Животные, изучаемые в лабораториях, имеют тела, состоящие из сотен и миллионов клеток. [31]
Плакозойные имеют внутри тела всего три анатомические части в виде тканевых слоев: верхний, промежуточный (средний) и нижний эпителий . Существует как минимум шесть различных типов клеток. [32] Верхний эпителий представляет собой самую тонкую часть и по существу состоит из плоских клеток, тела которых свисают под поверхностью, и каждая клетка имеет ресничку . [33] Кристаллические клетки редко распределены вблизи краевого края. Лишь немногие клетки имеют необычно большое количество митохондрий . [32] Средний слой является самым толстым и состоит из многочисленных волокнистых клеток, которые содержат митохондриальные комплексы, вакуоли и эндосимбиотические бактерии в эндоплазматической сети . Нижний эпителий состоит из многочисленных монореснитчатых цилиндрических клеток, а также нескольких эндокриноподобных железистых клеток и липофильных клеток. Каждая липофильная клетка содержит множество гранул среднего размера, одна из которых является секреторной . [34] [33]
Оси тела Hoilungia и Trichoplax явно похожи на орально-аборальную ось книдарий . [35] животные из другого типа, с которым они наиболее тесно связаны. [36] Структурно их нельзя отличить от других плакозой, поэтому идентификация проводится исключительно на основе генетических (митохондриальной ДНК) различий. [37] Секвенирование генома показало, что каждый вид имеет набор уникальных генов и несколько уникально недостающих генов. [14]
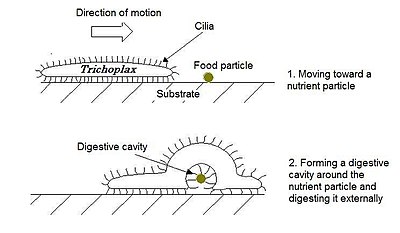
Трихоплакс — небольшое уплощенное животное диаметром около 1 мм (0,039 дюйма). Аморфное многоклеточное тело, аналогичное одноклеточной амебе , не имеет правильных очертаний, хотя нижняя поверхность несколько вогнута, а верхняя всегда уплощена. Тело состоит из внешнего слоя простого эпителия, окружающего рыхлый лист звездчатых клеток, напоминающий мезенхиму некоторых более сложных животных. Эпителиальные клетки несут реснички , которые животное использует, чтобы ползать по морскому дну. [11]
Нижняя поверхность поглощает мелкие частицы органического детрита, которыми животное питается . Все плакозоа могут размножаться бесполым путем, отпочковываясь от более мелких особей, а нижняя поверхность может также откладывать яйца в мезенхиму . [11] половое размножение Сообщается, что происходит в одной кладе плакозой. [38] [39] чей штамм H8, как позже выяснилось, принадлежал к роду Cladtertia , [2] где межгенная рекомбинация наблюдалась , а также другие признаки полового размножения.
Некоторые Trichoplax виды Rickettsiales содержат бактерии в качестве эндосимбионтов . [40] У одного из как минимум 20 описанных видов оказалось два бактериальных эндосимбионта; Грелия , которая живет в эндоплазматическом ретикулуме животных и, как предполагается, играет роль в производстве белка и мембран. Другой эндосимбионт – это первая описанная бактерия Маргулис , живущая внутри клеток, используемых для пищеварения водорослей . Похоже, что он поедает жиры и другие липиды водорослей и взамен снабжает своего хозяина витаминами и аминокислотами. [41] [42]
Исследования показывают, что кристаллы арагонита в кристаллических ячейках выполняют ту же функцию, что и статолиты, что позволяет использовать гравитацию для пространственной ориентации . [43]
В дорсальном эпителии расположены липидные гранулы, называемые блестящими сферами, которые выделяют смесь ядов и токсинов в качестве защиты от хищников и могут вызвать паралич или смерть у некоторых хищников. У трихоплакса были обнаружены гены, очень похожие на гены яда некоторых ядовитых змей, таких как американская медноголовая и западноафриканская ковровая гадюка. [44] [45]
Placozoa демонстрируют значительную эволюционную радиацию в отношении натриевых каналов , которых у них 5–7 различных типов, что больше, чем у любого другого вида беспозвоночных, изученных на сегодняшний день. [47]
Три режима динамики популяции зависели от источников питания, включая индукцию социального поведения, морфогенез и репродуктивные стратегии. [48]
Помимо деления, у представителей всех видов образовались «роевики» (отдельная стадия вегетативного размножения), которые также могли формироваться из нижнего эпителия с большим клеточным разнообразием. [49]
Эволюционные отношения
[ редактировать ]Убедительных окаменелостей плакозоа нет, хотя эдиакарский (докембрий, 550 миллионов лет назад ) организм дикинсонии кажется несколько похожим на плакозоа. [50] Кнауст (2021) сообщил о сохранении окаменелостей плакозоев в пласте микробиалита из среднего триаса Muschelkalk ( Германия ). [1]
Традиционно классификация основывалась на уровне их организации, т. е. у них нет тканей и органов. Однако это может быть результатом вторичной потери и, следовательно, недостаточно для исключения их из отношений с более сложными животными. В более поздних работах была предпринята попытка классифицировать их на основе последовательностей ДНК в их геноме; это поместило тип между губками и эуметазоа . [51] Считается, что в таком бедном типе молекулярные данные обеспечивают наиболее надежное приближение к филогении плакозой.
Их точное положение на филогенетическом дереве дало бы важную информацию о происхождении нейронов и мышц. Если отсутствие этих особенностей является исходной чертой Placozoa, это означало бы, что нервная система и мышцы развивались трижды, если placozoa и cnidarians были сестринской группой ; один раз в Ctenophora , один раз в Cnidaria и один раз в Bilateria . Если бы они разветвились до того, как разделились Cnidaria и Bilateria, нейроны и мышцы в двух последних группах имели бы одно и то же происхождение.
Функционально-морфологическая гипотеза
[ редактировать ]

Из-за своей простой структуры Placozoa часто рассматривались как модельный организм для перехода от одноклеточных организмов к многоклеточным животным ( Metazoa ) и, таким образом, считались сестринским таксоном для всех других многоклеточных животных:
| многоклеточные животные |
| ||||||||||||
Согласно функционально-морфологической модели, все или большинство животных произошли от галлертоида — свободноживущей ( пелагической ) сферы в морской воде, состоящей из одного ресничного слоя клеток, поддерживаемого тонким неклеточным разделительным слоем — базальной пластинкой . Внутренняя часть сферы заполнена сократительными волокнистыми клетками и студенистым внеклеточным матриксом . И современные Placozoa, и все другие животные затем произошли от этой начальной многоклеточной стадии посредством двух разных процессов: [53]
- Складывание эпителия привело к образованию внутренней системы протоков и, таким образом, к развитию видоизмененного галлертоида, из которого впоследствии развились губки ( Porifera ), Cnidaria и Ctenophora .
- Другие галлертоиды, согласно этой модели, со временем перешли к донному образу жизни; то есть их среда обитания переместилась из открытого океана на дно (бентическая зона). Это естественным образом приводит к избирательному преимуществу в уплощении тела, что, конечно, можно наблюдать у многих донных видов.
Хотя вероятность встречи с пищей, потенциальными половыми партнерами или хищниками одинакова во всех направлениях для животных, свободно плавающих в воде, на морском дне существует явная разница между функциями, полезными на сторонах тела, обращенных к субстрату и от него . заставляя свои сенсорные, защитные и собирающие пищу клетки дифференцироваться и ориентироваться по вертикали – направлению, перпендикулярному субстрату. В предложенной функционально-морфологической модели Placozoa и, возможно, несколько подобных организмов, известных только по окаменелостям, произошли от такой формы жизни, которую сейчас называют плакулоидной .
Три разные жизненные стратегии соответственно привели к трем различным возможным линиям развития:
- Животные, живущие в песке на дне океана, были ответственны за следы ползания окаменелостей, которые считаются самыми ранними свидетельствами существования животных; и их можно было обнаружить еще до начала эдиакарского периода в геологии . Их обычно приписывают двусторонне-симметричным червям, но представленная здесь гипотеза предполагает, что животные, полученные из плакулоидов и, следовательно, близкие родственники Trichoplax adhaerens . продуцентами следов являются
- Животные, включившие водоросли в качестве фотосинтетически активных эндосимбионтов , т.е. получавшие питательные вещества в первую очередь от своих партнеров по симбиозу , соответственно, были ответственны за загадочных существ эдиакарской фауны, не отнесенных ни к одному современному таксону животных и живших в эдиакарский период, до начала палеозоя . Однако недавняя работа показала, что некоторые эдиакарские сообщества (например, Мистейкен-Пойнт ) находились в глубокой воде, ниже фотической зоны , и, следовательно, эти особи не могли зависеть от эндосимбиотических фотосинтезаторов .
- Животные, которые паслись на водорослевых матах , в конечном итоге стали прямыми предками Placozoa. Таким образом, преимущества амебоидного разнообразия форм позволили имевшейся ранее базальной пластинке и студенистому внеклеточному матриксу потеряться вторично . Выраженная дифференциация между поверхностью, обращенной к субстрату ( вентральной ), и поверхностью, обращенной от него ( дорсальной ), соответственно привела к появлению физиологически различных клеточных слоев Trichoplax adhaerens , которые можно увидеть и сегодня. Следовательно, они аналогичны , но не гомологичны эктодерме – «внешнему» и «внутреннему» слоям клеток у эуметазоев – т.е. структуры , и энтодерме функционально соответствующие друг другу, не имеют, согласно предложенной гипотезе, общего эволюционного происхождения.
Если какой-либо из представленных выше анализов окажется верным, Trichoplax adhaerens станет древнейшей ветвью многоклеточных животных и реликтом эдиакарской фауны или даже доэдиакарской фауны. Несмотря на то, что эти животные очень преуспели в своей экологической нише , из-за отсутствия внеклеточного матрикса и базальной пластинки потенциал развития этих животных, конечно, был ограничен, что могло бы объяснить низкую скорость эволюции их фенотипа (их внешней формы во взрослом состоянии). чтобы так же бредительно . [ нужна ссылка ]
Эта гипотеза была подтверждена недавним анализом Trichoplax adhaerens митохондриального генома по сравнению с геномом других животных. [54] Однако эта гипотеза была отвергнута при статистическом анализе полногеномной последовательности Trichoplax adhaerens по сравнению с полногеномными последовательностями шести других животных и двух родственных видов, не являющихся животными, но только на уровне p = 0,07, что указывает на маргинальный уровень. уровень статистической значимости. [51]
Гипотеза эпителиозоев
[ редактировать ]Концепция, основанная на чисто морфологических характеристиках, рассматривает Placozoa как ближайшего родственника животных с настоящими тканями ( Eumetazoa ). Их общий таксон, называемый Epitheliozoa , сам по себе считается сестринской группой губок ( Porifera ):
Вышеупомянутая точка зрения может быть верной, хотя есть некоторые свидетельства того, что гребневики , традиционно рассматриваемые как Eumetazoa , могут быть сестрами всех других животных. [55] Сейчас это спорная классификация. [56] По оценкам, плакозойные появились 750–800 миллионов лет назад, а первый современный нейрон возник у общего предка книдарий и билатерий около 650 миллионов лет назад (многие гены, экспрессируемые в современных нейронах, отсутствуют в гребневиках, хотя некоторые из этих недостающих генов присутствуют у плакозойных). [57] [58]
Основным подтверждением таких взаимоотношений являются особые межклеточные соединения – ременные десмосомы – которые встречаются не только у Placozoa, но и у всех животных, за исключением губок: они позволяют клеткам соединяться в единый слой, подобный эпителиоиду Placozoa. . Trichoplax adhaerens также разделяет клетки вентральной железы с большинством эуметазоев. Обе характеристики можно считать эволюционно полученными признаками ( апоморфиями ) и, таким образом, составляют основу общего таксона для всех животных, обладающих ими. [ нужна ссылка ]
Один из возможных сценариев, вдохновленный предложенной гипотезой, начинается с идеи о том, что монореснитчатые клетки эпителиоида Trichoplax adhaerens эволюционировали в результате редукции воротников в воротниковых клетках ( хоаноцитах ) губок, когда предполагаемые предки Placozoa отказались от фильтрующего образа жизни. . Эпителиоид тогда послужил бы предшественником истинной эпителиальной ткани эуметазоев. [ нужна ссылка ]
В отличие от модели, основанной на функциональной морфологии, описанной ранее, в гипотезе Epitheliozoa вентральный и дорсальный клеточные слои Placozoa являются гомологами энтодермы и эктодермы — двух основных слоев эмбриональных клеток эуметазоа. Пищеварительный гастродермис у Cnidaria или эпителий кишечника у билатерально-симметричных животных ( Bilateria ), возможно, развился из энтодермы, тогда как эктодерма является предшественником внешнего слоя кожи ( эпидермиса ), среди прочего. Внутреннее пространство, заполненное синцитием волокон у Placozoa, тогда соответствовало бы соединительной ткани у других животных. Неясно, связаны ли ионы кальция, хранящиеся в синцитии, с известковыми скелетами многих книдарий. [ нужна ссылка ]
Как отмечалось выше, эта гипотеза была подтверждена статистическим анализом полногеномной последовательности Trichoplax adhaerens по сравнению с полногеномными последовательностями шести других животных и двух родственных видов, не являющихся животными. [51]
Гипотеза эуметазоа
[ редактировать ]Третья гипотеза, основанная в первую очередь на молекулярной генетике, рассматривает Placozoa как сильно упрощенных эуметазоа . Согласно этому, Trichoplax adhaerens произошел от значительно более сложных животных, у которых уже были мышцы и нервные ткани. Оба типа тканей, а также базальная пластинка эпителия были утрачены совсем недавно в результате радикального вторичного упрощения. [59]
Различные исследования в этом отношении до сих пор дают разные результаты для определения точной сестринской группы: в одном случае Placozoa можно было бы квалифицировать как ближайших родственников Cnidaria , тогда как в другом они были бы сестринской группой Ctenophora , и иногда их относят к прямо рядом с Bilateria . В настоящее время их обычно размещают в соответствии с кладограммой ниже: [60]
В этой кладограмме Epitheliozoa и Eumetazoa являются синонимами друг друга и диплобластов , а Ctenophora являются их базальными.
Аргументом против предложенного сценария является то, что он совершенно не учитывает морфологические особенности животных. Более того, крайняя степень упрощения, которую пришлось бы постулировать для Placozoa в этой модели, известна только для паразитических организмов, но ее было бы трудно функционально объяснить у свободноживущих видов, таких как Trichoplax adhaerens . [ нужна ссылка ]
Эта версия подтверждается статистическим анализом полногеномной последовательности Trichoplax adhaerens в сравнении с полногеномными последовательностями шести других животных и двух родственных видов, не являющихся животными. Однако гребневики не были включены в анализ, в результате чего плакозоа оказались за пределами отобранных Eumetazoas. [51] [61]
Гипотеза книдарии-сестры
[ редактировать ]Сравнение ДНК позволяет предположить, что плакозоа связаны с Cnidaria , происходящими от планулы (как это видно у некоторых Cnidaria). личинки [62] произошли от планулоидов. Bilateria Считается, что [63] [64] [65] [66] [67] [68] [69] [70] Ось тела Cnidaria и Placozoa явно схожа, а клетки Placozoa и Cnidarian реагируют на одни и те же нейропептидные антитела, несмотря на то, что у существующих Placozoa не развиваются нейроны. [71] [72]
| Чоанозоа |
| ||||||||||||||||||
| 950 млн лет назад |



Ссылки
[ редактировать ]- ^ Перейти обратно: а б Кнауст, Дирк. «Микробиалит с погребенным бентосным сообществом из группы Мюшелкальк среднего триаса (анизий-ладин) Германии» .
- ^ Перейти обратно: а б Тесслер, Майкл; Нойманн, Йоханнес С.; Камм, Кай; Осигус, Ханс-Юрген; Эшель, Гил; Наречанья, Апурва; Бернс, Джон А.; ДеСалле, Роб; Шируотер, Бернд (08 декабря 2022 г.). «Филогеномика и первая высшая таксономия Placozoa, древнего и загадочного типа животных» . Границы экологии и эволюции . 10 . дои : 10.3389/fevo.2022.1016357 .
- ^ Рюдигер Венер и Вальтер Геринг (июнь 2007 г.). Зоология (на немецком языке) (24-е изд.). Штутгарт: Тиме. п. 696.
- ^ Камм, Кай; Шируотер, Бернд; ДеСалле, Роб (2019). «Врожденный иммунитет у простейших животных — плакозой» . БМК Геномика . 20 (1): 5. дои : 10.1186/s12864-018-5377-3 . ПМК 6321704 . ПМИД 30611207 .
- ^ Placozoa в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- ^ Пенниси, Элизабет (2021). «Простейший из снов» . Наука . 374 (6567): 526–529. Бибкод : 2021Sci...374..526P . дои : 10.1126/science.acx9444 . ISSN 1095-9203 . ПМИД 34709907 . S2CID 240154099 .
- ^ Нойманн, Йоханнес С.; ДеСалле, Тесслер; ДеСалле, Роб; Майкл, Бернд (2021). «Современная систематика беспозвоночных» . В Шируотере, Бернд; ДеСалле, Роб (ред.). Зоология беспозвоночных: подход «Древо жизни» . ЦРК Пресс. п. 71. дои : 10.1201/9780429159053 . ISBN 978-1-4822-3582-1 . S2CID 260730525 .
- ^ Кавасима, Такеши; Ёсида, Маса-Аки; Миядзава, Хидеюки; Накано, Хироаки; Накано, Натуми; Сакамото, Тацуя; Хамада, Маюко (2022). «Наблюдение за разнообразием многоклеточных животных на уровне типов с помощью анализа ДНК окружающей среды в районе Ушимадо во Внутреннем море Сето» . Зоологическая наука . 39 (1): 157–165. дои : 10.2108/zs210073 . ISSN 0289-0003 . ПМИД 35107003 . S2CID 246297787 .
- ^ Шируотер, Бернд; Эйтель, Майкл (2015), Ваннингер, Андреас (редактор), «Placozoa» , Эволюционная биология развития беспозвоночных 1 , Вена: Springer Vienna, стр. 107–114, doi : 10.1007/978-3-7091-1862-7_5 , ISBN 978-3-7091-1861-0 , получено 2 июня 2023 г.
- ^ Перейти обратно: а б Ф. Е. Шульце « Trichoplax adhaerens ng, ns», Зоологический вестник (Эльзевир, Амстердам и Йена) 6 (1883), с. 92.
- ^ Перейти обратно: а б с Барнс, Роберт Д. (1982). Зоология беспозвоночных . Филадельфия: Холт-Сондерс Интернэшнл. стр. 84–85. ISBN 978-0-03-056747-6 .
- ^ Фойгт, О; Коллинз АГ; Пирс В.Б.; Пирс Дж.С.; Хадрис Х; Эндер А (23 ноября 2004 г.). «Placozoa — больше не отдельный тип» . Современная биология . 14 (22): R944–5. Бибкод : 2004CBio...14.R944V . дои : 10.1016/j.cub.2004.10.036 . PMID 15556848 . S2CID 11539852 .
- ^ Эйтель, Майкл; Осигус, Ханс-Юрген; ДеСалле, Роб; Шируотер, Бернд (2 апреля 2013 г.). «Глобальное разнообразие плакозоа» . ПЛОС ОДИН . 8 (4): e57131. Бибкод : 2013PLoSO...857131E . дои : 10.1371/journal.pone.0057131 . ПМЦ 3614897 . ПМИД 23565136 .
- ^ Перейти обратно: а б Тесслер, Майкл; Нойманн, Йоханнес С.; Камм, Кай; Осигус, Ханс-Юрген; Эшель, Гил; Наречанья, Апурва; Бернс, Джон А.; ДеСалле, Роб; Шируотер, Бернд (08 декабря 2022 г.). «Филогеномика и первая высшая таксономия Placozoa, древнего и загадочного типа животных» . Границы экологии и эволюции . 10 . дои : 10.3389/fevo.2022.1016357 . ISSN 2296-701X .
- ^ Шируотер, Бернд; Камм, Кай; Херцог, Ребекка; Рольфес, Сара; Осигус, Ханс-Юрген (04 марта 2019 г.). «Polyplacotoma mediterranea — новый разветвленный вид плакозойных» . Современная биология . 29 (5): Р148–Р149. Бибкод : 2019CBio...29.R148O . дои : 10.1016/j.cub.2019.01.068 . ISSN 0960-9822 . ПМИД 30836080 .
- ^ Шируотер, Бернд; Осигус, Ханс-Юрген; Бергманн, Тьярд; Блэкстоун, Нил В.; Хадрис, Хайке; Хауслаге, Йенс; Умберт, Патрик О.; Камм, Кай; и др. (2021). «Загадочные Placozoa, часть 1: Исследование эволюционных противоречий и плохих экологических знаний» . Биоэссе . 43 (10): e2100080. doi : 10.1002/bies.202100080 . ПМИД 34472126 . S2CID 237387715 .
- ^ Перейти обратно: а б с Сайед, Т.; Шируотер, Б. (2002). « Trichoplax adhaerens : обнаружен как недостающее звено, забыт как гидрозойное животное, вновь открыт как ключ к эволюции многоклеточных животных» . Vie Milieu . 52 (4): 177–187. Архивировано из оригинала 2 июня 2023 г. Проверено 2 июня 2023 г. - через HAL .
- ^ Романова Дарья Ю.; Варокко, Фредерик; Дарасп, Жан; Никитин Михаил А.; Эйтель, Майкл; Фассауэр, Дирк; Мороз, Леонид Л. (2021). «Скрытое клеточное разнообразие Placozoa: ультраструктурные данные Hoilungia hongkongensis» . Исследования клеток и тканей . 385 (3): 623–637. дои : 10.1007/s00441-021-03459-y . ПМЦ 8523601 . ПМИД 33876313 .
- ^ Каттанео-Вьетти, Р.; Руссо, GF (01 января 2019 г.). «Краткая история итальянской морской биологии» . Европейский зоологический журнал . 86 (1): 294–315. дои : 10.1080/24750263.2019.1651911 . ISSN 2475-0263 . S2CID 202372627 .
- ^ Тесслер, Майкл; Нойманн, Йоханнес С.; Камм, Кай; Осигус, Ханс-Юрген; Эшель, Гил; Наречанья, Апурва; Бернс, Джон А.; ДеСалле, Роб; Шируотер, Бернд (08 декабря 2022 г.). «Филогеномика и первая высшая таксономия Placozoa, древнего и загадочного типа животных» . Границы экологии и эволюции . 10 . дои : 10.3389/fevo.2022.1016357 . ISSN 2296-701X .
- ^ «WoRMS - Всемирный реестр морских видов - Treptoplax Reptans Monticelli, 1896» . www.marinespecies.org . Проверено 2 июня 2023 г.
- ^ Тесслер, Майкл; Нойманн, Йоханнес С.; Камм, Кай; Осигус, Ханс-Юрген; Эшель, Гил; Наречанья, Апурва; Бернс, Джон А.; ДеСалле, Роб; Шируотер, Бернд (08 декабря 2022 г.). «Филогеномика и первая высшая таксономия Placozoa, древнего и загадочного типа животных» . Границы экологии и эволюции . 10 . дои : 10.3389/fevo.2022.1016357 . ISSN 2296-701X .
- ^ Куль, Вилли; Куль, Гертруда (1966). «Исследования двигательного поведения Trichoplax adhaerens FE Schulze (временная трансформация: промежуток времени)» . Журнал морфологии и экологии животных (на немецком языке). 56 (4): 417–435. дои : 10.1007/BF00442291 . ISSN 0720-213X . S2CID 20206608 .
- ^ Грелль, К.Г. (1971). «Эмбриональное развитие Trichoplax adhaerens FE Schulze» . Естественные науки (на немецком языке). 58 (11): 570. Бибкод : 1971NW.....58..570G . дои : 10.1007/BF00598728 . ISSN 0028-1042 . S2CID 40022799 .
- ^ Грелль, Карл Г. (1972). «Формирование яиц и бороздки Trichoplax adhaerens FE Schulze (Placozoa)» . Журнал морфологии животных (на немецком языке). 73 (4): 297–314. дои : 10.1007/BF00391925 . ISSN 0720-213X . S2CID 22931046 .
- ^ Г, Грелль К. (1971). «Trichoplax adhaerens FE Schulze и происхождение многоклеточных животных» . Естественнонаучный обзор . 24 : 160-161.
- ^ Шируотер, Бернд; ДеСалле, Роб (2018). «Плакозоа» . Современная биология . 28 (3): Р97–Р98. Бибкод : 2018CBio...28..R97S . дои : 10.1016/j.cub.2017.11.042 . ПМИД 29408263 . S2CID 235331464 .
- ^ Мастерсон, Эндрю (01 августа 2018 г.). «В конце концов, простые организмы не так уж и просты» . Журнал «Космос» . Проверено 2 июня 2023 г.
- ^ Эйтель, Майкл; Фрэнсис, Уоррен Р.; Варокко, Фредерик; Дарасп, Жан; Осигус, Ханс-Юрген; Кребс, Стефан; Варгас, Серхио; Блюм, Гельмут; и др. (2018). «Сравнительная геномика и природа видов плакозойных» . ПЛОС Биология . 16 (7): e2005359. дои : 10.1371/journal.pbio.2005359 . ПМК 6067683 . ПМИД 30063702 .
- ^ Вуд, Чарли (06 октября 2018 г.). «Простейшее животное раскрывает скрытое разнообразие» . Научный американец . Проверено 2 июня 2023 г. - через журнал Quanta.
- ^ Исследователи из Стэнфорда раскрывают новый механизм сохранения нетронутых клеток животных.
- ^ Перейти обратно: а б Романова Дарья Ю.; Варокко, Фредерик; Дарасп, Жан; Никитин Михаил А.; Эйтель, Майкл; Фассауэр, Дирк; Мороз, Леонид Л. (2021). «Скрытое клеточное разнообразие Placozoa: ультраструктурные данные Hoilungia hongkongensis» . Исследования клеток и тканей . 385 (3): 623–637. дои : 10.1007/s00441-021-03459-y . ПМЦ 8523601 . ПМИД 33876313 .
- ^ Перейти обратно: а б Эйтель, Майкл; Фрэнсис, Уоррен Р.; Варокко, Фредерик; Дарасп, Жан; Осигус, Ханс-Юрген; Кребс, Стефан; Варгас, Серхио; Блюм, Гельмут; и др. (2018). «Сравнительная геномика и природа видов плакозойных» . ПЛОС Биология . 16 (7): e2005359. дои : 10.1371/journal.pbio.2005359 . ПМК 6067683 . ПМИД 30063702 .
- ^ Тесслер, Майкл; Нойманн, Йоханнес С.; Камм, Кай; Осигус, Ханс-Юрген; Эшель, Гил; Наречанья, Апурва; Бернс, Джон А.; ДеСалле, Роб; Шируотер, Бернд (08 декабря 2022 г.). «Филогеномика и первая высшая таксономия Placozoa, древнего и загадочного типа животных» . Границы экологии и эволюции . 10 . дои : 10.3389/fevo.2022.1016357 . ISSN 2296-701X .
- ^ ДюБук TQ, Райан Дж. Ф., Мартиндейл MQ (май 2019 г.). « Дорсально-вентральные» гены являются частью древней системы осевого паттерна: данные Trichoplax adhaerens (Placozoa)» . Молекулярная биология и эволюция . 36 (5): 966–973. дои : 10.1093/molbev/msz025 . ПМК 6501881 . ПМИД 30726986 .
- ^ Лаумер, Кристофер Э.; Фернандес, Роза; Лемер, Сара; Комбош, Дэвид; Кокот, Кевин М.; Риск, Анна; Андраде, Соня CS; Штеррер, Вольфганг; Соренсен, Мартин В.; Гирибет, Гонсало (10 июля 2019 г.). «Возврат к филогении многоклеточных животных с выборкой геномов всех типов» . Слушания. Биологические науки . 286 (1906): 20190831. doi : 10.1098/rspb.2019.0831 . ISSN 1471-2954 . ПМК 6650721 . ПМИД 31288696 .
- ^ Эйтель, Майкл; Фрэнсис, Уоррен Р.; Варокко, Фредерик; Дарасп, Жан; Осигус, Ханс-Юрген; Кребс, Стефан; Варгас, Серхио; Блюм, Гельмут; и др. (2018). «Сравнительная геномика и природа видов плакозойных» . ПЛОС Биология . 16 (7): e2005359. дои : 10.1371/journal.pbio.2005359 . ПМК 6067683 . ПМИД 30063702 .
- ^ Синьорович, А.Ю.; Деллапорта, СЛ; Басс, Л.В. (2005). «Молекулярные признаки пола у Placozoa» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (43): 15518–22. Бибкод : 2005PNAS..10215518S . дои : 10.1073/pnas.0504031102 . ПМК 1266089 . ПМИД 16230622 .
- ^ Чарльзворт, Д. (2006). «Популяционная генетика: использование рекомбинации для обнаружения полового размножения: контрастирующие случаи Placozoa и C. elegans ». Наследственность (Edinb.) . 96 (5): 341–342. дои : 10.1038/sj.hdy.6800809 . ПМИД 16552431 . S2CID 44333533 .
- ^ Камм, Кай; Шируотер, Бернд; ДеСалле, Роб (05 января 2019 г.). «Врожденный иммунитет у простейших животных – плакозой» . БМК Геномика . 20 (1): 5. дои : 10.1186/s12864-018-5377-3 . ISSN 1471-2164 . ПМК 6321704 . ПМИД 30611207 .
- ^ «Обманчиво просто: мелкие морские животные живут в сложном симбиозе с бактериями» . Phys.org (пресс-релиз). Общество Макса Планка. 10 июня 2019 г. Проверено 23 июня 2021 г.
- ^ Грубер-Водица, Харальд; Лейш, Нико; Кляйнер, Мануэль; Хинцке, Тьервен; Либеке, Мануэль; Макфолл-Нгай, Маргарет; и др. (2019). «Два внутриклеточных и клеточно-специфичных бактериальных симбионта у плакозойных Trichoplax H2» . Природная микробиология . 4 (9): 1465–1474. дои : 10.1038/s41564-019-0475-9 . ПМЦ 6784892 . ПМИД 31182796 .
- ^ Шируотер, Бернд; Осигус, Ханс-Юрген; Бергманн, Тьярд; Блэкстоун, Нил В.; Хадрис, Хайке; Хауслаге, Йенс; Умберт, Патрик О.; Камм, Кай; Квансакул, Марк; Высоцкий, Кэтрин; ДеСалле, Роб (2021). «Загадочные Placozoa, часть 2: Исследование эволюционных противоречий и многообещающих вопросов на Земле и в космосе» . Биоэссе . 43 (10): e2100083. doi : 10.1002/bies.202100083 . ISSN 0265-9247 . ПМИД 34490659 .
- ^ Живые тайны: познакомьтесь с самым простым животным Земли.
- ^ Куэрво-Гонсалес, Родриго (сентябрь 2017 г.). «Rhodope placozophagus (Heterobranchia) — новый вид турбелляриеподобных брюхоногих моллюсков, питающихся плакозойями» . Зоологический обзор . 270 : 43–48. Бибкод : 2017ЗооАн.270...43С . дои : 10.1016/j.jcz.2017.09.005 .
- ^ Эйтель, Майкл; Осигус, Ханс-Юрген; Десаль, Роб; Шируотер, Бернд (2013). «Глобальное разнообразие Placozoa» . ПЛОС ОДИН . 8 (4): e57131. Бибкод : 2013PLoSO...857131E . дои : 10.1371/journal.pone.0057131 . ПМЦ 3614897 . ПМИД 23565136 .
- ^ Романова Дарья Ю.; Смирнов Иван Васильевич; Никитин Михаил А.; Кон, Андреа Б.; Борман, Алиса И.; Малышев Алексей Юрьевич; и др. (29 октября 2020 г.). «Потенциалы действия натрия у плакозоа: понимание поведенческой интеграции и эволюции беснервных животных» . Связь с биохимическими и биофизическими исследованиями . 532 (1): 120–126. дои : 10.1016/j.bbrc.2020.08.020 . ПМЦ 8214824 . ПМИД 32828537 .
- ^ Романова Дарья; Никитин Михаил; Щенков Сергей; Мороз, Леонид (2022). «Расширение жизненных стратегий плакозоа: результаты долгосрочного культивирования трихоплакса и ойлунгии » . Границы клеточной биологии и биологии развития . 10 : 823283. doi : 10.3389/fcell.2022.823283 . ПМЦ 8864292 . ПМИД 35223848 .
- ^ Романова Дарья; Варокко, Фредерик; Дарасп, Жан; Никитин Михаил; Эйтель, Майкл; Фассауэр, Дирк; Мороз, Леонид (2021). «Расширение стратегии жизни Placozoa: результаты долгосрочного культивирования трихоплакса и хойлунгии » . Границы клеточной биологии и биологии развития . 10 : 623–637. дои : 10.3389/fcell.2022.823283 . ПМЦ 8864292 . ПМИД 35223848 .
- ^ Сперлинг, Эрик; Винтер, Якоб; Пизани, Давиде; Петерсон, Кевин (2008). «Сродство плакозой к дикинсонии и эволюция способов питания многоклеточных животных позднего докембрия» (PDF) . В Кьюсаке, М.; Оуэн, А.; Кларк, Н. (ред.). Программа с тезисами . Ежегодное собрание Палеонтологической ассоциации . Том. 52. Глазго, Великобритания. п. 81.
- ^ Перейти обратно: а б с д Шривастава, М.; Бегович, Эмина; Чепмен, Джаррод; Патнэм, Николас Х.; Хеллстен, Уффе; Кавасима, Такеши; и др. (21 августа 2008 г.). « Геном трихоплакса и природа плакозойных» . Природа . 454 (7207): 955–960. Бибкод : 2008Natur.454..955S . дои : 10.1038/nature07191 . ПМИД 18719581 . S2CID 4415492 .
- ^ Смит К.Л., Варокко Ф., Киттельманн М., Аззам Р.Н., Купер Б., Уинтерс К.А. и др. (июль 2014 г.). «Новые типы клеток, нейросекреторные клетки и строение тела ранних дивергентных многоклеточных животных Trichoplax adhaerens» . Современная биология . 24 (14): 1565–1572. Бибкод : 2014CBio...24.1565S . дои : 10.1016/j.cub.2014.05.046 . ПМЦ 4128346 . ПМИД 24954051 .
- ^ Грассхофф, Манфред; Гудо, Майкл (2002). «Происхождение многоклеточных животных и основные эволюционные линии животного царства: галлертоидная гипотеза в свете современных исследований». Сенкенбергиана Летея . 82 (1). ООО «Спрингер Сайенс энд Бизнес Медиа»: 295–314. дои : 10.1007/bf03043790 . ISSN 0037-2110 . S2CID 84989130 .
- ^ Деллапорта, СЛ; Сюй, А.; Сагассер, С.; Якоб, В.; Морено, Массачусетс; Басс, ЛВ; Шируотер, Б.; и др. (6 июня 2006 г.). «Митохондриальный геном Trichoplax adhaerens поддерживает Placozoa как базальный тип низших многоклеточных животных» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (23): 8751–8756. Бибкод : 2006PNAS..103.8751D . дои : 10.1073/pnas.0602076103 . ПМК 1470968 . ПМИД 16731622 .
- ^ Уилан, Натан В.; Кокот, Кевин М.; Мороз Татьяна П.; Мукерджи, Кришану; Уильямс, Питер; Паулай, Густав; и др. (09.10.2017). «Отношения гребневиков и их место в качестве сестринской группы для всех других животных» . Экология и эволюция природы . 1 (11): 1737–1746. Бибкод : 2017NatEE...1.1737W . дои : 10.1038/s41559-017-0331-3 . ISSN 2397-334X . ПМЦ 5664179 . ПМИД 28993654 .
- ^ «Губки и гребешки» . Новости и особенности. www.bristol.ac.uk (пресс-релиз). Бристольский университет. Ноябрь 2015 года . Проверено 11 марта 2023 г.
- ^ Крошечные морские существа раскрывают древнее происхождение нейронов.
- ^ Найл, Себастьян Р.; Грау-Бове, Ксавье; Элек, Анамария; Наваррете, Кристина; Чианферони, Дамиано; Чива, Кристина; Каньяс-Арментерос, Дидак; Маллабиабаррена, Аррате; Камм, Кай; Сабидо, Эдвард; Грубер-Водика, Харальд; Шируотер, Бернд; Серрано, Луис; Себе-Педрос, Арнау (2023 г.). «Поэтапное возникновение программы экспрессии нейрональных генов на ранней стадии эволюции животных» . Клетка . 186 (21): 4676–4693.e29. дои : 10.1016/j.cell.2023.08.027 . hdl : 10230/58738 . ПМИД 37729907 .
- ^ Печеник, Ян (2015). «Пориферы и плакозойцы». Биология беспозвоночных (7-е изд.). Макгроу-Хилл Образование. п. 90. ИСБН 978-0073524184 .
- ^ Лейден, Майкл Дж. (2018), «Гены Cnidarian Zic», семья Зик , Достижения в экспериментальной медицине и биологии, том. 1046, Спрингер, Сингапур, стр. 27–39, номер doi : 10.1007/978-981-10-7311-3_2 , ISBN. 9789811073106 , PMID 29442315
- ^ Вальберг, Андреас; Толлессон, Микаэль; Фаррис, Джеймс С.; Йонделиус, Ульф (1 декабря 2004 г.). «Филогенетическое положение гребневиков (Ctenophora) и значение таксономической выборки» . Кладистика . 20 (6): 558–578. дои : 10.1111/j.1096-0031.2004.00041.x . ISSN 1096-0031 . ПМИД 34892961 . S2CID 86185156 .
- ^ Лаумер, CE; Фернандес Р.; Лемер, С.; Комбош, Д.; Кокот, КМ; Рисго, А.; Андраде, SCS; Стеррер, В.; Соренсен, М.В.; Гирибет, Г. (2019). «Возврат к филогении многоклеточных животных с выборкой геномов всех типов» . Учеб. Биол. Наука . 286 (1906): 20190831. doi : 10.1098/rspb.2019.0831 . ПМК 6650721 . ПМИД 31288696 .
- ^ Алешин В.В.; Петров, Н.Б. (2002). «Молекулярные доказательства регресса в эволюции многоклеточных животных». Ж. Обще. Биол . 63 (3): 195–208. ПМИД 12070939 .
- ^ Лаумер, Кристофер Э.; Грубер-Водица, Харальд; Хэдфилд, Майкл Г.; Пирс, Вики Б.; Рисго, Ана; Мариони, Джон К.; Гирибет, Гонсало (30 октября 2018 г.). «Поддержка клады Placozoa и Cnidaria в генах с минимальной композиционной предвзятостью» . электронная жизнь . 7 . дои : 10.7554/elife.36278 . ISSN 2050-084X . ПМК 6277202 . ПМИД 30373720 .
- ^ Сайед, Тарек; Шируотер, Бернд (1 июня 2002 г.). «Эволюция плакозоа: новая морфологическая модель». Сенкенбергиана Летея . 82 (1): 315–324. дои : 10.1007/bf03043791 . ISSN 0037-2110 . S2CID 16870420 .
- ^ Хейнол, Андреас; Мартиндейл, Марк К. (27 апреля 2008 г.). «Разработка Acoel поддерживает простой урбилатерий, похожий на планулу» . Философские труды Королевского общества B: Биологические науки . 363 (1496): 1493–1501. дои : 10.1098/rstb.2007.2239 . ISSN 0962-8436 . ПМК 2614228 . ПМИД 18192185 .
- ^ Альзугарай, Мэри Евгения; Бруно, Мэри Сесилия; Вильялобос Самбукаро, Мария Хосе; Рондерос, Джордж Рафаэль (2019). «Эволюционная история семейства орексина/аллатотропина GPCR: от Placozoa и Cnidaria до позвоночных» . Научные отчеты 9 (1): 10217. Бибкод : 2019НатСР...910217А . bioRxiv 10.1101/403709 . дои : 10.1038/ s41598-019-46712-9 ПМК 6629687 . ПМИД 31308431 . S2CID 256990037 .
- ^ Сильва, Фернанда Бритто да; Мушнер, Валерия К.; Бонатто, Сандро Л. (2007). «Филогенетическое положение Placozoa на основе генов рРНК большой субъединицы (LSU) и малой субъединицы (SSU)» . Генетика и молекулярная биология . 30 (1): 127–132. дои : 10.1590/S1415-47572007000100022 . ISSN 1415-4757 .
- ^ Адл, Сина М.; Басс, Дэвид; Лейн, Кристофер Э.; Лукеш, Юлиус; Шох, Конрад Л.; Смирнов Алексей; Агата, Сабина; Берни, Седрик; Браун, Мэтью В. (26 сентября 2018 г.). «Пересмотр классификации, номенклатуры и разнообразия эукариот» . Журнал эукариотической микробиологии . 66 (1): 4–119. дои : 10.1111/jeu.12691 . ISSN 1066-5234 . ПМК 6492006 . ПМИД 30257078 .
- ^ Гирибет, Гонсало; Эджкомб, Грегори Д. (03 марта 2020 г.). Древо жизни беспозвоночных . Издательство Принстонского университета. ISBN 978-0-691-19706-7 .
- ^ ДюБук, Тимоти К.; Райан, Джозеф; Мартиндейл, Марк К. (06 февраля 2019 г.). Правда, Джон (ред.). « Дорсально-вентральные» гены являются частью древней системы формирования осевого паттерна: свидетельства Trichoplax adhaerens (Placozoa)» . Молекулярная биология и эволюция . 36 (5): 966–973. дои : 10.1093/molbev/msz025 . ISSN 0737-4038 . ПМК 6501881 . ПМИД 30726986 .
- ^ Шухерт, Питер (1 марта 1993 г.). « Trichoplax adhaerens (тип Placozoa) имеет клетки, которые реагируют с антителами против нейропептида RFамида». Акта Зоология . 74 (2): 115–117. дои : 10.1111/j.1463-6395.1993.tb01227.x . ISSN 1463-6395 .
Внешние ссылки
[ редактировать ]- Портал генома Trichoplax adhaerens Grell-BS-1999 v1.0 в Объединенном институте генома Министерства энергетики США.
- Проект генома трихоплакса в Йельском музее Пибоди
- Странный маленький зверь: Trichoplax adhaerens
- Научные статьи ITZ, TiHo Hannover
- Информационная страница Калифорнийского университета в Беркли.
- Эндер А., Шируотер Б. (январь 2003 г.). «Плакозоа не являются производными книдарий: данные молекулярной морфологии» . Мол. Биол. Эвол . 20 (1): 130–4. дои : 10.1093/molbev/msg018 . ПМИД 12519915 . – Анализ митохондриальной ДНК и 16S рРНК и филогения Trichoplax adhaerens.
- Исторический обзор трихоплакса исследований
- Science Daily: Геном простейшего животного раскрывает древнее происхождение и запутанный набор сложных способностей
- Вики Бухсбаум Пирс и Оливер Фойгт, 2007. «Полевая биология плакозойных (трихоплаксов): распространение, разнообразие, биотические взаимодействия. Интегративная и сравнительная биология» , дои : 10.1093/icb/icm015 .