Стимулятор генов интерферонов
| Стинг1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | Sting1 , Eris, Mita, Mpys, Net23, Savi, Sting, Hmita, HSTING, стимулятор генов интерферонов, трансмембранного белка 173, STING-BETA, TMEM173, стимулятор реакции интерферона CGAMP INGACTOR 1 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Омим : 612374 ; MGI : 1919762 ; Гомологен : 18868 ; GeneCards : Sting1 ; OMA : Sting1 - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викидид | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Стимулятор генов интерферонов ( STING ), также известный как трансмембранный белок 173 ( TMEM173 ) и MPYS / MITA / ERIS, является белком , который у людей кодируется геном STING1 . [ 5 ]
Стинг играет важную роль в врожденном иммунитете . Стинг индуцирует продукцию интерферона I типа , когда клетки инфицированы внутриклеточными патогенами, такими как вирусы , микобактерии и внутриклеточные паразиты . [ 6 ] Интерферон типа I , опосредованный укусом, защищает инфицированные клетки и близлежащие клетки от локальной инфекции путем связывания с той же клеткой, которая выделяет ее ( аутокринная передача сигналов) и близлежащие клетки ( сигнализация паракрина .) Таким образом, он играет важную роль, например, в контроле. норовирусная инфекция. [ 7 ]
STING работает как прямого цитозольного датчика ДНК (CDS), так и адаптерный белок в передаче передачи интерферонов I типа с помощью различных молекулярных механизмов. Было показано, что он активирует нижестоящие транскрипционные факторы STAT6 и IRF3 через TBK1 , которые ответственны за противовирусный ответ и врожденный иммунный ответ против внутриклеточного патогена . [ 8 ]
Структура
[ редактировать ]
Аминокислоты 1–379 укуса человека включают в себя 4 трансмембранные области (TMS) и C-концевой домен . С -концевой домен (CTD: аминокислоты 138–379) содержит домен димеризации (DD) и карбокси-концевой хвост (CTT: аминокислоты 340–379). [ 8 ]
Стинг образует симметричный димер в ячейке. Странс Димер напоминает бабочку, с глубокой щелью между двумя протомерами. Гидрофобные . остатки из каждого протомера укуса образуют гидрофобные взаимодействия между собой на границе раздела [ 8 ] [ 9 ]
Выражение
[ редактировать ]Стинг экспрессируется в гематопоэтических клетках в периферических лимфоидных тканях , включая Т -лимфоциты , NK -клетки , миелоидные клетки и моноциты . Также было показано, что укус высоко экспрессируется в легких, яичнике , сердце, гладких мышцах , сетчатке , костном мозге и влагалите. [ 10 ] [ 11 ]
Локализация
[ редактировать ]Субклеточная локализация укуса была выяснена как белок эндоплазматического ретикулума . Кроме того, вполне вероятно, что Стинг ассоциируется в непосредственной близости с митохондриями, связанными с мембраной ER (MAM) -границей между митохондрионом и ER. [ 12 ] Во время внутриклеточной инфекции укус способен пересекаться от эндоплазматического ретикулума в перинуклеарные везикулы, потенциально участвующие в транспорте, опосредованном экзоцистами . [ 12 ] Было также показано, что укус колокализуется с белками аутофагии, световой цепи белка 1, ассоциированной с микротрубочками, и белком 9A, связанного с аутофагией , после двухцепочечной стимуляции ДНК, что предполагает его присутствие в аутофагосоме . [ 13 ]
Функция
[ редактировать ]Стинг опосредует продукцию интерферона типа I в ответ на внутриклеточную ДНК и различные внутриклеточные патогены, включая вирусы , внутриклеточные бактерии и внутриклеточные паразиты . [ 14 ] После инфекции укус из инфицированных клеток может ощущать присутствие нуклеиновых кислот из внутриклеточных патогенов, а затем индуцировать интерферон β и более 10 форм продуцирования интерферона α . Интерферон типа I, продуцируемый инфицированными клетками, может найти и связываться с интерфероном-альфа-рецептором близлежащих клеток для защиты клеток от локальной инфекции.
Антивирусный иммунитет
[ редактировать ]Стинг вызывает мощный иммунитет интерферона I типа против вирусной инфекции. После вирусного въезда вирусные нуклеиновые кислоты в цитозоле инфицированных клеток присутствуют . Несколько датчиков ДНК, таких как DAI , РНК -полимераза III , IFI16 , DDX41 и CGA , могут обнаружить инородные нуклеиновые кислоты . После распознавания вирусной ДНК датчики ДНК инициируют сигнальные пути ниже по течению путем активации священного ответа интерферона. [ 15 ]
Было показано, что аденовирус , вирус простого простого герпеса , ВЕСВ-1 и HSV-2, а также вирус РНК отрицательной цепи , вирус везикулярного стоматита (VSV) способны активировать зависимый врожденный иммунный ответ . [ 14 ]
Дефицит укуса у мышей привел к смертельной восприимчивости к инфекции HSV-1 из-за отсутствия успешного ответа интерферона I типа. [ 16 ]
Точечная мутация серина -358 ослабляет активацию STING -IFN в летучих мышах и предлагается дает летучим мыши их способность служить хозяевами резервуара. [ 17 ]
Против внутриклеточных бактерий
[ редактировать ]внутриклеточные бактерии, Listeria Monocytogenes , стимулируют иммунный ответ хозяина посредством укуса. Было показано, что [ 18 ] Стинг может играть важную роль в производстве хемокинов MCP-1 и CCL7 . Моноциты с дефицитом укуса являются по своей природе дефектны при миграции в печень во время Listeria Monocytogenes инфекции . Таким образом, Sting защищает хозяина от Listeria Monocytogenes Infection, регулируя моноцитов миграцию . Активация укуса, вероятно, будет опосредована циклическим Di-AMP, секретируемым внутриклеточными бактериями. [ 18 ] [ 19 ]
Другой
[ редактировать ]Стинг может быть важной молекулой для защитного иммунитета против инфекционных организмов. Например, животные, которые не могут экспрессировать укус, более подвержены инфекции от VSV , HSV-1 и Listeria Monocytogenes , что предполагает его потенциальную корреляцию с инфекционными заболеваниями человека. [ 20 ]
Роль в иммунитете хозяина
[ редактировать ]Хотя IFN типа I абсолютно критичен для устойчивости к вирусам, существует растущая литература о негативной роли интерферона I типа в иммунитете хозяина, опосредованной укусом. что ДНК-мотива ДНК-петли с стеблем в плазмодие Falciparum и Plasmodium Berghei геноме и внеклеточной ДНК из туберкулеза Mycobacterium Было показано, активируют интерферон типа I через укус. [ 21 ] [ 22 ] Перфорация фагосомной мембраны, опосредованной системой секреции ESX1 , позволяет внеклеточной микобактериальной ДНК доступа к датчикам цитозольной ДНК -хозяина, тем самым вызывая продукцию интерферона типа I в макрофагах. Спись интерферона с высоким типом приводит к патогенезу M. tuberculosis и длительной инфекции. [ 22 ] Опосредованный STING-TBK1-IRF Реакция интерферона типа I является центральным в патогенезе экспериментальной мозговой малярии у лабораторных животных, инфицированных Plasmodium berghei . Лабораторные мыши с дефицитом в ответе интерферона I типа устойчивы к экспериментальной головной малярии. [ 21 ]
Механизмы сигнализации
[ редактировать ]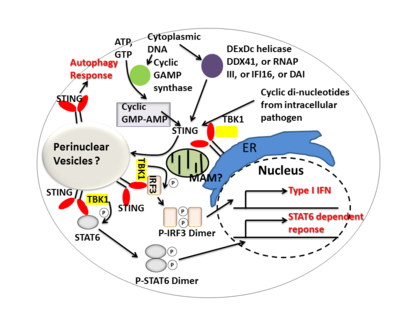
Стинг опосредует иммунный ответ типа I , функционируя как прямой датчик ДНК и белок сигнального адаптера . После активации укус стимулирует TBK1 активность для фосфорилирования IRF3 или STAT6 . Фосфорилированные IRF3S и STAT6S Dimerize, а затем введите ядро, чтобы стимулировать экспрессию генов, участвующих в иммунном ответе хозяина, таких как IFNB , CCL2 , CCL20 и т. Д. [ 8 ] [ 23 ]
Несколько сообщений свидетельствуют о том, что укус связан с активацией селективной аутофагии. [ 13 ] Mycobacterium tuberculosis Было показано, что продуцирует цитозольные ДНК -лиганды, которые активируют укусы, что приводит к убиквитинированию бактерий и последующему рекрутированию белков, связанных с аутофагией , которые необходимы для «селективного» аутофагического нацеливания и индингной защиты от М. Туберкулеза . [ 24 ]
Таким образом, STING координирует множественные иммунные реакции на инфекцию, включая индукцию интерферонов и STAT6-зависимый ответ и селективный ответ аутофагии. [ 8 ]
Как цитозольный датчик ДНК
[ редактировать ]Циклические динуклеотиды -секундные сигнальные молекулы, продуцируемые разнообразными видами бактерий, были обнаружены в цитозоле клеток млекопитающих во время внутриклеточной инфекции патогена; Это приводит к активации TBK1 - IRF3 и нижестоящему производству I. интерферона типа [ 8 ] [ 25 ] Было показано, что укус связывается непосредственно с циклическим Di-GMP , и это распознавание приводит к производству цитокинов , таких как интерферон типа I , которые необходимы для успешной элиминации патогенов. [ 26 ]
Как сигнальный адаптер
[ редактировать ]DDX41 , член семейства геликаз DEXDC , в миелоидных дендритных клетках распознает внутриклеточную ДНК и опосредует врожденный иммунный ответ посредством прямой связи с укусом. [ 27 ] Также было показано , что другие датчики ДНК- дай , РНК-полимераза III , IFI16 , активируют укусы через прямые или косвенные взаимодействия. [ 15 ]
Циклическая GMP-AMP-синтаза (CGAS), которая принадлежит к семейству нуклеотидилтрансферазы , способна распознавать содержание цитозольной ДНК и индуцировать стризависимый ответ интерферона путем продуцирования вторичного циклического гуанозин-монофосфата-монофосфата (циклический GMP-amp или cgamp). После циклического GMP-AMP активации IRF3 Sting Activation он усиливает активность TBK1 до фосфорилирования и STAT6 для отклика интерферона вниз по течению I. типа [ 28 ] [ 29 ]
Было предложено, что внутриклеточный кальций играет важную роль в реакции пути укуса. [ 30 ]
Смотрите также
[ редактировать ]- Агонист укуса - компонент страниц врожденной иммунной системы,
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в ENSG00000288243 GRCH38: ENSEMBL Release 89: ENSG00000184584, ENSG00000288243 - ENSEMBL , май 2017
- ^ Jump up to: а беременный в GRCM38: Ensembl Release 89: Ensmusg00000024349 - Ensembl , май 2017 г.
- ^ «Человеческая PubMed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ «Мышь Pubmed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ «Стинг1 стимулятор интерферона ответа CGAMP Interactor 1 [Homo Sapiens (Human)]» .
- ^ Nakhaei P, Hiscott J, Lin R (Jun 2010). «Стинг по антивирусному пути» . Журнал молекулярной клеточной биологии . 2 (3): 110–2. doi : 10.1093/jmcb/mjp048 . PMID 20022884 .
- ^ NYU P, Miao Z, Li Y, Bansal R, Peppelenbosch MP, Pan Q (2021). «CGAS-Sting эффективно ограничивает мышиную инфекцию норовируса, но противодействует противовирусному действию N-конце RIG-I в макрофагах мыши» . Кишечные микробы . 13 (1): 1959839. DOI : 10.1080/19490976.2021.1959839 . ISSN 1949-0976 . PMC 8344765 . PMID 34347572 .
- ^ Jump up to: а беременный в дюймовый и фон Burdette DL, Vance Re (январь 2013 г.). «Стинг и врожденный иммунный ответ на нуклеиновые кислоты в цитозоле». Природа иммунология . 14 (1): 19–26. doi : 10.1038/ni.2491 . PMID 23238760 . S2CID 7968532 .
- ^ Shu C, Yi G, Watts T, Kao CC, Li P (июль 2012). «Структура укуса, связанная с циклическим DI-GMP, выявляет механизм циклического распознавания динуклеотидов иммунной системой» . Природа структурная и молекулярная биология . 19 (7): 722–4. doi : 10.1038/nsmb.2331 . PMC 3392545 . PMID 22728658 .
- ^ «Эст выражения профиля выражения TMEM173» . Biogps org . biogps.org.
- ^ «NCBI TMEM173 Экспрессия Геопрофиль» . NCBI . www.ncbi.nlm.nih.gov/geoprofiles.
- ^ Jump up to: а беременный Ишикава Х, Барбер Г.Н. (октябрь 2008 г.). «Стинг - это эндоплазматический адаптер ретикулума, который облегчает врожденную иммунную передачу сигналов» . Природа . 455 (7213): 674–8. Bibcode : 2008natur.455..674i . doi : 10.1038/nature07317 . PMC 2804933 . PMID 18724357 .
- ^ Jump up to: а беременный Сайто Т., Фудзита Н., Хаяси Т., Такахара К., Сато Т., Ли Х, Мацунага К., Кагеяма С., Омори Х, Нода Т., Ямамото Н., Каваи Т., Ишии К., Такеучи О, Йошимори Т., Акира С. (декабрь 2009 ) «ATG9A контролирует динамическую транслокацию укуса и врожденного иммунного ответа, управляемая дцДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (49): 20842–6. Bibcode : 2009pnas..10620842S . doi : 10.1073/pnas.0911267106 . PMC 2791563 . PMID 19926846 .
- ^ Jump up to: а беременный Барбер Г.Н. (февраль 2011 г.). «Пути чувствительности врожденной иммунной ДНК: Sting, AIMII и регуляция продукции интерферона и воспалительных реакций» . Текущее мнение в иммунологии . 23 (1): 10–20. doi : 10.1016/j.coi.2010.12.015 . PMC 3881186 . PMID 21239155 .
- ^ Jump up to: а беременный Keating SE, Baran M, Bowie AG (Dec 2011). «Цитозольные датчики ДНК, регулирующие индукцию интерферона типа I типа» (PDF) . Тенденции в иммунологии . 32 (12): 574–81. doi : 10.1016/j.it.2011.08.004 . HDL : 2262/68041 . PMID 21940216 .
- ^ MA Z, Damania B (февраль 2016 г.). «Проход защиты CGAS Sting и его противодействие вирусами» . Ячейка и микроб . 19 (2): 150–8. doi : 10.1016/j.chom.2016.01.010 . PMC 4755325 . PMID 26867174 .
- ^ Xie J, Li Y, Shen X, Got G, Zhu Y, Cui J, Wang L, Shi Z, Zhou P (март 2018 г.). «Украшенная активация интерферона в летучих мышах» . Ячейка и микроб . 23 (3): 297–301.e4. doi : 10.1016/j.chom.2018.01.006 . PMC 7104992 . PMID 29478775 .
- ^ Jump up to: а беременный Джин Л., Гетахун А., Ноулз Х.М., Моган Дж., Акерлунд Л.Дж., Паккард Т.А., Перро А.Л., Камбиер Дж.С. (март 2013 г.). «Sting/Mpys опосредует защиту хозяина от инфекции Listeria Monocytogenes, регулируя миграцию моноцитов LY6C (HI)» . Журнал иммунологии . 190 (6): 2835–43. doi : 10.4049/jimmunol.1201788 . PMC 3593745 . PMID 23378430 .
- ^ Woodward JJ, Iavarone AT, Portnoy DA (Jun 2010). «C-DI-AMP, секретируемый внутриклеточными Listeria Monocytogenes, активирует реакцию интерферона типа I-хозяина» . Наука . 328 (5986): 1703–5. Bibcode : 2010sci ... 328.1703W . doi : 10.1126/science.1189801 . PMC 3156580 . PMID 20508090 .
- ^ Ишикава H, Ma Z, Barber GN (октябрь 2009 г.). «Стинг регулирует внутриклеточный ДНК-опосредованный, интерферон-зависимый врожденный иммунитет типа I» . Природа . 461 (7265): 788–92. Bibcode : 2009natur.461..788i . doi : 10.1038/nature08476 . PMC 4664154 . PMID 19776740 .
- ^ Jump up to: а беременный Sharma S, Deoliveira RB, Kalantari P, Parroche P, Goutagny N, Jiang Z, Chan J, Bartholomeu DC, Lauw F, Hall JP, Barber GN, Gazzinelli RT, Fitzgerald KA, Golenbock DT (август 2011 г.). «Врожденное иммунное распознавание богатого AT-мотива ДНК ствола в геноме Plasmodium falciparum» . Иммунитет . 35 (2): 194–207. doi : 10.1016/j.immuni.2011.05.016 . PMC 3162998 . PMID 21820332 .
- ^ Jump up to: а беременный Manzanillo PS, Shiloh Mu, Portnoy Da, Cox JS (май 2012). «Mycobacterium tuberculosis активирует ДНК-зависимый путь цитозольного наблюдения в макрофагах» . Ячейка и микроб . 11 (5): 469–80. doi : 10.1016/j.chom.2012.03.007 . PMC 3662372 . PMID 22607800 .
- ^ Chen H, Sun H, You F, Sun W, Zhou X, Chen L, Yang J, Wang Y, Tang H, Guan Y, Xia W, Gu J, Ishikawa H, Gutman D, Barber G, Qin Z, Jiang Z (Октябрь 2011). «Активация STAT6 STING имеет решающее значение для противовирусного врожденного иммунитета» . Клетка . 147 (2): 436–46. doi : 10.1016/j.cell.2011.09.022 . PMID 22000020 .
- ^ Watson Ro, Manzanillo PS, Cox JS (август 2012 г.). «Внеклеточный M. DNA Tuberculosis нацелен на бактерии для аутофагии путем активации пути чувствительного ДНК-хозяина» . Клетка . 150 (4): 803–15. doi : 10.1016/j.cell.2012.06.040 . PMC 3708656 . PMID 22901810 .
- ^ McWhirter SM, Barbalat R, Monroe KM, Fontana MF, Hyodo M, Joncker NT, Ishii KJ, Akira S, Colonna M, Chen ZJ, Fitzgerald Ka, Hayakawa Y, Vance Re (август 2009 г.). «Ответ интерферона типа IS-типа I-типа индуцируется цитозольным восприятием бактериального второго мессенджера Cyclic-Di-GMP» . Журнал экспериментальной медицины . 206 (9): 1899–911. doi : 10.1084/jem.20082874 . PMC 2737161 . PMID 19652017 .
- ^ Burdette DL, Monroe KM, Sotelo-Roha K, Iwig JS, Eckert B, Hyodo M, Hayakawa Y, Vance Re (октябрь 2011). «Стинг-это прямой врожденный иммунный датчик циклического DI-GMP» . Природа . 478 (7370): 515–8. Bibcode : 2011natur.478..515b . doi : 10.1038/nature10429 . PMC 3203314 . PMID 21947006 .
- ^ Zhang Z, Yuan B, Bao M, Lu N, Kim T, Liu YJ (октябрь 2011 г.). «Геликаза DDX41 ощущает внутриклеточную ДНК, опосредованную адаптером в дендритных клетках» . Природа иммунология . 12 (10): 959–65. doi : 10.1038/ni.2091 . PMC 3671854 . PMID 21892174 .
- ^ Wu J, Sun L, Chen X, Du F, Shi H, Chen C, Chen ZJ (февраль 2013 г.). «Циклический GMP-AMP-это эндогенный второй посланник в врожденной иммунной передаче сигналов цитозольной ДНК» . Наука . 339 (6121): 826–30. Bibcode : 2013sci ... 339..826W . doi : 10.1126/science.1229963 . PMC 3855410 . PMID 23258412 .
- ^ Sun L, Wu J, Du F, Chen X, Chen ZJ (февраль 2013 г.). «Циклическая GMP-AMP-синтаза представляет собой цитозольный датчик ДНК, который активирует путь интерферона типа I» . Наука . 339 (6121): 786–91. Bibcode : 2013sci ... 339..786s . doi : 10.1126/science.1232458 . PMC 3863629 . PMID 23258413 .
- ^ Ким С., Кох П., Ли Л., Пишкин Л., Митчисон Т.Дж. (4 июня 2017 г.). «Свидетельство о роли кальция в сигнализации STING» Biorxiv 10.1101/ 1
Дальнейшее чтение
[ редактировать ]- Wang Y, Tong X, Omoregie ES, Liu W, Meng S, Ye X (октябрь 2012 г.). «Тетраспанин 6 (TSPAN6) отрицательно регулирует рецептор-рецептор-опосредованную, опосредованную рецептором, опосредованной рецептором, в зависимости от убиквитинирования» . Журнал биологической химии . 287 (41): 34626–34. doi : 10.1074/jbc.m112.390401 . PMC 3464568 . PMID 22908223 .
- Инь К., Тянь Ю., Кабалесваран В., Цзян Х, Ту Д., Эк М.Дж., Чен З.Дж., Ву Х (июнь 2012 г.). «Циклическое ощущение DI-GMP через врожденное иммунное сигнальное утирование белка» . Молекулярная клетка . 46 (6): 735–45. doi : 10.1016/j.molcel.2012.05.029 . PMC 3697849 . PMID 22705373 .
- Aguirre S, Maestre AM, Pagni S, Patel JR, Savage T, Gutman D, Maringer K, Bernal-Rubio D, Shabman RS, Simon V, Rodriguez-Madoz JR, Mulder LC, Barber GN, Fernandez-Sesma A (2012) Полем «DENV ингибирует выработку IFN типа в инфицированных клетках, расщепляя укус человека» . PLO -патогены . 8 (10): E1002934. doi : 10.1371/journal.ppat.1002934 . PMC 3464218 . PMID 23055924 .
- Li Y, Li C, Xue P, Zhong B, Mao AP, Ran Y, Chen H, Wang Yy, Yang F, Shu HB (май 2009 г.). «ISG56 является регулятором отрицательной обратной связи передаваемой вирусом передачи сигналов и клеточного противовирусного ответа» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (19): 7945–50. Bibcode : 2009pnas..106.7945L . doi : 10.1073/pnas.0900818106 . PMC 2683125 . PMID 19416887 .
- Конлон Дж., Бурдетт Д.Л., Шарма С., Бхат Н., Томпсон М., Цзян З., Ратинам В.А., Монахи Б., Джин Т., Сяо Т.С., Фогель С.Н., Вэнс Р.Е., Фицджеральд К.А. (май 2013). «Мышь, но не укус человека, связывает и сигнализирует в ответ на сосудистый агент, нарушающий 5,6-диметилксантенон-4-уксусную кислоту» . Журнал иммунологии . 190 (10): 5216–25. doi : 10.4049/jimmunol.1300097 . PMC 3647383 . PMID 23585680 .
- Абэ Т., Харасима А., Ся Т., Конно Х., Конно К., Моралес А., Ан Дж, Гутман Д., Барбер Г.Н. (апрель 2013). «Опознание укуса цитоплазматической ДНК спровоцирует клеточную защиту» . Молекулярная клетка . 50 (1): 5–15. doi : 10.1016/j.molcel.2013.01.039 . PMC 3881179 . PMID 23478444 .
- Назми А., Мухопадхьяй Р., Датта К., Басу А. (2012). «Стинг опосредует нейрональный врожденный иммунный ответ после инфекции вируса японского энцефалита» . Научные отчеты . 2 : 347. Bibcode : 2012natsr ... 2e.347n . doi : 10.1038/srep00347 . PMC 3317237 . PMID 22470840 .
- Zhang J, Hu MM, Wang Yy, Shu HB (август 2012 г.). «Белок TRIM32 модулирует индукцию интерферона I типа и клеточный противовирусный ответ, нацеляя MITA/STING-белок для K63-связанного убиквитинирования» . Журнал биологической химии . 287 (34): 28646–55. doi : 10.1074/jbc.m112.362608 . PMC 3436586 . PMID 22745133 .
- Ишикава Х, Барбер Г.Н. (октябрь 2008 г.). «Стинг - это эндоплазматический адаптер ретикулума, который облегчает врожденную иммунную передачу сигналов» . Природа . 455 (7213): 674–8. Bibcode : 2008natur.455..674i . doi : 10.1038/nature07317 . PMC 2804933 . PMID 18724357 .