Белок 1 распознавания пептидогликана
| PGLYRP1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | PGLYRP1 , PGLYRP, PGRP, PGRP-S, PGRPS, TAG7, TNFSF3L, белок распознавания пептидогликана 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 604963 ; МГИ : 1345092 ; Гомологен : 74539 ; Генные карты : PGLYRP1 ; ОМА : PGLYRP1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||






Белок распознавания пептидогликана 1 , PGLYRP1 , также известный как TAG7 , представляет собой антибактериальный и провоспалительный врожденного иммунитета белок , который у человека кодируется PGLYRP1 геном . [ 5 ] [ 6 ] [ 7 ] [ 8 ]
Открытие
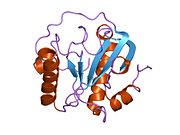
[ редактировать ]PGLYRP1 был открыт независимо двумя лабораториями в 1998 году. [ 5 ] [ 7 ] Хокан Штайнер и его коллеги, используя экран дифференциального дисплея, идентифицировали и клонировали белок распознавания пептидогликана (PGRP) у бабочки ( Trichoplusia ni ) и на основе этой последовательности обнаружили и клонировали ортологи PGRP мыши и человека. [ 5 ] Сергей Киселев и его коллеги обнаружили и клонировали белок аденокарциномы мыши с той же последовательностью, что и мышиный PGRP, который они назвали Tag7. [ 7 ] Человеческий PGRP был одним из основателей семейства из четырех генов PGRP, обнаруженных у человека, которые были названы PGRP-S, PGRP-L, PGRP-Iα и PGRP-Iβ (для коротких, длинных и средних транскриптов по аналогии с транскриптами насекомых). PGRP). [ 9 ] на PGLYRP1 (белок распознавания пептидогликана 1), PGLYRP2 ( белок распознавания пептидогликана 2 ), PGLYRP3 ( белок распознавания пептидогликана 3 ) и PGLYRP4 ( белок распознавания пептидогликана 4 Их генные символы были впоследствии изменены Комитетом по номенклатуре генов организации генома человека ) соответственно. , и эта номенклатура в настоящее время также используется для других PGRP млекопитающих . В 2005 году Рой Мариуцца и его коллеги кристаллизовали человеческий PGLYRP1 и расшифровали его структуру. [ 10 ]
Распределение и секреция в тканях
[ редактировать ]PGLYRP1 имеет самый высокий уровень экспрессии среди всех PGRP млекопитающих. PGLYRP1 высоко конститутивно экспрессируется в костном мозге. [ 5 ] [ 9 ] [ 11 ] и в третичных гранулах полиморфно-ядерных лейкоцитов ( нейтрофилов и эозинофилов ), [ 11 ] [ 12 ] [ 13 ] [ 14 ] [ 15 ] [ 16 ] и в меньшей степени в активированных макрофагах [ 15 ] [ 16 ] и печень плода . [ 9 ] PGLYRP1 также экспрессируется в лактирующей молочной железе . [ 17 ] и в гораздо более низком уровне в эпителии глаза роговицы , [ 18 ] в воспаленной коже , [ 19 ] [ 20 ] селезенка , [ 5 ] тимус , [ 5 ] и в эпителиальных клетках дыхательных путей [ 15 ] [ 16 ] и кишечного тракта. [ 5 ] [ 21 ] PGLYRP1 заметно экспрессируется в кишечных пейеровых бляшках в микроскладчатых (М) клетках. [ 22 ] [ 23 ] а также является одним из маркеров дифференцировки Т-хелперов 17 (Th17) в Т-регуляторные (Treg) клетки у мышей. [ 24 ] Экспрессия PGLYRP1 в Т-клетках увеличивается под действием IL-27 . [ 25 ] Мышиный PGLYRP1 экспрессируется в развивающемся мозге , и на эту экспрессию влияет кишечный микробиом . [ 26 ] Экспрессия PGLYRP1 в мозге крыс индуцируется лишением сна [ 27 ] и в мозге мыши в результате ишемии . [ 28 ]
Человеческий PGLYRP1 также обнаруживается в сыворотке после высвобождения из лейкоцитарных гранул путем экзоцитоза . [ 29 ] [ 30 ] PGLYRP1 присутствует в верблюжьем молоке в концентрации 120 мкг/мл. [ 17 ] а в гранулах полиморфно-ядерных лейкоцитов - 2,9 мг/10 9 клетки. [ 11 ]
Структура
[ редактировать ]Как и большинство PGRP, PGLYRP1 имеет один карбокси-концевой пептидогликан-связывающий амидазный домен типа 2 (также известный как домен PGRP), который, однако, не обладает ферментативной амидазной активностью. [ 31 ] Этот домен PGRP состоит из трех альфа-спиралей, пяти бета-цепей и катушек, а также N-концевого сегмента (остатки 1–30, PGRP-специфического сегмента), структура которого существенно различается среди PGRP. [ 10 ] PGLYRP1 имеет три пары консервативных цистеинов , которые образуют три дисульфидные связи в положениях 9 и 133, 25 и 70, а также 46 и 52 в PGLYRP1 человека. [ 10 ] Дисульфид Cys46-Cys52 широко консервативен в PRGP беспозвоночных и позвоночных , дисульфид Cys9-Cys133 консервативен во всех PGRP млекопитающих, а дисульфид Cys25-Cys70 уникален для PGLYRP1, PGLYRP3 и PGLYRP4 млекопитающих, но не обнаружен в амидазо-активном PGLYRP2. [ 9 ] [ 10 ] PGLYRP1 человека имеет пептидогликансвязывающую щель длиной 25 Å, стенки которой образованы двумя α-спиралями, а дно - β-листом. [ 10 ]
PGLYRP1 человека секретируется и образует гомодимеры с дисульфидной связью. [ 30 ] [ 32 ] Структура димера с дисульфидной связью неизвестна, поскольку была решена кристаллическая структура только мономерного человеческого PGLYRP1, поскольку в кристаллизованном белке отсутствовали 8 N-концевых аминокислот, включая Cys8, [ 10 ] который, вероятно, участвует в образовании димера с дисульфидной связью. Крысиный PGLYRP1 также, вероятно, образует димеры с дисульфидной связью, поскольку он содержит Cys в том же положении, что и Cys8 в PGLYRP1 человека. [ 27 ] тогда как мышь [ 5 ] и бычий [ 11 ] PGLYRP1 не содержит этого Cys и, вероятно, не образует димеров с дисульфидной связью.
Camel PGLYRP1 может образовывать два димера, не связанных дисульфидной связью: первый с сайтами связывания пептидогликана двух участвующих молекул, полностью экспонированными на противоположных концах димера, а второй с сайтами связывания пептидогликана, скрытыми на границе раздела и открытыми противоположными сторонами. на концах димера. [ 33 ] Такое расположение уникально для верблюда PGLYRP1. [ 33 ]
PGLYRP1 гликозилирован , и гликозилирование необходимо для его бактерицидной активности. [ 32 ] [ 34 ]
Функции
[ редактировать ]Белок PGLYRP1 играет важную роль во врожденном иммунном ответе.
Связывание пептидогликана
[ редактировать ]PGLYRP1 связывает пептидогликан , полимер β(1-4)-связанного N -ацетилглюкозамина (GlcNAc) и N -ацетилмураминовой кислоты (MurNAc), сшитый короткими пептидами , основной компонент бактерий клеточной стенки . [ 5 ] [ 9 ] [ 12 ] [ 14 ] [ 35 ] [ 36 ] [ 37 ] PGLYRP1 человека связывает трипептид GlcNAc-MurNAc с высоким сродством (Kd = 5,5 x 10 −8 М) [ 14 ] и MurNAc-трипептид, MurNAc-тетрапептид и MurNAc-пентапептид с Kd = 0,9–3,3 x 10. −7 М [ 36 ] с предпочтением мезодиаминопимелиновой кислоты ( м -ДАФ) перед L- лизинсодержащими фрагментами пептидогликана. [ 36 ] m -DAP присутствует в третьем положении пептидогликана у грамотрицательных бактерий и грамположительных бацилл, тогда как L-лизин находится в этом положении у пептидогликана у грамположительных кокков. Меньшие фрагменты пептидогликана не связываются или связываются с гораздо меньшим сродством. [ 14 ] [ 36 ] PGLYRP1 также связывается с пептидогликаном с орнитином в третьем положении пептидогликана, обнаруженного в спирохете Borrelia burgdorferi . [ 37 ]
Camel PGLYRP1 связывает MurNAc-дипептид с низким сродством (Kd = 10 −7 М) [ 38 ] а также связывает бактериальный липополисахарид с Kd = 1,6 x 10 −9 М и липотейхоевая кислота с Kd = 2,4 х 10 −8 M в сайтах связывания за пределами канонической щели, связывающей пептидогликан, с лигандами и PGLYRP1, образующими тетрамеры. [ 39 ] Такие тетрамеры уникальны для верблюжьего PGLYRP1 и не обнаружены в человеческом PGLYRP1 из-за стеариновых затруднений. [ 39 ]
Бактерицидная активность
[ редактировать ]PGLYRP1 человека оказывает прямое бактерицидное действие как на грамположительные ( Bacillus subtilis , Bacillus licheniformis , Lactobacillus acidophilus , Staphylococcus aureus , Streptococcus pyogenes ), так и на грамотрицательные ( Escherichia coli , Proteus vulgaris , Salmonella enterica , Shigella sonnei , Pseudomonas) . aeruginosa ) бактерии, [ 12 ] [ 14 ] [ 32 ] [ 34 ] [ 40 ] и спирохета Borrelia burgdorferi . [ 37 ] PGLYRP1 ограничивает внутриклеточную выживаемость Listeria monocytogenes в макрофагах [ 41 ] а также активен в отношении Chlamydia trachomatis . [ 42 ] Мышь [ 12 ] [ 13 ] и бычий [ 11 ] [ 29 ] PGLYRP1 обладает антибактериальной активностью в отношении Bacillus megaterium , Staphylococcus hemolyticus , S. aureus , E. coli и S. enterica , а бычий PGLYRP1 также обладает противогрибковой активностью в отношении Cryptococcus neoformans . [ 11 ]
В грамположительных бактериях человеческий PGLYRP1 связывается с сайтами разделения новообразованных дочерних клеток, созданными бактериальными пептидогликанлитическими эндопептидазами, LytE и LytF в B. subtilis , которые отделяют дочерние клетки после клеточного деления. [ 40 ] Эти эндопептидазы, разделяющие клетки, вероятно, подвергают PGLYRP1-связывающим мурамиловым пептидам, о чем свидетельствует совместная локализация PGLYRP1, LytE и LytF в сайтах разделения клеток и отсутствие связывания PGLYRP1 с другими областями клеточной стенки с сильно сшитым пептидогликаном. . [ 40 ] Эта локализация необходима для уничтожения бактерий, поскольку мутанты , у которых отсутствуют эндопептидазы LytE и LytF и которые не разделяются после деления клеток, не связывают PGLYRP1, а также с трудом уничтожаются PGLYRP1. [ 40 ] У грамотрицательных бактерий ( E. coli ) PGLYRP1 связывается с внешней мембраной. [ 40 ] Как у грамположительных, так и у грамотрицательных бактерий PGLYRP1 остается связанным с клеточной оболочкой и не проникает в цитоплазму . [ 40 ]
Механизм уничтожения PGLYRP1 основан на индукции летального стресса оболочки и продукции активных форм кислорода у бактерий и последующем отключении транскрипции и трансляции . [ 40 ] Уничтожение бактерий, индуцированное PGLYRP1, не включает проницаемость клеточной мембраны, которая типична для дефензинов и других антимикробных пептидов , гидролиз клеточной стенки или осмотический шок . [ 32 ] [ 34 ] [ 40 ]
Человеческий PGLYRP1 обладает синергической бактерицидной активностью с лизоцимом. [ 14 ] и антибактериальные пептиды. [ 34 ] Стрептококки продуцируют белок (SP1), который ингибирует антибактериальную активность человеческого PGLYRP1. [ 43 ]
Защита от инфекций
[ редактировать ]PGLYRP1 играет ограниченную роль в защите хозяина от большинства инфекций. Мыши с дефицитом PGLYRP1 более чувствительны к системным инфекциям, вызванным непатогенными бактериями ( Micrococcus luteus и B. subtilis ). [ 13 ] и P. aeruginosa , вызванному кератиту , [ 18 ] но не системным инфекциям патогенными бактериями ( S. aureus и E. coli ). [ 13 ] Внутривенное введение PGLYRP1 защищает мышей от системной инфекции Listeria monocytogenes . [ 44 ]
PGLYRP1 также защищает от инфекции B. burgdorferi , поскольку у мышей, лишенных PGLYRP1, увеличивается количество спирохет в сердце и суставах, но не в коже, что указывает на роль PGLYRP1 в контроле распространения B. burgdorferi во время системной фазы инфекции. [ 37 ]
Поддержание микробиома
[ редактировать ]Мышиный PGLYRP1 играет роль в поддержании здорового микробиома, поскольку у мышей с дефицитом PGLYRP1 наблюдаются значительные изменения в составе микробиомов кишечника и легких, которые влияют на их чувствительность к колиту и воспалению легких. [ 21 ] [ 45 ] [ 46 ]
Влияние на воспаление
[ редактировать ]Мышиный PGLYRP1 играет роль в поддержании противовоспалительного гомеостаза в кишечнике, коже, легких, суставах, лимфоидных органах, глазах и мозге. Мыши с дефицитом PGLYRP1 более чувствительны, чем мыши дикого типа, к колиту, индуцированному декстран-сульфатом натрия (DSS), что указывает на то, что PGLYRP1 защищает мышей от колита, индуцированного DSS. [ 21 ] [ 45 ] Однако в мышиной модели артрита у мышей с дефицитом PGLYRP1 развивается более тяжелый артрит, чем у мышей дикого типа. [ 47 ] Кроме того, у мышей с дефицитом как PGLYRP1 , так и PGLYRP2 развивается более тяжелый артрит, чем у мышей с дефицитом PGLYRP2 , которые устойчивы к артриту. [ 47 ] Эти результаты показывают, что PGLYRP2 способствует развитию артрита и что PGLYRP1 противодействует провоспалительному эффекту PGLYRP2. [ 47 ] У мышей с дефицитом PGLYRP1 также наблюдается нарушение заживления ран роговицы по сравнению с мышами дикого типа, что указывает на то, что PGLYRP1 способствует заживлению ран роговицы. [ 18 ]
Мыши с дефицитом PGLYRP1 более устойчивы, чем мыши дикого типа, к экспериментально индуцированной аллергической астме . [ 15 ] [ 16 ] атопический дерматит , [ 20 ] контактный дерматит , [ 20 ] и псориазоподобное воспаление кожи. [ 19 ] Эти результаты показывают, что мышиный PGLYRP1 способствует воспалению легких и кожи. Эти провоспалительные эффекты обусловлены увеличением количества и активности Т-хелперов 17 (Th17) и уменьшением количества Т-регуляторных (Treg) клеток. [ 15 ] [ 19 ] [ 20 ] а в случае астмы также увеличивается количество Т-хелперов 2 (Th2) и снижается количество плазмацитоидных дендритных клеток . [ 15 ] Провоспалительное действие PGLYRP1 на астму зависит от регулируемого PGLYRP1 кишечного микробиома, поскольку эта повышенная устойчивость к экспериментально индуцированной аллергической астме может быть передана стерильным мышам дикого типа путем трансплантации микробиома от мышей с дефицитом PGLYRP1 . [ 46 ]
Мыши с дефицитом PGLYRP1 также защищены от экспериментального аутоиммунного энцефаломиелита (EAE), а PGLYRP1 способствует патологии заболевания EAE. [ 25 ] Экспрессия PGLYRP1 в моноцитах/макрофагах и нейтрофилах, но не в Т-клетках, необходима для оптимальной презентации антигена и праймирования CD4. + Т-клетки в патогенезе ЭАЭ. [ 25 ]
Однако PGLYRP1 выполняет противоположную (т.е. ингибирующую) функцию в Т-клетках. Мыши с дефицитом PGLYRP1 обладают повышенным противоопухолевым иммунитетом и замедлением роста опухоли. [ 25 ] PGLYRP1 экспрессируется совместно с ингибирующими генами CD8. + инфильтрирующие опухоль Т-лимфоциты (TIL) с признаком истощения CD8 + Т-клетки как у мышей, так и у людей, [ 25 ] что делает эти клетки менее эффективными в противоопухолевом иммунитете. Таким образом, удаление PGLYRP1 в Т-клетках позволяет повысить противоопухолевый иммунитет. [ 25 ]
У мышей, лишенных PGLYRP1, инфицированных B. burgdorferi, также наблюдаются признаки иммунной дисрегуляции, включая более низкие уровни IgG в сыворотке и более высокие уровни провоспалительных цитокинов и хемокинов, IFNγ , CXCL9 и CXCL10 . [ 37 ] Таким образом, отсутствие Pglyrp1 у этих мышей приводит к цитокиновому ответу Th1, одновременно ухудшая ответ антител к B. burgdorferi . [ 37 ]
PGLYRP1, слитый с Fc-областью мышиного IgG2a, увеличивает выживаемость и облегчает повреждение и воспаление легких в мышиной модели E. coli , вызванного острого респираторного дистресс-синдрома , не влияя на клиренс бактерий. [ 48 ] Эта конструкция PGLYRP1-Fc подавляет активацию макрофагов посредством механизма, зависимого от гамма-рецептора Fc (FcγR), тем самым уменьшая воспалительное повреждение легких. [ 48 ]
Цитотоксичность
[ редактировать ]Сообщалось, что мышиный PGLYRP1 (Tag7) является цитотоксичным для опухолевых клеток и действует как подобный фактору некроза опухоли-α (TNF-α) цитокин, . [ 7 ] Последующие эксперименты показали, что PGLYRP1 (Tag7) сам по себе не обладает цитотоксической активностью. [ 12 ] [ 49 ] но что PGLYRP1 (Tag7) образует комплекс с белком теплового шока 70 ( Hsp70 ) и что только эти комплексы цитотоксичны для опухолевых клеток, [ 49 ] тогда как PGLYRP1 (Tag7) сам по себе действует как антагонист цитотоксичности комплексов PGLYRP1-Hsp70. [ 50 ]
Взаимодействие с белками и рецепторами хозяина
[ редактировать ]PGLYRP1 человека и мыши (Tag7) связывается с белком теплового шока 70 ( Hsp70 ) в растворе, а комплексы PGLYRP1-Hsp70 также секретируются цитотоксическими лимфоцитами, и эти комплексы цитотоксичны для опухолевых клеток. [ 49 ] [ 51 ] Этой цитотоксичности противодействует метастазин ( S100A4 ). [ 52 ] и белок, связывающий тепловой шок HspBP1 . [ 53 ] Комплексы PGLYRP1-Hsp70 связываются с TNFR1 ( рецептором фактора некроза опухоли-1 , который является рецептором смерти) и вызывают цитотоксический эффект посредством апоптоза и некроптоза . [ 50 ] Эта цитотоксичность связана с проницаемостью лизосом и митохондрий . [ 54 ] Напротив, свободный PGLYRP1 действует как антагонист TNFR1, связываясь с TNFR1 и ингибируя его активацию комплексами PGLYRP1-Hsp70. [ 50 ] Пептиды человеческого PGLYRP1 ингибируют цитотоксические эффекты комплексов TNF-α и PGLYRP1-Hsp70. [ 55 ] и продукция цитокинов в мононуклеарных клетках периферической крови человека. [ 56 ] Они также уменьшают воспалительные реакции на мышиной модели острого повреждения легких. [ 56 ] и при полном артрите, вызванном адъювантом Фрейнда, у мышей. [ 57 ]
Человеческий PGLYRP1 в комплексе с пептидогликаном или мультимеризованный связывается и стимулирует TREM-1 (триггерный рецептор, экспрессируемый на миелоидных клетках-1), рецептор, присутствующий на нейтрофилах, моноцитах и макрофагах, который индуцирует выработку провоспалительных цитокинов. [ 25 ] [ 58 ]
Медицинская значимость
[ редактировать ]Генетические варианты PGLYRP1 или измененная экспрессия PGLYRP1 часто связаны с различными заболеваниями. Пациенты с воспалительным заболеванием кишечника (ВЗК), включая болезнь Крона и язвенный колит , имеют значительно более частые миссенс-варианты в гене PGLYRP1 (а также в трех других генах PGLYRP ), чем здоровые люди из контрольной группы. [ 59 ] Эти результаты позволяют предположить, что PGLYRP1 защищает людей от этих воспалительных заболеваний и что мутации гена PGLYRP1 являются одними из генетических факторов, предрасполагающих к этим заболеваниям. Варианты PGLYRP1 также связаны с повышением уровня гемоглобина плода при серповидно-клеточной анемии . [ 60 ]
Несколько заболеваний связаны с повышенной экспрессией PGLYRP1, в том числе: атеросклероз , [ 61 ] [ 62 ] инфаркт миокарда , [ 63 ] [ 64 ] ишемическая болезнь сердца , [ 65 ] [ 66 ] сердечная недостаточность , [ 66 ] сепсис , [ 67 ] воспаленные ткани при болезни Крона и язвенном колите , [ 68 ] [ 69 ] фиброз легких , [ 70 ] астма, [ 71 ] хроническая болезнь почек , [ 72 ] ревматоидный артрит , [ 73 ] воспаление десен , [ 74 ] [ 75 ] [ 76 ] [ 77 ] [ 78 ] [ 79 ] кариес и заболевания мышц и суставов, [ 80 ] остеоартрит , [ 81 ] сердечно-сосудистые события и смерть у перенесших трансплантацию почки , пациентов, [ 82 ] алопеция , [ 83 ] сердечная недостаточность , [ 84 ] диабет I типа , [ 85 ] инфекционные осложнения при гемодиализе , [ 86 ] и тромбоз , [ 87 ] согласуется с провоспалительным действием PGLYRP1. Более низкая экспрессия PGLYRP1 была обнаружена при эндометриозе . [ 88 ] Концентрация PGLYRP1 в сыворотке пуповинной крови обратно пропорциональна детской астме и функции легких в подростковом возрасте. [ 89 ]
См. также
[ редактировать ]- Белок распознавания пептидогликана
- Белок 2 распознавания пептидогликана
- Белок 3 распознавания пептидогликана
- Белок 4 распознавания пептидогликана
- Пептидогликан
- Врожденная иммунная система
- Стенки бактериальных клеток
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000008438 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000030413 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Перейти обратно: а б с д и ж г час я Канг Д., Лю Г., Лундстрем А., Гелиус Э., Штайнер Х. (август 1998 г.). «Белок, распознающий пептидогликан, во врожденном иммунитете, сохраняющийся от насекомых до человека» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (17): 10078–10082. Бибкод : 1998PNAS...9510078K . дои : 10.1073/pnas.95.17.10078 . ПМК 21464 . ПМИД 9707603 .
- ^ «Ген Энтрез: белок 1 распознавания пептидогликана PGLYRP1» .
- ^ Перейти обратно: а б с д Киселев С.Л., Кустикова О.С., Коробко Е.В., Прохорчук Е.Б., Кабышев А.А., Луканидин Е.М., Георгиев Г.П. (июль 1998 г.). «Молекулярное клонирование и характеристика мышиного гена tag7, кодирующего новый цитокин» . Журнал биологической химии . 273 (29): 18633–18639. дои : 10.1074/jbc.273.29.18633 . ПМИД 9660837 . S2CID 11417742 .
- ^ «Белок 1 распознавания пептидогликана Pglyrp1 [Mus musculus (домовая мышь)] - Ген - NCBI» . www.ncbi.nlm.nih.gov . Проверено 3 ноября 2020 г.
- ^ Перейти обратно: а б с д и Лю С, Сюй З, Гупта Д, Дзиарски Р (сентябрь 2001 г.). «Белки распознавания пептидогликана: новое семейство из четырех молекул распознавания образов врожденного иммунитета человека» . Журнал биологической химии . 276 (37): 34686–34694. дои : 10.1074/jbc.M105566200 . ПМИД 11461926 . S2CID 44619852 .
- ^ Перейти обратно: а б с д и ж Гуан Р., Ван К., Сундберг Э.Дж., Мариуцца Р.А. (апрель 2005 г.). «Кристаллическая структура белка S распознавания пептидогликана человека (PGRP-S) с разрешением 1,70 А». Журнал молекулярной биологии . 347 (4): 683–691. дои : 10.1016/j.jmb.2005.01.070 . ПМИД 15769462 .
- ^ Перейти обратно: а б с д и ж Тайделл CC, Йонт Н., Тран Д., Юань Дж., Селстед М.Э. (май 2002 г.). «Выделение, характеристика и антимикробные свойства бычьего олигосахаридсвязывающего белка. Микробицидный гранулярный белок эозинофилов и нейтрофилов» . Журнал биологической химии . 277 (22): 19658–19664. дои : 10.1074/jbc.M200659200 . ПМИД 11880375 . S2CID 904536 .
- ^ Перейти обратно: а б с д и Лю С., Гелиус Э., Лю Г., Штайнер Х., Дзиарски Р. (август 2000 г.). «Белок распознавания пептидогликана млекопитающих связывает пептидогликан с высоким сродством, экспрессируется в нейтрофилах и ингибирует рост бактерий» . Журнал биологической химии . 275 (32): 24490–24499. дои : 10.1074/jbc.M001239200 . ПМИД 10827080 . S2CID 24226481 .
- ^ Перейти обратно: а б с д Дзиарский Р., Платт К.А., Гелиус Э., Штайнер Х., Гупта Д. (июль 2003 г.). «Нарушение уничтожения нейтрофилов и повышенная восприимчивость к инфекции непатогенными грамположительными бактериями у мышей с дефицитом белка-S распознавания пептидогликана (PGRP-S)» . Кровь . 102 (2): 689–697. дои : 10.1182/кровь-2002-12-3853 . ПМИД 12649138 .
- ^ Перейти обратно: а б с д и ж Чо Дж.Х., Фрейзер И.П., Фукасе К., Кусумото С., Фудзимото Ю., Шталь Г.Л., Иезековиц Р.А. (октябрь 2005 г.). «Белок S распознавания пептидогликана человека является эффектором врожденного иммунитета, опосредованного нейтрофилами» . Кровь . 106 (7): 2551–2558. дои : 10.1182/кровь-2005-02-0530 . ПМЦ 1895263 . ПМИД 15956276 .
- ^ Перейти обратно: а б с д и ж Пак С.Ю., Цзин Икс, Гупта Д., Дзиарски Р. (апрель 2013 г.). «Белок 1, распознающий пептидогликан, усиливает экспериментальную астму, стимулируя Th2 и Th17 и ограничивая ответы регуляторных Т-клеток и плазмоцитоидных дендритных клеток» . Журнал иммунологии . 190 (7): 3480–3492. doi : 10.4049/jimmunol.1202675 . ПМЦ 3608703 . ПМИД 23420883 .
- ^ Перейти обратно: а б с д Яо X, Гао М, Дай С, Мейер К.С., Чен Дж., Киран К.Дж. и др. (декабрь 2013 г.). «Белок 1, распознающий пептидогликан, способствует воспалению дыхательных путей у мышей, вызванному клещами домашней пыли» . Американский журнал респираторной клеточной и молекулярной биологии . 49 (6): 902–911. дои : 10.1165/rcmb.2013-0001OC . ПМЦ 3931111 . ПМИД 23808363 .
- ^ Перейти обратно: а б Каппелер С.Р., Хойбергер С., Фара З., Пухан З. (август 2004 г.). «Экспрессия белка распознавания пептидогликана, PGRP, в лактирующей молочной железе» . Журнал молочной науки . 87 (8): 2660–2668. doi : 10.3168/jds.S0022-0302(04)73392-5 . ПМИД 15328291 .
- ^ Перейти обратно: а б с Гош А., Ли С., Дзиарски Р., Чакраварти С. (сентябрь 2009 г.). «Новый антимикробный белок, распознающий пептидогликан в роговице» . Исследовательская офтальмология и визуальные науки . 50 (9): 4185–4191. дои : 10.1167/iovs.08-3040 . ПМК 3052780 . ПМИД 19387073 .
- ^ Перейти обратно: а б с Пак С.Ю., Гупта Д., Хурвич Р., Ким Ч.Х., Дзиарски Р. (декабрь 2011 г.). «Белок распознавания пептидогликана Pglyrp2 защищает мышей от псориазоподобного воспаления кожи, стимулируя регуляторные Т-клетки и ограничивая ответы Th17» . Журнал иммунологии . 187 (11): 5813–5823. doi : 10.4049/jimmunol.1101068 . ПМК 3221838 . ПМИД 22048773 .
- ^ Перейти обратно: а б с д Пак С.Ю., Гупта Д., Ким Ч., Дзиарски Р. (2011). «Дифференциальное влияние белков распознавания пептидогликана на экспериментальный атопический и контактный дерматит, опосредованный клетками Treg и Th17» . ПЛОС ОДИН . 6 (9): e24961. Бибкод : 2011PLoSO...624961P . дои : 10.1371/journal.pone.0024961 . ПМК 3174980 . ПМИД 21949809 .
- ^ Перейти обратно: а б с Саха С., Цзин Х, Пак С.Ю., Ван С., Ли Х., Гупта Д., Дзиарски Р. (август 2010 г.). «Белки, распознающие пептидогликаны, защищают мышей от экспериментального колита, способствуя нормальной флоре кишечника и предотвращая индукцию гамма-интерферона» . Клетка-хозяин и микроб . 8 (2): 147–162. дои : 10.1016/j.chom.2010.07.005 . ПМЦ 2998413 . ПМИД 20709292 .
- ^ Ло Д., Тайнан В., Дикерсон Дж., Менди Дж., Чанг Х.В., Шарф М. и др. (июль 2003 г.). «Экспрессия белка распознавания пептидогликана в эпителии, связанном с фолликулами Пейеровых пятен мыши, предполагает функциональную специализацию». Клеточная иммунология . 224 (1): 8–16. дои : 10.1016/s0008-8749(03)00155-2 . ПМИД 14572796 .
- ^ Ван Дж., Густи В., Сарасвати А., Ло Д.Д. (ноябрь 2011 г.). «Конвергентное и дивергентное развитие линий М-клеток в эпителии слизистой оболочки мыши» . Журнал иммунологии . 187 (10): 5277–5285. doi : 10.4049/jimmunol.1102077 . ПМК 3208058 . ПМИД 21984701 .
- ^ Даунс-Каннер С., Берки С., Дельгофф Г.М., Эдвардс Р.П., Куриэль Т., Одунси К. и др. (март 2017 г.). «Подавляющий Ил-17А + Фоксp3 + и бывший Th17 Ил-17А отрицательный Фоксp3 + T- reg -клетки являются источником опухолеассоциированных T -reg -клеток» . Nature Communications . 8 : 14649. Bibcode : ...814649D . doi : 10.1038/ncomms14649 . PMC 5355894. . PMID 28290453 2017NatCo
- ^ Перейти обратно: а б с д и ж г Шнелл А., Хуанг Л., Риган Б.М., Сингх В., Фонфихт Д., Боллхаген А. и др. (октябрь 2023 г.). «Нацеливание на PGLYRP1 способствует противоопухолевому иммунитету и одновременно подавляет аутоиммунное нейровоспаление» . Природная иммунология . 24 (11): 1908–1920. дои : 10.1038/s41590-023-01645-4 . ПМЦ 10864036 . ПМИД 37828379 . S2CID 263963953 .
- ^ Арентсен Т., Цянь Ю., Гкоцис С., Фемения Т., Ван Т., Удекву К. и др. (февраль 2017 г.). «Бактериальная молекула Pglyrp2, чувствительная к пептидогликанам, модулирует развитие и поведение мозга» . Молекулярная психиатрия . 22 (2): 257–266. дои : 10.1038/mp.2016.182 . ПМЦ 5285465 . ПМИД 27843150 .
- ^ Перейти обратно: а б Рехман А., Тайши П., Фанг Дж., Мажде Дж.А., Крюгер Дж.М. (январь 2001 г.). «Клонирование крысиного белка распознавания пептидогликана (PGRP) и его индукция в мозге при лишении сна». Цитокин . 13 (1): 8–17. дои : 10.1006/cyto.2000.0800 . ПМИД 11145837 .
- ^ Ланг М.Ф., Шнайдер А., Крюгер С., Шмид Р., Дзиарски Р., Шванингер М. (январь 2008 г.). «Белок-S распознавания пептидогликана (PGRP-S) активируется NF-kappaB». Письма по неврологии . 430 (2): 138–141. дои : 10.1016/j.neulet.2007.10.027 . ПМИД 18035491 . S2CID 54406942 .
- ^ Перейти обратно: а б Тайделл CC, Юань Дж., Тран П., Селстед М.Э. (январь 2006 г.). «Бычий белок-S, распознающий пептидогликан: антимикробная активность, локализация, секреция и связывающие свойства» . Журнал иммунологии . 176 (2): 1154–1162. дои : 10.4049/jimmunol.176.2.1154 . ПМИД 16394004 . S2CID 11173657 .
- ^ Перейти обратно: а б Де Марци MC, Тодоне М., Ганем М.Б., Ван К., Мариуцца Р.А., Фернандес М.М., Мальчиоди Э.Л. (июль 2015 г.). «Комплексы белок-пептидогликан, распознающие пептидогликан, увеличивают активацию моноцитов/макрофагов и усиливают воспалительную реакцию» . Иммунология . 145 (3): 429–442. дои : 10.1111/imm.12460 . ПМЦ 4479541 . ПМИД 25752767 .
- ^ Ван З.М., Ли Х., Коклин Р.Р., Ван М., Ван М., Фукасе К. и др. (декабрь 2003 г.). «Белок L-распознавания пептидогликана человека представляет собой N-ацетилмурамоил-L-аланинамидазу» . Журнал биологической химии . 278 (49): 49044–49052. дои : 10.1074/jbc.M307758200 . ПМИД 14506276 . S2CID 35373818 .
- ^ Перейти обратно: а б с д Лу X, Ван М, Ци Дж, Ван Х, Ли X, Гупта Д, Дзиарски Р (март 2006 г.). «Белки, распознающие пептидогликаны, представляют собой новый класс бактерицидных белков человека» . Журнал биологической химии . 281 (9): 5895–5907. дои : 10.1074/jbc.M511631200 . ПМИД 16354652 . S2CID 21943426 .
- ^ Перейти обратно: а б Шарма П., Сингх Н., Синха М., Шарма С., Пербандт М., Бетцель С. и др. (май 2008 г.). «Кристаллическая структура белка распознавания пептидогликана при разрешении 1,8 А демонстрирует двойную стратегию борьбы с инфекцией с помощью двух независимых функциональных гомодимеров». Журнал молекулярной биологии . 378 (4): 923–932. дои : 10.1016/j.jmb.2008.03.018 . ПМИД 18395744 .
- ^ Перейти обратно: а б с д Ван М., Лю Л.Х., Ван С., Ли X, Лу X, Гупта Д., Дзиарски Р. (март 2007 г.). «Белкам, распознающим пептидогликаны человека, требуется цинк для уничтожения как грамположительных, так и грамотрицательных бактерий, и они синергичны с антибактериальными пептидами» . Журнал иммунологии . 178 (5): 3116–3125. дои : 10.4049/jimmunol.178.5.3116 . ПМИД 17312159 . S2CID 22160694 .
- ^ «Реактом | PGLYRP1 связывает бактериальный пептидогликан» . http://reactome.org . Проверено 3 ноября 2020 г.
- ^ Перейти обратно: а б с д Кумар С., Ройчоудхури А., Эмбер Б., Ван К., Гуан Р., Мариуцца Р.А., Бунс Г.Дж. (ноябрь 2005 г.). «Селективное распознавание синтетических фрагментов пептидогликана типа лизина и мезодиаминопимелиновой кислоты белками распознавания пептидогликана человека I {альфа} и S» . Журнал биологической химии . 280 (44): 37005–37012. дои : 10.1074/jbc.M506385200 . ПМИД 16129677 . S2CID 44913130 .
- ^ Перейти обратно: а б с д и ж Гупта А., Арора Г., Розен С.Э., Клоос З., Цао Ю., Черни Дж. и др. (ноябрь 2020 г.). Коберн Дж. (ред.). «Скрининг библиотеки секретомов человека выявил роль белка распознавания пептидогликана 1 при Лайм-боррелиозе» . ПЛОС Патогены . 16 (11): e1009030. дои : 10.1371/journal.ppat.1009030 . ПМЦ 7657531 . ПМИД 33175909 .
- ^ Шарма П., Дубе Д., Синха М., Мишра Б., Дей С., Мал Г. и др. (сентябрь 2011 г.). «Мультилигандная специфичность сайта связывания молекулярного паттерна, связанного с патогеном, в белке распознавания пептидогликана» . Журнал биологической химии . 286 (36): 31723–31730. дои : 10.1074/jbc.M111.264374 . ПМК 3173064 . ПМИД 21784863 .
- ^ Перейти обратно: а б Шарма П., Дубе Д., Сингх А., Мишра Б., Сингх Н., Синха М. и др. (май 2011 г.). «Структурные основы распознавания патоген-ассоциированных молекулярных структур и ингибирования провоспалительных цитокинов верблюжьим белком распознавания пептидогликана» . Журнал биологической химии . 286 (18): 16208–16217. дои : 10.1074/jbc.M111.228163 . ПМК 3091228 . ПМИД 21454594 .
- ^ Перейти обратно: а б с д и ж г час Кашьяп Д.Р., Ван М., Лю Л.Х., Бунс Г.Дж., Гупта Д., Дзиарски Р. (июнь 2011 г.). «Белки, распознающие пептидогликаны, убивают бактерии, активируя белок-чувствительные двухкомпонентные системы» . Природная медицина . 17 (6): 676–683. дои : 10.1038/нм.2357 . ПМК 3176504 . ПМИД 21602801 .
- ^ Слонова Д., Посвятенко А., Кибардин А., Сысолятина Е., Лыссюк Е., Ермолаева С. и др. (23 декабря 2020 г.). «Человеческий белок распознавания короткого пептидогликана PGLYRP1 / Tag-7 / PGRP-S ингибирует внутриклеточное выживание Listeria monocytogenes в макрофагах» . Границы клеточной и инфекционной микробиологии . 10 : 582803. дои : 10.3389/fcimb.2020.582803 . ПМЦ 7785527 . ПМИД 33425777 .
- ^ Бобровский П., Манувера В., Полина Н., Подгорный О., Прусаков К., Говорун В., Лазарев В. (июль 2016 г.). «Рекомбинантные белки распознавания пептидогликана человека проявляют антихламидийную активность» . Инфекция и иммунитет . 84 (7): 2124–2130. дои : 10.1128/IAI.01495-15 . ПМЦ 4936355 . ПМИД 27160295 .
- ^ Ван Дж., Фэн Ю., Ван С., Сринивас С., Чен С., Ляо Х. и др. (июль 2017 г.). «Штаммы патогенных стрептококков используют новую стратегию побега для подавления бактериостатического эффекта, опосредованного белком распознавания пептидогликана млекопитающих» . Клеточная микробиология . 19 (7): e12724. дои : 10.1111/cmi.12724 . ПМИД 28092693 . S2CID 3534029 .
- ^ Осанаи А., Сашинами Х., Асано К., Ли С.Дж., Ху Д.Л., Накане А. (февраль 2011 г.). «Белок распознавания пептидогликана мыши PGLYRP-1 играет роль во врожденном иммунном ответе хозяина против инфекции Listeria monocytogenes» . Инфекция и иммунитет . 79 (2): 858–866. дои : 10.1128/IAI.00466-10 . ПМК 3028829 . ПМИД 21134971 .
- ^ Перейти обратно: а б Дзиарски Р., Пак С.Ю., Кашьяп Д.Р., Дауд С.Е., Гупта Д. (2016). «Pglyrp-регулируемая микрофлора кишечника Prevotella falsenii, Parabacteroides distasonis и Bacteroides eggerthii усиливается, а Alistipes Finegoldii ослабляет колит у мышей» . ПЛОС ОДИН 11 (1): e0146162. Бибкод : 2016PLoSO..1146162D . дои : 10.1371/journal.pone.0146162 . ПМК 4699708 . ПМИД 26727498 .
- ^ Перейти обратно: а б Банскар С., Децнер А.А., Хуарес-Родригес М.Д., Хозо И., Гупта Д., Дзиарски Р. (декабрь 2019 г.). « Микробиом, регулируемый Pglyrp1, усиливает экспериментальную аллергическую астму» . Журнал иммунологии . 203 (12): 3113–3125. doi : 10.4049/jimmunol.1900711 . ПМИД 31704882 . S2CID 207942798 .
- ^ Перейти обратно: а б с Саха С., Ци Дж., Ван С., Ван М., Ли Х, Ким Ю.Г. и др. (февраль 2009 г.). «PGLYRP-2 и Nod2 необходимы при артрите, вызванном пептидогликанами, и местном воспалении» . Клетка-хозяин и микроб . 5 (2): 137–150. дои : 10.1016/j.chom.2008.12.010 . ПМК 2671207 . ПМИД 19218085 .
- ^ Перейти обратно: а б Цзя Ю, Рен С., Сонг Л., Ван С., Хан В., Ли Дж. и др. (май 2023 г.). «PGLYRP1-mIgG2a-Fc ингибирует активацию макрофагов посредством передачи сигналов AKT/NF-κB и защищает от смертельного повреждения легких во время бактериальной инфекции» . iScience . 26 (5): 106653. Бибкод : 2023iSci...26j6653J . дои : 10.1016/j.isci.2023.106653 . ПМЦ 10102533 . ПМИД 37113764 .
- ^ Перейти обратно: а б с Сащенко Л.П., Духанина Е.А., Яшин Д.В., Шаталов Ю.В., Романова Е.А., Коробко Е.В. и др. (январь 2004 г.). «Белок распознавания пептидогликана tag7 образует цитотоксический комплекс с белком теплового шока 70 в растворе и в лимфоцитах» . Журнал биологической химии . 279 (3): 2117–2124. дои : 10.1074/jbc.M307513200 . ПМИД 14585845 . S2CID 23485070 .
- ^ Перейти обратно: а б с Яшин Д.В., Иванова О.К., Сошникова Н.В., Шелудченков А.А., Романова Е.А., Духанина Е.А. и др. (август 2015 г.). «Tag7 (PGLYRP1) в комплексе с Hsp70 индуцирует альтернативные цитотоксические процессы в опухолевых клетках через рецептор TNFR1» . Журнал биологической химии . 290 (35): 21724–21731. дои : 10.1074/jbc.M115.639732 . ПМЦ 4571894 . ПМИД 26183779 .
- ^ Сащенко Л.П., Духанина Е.А., Шаталов Ю.В., Яшин Д.В., Лукьянова Т.И., Кабанова О.Д. и др. (сентябрь 2007 г.). «Цитотоксические Т-лимфоциты, несущие белок распознавания образов Tag7, могут обнаруживать уклончивые, HLA-отрицательные, но экспонирующие Hsp70 опухолевые клетки, тем самым обеспечивая контактное уничтожение, опосредованное FasL/Fas». Кровь . 110 (6): 1997–2004. дои : 10.1182/blood-2006-12-064444 . ПМИД 17551095 . S2CID 14869208 .
- ^ Духанина Е.А., Кабанова О.Д., Лукьянова Т.И., Шаталов Ю.В., Яшин Д.В., Романова Е.А. и др. (август 2009 г.). «Противоположные роли метастазина (S100A4) в двух потенциально противоопухолевых механизмах с участием белка лимфоцитов человека Tag7 и Hsp70» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (33): 13963–13967. Бибкод : 2009PNAS..10613963D . дои : 10.1073/pnas.0900116106 . ПМК 2729003 . ПМИД 19666596 .
- ^ Яшин Д.В., Духанина Е.А., Кабанова О.Д., Романова Е.А., Лукьянова Т.И., Тоневицкий А.Г. и др. (март 2011 г.). «Белок, связывающий тепловой шок (HspBP1), защищает клетки от цитотоксического действия комплекса Tag7-Hsp70» . Журнал биологической химии . 286 (12): 10258–10264. дои : 10.1074/jbc.M110.163436 . ПМК 3060480 . ПМИД 21247889 .
- ^ Яшин Д.В., Романова Е.А., Иванова О.К., Сащенко Л.П. (апрель 2016). «Цитотоксический комплекс Tag7-Hsp70 индуцирует некроптоз опухолевых клеток посредством пермеабилизации лизосом и митохондрий». Биохимия . 123 : 32–36. дои : 10.1016/j.biochi.2016.01.007 . ПМИД 26796882 .
- ^ Романова Е.А., Шарапова Т.Н., Телегин Г.Б., Минаков АН, Чернов А.С., Иванова ОК и др. (февраль 2020 г.). «12-мерный пептид Tag7 (PGLYRP1) образует цитотоксический комплекс с Hsp70 и ингибирует гибель клеток, индуцированную TNF-альфа» . Клетки . 9 (2): 488. doi : 10.3390/cells9020488 . ПМК 7072780 . ПМИД 32093269 .
- ^ Перейти обратно: а б Шарапова Т.Н., Романова Е.А., Чернов А.С., Минаков АН, Казаков В.А., Кудряева А.А. и др. (октябрь 2021 г.). «Белковые пептиды PGLYRP1/Tag7 снижают провоспалительную реакцию в клетках крови человека и мышиную модель диффузного альвеолярного повреждения легких посредством блокады рецепторов TREM-1 и TNFR1» . Международный журнал молекулярных наук . 22 (20): 11213. doi : 10.3390/ijms222011213 . ПМЦ 8538247 . ПМИД 34681871 .
- ^ Телегин Г.Б., Чернов А.С., Казаков В.А., Романова Е.А., Шарапова Т.Н., Яшин Д.В. и др. (07.06.2021). «8-мерный пептид белка врожденного иммунитета PGLYRP1/Tag7 связывается с рецептором TNFR1 и ингибирует индуцированный TNFα цитотоксический эффект и воспаление» . Границы в иммунологии . 12 : 622471. дои : 10.3389/fimmu.2021.622471 . ПМЦ 8215708 . ПМИД 34163464 .
- ^ Прочтите CB, Kuijper JL, Hjorth SA, Heipel MD, Tang X, Fleetwood AJ и др. (февраль 2015 г.). «Передовой край: идентификация нейтрофила PGLYRP1 как лиганда TREM-1» . Журнал иммунологии . 194 (4): 1417–1421. doi : 10.4049/jimmunol.1402303 . ПМЦ 4319313 . ПМИД 25595774 .
- ^ Зульфикар Ф, Хозо И, Рангараджан С, Мариуцца Р.А., Дзиарски Р., Гупта Д. (2013). «Генетическая ассоциация вариантов белка распознавания пептидогликана с воспалительным заболеванием кишечника» . ПЛОС ОДИН 8 (6): e67393. Бибкод : 2013PLoSO...867393Z . дои : 10.1371/journal.pone.0067393 . ПМЦ 3686734 . ПМИД 23840689 .
- ^ Нкья С., Мвита Л., Мгая Дж., Кумбуру Х., ван Цветселар М., Мензель С. и др. (июнь 2020 г.). «Идентификация генетических вариантов и путей, связанных с экстремальными уровнями фетального гемоглобина при серповидно-клеточной анемии в Танзании» . BMC Медицинская генетика . 21 (1): 125. дои : 10.1186/s12881-020-01059-1 . ПМЦ 7275552 . ПМИД 32503527 .
- ^ Рохатги А., Айерс Ч.Р., Кера А., Макгуайр Д.К., Дас С.Р., Матулявичус С. и др. (апрель 2009 г.). «Связь между белком-1, распознающим пептидогликан, и коронарным и периферическим атеросклерозом: наблюдения из исследования сердца в Далласе». Атеросклероз . 203 (2): 569–575. doi : 10.1016/j.atherosclerosis.2008.07.015 . ПМИД 18774573 .
- ^ Браунелл Н.К., Кера А., де Лемос Дж.А., Айерс Ч.Р., Рохатги А. (май 2016 г.). «Связь между белком-1, распознающим пептидогликан, и возникновением атеросклеротических сердечно-сосудистых заболеваний: исследование сердца в Далласе» . Журнал Американского колледжа кардиологов . 67 (19): 2310–2312. дои : 10.1016/j.jacc.2016.02.063 . ПМИД 27173041 .
- ^ Пак Х.Дж., Но Дж.Х., Ын Дж.В., Ко Ю.С., Со СМ, Пак В.С. и др. (май 2015 г.). «Оценка и диагностическая значимость новых сывороточных биомаркеров для раннего принятия решения об инфаркте миокарда с подъемом сегмента ST» . Онкотаргет . 6 (15): 12970–12983. дои : 10.18632/oncotarget.4001 . ПМЦ 4536992 . ПМИД 26025919 .
- ^ Ратнаяке Н., Густафссон А., Сорса Т., Норхаммар А., Бостанчи Н. (сентябрь 2022 г.). «Связь белка распознавания пептидогликана 1 с постинфарктным инфарктом миокарда и воспалением пародонта: отчет подгруппы исследования PAROKRANK (заболевания пародонта и связь с инфарктом миокарда)» . Журнал пародонтологии . 93 (9): 1325–1335. дои : 10.1002/JPER.21-0595 . ПМЦ 9796725 . ПМИД 35344208 .
- ^ Цзинь Ю, Хуан Х, Шу Х, Лю З, Лу Л, Дай Ю, У Цзы (октябрь 2021 г.). «Белок 1, распознающий пептидогликан, ослабляет атеросклероз путем подавления адгезии эндотелиальных клеток». Журнал сердечно-сосудистой фармакологии . 78 (4): 615–621. doi : 10.1097/FJC.0000000000001100 . ПМИД 34269701 . S2CID 235962339 .
- ^ Перейти обратно: а б Хан Ю, Хуа С, Чен Ю, Ян В, Чжао В, Хуан Ф и др. (май 2021 г.). «Циркулирующие уровни PGLYRP1 как потенциальный биомаркер ишемической болезни сердца и сердечной недостаточности». Журнал сердечно-сосудистой фармакологии . 77 (5): 578–585. doi : 10.1097/FJC.0000000000000996 . ПМИД 33760799 . S2CID 232356516 .
- ^ Чжан Дж, Ченг Ю, Дуань М, Ци Н, Лю Дж (май 2017 г.). «Выявление дифференциально экспрессируемых генов при регуляции факторов транскрипции при сепсисе» . 3 Биотехнологии . 7 (1): 46. дои : 10.1007/s13205-017-0713-x . ПМК 5428098 . ПМИД 28444588 .
- ^ Бриньольфссон С.Ф., Магнуссон М.К., Конг П.Л., Йенсен Т., Куйпер Дж.Л., Хоканссон К. и др. (август 2016 г.). «Антитело против триггерного рецептора, экспрессируемое на миелоидных клетках 1 (TREM-1), подавляет секрецию провоспалительных цитокинов клетками собственной пластинки у пациентов с ВЗК» . Воспалительные заболевания кишечника . 22 (8): 1803–1811. doi : 10.1097/MIB.0000000000000822 . ПМИД 27243593 . S2CID 3637291 .
- ^ Соомро С., Венкатешваран С., Ванарса К., Харбутли М., Нидхи М., Сусарла Р. и др. (июнь 2021 г.). «Прогнозирование течения заболевания при язвенном колите с использованием белков стула, выявленных с помощью скрининга на основе аптамеров» . Природные коммуникации . 12 (1): 3989. Бибкод : 2021NatCo..12.3989S . дои : 10.1038/s41467-021-24235-0 . ПМК 8239008 . ПМИД 34183667 .
- ^ Молино П.Л., Уиллис-Оуэн С.А., Кокс М.Дж., Джеймс П., Коуман С., Лебингер М. и др. (июнь 2017 г.). «Взаимодействие хозяина и микробов при идиопатическом легочном фиброзе» . Американский журнал респираторной медицины и интенсивной терапии . 195 (12): 1640–1650. дои : 10.1164/rccm.201607-1408OC . ПМЦ 5476909 . ПМИД 28085486 .
- ^ Касаян М.Т., Ли Дж., Бреннан А., Данто С.И., Блэк К.Е., Фитц Л., Диксон А.Е. (июль 2018 г.). «Протеомный анализ анализируемых веществ сыворотки и мокроты различает контролируемых и плохо контролируемых астматиков». Клиническая и экспериментальная аллергия . 48 (7): 814–824. дои : 10.1111/cea.13151 . ПМИД 29665127 . S2CID 4938216 .
- ^ Нюлунд К.М., Руоконен Х., Сорса Т., Хейккинен А.М., Меурман Дж.Х., Ортис Ф. и др. (январь 2018 г.). «Связь рецептора, запускающего слюну, экспрессируемого на миелоидных клетках / оси белка 1 узнавания лиганда пептидогликана, с воспалением полости рта при заболевании почек». Журнал пародонтологии . 89 (1): 117–129. дои : 10.1902/jop.2017.170218 . ПМИД 28846062 . S2CID 21830535 .
- ^ Луо Ц, Ли Х, Чжан Л, Яо Ф, Дэн З, Цин С и др. (январь 2019 г.). «Сывороточный PGLYRP‑1 является высокодискриминационным биомаркером для диагностики ревматоидного артрита» . Отчеты о молекулярной медицине . 19 (1): 589–594. дои : 10.3892/ммр.2018.9632 . ПМИД 30431075 .
- ^ Силберайсен А., Халлак А.К., Насименто Г.Г., Сорса Т., Белибасакис Г.Н., Лопес Р., Бостанчи Н. (октябрь 2019 г.). «Регуляция PGLYRP1 и TREM-1 во время прогрессирования и разрешения воспаления десен». JDR Клинические и трансляционные исследования . 4 (4): 352–359. дои : 10.1177/2380084419844937 . ПМИД 31013451 . S2CID 129941967 .
- ^ Райвисто Т., Хейккинен А.М., Силберайзен А., Кованен Л., Руоконен Х., Тервахартиала Т. и др. (октябрь 2020 г.). «Регуляция белка 1 распознавания пептидогликана слюны у подростков». JDR Клинические и трансляционные исследования . 5 (4): 332–341. дои : 10.1177/2380084419894287 . ПМИД 31860804 . S2CID 209434091 .
- ^ Юсель З.П., Силберайзен А., Эмингил Г., Токгоз Ю., Козе Т., Сорса Т. и др. (октябрь 2020 г.). «Биомаркеры слюны в контексте воспаления десен у детей с муковисцидозом». Журнал пародонтологии . 91 (10): 1339–1347. дои : 10.1002/JPER.19-0415 . hdl : 10138/327022 . ПМИД 32100289 . S2CID 211523360 .
- ^ Карсияка Хендек М., Киса У., Ольгун Э. (январь 2020 г.). «Влияние курения на уровень белка-1 распознавания пептидогликана десневой жидкости после начальной пародонтальной терапии при хроническом пародонтите». Заболевания полости рта . 26 (1): 166–172. дои : 10.1111/odi.13207 . ПМИД 31587460 . S2CID 203850763 .
- ^ Тейшейра М.К., Лира-Жуниор Р., Лоуренсу Э.Дж., Теллес Д.М., Бострём Э.А., Фигередо К.М., Бостанчи Н. (май 2020 г.). «Модуляция оси TREM-1/PGLYRP1/MMP-8 при периимплантатных заболеваниях» . Клинические оральные исследования . 24 (5): 1837–1844. дои : 10.1007/s00784-019-03047-z . ПМИД 31444693 . S2CID 201283050 .
- ^ Инанк Н., Мумку Г., Джан М., Яй М., Зильберайзен А., Манойл Д. и др. (февраль 2021 г.). «Повышение уровня TREM-1 в сыворотке связано с пародонтитом и активностью заболевания при ревматоидном артрите» . Научные отчеты . 11 (1): 2888. Бибкод : 2021NatSR..11.2888I . дои : 10.1038/s41598-021-82335-9 . ПМЦ 7859204 . ПМИД 33536478 .
- ^ Зильберайзен А., Лира-Джуниор Р., Окерман С., Клинге Б., Бострём Э.А., Бостанчи Н. (ноябрь 2023 г.). «Ассоциация воспалительных маркеров TREM-1 и PGLYRP1 слюны с неинфекционными заболеваниями» . Журнал клинической пародонтологии . 50 (11): 1467–1475. дои : 10.1111/jcpe.13858 . ПМИД 37524498 . S2CID 260349050 .
- ^ Ян З, Ни Дж, Куанг Л, Гао Й, Тао С (сентябрь 2020 г.). «Идентификация генов и путей, связанных с субхондральной костью при остеоартрите, посредством биоинформатического анализа» . Лекарство . 99 (37):e2 дои : 10.1097/MD.0000000000022142 . ПМЦ 7489699 . ПМИД 32925767 .
- ^ Ортис Ф., Нюлунд К.М., Руоконен Х., Меурман Дж. Х., Фурухольм Дж., Бостанчи Н., Сорса Т. (декабрь 2020 г.). «Слюнные биомаркеры воспаления полости рта связаны с сердечно-сосудистыми заболеваниями и смертью среди пациентов, перенесших трансплантацию почки». Процедура трансплантации . 52 (10): 3231–3235. doi : 10.1016/j.transproceed.2020.07.007 . ПМИД 32768288 . S2CID 225451024 .
- ^ Гликман Дж.В., Дубин С., Ренерт-Юваль Ю., Дахабре Д., Киммел Г.В., Ауеунг К. и др. (февраль 2021 г.). «Поперечное исследование биомаркеров крови пациентов с очаговой алопецией от умеренной до тяжелой степени выявляет системную дисрегуляцию иммунных и сердечно-сосудистых биомаркеров». Журнал Американской академии дерматологии . 84 (2): 370–380. дои : 10.1016/j.jaad.2020.04.138 . ПМИД 32376430 . S2CID 218532915 .
- ^ Климчак-Томаниак Д., Боувенс Э., Шуурман А.С., Аккерхейс К.М., Константинеску А., Бругтс Дж. и др. (июнь 2020 г.). «Временные закономерности маркеров, связанных с макрофагами и нейтрофилами, связаны с клиническим исходом у пациентов с сердечной недостаточностью» . ЭСК Сердечная недостаточность . 7 (3): 1190–1200. дои : 10.1002/ehf2.12678 . ПМК 7261550 . ПМИД 32196993 .
- ^ Ян С., Цао С., Се Z, Чжоу Z (март 2020 г.). «Анализ потенциальных хаб-генов, участвующих в патогенезе китайских пациентов с диабетом 1 типа» . Анналы трансляционной медицины . 8 (6): 295. дои : 10.21037/атм.2020.02.171 . ПМЦ 7186604 . ПМИД 32355739 .
- ^ Арениус И., Руоконен Х., Ортис Ф., Фурухольм Дж., Валимаа Х., Бостанчи Н. и др. (июль 2020 г.). «Взаимосвязь между заболеваниями полости рта и инфекционными осложнениями у пациентов, находящихся на диализе». Заболевания полости рта . 26 (5): 1045–1052. дои : 10.1111/odi.13296 . hdl : 10138/325947 . ПМИД 32026534 . S2CID 211045697 .
- ^ Го Ц, Ли З (декабрь 2019 г.). «Биоинформатический анализ ключевых генов и путей, связанных с тромбозом при эссенциальной тромбоцитемии» . Монитор медицинских наук . 25 : 9262–9271. дои : 10.12659/MSM.918719 . ПМК 6911306 . ПМИД 31801935 .
- ^ Гранде Дж., Винченцони Ф., Миларди Д., Помпа Дж., Риккарди Д., Фрушелла Е. и др. (2017). «Протеом цервикальной слизи при эндометриозе» . Клиническая протеомика . 14 :7 дои : 10.1186/s12014-017-9142-4 . ПМК 5290661 . ПМИД 28174513 .
- ^ Туртурис Б.А., Теорелл Дж., Кениг М.Д., Тассинг-Хамфрис Л., Голд Д.Р., Литоньюа А.А. и др. (февраль 2021 г.). «Перинатальный гранулопоэз и риск детской астмы» . электронная жизнь . 10 . doi : 10.7554/eLife.63745 . ПМЦ 7889076 . ПМИД 33565964 .
Дальнейшее чтение
[ редактировать ]- Дзиарски Р., Ройет Дж., Гупта Д. (2016). «Белки, распознающие пептидогликаны, и лизоцим». В Рэтклиффе MJ (ред.). Энциклопедия иммунобиологии . Том. 2. Elsevier Ltd., стр. 389–403. дои : 10.1016/B978-0-12-374279-7.02022-1 . ISBN 978-0-12-374279-7 .
- Ройет Дж., Гупта Д., Дзиарски Р. (ноябрь 2011 г.). «Белки, распознающие пептидогликаны: модуляторы микробиома и воспаления». Обзоры природы. Иммунология . 11 (12): 837–851. дои : 10.1038/nri3089 . ПМИД 22076558 . S2CID 5266193 .
- Ройет Дж., Дзиарски Р. (апрель 2007 г.). «Белки-распознающие пептидогликаны: плейотропные сенсоры и эффекторы антимикробной защиты». Обзоры природы. Микробиология . 5 (4): 264–277. дои : 10.1038/nrmicro1620 . ПМИД 17363965 . S2CID 39569790 .
- Дзиарский Р., Гупта Д. (2006). «Белки, распознающие пептидогликаны (PGRP)» . Геномная биология . 7 (8): 232. doi : 10.1186/gb-2006-7-8-232 . ПМЦ 1779587 . ПМИД 16930467 .
- Бастос П.А., Уиллер Р., Бонека И.Г. (январь 2021 г.). «Поглощение, распознавание и реакция на пептидогликан у млекопитающего-хозяина» . Обзоры микробиологии FEMS . 45 (1). дои : 10.1093/femsre/fuaa044 . ПМК 7794044 . ПМИД 32897324 .
- Вольф Эй Джей, Андерхилл DM (апрель 2018 г.). «Распознавание пептидогликана врожденной иммунной системой». Обзоры природы. Иммунология . 18 (4): 243–254. дои : 10.1038/nri.2017.136 . ПМИД 29292393 . S2CID 3894187 .
- Ламан Дж.Д., 'т Харт Б.А., Пауэр С., Дзиарски Р. (июль 2020 г.). «Бактериальный пептидогликан как причина хронического воспаления головного мозга» (PDF) . Тенденции молекулярной медицины . 26 (7): 670–682. doi : 10.1016/j.molmed.2019.11.006 . ПМИД 32589935 . S2CID 211835568 .
- Гонсалес-Сантана А., Диас Хейц Р. (август 2020 г.). «Бактериальные пептидогликаны из микробиоты в развитии нервной системы и поведении» . Тенденции молекулярной медицины . 26 (8): 729–743. doi : 10.1016/j.molmed.2020.05.003 . ПМИД 32507655 .
галерея PDB |
|---|