Генетическая кластеризация человека
Генетическая кластеризация человека относится к закономерностям относительного генетического сходства между отдельными людьми и популяциями, а также к широкому спектру научных и статистических методов, используемых для изучения этого аспекта генетической изменчивости человека .
Кластерные исследования считаются ценными для характеристики общей структуры генетических вариаций среди человеческих популяций, а также способствуют изучению происхождения предков, истории эволюции и точной медицины. После картирования человеческого генома и доступности все более мощных аналитических инструментов кластерный анализ выявил ряд наследственных и миграционных тенденций среди человеческих популяций и отдельных людей. [1] Генетические кластеры человека, как правило, организованы по географическому происхождению, при этом разделения между кластерами в основном совпадают с географическими барьерами, такими как океаны или горные хребты. [2] [3] Кластерные исследования были применены к населению всего мира, [4] а также к подгруппам населения, таким как постколониальная Северная Америка. [5] [6] Примечательно, что практика определения кластеров среди современных человеческих популяций во многом произвольна и изменчива из-за непрерывной природы человеческих генотипов; хотя отдельные генетические маркеры можно использовать для создания меньших групп, не существует моделей, которые создавали бы совершенно разные подгруппы при использовании большего количества генетических маркеров. [2] [7] [8]
Многие исследования генетической кластеризации человека были вовлечены в дискуссии о расе , этнической принадлежности и научном расизме , поскольку некоторые противоречиво предположили, что генетически полученные кластеры можно понимать как доказательство генетически детерминированных рас. [9] [10] Хотя кластерный анализ неизменно объединяет людей (или группы людей) в подгруппы, продолжаются дебаты о том, как интерпретировать эти генетические кластеры в отношении расы, ее социальных и фенотипических особенностей. А поскольку в целом доля генетических вариаций между генотипами человека очень мала, подходы к генетической кластеризации сильно зависят от выборочных данных, генетических маркеров и статистических методов, применяемых для их построения.
Алгоритмы и методы генетической кластеризации
[ редактировать ]Разработан широкий спектр методов оценки структуры популяций человека с использованием генетических данных. В ранних исследованиях внутригрупповых и межгрупповых генетических вариаций использовались физические фенотипы и группы крови, а в современных генетических исследованиях используются генетические маркеры, такие как последовательности Alu , полиморфизмы коротких тандемных повторов и однонуклеотидные полиморфизмы (SNP) и другие. [11] Модели генетической кластеризации также различаются в зависимости от алгоритмов и программ, используемых для обработки данных. Наиболее сложные методы определения кластеров можно отнести к методам кластеризации на основе моделей (например, алгоритм STRUCTURE [12] ) или многомерные сводки (обычно посредством анализа главных компонентов). [1] [13] Обрабатывая большое количество SNP (или других данных генетических маркеров) разными способами, оба подхода к генетической кластеризации имеют тенденцию сходиться к схожим закономерностям путем выявления сходства между SNP и/или трактами гаплотипов для выявления наследственных генетических сходств. [13]
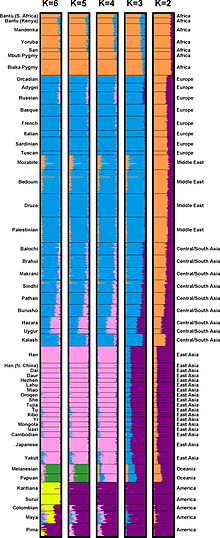
Общие алгоритмы кластеризации на основе моделей включают STRUCTURE, ADMIXTURE и HAPMIX. Эти алгоритмы работают путем поиска наилучшего соответствия генетических данных среди произвольного или математически полученного количества кластеров, так что различия внутри кластеров сводятся к минимуму, а различия между кластерами максимизируются. Этот метод кластеризации также называют « выводом о примесях », поскольку отдельные геномы (или особи внутри популяции) могут быть охарактеризованы пропорциями аллелей , связанных с каждым кластером. [1] Другими словами, такие алгоритмы, как STRUCTURE, генерируют результаты, которые предполагают существование дискретных предковых популяций, реализуемых с помощью уникальных генетических маркеров, которые со временем объединились, чтобы сформировать смешанные популяции современных дней.
Многомерная сводная статистика
[ редактировать ]В то время как кластеризация на основе моделей характеризует популяции с использованием пропорций предполагаемых предковых кластеров, многомерная сводная статистика характеризует популяции в непрерывном спектре. Наиболее распространенным многомерным статистическим методом, используемым для генетической кластеризации, является анализ главных компонентов (PCA), который отображает людей по двум или более осям (их «главным компонентам»), которые представляют собой скопления генетических маркеров, которые отвечают за наибольшую дисперсию. Затем кластеры можно идентифицировать путем визуальной оценки распределения данных; при более крупных выборках генотипов человека данные имеют тенденцию группироваться в отдельные группы, а также в смешанные позиции между группами. [1] [13]
Предостережения и ограничения
[ редактировать ]Существуют предостережения и ограничения для методов генетической кластеризации любого типа, учитывая степень примеси и относительного сходства внутри человеческой популяции. Все результаты генетических кластеров искажаются процессом выборки, используемым для сбора данных, а также качеством и количеством этих данных. Например, во многих исследованиях кластеризации используются данные, полученные от популяций, которые географически различаются и находятся далеко друг от друга, что может создавать иллюзию дискретных кластеров, тогда как в действительности популяции гораздо более смешаны друг с другом, когда включены промежуточные группы. [1] Размер выборки также играет важную сдерживающую роль в результатах кластеризации, поскольку разные исходные данные по размеру выборки могут влиять на распределение кластеров, а более тонкие связи между генотипами могут возникнуть только при увеличении размера выборки. [1] [8] В частности, использование STRUCTURE подверглось широкой критике как потенциально вводящее в заблуждение, поскольку требует сортировки данных в заранее определенное количество кластеров, которые могут отражать, а могут и не отражать фактическое распределение населения. [8] [14] Создатели STRUCTURE первоначально описали алгоритм как « исследовательский » метод, который следует интерпретировать с осторожностью, а не как тест со статистически значимой мощностью. [12] [15]
Известные приложения к генетическим данным человека
[ редактировать ]Современные применения методов генетической кластеризации к генетическим данным глобального масштаба были впервые отмечены исследованиями, связанными с данными Проекта разнообразия генома человека (HGDP). [1] Эти ранние исследования HGDP, например, Rosenberg et al. (2002), [4] [16] внес свой вклад в теорию эффекта серийного основателя и ранней миграции людей из Африки, а методы кластеризации особенно применялись для описания смешанных континентальных популяций. [5] [6] [17] Генетическая кластеризация и исследования HGDP также способствовали разработке методов и критике индустрии потребительского тестирования генетического происхождения . [18]
С 2002 года на популяциях людей по всему миру был проведен ряд знаковых генетических кластерных исследований, в том числе следующие:
| Авторы | Год | Заголовок | Размер выборки/количество отобранных групп населения | Образец | Маркеры |
|---|---|---|---|---|---|
| Розенберг и др. | 2002 | Генетическая структура человеческих популяций [19] | 1056 / 52 | Проект разнообразия генома человека (HGDP-CEPH) | 377 СПО |
| Серре и Паабо | 2004 | Человеческие отношения во всем мире, выведенные из общегеномных закономерностей изменчивости [20] | 89 / 15 | а: ВВПП | 20 СПО |
| 90 / географически распределенных лиц | б: Земля 1997 г. | ||||
| Розенберг и др. | 2005 | Клины, кластеры и влияние дизайна исследования на структуру человеческой популяции [21] | 1056 / 52 | Проект разнообразия генома человека (HGDP-CEPH) | 783 STR + 210 удалений |
| Ли и др. | 2008 | Человеческие отношения во всем мире, выведенные из общегеномных закономерностей изменчивости [22] | 938 / 51 | Проект разнообразия генома человека (HGDP-CEPH) | 650 000 SNP |
| Тишкофф и др. | 2009 | Генетическая структура и история африканцев и афроамериканцев [23] | ~3400 / 185 | HGDP-CEPH плюс 133 дополнительных африканских населения и индейцев | 1327 STR + делений |
| Син и др. | 2010 | На пути к более единообразной выборке генетического разнообразия человека: исследование населения мира с помощью генотипирования высокой плотности [24] | 850 / 40 | HapMap плюс 296 человек | 250 000 SNP |
Генетическая кластеризация и раса
[ редактировать ]Группы людей часто имеют географическую структуру . Например, при кластеризации населения Восточной Азии и европейцев каждая группа, скорее всего, сформирует свой собственный кластер на основе схожих частот аллелей . [25] Таким образом, кластеры могут коррелировать с традиционными представлениями о расе и самоидентифицированном происхождении; в некоторых случаях, например, в медицинских анкетах, последние переменные могут использоваться в качестве показателя генетического происхождения, когда генетические данные недоступны. [9] [4] Однако генетические вариации распространяются сложным, непрерывным и перекрывающимся образом, поэтому эта корреляция несовершенна, и использование расовых категорий в медицине может создать дополнительные опасности. [9]
Некоторые ученые [ ВОЗ? ] поставили под сомнение идею о том, что расу можно определить по генетическим кластерам, проводя различия между произвольно назначенными генетическими кластерами, происхождением и расой. Одним из повторяющихся предостережений против рассмотрения человеческих популяций в терминах кластеров является представление о том, что генотипические вариации и черты распределяются между популяциями равномерно, вдоль постепенных линий, а не вдоль дискретных границ популяций; поэтому, хотя генетическое сходство обычно организовано географически, лежащие в его основе популяции никогда не были полностью отделены друг от друга. Из-за миграции, потока генов и исходной однородности особенности между группами сильно перекрываются и переплетаются. [2] [9] Более того, генетические кластеры обычно не соответствуют социально определенным расовым группам; многие общепонятные расы не могут быть отнесены к одному и тому же генетическому кластеру, и многие генетические кластеры состоят из людей, которые имеют различную расовую идентичность. [7] В общем, кластеры проще всего понимать как продукты методов, используемых для выборки и анализа генетических данных; не лишено значения для понимания происхождения и генетических характеристик, но недостаточно для полного объяснения концепции расы, которую чаще описывают с точки зрения социальных и культурных сил.
В контексте персонализированной медицины раса в настоящее время считается фактором риска широкого спектра заболеваний, имеющих генетические и негенетические причины. Возникли вопросы относительно того, поддерживают ли генетические кластеры идею расы как действительную конструкцию, применимую к медицинским исследованиям и лечению заболеваний, поскольку существует множество заболеваний, которые соответствуют конкретным генетическим маркерам и/или конкретным популяциям, как это видно на примере Тэя. -Болезнь Сакса или серповидно-клеточная анемия . [3] [26] Исследователи стараются подчеркнуть, что происхождение, частично выявляемое с помощью кластерного анализа, играет важную роль в понимании риска заболеваний. Но расовая или этническая идентичность не полностью соответствует генетическому происхождению, поэтому раса и этническая принадлежность не дают достаточно информации для постановки медицинского диагноза. [26] Раса как переменная в медицине с большей вероятностью отражает социальные факторы, где информация о происхождении с большей вероятностью будет иметь значение при рассмотрении генетического происхождения. [2] [26]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г Новембре, Джон; Рамачандран, Сохини (22 сентября 2011 г.). «Перспективы структуры человеческой популяции на пороге эры секвенирования» . Ежегодный обзор геномики и генетики человека . 12 (1): 245–274. doi : 10.1146/annurev-genom-090810-183123 . ISSN 1527-8204 . ПМИД 21801023 .
- ^ Перейти обратно: а б с д Магло, Коффи Н.; Мерша, Тесфайе Б.; Мартин, Лиза Дж. (17 февраля 2016 г.). «Популяционная геномика и статистические значения расы: междисциплинарный взгляд на биологическую классификацию человеческих популяций и значение для клинических генетических эпидемиологических исследований» . Границы генетики . 7:22 . дои : 10.3389/fgene.2016.00022 . ISSN 1664-8021 . ПМЦ 4756148 . ПМИД 26925096 .
- ^ Перейти обратно: а б Гудман, Алан Х.; Моисей, Иоланда Т.; Джонс, Джозеф Л., ред. (29 октября 2012 г.). Раса . дои : 10.1002/9781118233023 . ISBN 9781118233023 .
- ^ Перейти обратно: а б с Розенберг, Н.А. (20 декабря 2002 г.). «Генетическая структура человеческих популяций» . Наука . 298 (5602): 2381–2385. Бибкод : 2002Sci...298.2381R . дои : 10.1126/science.1078311 . ISSN 0036-8075 . ПМИД 12493913 . S2CID 8127224 .
- ^ Перейти обратно: а б Хан, Ынчжон; Карбонетто, Питер; Кертис, Росс Э.; Ван, Юн; Гранка, Джули М.; Бирнс, Джейк; Ното, Кейт; Кермани, Амир Р.; Майрес, Натали М.; Барбер, Мэтью Дж.; Рэнд, Кристин А. (07 февраля 2017 г.). «Кластеризация 770 000 геномов раскрывает постколониальную структуру населения Северной Америки» . Природные коммуникации . 8 (1): 14238. Бибкод : 2017NatCo...814238H . дои : 10.1038/ncomms14238 . ISSN 2041-1723 . ПМК 5309710 . ПМИД 28169989 .
- ^ Перейти обратно: а б Джордан, И. Кинг; Ришишвар, Лаванья; Конли, Эндрю Б. (сентябрь 2019 г.). «Примесь коренных американцев повторяет миграцию и расселение населения континентальной части Соединенных Штатов» . ПЛОС Генетика . 15 (9): e1008225. дои : 10.1371/journal.pgen.1008225 . ISSN 1553-7404 . ПМЦ 6756731 . ПМИД 31545791 .
- ^ Перейти обратно: а б Бамшад, Майкл Дж.; Олсон, Стив Э. (декабрь 2003 г.). «Существует ли раса?» . Научный американец . 289 (6): 78–85. Бибкод : 2003SciAm.289f..78B . doi : 10.1038/scientificamerican1203-78 . ISSN 0036-8733 . ПМИД 14631734 .
- ^ Перейти обратно: а б с Калиновский, СТ (4 августа 2010 г.). «Компьютерная программа STRUCTURE не позволяет достоверно определить основные генетические кластеры внутри видов: моделирование и последствия для структуры человеческой популяции» . Наследственность . 106 (4): 625–632. дои : 10.1038/hdy.2010.95 . ISSN 0018-067X . ПМК 3183908 . ПМИД 20683484 .
- ^ Перейти обратно: а б с д Джорд, Линн Б; Вудинг, Стивен П. (26 октября 2004 г.). «Генетическая изменчивость, классификация и «раса» » . Природная генетика . 36 (С11): С28–С33. дои : 10.1038/ng1435 . ISSN 1061-4036 . ПМИД 15508000 .
- ^ Маркс, Джонатан (27 февраля 2017 г.). Является ли наука расистской? . Джон Уайли и сыновья. ISBN 978-0-7456-8925-8 . OCLC 1037867598 .
- ^ Бамшад, Майкл; Вудинг, Стивен; Солсбери, Бенджамин А.; Стивенс, Дж. Клэйборн (август 2004 г.). «Деконструкция связи между генетикой и расой» . Обзоры природы Генетика . 5 (8): 598–609. дои : 10.1038/nrg1401 . ISSN 1471-0056 . PMID 15266342 . S2CID 12378279 .
- ^ Перейти обратно: а б Причард, Джонатан К; Стивенс, Мэтью; Доннелли, Питер (1 июня 2000 г.). «Вывод о структуре населения с использованием данных мультилокусного генотипа» . Генетика . 155 (2): 945–959. дои : 10.1093/генетика/155.2.945 . ISSN 1943-2631 . ПМК 1461096 . ПМИД 10835412 .
- ^ Перейти обратно: а б с Лоусон, Дэниел Джон; Фалуш, Дэниел (22 сентября 2012 г.). «Идентификация населения с использованием генетических данных» . Ежегодный обзор геномики и генетики человека . 13 (1): 337–361. doi : 10.1146/annurev-genom-082410-101510 . ISSN 1527-8204 . ПМИД 22703172 .
- ^ Лоусон, Дэниел Дж.; ван Дорп, Люси; Фалуш, Дэниел (14 августа 2018 г.). «Учебное пособие о том, как не переоценивать гистограммы СТРУКТУРЫ и ПРИМЕШЕНИЯ» . Природные коммуникации . 9 (1): 3258. Бибкод : 2018NatCo...9.3258L . дои : 10.1038/s41467-018-05257-7 . ISSN 2041-1723 . ПМК 6092366 . ПМИД 30108219 .
- ^ Новембре, Джон (01 октября 2016 г.). «Притчард, Стивенс и Доннелли о структуре населения» . Генетика . 204 (2): 391–393. дои : 10.1534/genetics.116.195164 . ISSN 1943-2631 . ПМК 5068833 . ПМИД 27729489 .
- ^ Розенберг, Ной А; Махаджан, Саураб; Рамачандран, Сохини ; Чжао, Чэнфэн; Причард, Джонатан К; Фельдман, Маркус В. (9 декабря 2005 г.). «Клины, кластеры и влияние дизайна исследования на структуру человеческой популяции» . ПЛОС Генетика . 1 (6): е70. дои : 10.1371/journal.pgen.0010070 . ISSN 1553-7404 . ПМК 1310579 . ПМИД 16355252 .
- ^ Лесли, Стивен; Винни, Брюс; Хеллентал, Гарретт; Дэвисон, Дэн; Бумертит, Абдельхамид; Дэй, Тэмми; Хутник, Катажина; Ройрвик, Эллен К.; Канлифф, Барри; Лоусон, Дэниел Дж.; Фалуш, Дэниел (март 2015 г.). «Мелкомасштабная генетическая структура британского населения» . Природа . 519 (7543): 309–314. Бибкод : 2015Natur.519..309. . дои : 10.1038/nature14230 . ISSN 1476-4687 . ПМК 4632200 . ПМИД 25788095 .
- ^ Роял, Чармейн Д.; Новембре, Джон; Фуллертон, Стефани М.; Гольдштейн, Дэвид Б.; Лонг, Джеффри С.; Бамшад, Майкл Дж.; Кларк, Эндрю Г. (14 мая 2010 г.). «Вывод о генетическом происхождении: возможности, проблемы и последствия» . Американский журнал генетики человека . 86 (5): 661–673. дои : 10.1016/j.ajhg.2010.03.011 . ISSN 0002-9297 . ПМК 2869013 . ПМИД 20466090 .
- ^ Розенберг, Ной А.; Причард, Джонатан К.; Вебер, Джеймс Л.; Канн, Ховард М.; Кидд, Кеннет К.; Животовский Лев А.; Фельдман, Маркус В. (20 декабря 2002 г.). «Генетическая структура человеческих популяций». Наука . 298 (5602): 2381–2385. Бибкод : 2002Sci...298.2381R . дои : 10.1126/science.1078311 . ISSN 0036-8075 . ПМИД 12493913 . S2CID 8127224 .
- ^ Серр, Дэвид; Паабо, Сванте (сентябрь 2004 г.). «Доказательства градиентов генетического разнообразия человека внутри и между континентами» . Геномные исследования . 14 (9): 1679–1685. дои : 10.1101/гр.2529604 . ISSN 1088-9051 . ПМК 515312 . ПМИД 15342553 .
- ^ Розенберг, Н.А.; Махаджан, С; Рамачандран, С; Чжао, К; Притчард, Дж. К. ; и др. (2005). «Клины, кластеры и влияние дизайна исследования на структуру человеческой популяции» . ПЛОС Генет . 1 (6): е70. дои : 10.1371/journal.pgen.0010070 . ПМЦ 1310579 . ПМИД 16355252 .
- ^ Ли, Цзюнь З.; Абшер, Девин М.; Тан, Хуа; Саутвик, Одри М.; Касто, Аманда М.; Рамачандран, Сохини; Канн, Ховард М.; Барш, Грегори С.; Фельдман, Маркус; Кавалли-Сфорца, Луиджи Л.; Майерс, Ричард М. (22 февраля 2008 г.). «Всемирные человеческие отношения, выведенные из общегеномных закономерностей вариаций». Наука . 319 (5866): 1100–1104. Бибкод : 2008Sci...319.1100L . дои : 10.1126/science.1153717 . ISSN 0036-8075 . ПМИД 18292342 . S2CID 53541133 .
- ^ Тишкофф, Сара А; Рид, Флойд А; Фридлендер, Франсуаза Р; Эрет, Кристофер; Ранчиаро, Алессия; Фроман, Ален; Хирбо, Джибрил Б; Авомойи, Агнес А; Бодо, Жан-Мари; Думбо, Огобара; Ибрагим, Мунтасер; Джума, Абдалла Т; Котце, Марита Дж; Лема, Годфри; Мур, Джейсон Х; Мортенсен, Холли; Ньямбо, Томас Б; Омар, Сабах А; Пауэлл, Квели; Преториус, Гидеон С; Смит, Майкл В.; Тера, Махамаду А; Вамбебе, Чарльз; Вебер, Джеймс Л; Уильямс, Скотт М. (22 мая 2009 г.). «Генетическая структура и история африканцев и афроамериканцев» . Наука . 324 (5930): 1035–1044. Бибкод : 2009Sci...324.1035T . дои : 10.1126/science.1172257 . ISSN 0036-8075 . ПМЦ 2947357 . ПМИД 19407144 .
- ^ Син, Цзиньчуань; Уоткинс, В. Скотт; Шлиен, Адам; Уокер, Эрин; Хафф, Чад Д.; Уизерспун, Дэвид Дж.; Чжан, Юхуа; Симонсон, Татум С.; Вайс, Роберт Б.; Шиффман, Джошуа Д.; Малкин, Дэвид; Вудворд, Скотт Р.; Джорд, Линн Б. (октябрь 2010 г.). «На пути к более единообразной выборке генетического разнообразия человека: исследование населения мира с помощью генотипирования высокой плотности» . Геномика . 96 (4): 199–210. дои : 10.1016/j.ygeno.2010.07.004 . ISSN 0888-7543 . ПМК 2945611 . ПМИД 20643205 .
- ^ Спенсер, Куэйшон (2014). «Радикальное решение расовой проблемы» . Философия науки . 81 (5): 1029-30. дои : 10.1086/677694 .
- ^ Перейти обратно: а б с Кениг, Барбара А. Ли; Су-Джин, Сандра; Ричардсон, Сара С. (2008). Возвращение к расе в эпоху генома . Издательство Университета Рутгерса. ISBN 978-0-8135-4323-9 . OCLC 468194495 .