альвеоляты
| альвеоляты Временной диапазон:
| |
|---|---|

| |
| Цератиевая вилка | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | потогонные средства |
| Клэйд : | ЦАРЬ |
| Клэйд : | САР |
| Клэйд : | Альвеолата Кавальер-Смит , 1991 г. |
| Фила | |
| Синонимы | |
| |
Альвеоляты (что означает «пористые , как соты») [ 2 ] — группа протистов , считающаяся основной кладой [ 3 ] и суперфилум [ 4 ] внутри Эукарьи . В настоящее время они группируются вместе со страменопилами и ризариями среди протистов с тубулокристатными митохондриями в супергруппу SAR .
Характеристики
[ редактировать ]Наиболее примечательной общей характеристикой является наличие кортикальных (около поверхности) альвеол (мешочков) . Это уплощенные везикулы (мешочки), расположенные в виде слоя непосредственно под мембраной и поддерживающие ее, обычно образующие гибкую пленку (тонкую кожу). У панцирных динофлагеллят они могут содержать жесткие пластинки. Альвеолаты имеют митохондрии с трубчатыми кристами ( инвагинациями ), а клетки часто имеют порообразные внедрения через клеточную поверхность. В группу входят свободноживущие и паразитические организмы, хищные жгутиконосцы и фотосинтезирующие организмы.

Почти все секвенированные митохондриальные геномы инфузорий и апикомплексов линейны. [ 5 ] Почти все митохондрии несут мтДНК собственную , но со значительно уменьшенным размером генома. Исключением являются Cryptosporidium , у которых осталась только митосома , круглые митохондриальные геномы Acavomonas и Babesia microti . [ 6 ] [ 7 ] и митохондриальный геном токсоплазмы высокофрагментированный , состоящий из 21 блока последовательностей, которые рекомбинируются с образованием более длинных сегментов. [ 8 ] [ 9 ]
История
[ редактировать ]Взаимосвязь апикомплексов, динофлагеллят и инфузорий была предположена в 1980-х годах и подтверждена в начале 1990-х годов сравнением последовательностей рибосомальных РНК, в первую очередь Gajadhar et al . [ 10 ] Кавальер-Смит представил официальное название Alveolata в 1991 году. [ 11 ] хотя в то время он считал эту группу парафилетической совокупностью. Многие биологи предпочитают использовать разговорное название «альвеолат». [ 12 ]
Классификация
[ редактировать ]Альвеолы включают около девяти основных и второстепенных групп. Они разнообразны по форме и, как известно, связаны различными ультраструктурными и генетическими сходствами: [ 13 ]
- Инфузории - очень распространенные простейшие с множеством коротких ресничек, расположенных рядами, и двумя ядрами.
- Акавомонидии [ 13 ]
- Колпонемидии [ 13 ]
- Динофлагелляты sl - в основном морские жгутиконосцы, многие из которых имеют хлоропласты.
- Перкинсоз
- Chromerida - морской тип фотосинтезирующих простейших.
- Колподеллида
- В Воромонаде
- Apicomplexa - паразитические и вторичные нефотосинтезирующие простейшие, у которых отсутствуют аксонемные двигательные структуры, за исключением гамет.
Acavomonidia и Colponemidia ранее были сгруппированы вместе как кольпонемиды, теперь этот таксон разделен, поскольку каждый из них имеет отличительную организацию или ультраструктурную идентичность . Acavomonidia ближе к группе динофлагеллят/перкинсид, чем Colponemidia. [ 13 ] Таким образом, неофициальный термин «кольпонемиды» в его нынешнем виде охватывает две несестринские группы внутри Alveolata: Acavomonidia и Colponemidia. [ 13 ]
Apicomplexa и динофлагелляты могут быть более тесно связаны друг с другом, чем с инфузориями. Оба имеют пластиды , и большинство из них имеют общий пучок или конус микротрубочек в верхней части клетки. У apicomplexans он образует часть комплекса, используемого для проникновения в клетки-хозяева, тогда как у некоторых бесцветных динофлагеллят он образует ножку, используемую для заглатывания добычи. С этими двумя группами тесно связаны и другие роды, в основном жгутиконосцы со сходным апикальным строением. К ним относятся свободноживущие представители Oxyrris и Colponema , а также паразиты Perkinsus , [ 14 ] Parvilucifera , Rastrimonas и эллобиопсиды . В 2001 году прямая амплификация гена рРНК в образцах морского пикопланктона выявила наличие двух новых альвеолятных линий, названных группой I и II. [ 15 ] [ 16 ] Группа I не имеет культивируемых родственников, а группа II связана с динофлагеллятным паразитом Amoebophrya , который до сих пор относился к отряду динофлагеллят Syndiniales .
Некоторые исследования предположили, что сюда могут принадлежать гаплоспориды , в основном паразиты морских беспозвоночных, но у них нет альвеол, и теперь их относят к Cercozoa .
Эллобиопсиды имеют неопределенное родство внутри альвеолятов. анализа установили, что эллобиопсиды рода Thalassomyces являются альвеолятами Зильберман и др. (2004 г.) с помощью филогенетического , однако по состоянию на 2016 г. [update] на их месте больше нет уверенности. [ 17 ] [ 18 ]
Филогения
[ редактировать ]В 2017 году Томас Кавальер-Смит описал филогению альвеол следующим образом: [ 19 ]
Таксономия
[ редактировать ]Альвеолата Кавальер-Смит 1991 [Alveolatobiontes]
- Тип Ciliophora Doflein 1901 стат. н. Коупленд, 1956 г. [Ciliata Perty, 1852 г .; Инфузория Бючли 1887 ; Реснички, Ciliozoa, Cytoidea, Eozoa, Heterokaryota, Heterokaryota]
- Подтип Postciliodesmatophora Герасимова и Серавин 1976
- Класс Heterotrichea Stein 1859 г.
- Класс Karyorelictea Corliss 1974.
- Подтип Intramacronucleata Lynn 1996
- Сорт ? Мезодинеа Чен и др. 2015 год
- Инфрафилум ламелликортиката
- Класс Litostomatea Small & Lynn, 1981 г.
- Класс Армофория Линн, 2004 г.
- Класс Cariacotricea Orsi et al. 2011 год
- Класс Spirotricea Bütschli 1889.
- Infraphylum Ventrata Cavalier-Smith 2004 [Conthreep Lynn 2012 ]
- Заказ? Дискотрихида Чен и др. 2015 год
- Класс Protocruziea Chen et al. 2015 [Protocruziidia de Puytorac, Grain & Mignot 1987 ]
- Класс Colpodea Small & Lynn, 1981 г.
- Класс Насофорея Смолл и Линн, 1981 г.
- Класс Phyllopharyngea de Puytorac et al. 1974 год
- Class Prostomatea Schewiakoff 1896
- Класс Plagiopylea Small & Lynn 1985 по версии Lynn 2008
- Класс Oligohymenophorea de Puytorac et al. 1974 год
- Подтип Postciliodesmatophora Герасимова и Серавин 1976
- Тип Miozoa Кавальер-Смит 1987 г.
- Подтип Colponemidia Тихоненков, Мыльников и Килинг 2013
- Класс Колпонема Кавальер-Смит 1993 г.
- Подтип Acavomonadia Тихоненков и др. 2014 год
- Класс Acavomonadea Тихоненков и др. 2014 год
- Подтип Myzozoa Cavalier-Smith 2004 г.
- Infraphylum Apicomplexa Levine 1970, исправление. Аддл и др. 2005 г.
- Заказ ? Витреллида Кавальер-Смит 2017
- Сорт ? Myzomonadea Cavalier-Smith & Chao 2004 sensu Ruggiero et al. 2015 год
- Класс Хромрея
- Order Colpodellida Patterson & Zölffel 1991 [Spiromonadida Krylov & Mylnikov 1986 ]
- Надкласс Sporozoa Leuckart 1879 стат. ноябрь Кавалер-Смит 2013 [Гамонтозоа]
- Класс Бластогрегариниды Chatton & Villeneuve 1936 [Blastoggregarinina; Бластогрегаринорина Чаттон и Вильнёв, 1936 ]
- Класс Парагрегареа Кавальер-Смит 2014 г.
- Класс Gregarinomorphea Грассе, 1953 г.
- Класс Coccidiomorphea Doflein 1901.
- Infraphylum Dinozoa Cavalier-Smith, 1981, исправление. 2003 г.
- Заказ ? Акроцелида Кавальер-Смит и Чао, 2004 г.
- Заказ ? Растромонадида Кавальер-Смит и Чао 2004 г.
- Класс Squirmidea Norén 1999 г., стат. ноябрь Кавалер-Смит 2014
- Надкласс Perkinsozoa Norén et al. 1999 сс
- Класс Перкинси Левин 1978 г. [Перкинсасида Левин 1978 г. ]
- Надкласс Dinoflagellata Butschli 1885 стат. ноябрь Кавалер-Смит 1999 в смысле Кавалер-Смит 2013 [Dinozoa Cavalier-Smith 1981 ]
- Класс Проноктилуцеа
- Класс Ellobiopsea Cavalier-Smith 1993 [Ellobiophyceae Loeblich III 1970 ; Эллобиопсида Уислер 1990 ]
- Класс Мизодинея Кавальер-Смит 2017 г.
- Класс Оксирея Кавальер-Смит 1987 г.
- Класс Syndinea Chatton 1920 SL [Syndiniophyceae Loeblich III 1970 SS ; Синдина Кавальер-Смит ]
- Класс Endodinea Cavalier-Smith 2017
- Класс Noctiluciphyceae Fensome et al. 1993 [Noctilucae Haeckel 1866 ; Noctilucea Haeckel 1866 стат. ноябрь ; Cystoflagellata Haeckel 1873 stat. ноябрь Бучли 1887 ]
- Класс Dinophyceae Pascher 1914 [Peridinea Ehrenberg 1830 stat. Ноябрь Веттштайн ]
- Infraphylum Apicomplexa Levine 1970, исправление. Аддл и др. 2005 г.
- Подтип Colponemidia Тихоненков, Мыльников и Килинг 2013
Разработка
[ редактировать ]Интригует развитие пластид среди альвеол. Кавалье-Смит предположил, что альвеолаты развились от предка, содержащего хлоропласты, который также дал начало Chromista ( гипотеза хромальвеолата ). Другие исследователи предположили, что у альвеолятов изначально отсутствовали пластиды и, возможно, динофлагелляты и Apicomplexa приобрели их отдельно. Однако теперь оказывается, что альвеолаты, динофлагелляты, хромериды и гетероконтовые водоросли получили свои пластиды от красной водоросли, что свидетельствует об общем происхождении этой органеллы во всех этих четырех кладах. [ 20 ]
Эволюция
[ редактировать ]По байесовским оценкам, эволюция альвеолятной группы произошла примерно 850 миллионов лет назад . [ 21 ] Альвеолы состоят из мизозоа , инфузорий и кольпонемид. Другими словами, термин Myzozoa, означающий «выкачивать содержимое из добычи», может неофициально применяться к общему предку подмножества альвеолятов, которые не являются ни инфузориями, ни колпонемидами. Хищничество водорослей является важным фактором эволюции альвеол, поскольку оно может служить источником эндосимбиоза новых пластид. Таким образом, термин Myzozoa является удобной концепцией для отслеживания истории альвеолятного типа.
Предки альвеолятной группы могли быть фотосинтезирующими. [ 22 ] Предковая альвеолята, вероятно, содержала пластиду . Хромриды, апикомплексаны и динофлагелляты перидинина сохранили эту органеллу . [ 23 ] Сделав еще один шаг назад, можно утверждать, что хромриды, перидининовые динофлагелляты и гетероконтовые водоросли обладают общей монофилетической пластидной линией, то есть приобрели свои пластиды от красной водоросли . [ 20 ] поэтому вполне вероятно, что общий предок альвеолятов и гетероконтов также был фотосинтезирующим.
В одной школе мысли общий предок динофлагеллят , апикомплексов , Colpodella , Chromerida и Voromonas был мизоцитозным хищником с двумя гетеродинамическими жгутиками , микропорами , трихоцистами , роптриями , микронемами , полярным кольцом и свернутым открытым коноидом . [ 24 ] Хотя общий предок альвеолятов также мог обладать некоторыми из этих характеристик, утверждалось, что мизоцитоз не был одной из этих характеристик, поскольку инфузории поглощают добычу по другому механизму. [ 13 ]
Продолжающиеся дебаты касаются количества мембран, окружающих пластиду у апикомплексов и некоторых динофлагеллят, а также происхождения этих мембран. Этот ультраструктурный признак можно использовать для группировки организмов, и если этот признак является общим, это может означать, что у типов был общий фотосинтетический предок. На основании того, что апикомплексаны обладают пластидой, окруженной четырьмя мембранами, а перидининовые динофлагелляты обладают пластидой, окруженной тремя мембранами, Petersen et al. [ 25 ] не смогли исключить, что общий страменопил-альвеолятный пластид мог многократно перерабатываться в альвеолятном типе, источником которого были доноры страменопила-альвеолата, посредством механизма проглатывания и эндосимбиоза .
Инфузории являются модельными альвеолятами, которые генетически изучались очень глубоко в течение самого длительного периода среди всех альвеолятных линий. Они необычны среди эукариот тем, что в размножении участвуют микронуклеус и макронуклеус . Их размножение легко изучить в лаборатории, что исторически сделало их моделью эукариот. Будучи полностью хищными и лишенными остатков пластид, их развитие как типа иллюстрирует, как хищничество и автотрофность [ 22 ] находятся в динамическом равновесии и что баланс может колебаться в ту или иную сторону в момент возникновения нового типа от миксотрофных предков, приводя к потере одной способности.
-

-
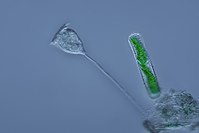 Вортицелла ( Ciliophora ) (слева)
Вортицелла ( Ciliophora ) (слева) -
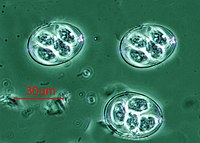
 Plasmodium falciparum ( Apicomplexa ) в крови
Plasmodium falciparum ( Apicomplexa ) в крови -
-






Эпигенетика
[ редактировать ]Лишь немногие водоросли были изучены с точки зрения эпигенетики . [ 26 ] Те, по которым доступны эпигенетические данные, включают некоторые альвеолаты водорослей. [ 26 ]
Ссылки
[ редактировать ]- ^ Ли CW и др. (2007). «Ресничные простейшие из докембрийской формации Душаньтуо, Вэньгань, Южный Китай». Геологическое общество, Лондон, специальные публикации . 286 (1): 151–6. Бибкод : 2007GSLSP.286..151L . дои : 10.1144/SP286.11 . S2CID 129584945 .
- ^ «альвеолятный» . Словарь/тезаурус Memidex (WordNet) . Архивировано из оригинала 11 апреля 2016 г. Проверено 26 января 2011 г.
- ^ Адл С. и др. (2012). «Пересмотренная классификация эукариот» . Журнал эукариотической микробиологии . 59 (5): 429–514. дои : 10.1111/j.1550-7408.2012.00644.x . ПМЦ 3483872 . ПМИД 23020233 .
- ^ Руджеро М.А., Гордон Д.П., Оррелл Т.М., Байи Н., Бургуэн Т., Бруска Р.К., Кавальер-Смит Т., Гири М.Д., Кирк П.М. (2015). «Классификация всех живых организмов более высокого уровня» . ПЛОС ОДИН . 10 (4): e0119248. Бибкод : 2015PLoSO..1019248R . дои : 10.1371/journal.pone.0119248 . ПМЦ 4418965 . ПМИД 25923521 .
- ^ Barth D, Berendonk Tu (2011). «Последовательность митохондриального генома ресничного парамециума Caudatum показывает сдвиг в нуклеотидном составе и использование кодона в рамках рода Paramecium » . BMC Genomics . 12 : 272. DOI : 10.1186/1471-2164-12-272 . PMC 3118789 . PMID 21627782 .
- ^ Оборнин М., Люкш Дж. (2015-10-15). «Органеллярные геномы хромеры и витраллы, фототрофные родственники паразитов апикомплексана» . Ежегодный обзор микробиологии . 69 : 129–144. doi : 10.1146/annurev-micro-091014-104449 . ISSN 0066-4227 . PMID 26092225 . Получено 2024-04-15 .
- ^ Корнильо Э, Хадж-Каддур К, Дассули А, Ноэль Б, Ранвез В, Вашери Б, Оганьёр Ю, Брес В, Дюкло А, Рандаццо С, Карси Б, Дебьер-Грокьего Ф, Дельбек С, Мубри-Менаж К, Шамс- Элдин Х., Усмани-Браун С., Бринго Ф., Винкер П., Виварес КП, Шварц Р.Т., Шеттерс Т.П., Краузе П.Дж., Горенфлот А., Берри В., Барбе В., Бен Мамун С. (2012). «Секвенирование наименьшего генома апикомплекса человеческого патогена Babesia microti » . Нуклеиновые кислоты Рез . 40 (18): 9102–14. дои : 10.1093/nar/gks700 . ПМК 3467087 . ПМИД 22833609 .
- ^ Намасиваям С., Баптиста Р.П., Сяо В., Холл Э.М., Доггетт Дж.С., Троэлл К., Киссинджер Дж.К. (май 2021 г.). «Новый фрагментированный митохондриальный геном возбудителя простейших Toxoplasma gondii и родственных ему тканевых кокцидий» . Геномные исследования . 31 (5): 852–865. дои : 10.1101/гр.266403.120 . ПМК 8092004 . ПМИД 33906963 .
- ^ Намасиваям С., Сан С., Бах А.Б., Оберсталлер Дж., Пьер-Луи Э., Этеридж Р.Д., Фешотт С., Притам Э.Дж., Киссинджер Дж.К. (07.11.2023). «Массивная инвазия органеллярной ДНК стимулирует эволюцию ядерного генома токсоплазмы» . Труды Национальной академии наук . 120 (45): –2308569120. Бибкод : 2023PNAS..12008569N . дои : 10.1073/pnas.2308569120 . ПМЦ 10636329 . ПМИД 37917792 .
- ^ Гаджадхар А.А. и др. (1991). «Последовательности рибосомальной РНК Sarcocystis muris , Theilera annulata и Crypthecodinium cohnii раскрывают эволюционные связи между апикомплексами, динофлагеллятами и инфузориями». Молекулярная и биохимическая паразитология . 45 (1): 147–153. дои : 10.1016/0166-6851(91)90036-6 . ПМИД 1904987 . .
- ^ Кавалер-Смит Т. (1991). «Диверсификация клеток гетеротрофных жгутиконосцев» . Паттерсон Дж., Ларсен Дж., Ассоциация систематики (ред.). Биология свободноживущих гетеротрофных жгутиконосцев . Издательство Оксфордского университета. стр. 113–131. ISBN 978-0-19-857747-8 .
- ^ Кумар, С. и Ржецкий, А. 1996. Эволюционные взаимоотношения эукариотических царств. Журнал молекулярной эволюции, 42: 183–193.
- ^ Jump up to: а б с д и ж Тихоненков Д.В., Янушковец Ю., Мыльников А.П., Михайлов К.В., Симдянов Т.Г., Алеошин В.В., Килинг П.Дж. (2014). «Описание Colponema vietnamica sp.n. и Acavomonas peruviana n. gen. n. sp., двух новых типов альвеолятов (Colponemidia nom. nov. и Acavomonidia nom. nov.) и их вклад в реконструкцию предкового состояния альвеолятов и эукариот. " . ПЛОС ОДИН . 9 (4): е95467. Бибкод : 2014PLoSO...995467T . дои : 10.1371/journal.pone.0095467 . ПМЦ 3989336 . ПМИД 24740116 .
- ^ Чжан Х., Кэмпбелл Д.А., Штурм Н.Р., Дунган К.Ф., Лин С. (2011). «Сплайсинг лидерных РНК, сдвиг рамки митохондриальных генов и мультибелковая филогения расширяют поддержку рода Perkinsus как уникальной группы альвеолатов» . ПЛОС ОДИН . 6 (5): e19933. Бибкод : 2011PLoSO...619933Z . дои : 10.1371/journal.pone.0019933 . ПМК 3101222 . ПМИД 21629701 .
- ^ Лопес-Гарсиа П., Родригес-Валера Ф., Педрос-Алио К., Морейра Д. (2001). «Неожиданное разнообразие мелких эукариот в глубоководном антарктическом планктоне». Природа . 409 (6820): 603–7. Бибкод : 2001Natur.409..603L . дои : 10.1038/35054537 . ПМИД 11214316 . S2CID 11550698 .
- ^ Мун-ван дер Стаай С.Ю., Де Вахтер Р., Валот Д. (2001). «Последовательности океанической 18S рДНК пикопланктона обнаруживают непредвиденное эукариотическое разнообразие». Природа . 409 (6820): 607–10. Бибкод : 2001Natur.409..607M . дои : 10.1038/35054541 . ПМИД 11214317 . S2CID 4362835 .
- ^ Хоппенрат М (29 апреля 2016 г.). «Таксономия динофлагеллят — обзор и предложение пересмотренной классификации». Морское биоразнообразие . 47 (2). Институт Зенкенберга ( Шпрингер ): 381–403. doi : 10.1007/s12526-016-0471-8 (неактивен 3 мая 2024 г.). ISSN 1867-1616 . S2CID 42100119 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на май 2024 г. ( ссылка ) - ^ Тейлор Ф.Дж. (2004). «Освещение или путаница? Молекулярно-филогенетические данные динофлагеллят, рассматриваемые преимущественно с морфологической точки зрения». Психологические исследования . 52 (4). Японское общество психологии ( Уайли ): 308–324. дои : 10.1111/j.1440-183.2004.00360.x . ISSN 1322-0829 . S2CID 86797666 .
- ^ Кавалер-Смит Т. (5 сентября 2017 г.). «Королевство Chromista и его восемь типов: новый синтез, подчеркивающий нацеливание на перипластидные белки, эволюцию цитоскелета и перипластид, а также древние расхождения» . Протоплазма . 255 (1): 297–357. doi : 10.1007/s00709-017-1147-3 (неактивен 3 мая 2024 г.). ПМЦ 5756292 . ПМИД 28875267 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на май 2024 г. ( ссылка ) - ^ Jump up to: а б Янушковец Дж., Горак А., Оборник М., Люкес Дж., Килинг П.Дж. (2010). «Общее красноводорослевое происхождение пластид апикомплексана, динофлагелляты и гетероконта» . Proc Natl Acad Sci США . 107 (24): 10949–54. Бибкод : 2010PNAS..10710949J . дои : 10.1073/pnas.1003335107 . ПМК 2890776 . ПМИД 20534454 .
- ^ Берни С., Павловски Дж. (2006). «Молекулярная шкала времени эволюции эукариот, перекалиброванная с помощью непрерывной записи микроокаменелостей» . Учебник по биологическим наукам . 273 (1596): 1867–72. дои : 10.1098/rspb.2006.3537 . ПМЦ 1634798 . ПМИД 16822745 .
- ^ Jump up to: а б Рейес-Прието А., Мустафа А., Бхаттачарья Д. (2008). «Множество генов, очевидно, происходящих из водорослей, позволяют предположить, что инфузории когда-то могли быть фотосинтезирующими» . Курс. Биол . 18 (13): 956–62. Бибкод : 2008CBio...18..956R . дои : 10.1016/j.cub.2008.05.042 . ПМК 2577054 . ПМИД 18595706 .
- ^ Мур Р.Б., Оборник М., Янушковец Дж., Хрудимский Т., Ванцова М., Грин Д.Х., Райт С.В., Дэвис Н.В., Болч С.Дж., Хейманн К., Слапета Дж., Хоег-Гульдберг О., Логсдон Дж.М., Картер Д.А. (2008). «Фотосинтетический альвеолят, тесно связанный с апикомплексными паразитами». Природа . 451 (7181): 959–963. Бибкод : 2008Natur.451..959M . дои : 10.1038/nature06635 . ПМИД 18288187 . S2CID 28005870 .
- ^ Кувардина О.Н., Леандер Б.С., Алешин В.В., Мыльников А.П., Килинг П.Дж., Симдянов Т.Г. (2002). «Филогения колподеллид (Alveolata) с использованием последовательностей генов рРНК малых субъединиц предполагает, что они являются свободноживущей сестринской группой апикомплексов» . J Эукариотная микробиол . 49 (6): 498–504. дои : 10.1111/j.1550-7408.2002.tb00235.x . ПМИД 12503687 . S2CID 4283969 .
- ^ Петерсен Дж., Людвиг А.К., Михаэль В., Банк Б., Ярек М., Баурен Д., Бринкманн Х. (2014). « Chromera velia , эндосимбиозы и гипотеза родоплекса - пластидная эволюция у криптофитов, альвеолят, страменопилей и гаптофитов (линии CASH)» . Геном Биол Эвол . 6 (3): 666–684. дои : 10.1093/gbe/evu043 . ПМЦ 3971594 . ПМИД 24572015 .
- ^ Jump up to: а б Пайферрер Ф., Ван Х.П., ред. (2023). Эпигенетика в аквакультуре . Уайли. стр. 100-1 383–411. дои : 10.1002/9781119821946 . hdl : 10261/191758 . ISBN 978-1-119-82191-5 .