Орбитофронтальная кора
| Орбитофронтальная кора | |
|---|---|
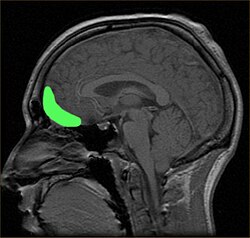 Примерное расположение OFC, показанное на сагиттальной МРТ. | |
 Орбитальная поверхность левой лобной доли. | |
| Подробности | |
| Часть | Лобная доля |
| Идентификаторы | |
| латинский | орбитофронтальная кора |
| Нейроимена | 91 |
| НейроЛекс ID | бирнлекс_1049 |
| ФМА | 242003 |
| Анатомические термины нейроанатомии | |
Орбитофронтальная кора ( ОФК ) — это область префронтальной коры в лобных долях головного мозга , которая участвует в когнитивном процессе принятия решений . У приматов, кроме человека, он состоит из ассоциативных областей коры Бродмана 11 , 12 и 13 ; у человека он состоит из зон Бродмана 10 , 11 и 47 . [1]
OFC функционально связан с вентромедиальной префронтальной корой . [2] Таким образом, этот регион отличается наличием различных нейронных связей и различными функциями, которые он выполняет. [3] Он определяется как часть префронтальной коры, которая получает проекции из дорсального ядра таламуса медиального и, как полагают, представляет эмоции , вкус, запах и вознаграждение при принятии решений. [4] [5] [6] [7] [8] [9] [10] [11] Свое название он получил из-за своего положения непосредственно над орбитами, в которых глаза расположены . В ОФК человека обнаружена значительная индивидуальная изменчивость. [12] Соответствующая область обнаружена у грызунов . [13]
Структура
[ редактировать ]OFC разделен на множество широких областей, отличающихся цитоархитектурой, включая область Бродмана 47/12 , область Бродмана 11 , область Бродмана 14 , область Бродмана 13 и область Бродмана 10 . [14] Четыре извилины разделены комплексом борозд, который чаще всего напоминает узор «H» или «K». Проходя вдоль ростро-каудальной оси, две борозды, латеральная и глазничная, обычно соединяются поперечной глазничной бороздой, которая проходит вдоль медиально-латеральной оси. Медиальнее медиальная глазничная извилина отделена от прямой извилины обонятельной бороздой. [15] Спереди и прямая извилина, и медиальная часть медиальной глазничной извилины состоят из области 11(m), сзади — из области 14. Задняя глазничная извилина состоит в основном из области 13 и граничит медиально и латерально с передними конечностями глазницы. медиальная и латеральная орбитальные борозды. Область 11 составляет большую часть ОФК, включающую как латеральные части медиальной глазничной извилины, так и переднюю глазничную извилину. Латеральная глазничная извилина состоит преимущественно из области 47/12. [14] Большая часть OFC зернистая , хотя каудальные части областей 13 и 14 агранулярны. [16] Эти каудальные области, которые иногда включают в себя части островковой коры , реагируют в первую очередь на необработанные сенсорные сигналы. [17]
Соединения
[ редактировать ]Связь OFC несколько варьируется вдоль рострально-каудальной оси. Каудальная OFC более тесно связана с сенсорными областями, в частности, получая прямую информацию от грушевидной коры . Каудальный OFC также наиболее тесно связан с миндалевидным телом . [18] Рострально OFC получает меньше прямых сенсорных проекций и меньше связан с миндалевидным телом, но взаимосвязан с латеральной префронтальной корой и парагиппокампом . [17] Соединение OFC также было задумано как состоящее из двух сетей; орбитальная сеть, состоящая из большей части центральных частей OFC, включая большую часть областей 47/12, 13 и 11; медиальная сеть, состоящая из медиальной самой и каудолатеральной областей OFC, а также областей 24 , 25 и 32 медиальной префронтальной коры. [19] Медиальную и орбитальную сети иногда называют «висцеромоторной сетью» и «сенсорной сетью» соответственно. [20]
Афференты
[ редактировать ]OFC получает проекции от нескольких сенсорных модальностей. Первичная обонятельная кора , вкусовая кора , вторичная соматосенсорная кора , верхняя и нижняя височная извилина (переносящая визуальную информацию) — все они проецируются на OFC. [16] [21] [22] Доказательства слуховых сигналов слабы, хотя некоторые нейроны реагируют на слуховые стимулы, что указывает на возможность существования косвенной проекции. [19] OFC также получает информацию от медиального дорсального ядра , островковой коры , энторинальной коры , периринальной коры , гипоталамуса и миндалевидного тела . [21] [23]
Эфференты
[ редактировать ]Орбитофронтальная кора взаимно связана с периринальной и энторинальной корой. [23] миндалевидное тело, гипоталамус и части медиальной височной доли. В дополнение к этим выводам OFC также проецируется на полосатое тело , включая прилежащее ядро , хвостатое ядро и вентральную скорлупу , а также на области среднего мозга, включая периакведуктальную серую часть и вентральную область покрышки . [21] [24] OFC воздействует на синапс миндалины по нескольким мишеням, включая два надежных пути к базолатеральной миндалине и вставочным клеткам миндалины , а также более слабую прямую проекцию на центральное ядро миндалины . [18]
Функция
[ редактировать ]OFC приписывают множество функций, включая посредничество, реагирование на конкретные ситуации, [25] гибкое кодирование непредвиденных обстоятельств, значение кодирования, кодирование предполагаемого значения, сдерживающие реакции , изменения в обучении в непредвиденных обстоятельствах, эмоциональная оценка, [26] изменение поведения с помощью соматических маркеров, управление социальным поведением и представление пространств состояний. [27] [28] Хотя большинство этих теорий объясняют определенные аспекты электрофизиологических наблюдений и изменения в поведении, связанные с поражением, они часто не объясняют или противоречат другим данным. Одно из предположений, объясняющих разнообразие функций OFC, заключается в том, что OFC кодирует пространства состояний или дискретную конфигурацию внутренних и внешних характеристик, связанных с ситуацией и ее непредвиденными обстоятельствами. [29] Например, предложение о том, что OFC кодирует экономическую ценность, может быть отражением значения состояния задачи кодирования OFC. [25] Представление состояний задач могло бы также объяснить предложение о том, что OFC действует как гибкая карта непредвиденных обстоятельств, поскольку переключение в состоянии задачи позволит кодировать новые непредвиденные обстоятельства в одном состоянии с сохранением старых непредвиденных обстоятельств в отдельном состоянии, что позволяет переключение непредвиденных обстоятельств, когда старое состояние задачи снова становится актуальным. [28] Представление состояний задачи подтверждается электрофизиологическими данными, демонстрирующими, что OFC реагирует на разнообразный набор функций задачи и способен быстро переназначать карту во время непредвиденных сдвигов. [28] Представление состояний задачи может влиять на поведение посредством множества потенциальных механизмов. Например, OFC необходим нейронам вентральной покрышки (VTA) для выработки ошибки прогнозирования дофаминергического вознаграждения, а OFC может кодировать ожидания для вычисления RPE в VTA. [25]
Конкретные функции были возложены на субрегионы OFC. Было предложено, чтобы латеральный OFC отражал потенциальную ценность выбора, позволяя фиктивным (контрфактическим) ошибкам прогнозирования потенциально опосредовать переключение выбора во время разворота, исчезновения и девальвации. [30] Оптогенетическая активация lOFC усиливает цель, направленную на привычное поведение, возможно, отражая повышенную чувствительность к потенциальному выбору и, следовательно, более частое переключение. С другой стороны, было предложено, чтобы mOFC отражал относительную субъективную ценность. [26] У грызунов аналогичная функция была приписана mOFC, кодируя ценность действия ступенчатым образом, в то время как lOFC было предложено кодировать специфические сенсорные характеристики результатов. [31] Также было предложено, чтобы lOFC кодировал ассоциации результатов стимула, которые затем сравнивались по значению в mOFC. [32] Метаанализ исследований нейровизуализации на людях показывает, что существует медиально-латеральный градиент валентности, при этом медиальный OFC чаще всего реагирует на вознаграждение, а латеральный OFC чаще всего реагирует на наказание. Также был обнаружен задне-передний градиент абстрактности: задний OFC реагирует на более простое вознаграждение, а передний OFC больше реагирует на абстрактное вознаграждение. [33] Аналогичные результаты были получены в ходе метаанализа исследований первичных и вторичных вознаграждений. [34]
OFC и базолатеральная миндалина (BLA) тесно взаимосвязаны, и их связь необходима для задач девальвации. Повреждение BLA или OFC раньше, но только OFC после девальвации ухудшает производительность. [35] В то время как BLA реагирует только на сигналы, предсказывающие значимые результаты в градационной форме в соответствии с ценностью, OFC реагирует как на ценность, так и на конкретные сенсорные атрибуты ассоциаций сигнал-результат. В то время как нейроны OFC, которые на ранних этапах обучения реагируют на получение результата, обычно передают свой ответ на появление сигналов, предсказывающих результат, повреждение BLA ухудшает эту форму обучения. [36]
Задняя орбитофронтальная кора (pOFC) связана с миндалевидным телом несколькими путями, которые способны как повышать, так и подавлять активность вегетативной нервной системы. [37] Предварительные данные свидетельствуют о том, что нейромодулятор дофамин играет роль в обеспечении баланса между тормозными и возбуждающими путями, при этом высокий уровень дофамина стимулирует вегетативную активность. [38]
Было высказано предположение, что медиальный OFC участвует в создании ассоциаций стимул-вознаграждение и в подкреплении поведения, тогда как латеральный OFC участвует в ассоциациях стимул-результат, а также в оценке и, возможно, изменении поведения. [39] Активность латерального OFC обнаруживается, например, когда субъекты кодируют новые ожидания наказания и социальных репрессий. [40] [41]
В исследованиях нейровизуализации было обнаружено, что средний передний OFC постоянно отслеживает субъективное удовольствие . В передней части OFC была обнаружена гедоническая горячая точка , которая способна усиливать реакцию симпатии к сахарозе. OFC также способен смещать аффективные реакции, вызванные антагонизмом α-амино-3-гидрокси-5-метил-4-изоксазолпропионовой кислоты (АМРА) в прилежащем ядре, в сторону аппетитных реакций. [42]
OFC способен модулировать агрессивное поведение посредством проекций на интернейроны миндалины, которые ингибируют глутаминергические проекции на вентромедиальный гипоталамус . [43]
Электрофизиология
[ редактировать ]Нейроны OFC реагируют как на первичные подкрепления, так и на сигналы, предсказывающие вознаграждение в нескольких сенсорных областях. Доказательства реакций на зрительные, вкусовые, соматосенсорные и обонятельные стимулы надежны, но доказательства слуховых реакций слабее. В подмножестве OFC-нейронов нейронные реакции на вознаграждение или сигналы вознаграждения модулируются индивидуальными предпочтениями и внутренними мотивационными состояниями, такими как голод. Часть нейронов, которые реагируют на сенсорные сигналы, предсказывающие вознаграждение, избирательны в отношении вознаграждения и демонстрируют обратное поведение, когда отношения результатов сигнала меняются местами. Нейроны OFC также реагируют на отсутствие ожидаемого вознаграждения и наказания. Другая популяция нейронов реагирует на новые стимулы и может «помнить» знакомые стимулы до суток. [44]
Во время задач вознаграждения по сигналу или инструментального вознаграждения по сигналу нейроны OFC демонстрируют три основных паттерна срабатывания; стрельба в ответ на сигналы; стрельба до получения награды; стрельба в ответ на получение вознаграждения. В отличие от медиальной префронтальной коры и полосатого тела , нейроны OFC не проявляют возбуждения, опосредованного движением. Однако их реакции, прогнозирующие вознаграждение, формируются вниманием: при переключении внимания между двумя альтернативами одна и та же популяция OFC будет положительно представлять ценность предмета, который в данный момент посещается, но отрицательно — ценность предмета, оставленного без внимания. [45] Кодирование величины вознаграждения также является гибким и учитывает относительную стоимость существующих вознаграждений. [46]
Люди
[ редактировать ]OFC человека является одной из наименее изученных областей человеческого мозга. Было высказано предположение, что OFC участвует в сенсорной интеграции, в представлении аффективной ценности подкреплений, а также в принятии решений и ожиданиях. [1] В частности, OFC, по-видимому, важен для сигнализации об ожидаемом вознаграждении/наказании за действие с учетом конкретных деталей ситуации. [47] При этом мозг способен сравнивать ожидаемое вознаграждение/наказание с фактическим получением вознаграждения/наказания, что делает OFC критически важным для адаптивного обучения. Это подтверждается исследованиями на людях, приматах и грызунах.
Психические расстройства
[ редактировать ]Орбитофронтальная кора вовлечена в пограничное расстройство личности . [48] шизофрения , большое депрессивное расстройство , биполярное расстройство , обсессивно-компульсивное расстройство , зависимость , посттравматическое стрессовое расстройство , аутизм , [49] и паническое расстройство . Хотя исследования нейровизуализации предоставили доказательства дисфункции при широком спектре психических расстройств, загадочная природа роли OFC в поведении усложняет понимание его роли в патофизиологии психических расстройств. [50] Функция OFC неизвестна, но его анатомические связи с вентральным полосатым телом, миндалевидным телом, гипоталамусом, гиппокампом и околоводопроводным серым цветом подтверждают роль в опосредовании поведения, связанного с вознаграждением и страхом. [51]
Обсессивно-компульсивное расстройство
[ редактировать ]Мета-анализ нейровизуализационных исследований при ОКР сообщает о гиперактивности в областях, которые обычно считаются частью орбитофронтального сегмента корково-базальных ганглиев-таламо-кортикальной петли, таких как хвостатое ядро , таламус и орбитофронтальная кора. Было предложено, чтобы ОКР отражало петлю положительной обратной связи из-за взаимного возбуждения OFC и подкорковых структур. [52] Хотя OFC обычно сверхактивен во время задач, провоцирующих симптомы, когнитивные задачи обычно вызывают гипоактивность OFC; [53] это может отражать различие между эмоциональными и неэмоциональными задачами, латеральной и медиальной OFC, [54] или просто противоречивые методологии. [55]
Зависимость
[ редактировать ]Модели на животных и специфические для клеток манипуляции в отношении поведения, связанного с поиском наркотиков, указывают на дисфункцию OFC при зависимости. [56] Расстройства, связанные с употреблением психоактивных веществ, связаны с различными нарушениями, связанными с гибким целенаправленным поведением и принятием решений. Эти дефициты совпадают с симптомами, связанными с поражениями OFC, а также связаны с уменьшением орбитофронтального серого вещества , гипометаболизмом в состоянии покоя и притуплением активности OFC во время задач, связанных с принятием решений или целенаправленным поведением. В отличие от состояния покоя и активности, связанной с принятием решений, сигналы, связанные с наркотиками, вызывают сильную активность OFC, которая коррелирует с тягой. [57] Исследования на грызунах также показывают, что проекции lOFC на BLA необходимы для восстановления самоуправления, индуцированного сигналом. Все эти результаты согласуются с ролью, которую OFC играет в кодировании результатов, связанных с определенными стимулами. [58] [59] [60] Прогресс в сторону компульсивного злоупотребления психоактивными веществами может отражать переход от принятия решений на основе модели, когда внутренняя модель будущих результатов определяет решения, к модели свободного обучения, где решения основаны на истории подкрепления. Обучение на основе моделей включает OFC и является гибким и целенаправленным, в то время как обучение без моделей более жесткое; поскольку переход к более свободному от модели поведению из-за дисфункции OFC, например, вызванной злоупотреблением наркотиками, может лежать в основе привычек, связанных с поиском наркотиков. [61]
Поведенческие расстройства
[ редактировать ]Расстройство поведения связано как со структурными отклонениями, так и с функциональными нарушениями при выполнении аффективных задач. [62] Аномалии в структуре, активности и функциональных связях OFC наблюдались в связи с агрессией. [63]
Аффективные расстройства
[ редактировать ]Нейровизуализационные исследования выявили нарушения OFC при БДР и биполярном расстройстве. В соответствии с градиентом медиальный/награда и латеральный/наказание, обнаруженным в исследованиях нейровизуализации, в некоторых исследованиях нейровизуализации наблюдалась повышенная активность латеральной OFC при депрессии, а также снижение взаимосвязанности медиальной OFC и усиление взаимосвязи в латеральной OFC. [64] Гипоактивность латеральной OFC часто наблюдалась при биполярном расстройстве, особенно во время маниакальных эпизодов. [64]
Исследовать
[ редактировать ]Визуализация
[ редактировать ]Использование функциональной магнитно-резонансной томографии (фМРТ) для визуализации ОФК человека является сложной задачей, поскольку эта область мозга находится в непосредственной близости от наполненных воздухом пазух . Это означает, что могут возникать артефактные ошибки при обработке сигнала , вызывающие, например, геометрические искажения, которые часто встречаются при использовании эхопланарной визуализации (EPI) при более высоких напряженностях магнитного поля. Поэтому рекомендуется проявлять особую осторожность для получения хорошего сигнала от орбитофронтальной коры, и был разработан ряд стратегий, таких как автоматическое шиммирование при высокой напряженности статического магнитного поля. [65]
Грызуны
[ редактировать ]У грызунов OFC полностью агранулярный или дисгранулярный. [16] OFC делится на вентролатеральную (VLO), латеральную (LO), медиальную (MO) и дорсолатеральную (DLO) области. [19] Используя весьма специфические методы манипулирования схемами, такие как оптогенетика , OFC был вовлечен в поведение, подобное ОКР. [66] и в способности использовать скрытые переменные в задаче принятия решений. [67]
Клиническое значение
[ редактировать ]Повреждать
[ редактировать ]Разрушение OFC в результате приобретенной травмы головного мозга обычно приводит к расторможенному поведению. Примеры включают чрезмерную ругань, гиперсексуальность, плохое социальное взаимодействие, пристрастие к азартным играм, употребление наркотиков (включая алкоголь и табак) и плохую способность к сопереживанию. Считается , что расторможенное поведение пациентов с некоторыми формами лобно-височной деменции вызвано дегенерацией ОФК. [68]
Нарушение
[ редактировать ]Когда OFC-связи нарушаются, может возникнуть ряд когнитивных, поведенческих и эмоциональных последствий. Исследования подтверждают, что основные расстройства, связанные с нарушением регуляции связей/схем OFC, сосредоточены вокруг принятия решений, регуляции эмоций, импульсивного контроля и ожидания вознаграждения. [69] [70] [71] [72] Недавнее мультимодальное исследование нейровизуализации человека показывает, что нарушение структурной и функциональной связи OFC с подкорковыми лимбическими структурами (например, миндалевидным телом или гиппокампом) и другими лобными областями (например, дорсальной префронтальной корой или передней поясной извилиной корой) коррелирует с аномальным эффектом OFC ( например, страх) у клинически тревожных взрослых. [73]
Одним из явных проявлений проблем с принятием решений является наркомания/ зависимость от психоактивных веществ , которые могут возникнуть в результате нарушения стриато-таламо-орбитофронтального контура. [72] [70] [74] Синдром дефицита внимания и гиперактивности (СДВГ) также связан с дисфункцией нейронных цепей вознаграждения, контролирующих мотивацию, вознаграждение и импульсивность, включая системы OFC. [71] другие нарушения исполнительного функционирования и контроля импульсов Дисрегуляция схемы OFC может влиять на , такие как обсессивно-компульсивное расстройство и трихотилломания. [75] [76] [77]
Некоторые виды деменции также связаны с нарушениями связи OFC. Поведенческий вариант лобно-височной деменции [78] связан с паттернами нейронной атрофии проекционных волокон белого и серого вещества, участвующих в соединении OFC. [79] Наконец, некоторые исследования показывают, что на более поздние стадии болезни Альцгеймера влияет изменение связности систем OFC. [77]
Орбитофронтальная эпилепсия
[ редактировать ]Орбитофронтальная эпилепсия встречается редко, но встречается. Проявления ОФК-эпилепсии довольно разнообразны, хотя общие характеристики включают симптомы, связанные со сном, автоматизмы и гипермоторные симптомы. В одном обзоре сообщалось, что ауры, как правило, не были распространены или неспецифичны, тогда как в другом сообщалось, что ОФК-эпилепсия ассоциировалась с аурами, включающими соматосенсорные явления и страх. [80] [81] [82]
Оценка
[ редактировать ]Тест на зрительную дискриминацию
[ редактировать ]Тест на зрительную дискриминацию состоит из двух компонентов. В первом компоненте, «обратное обучение», участникам предоставляется одна из двух картинок, A и B. Они узнают, что будут вознаграждены, если нажмут кнопку, когда отображается изображение A, но будут наказаны, если они нажмут кнопку, когда появится изображение A. Отображается буква Б. Как только это правило установлено, оно меняется. Другими словами, теперь правильно нажимать кнопку для изображения Б, а не для изображения А. Большинство здоровых участников практически сразу замечают изменение этого правила, но пациенты с повреждением OFC продолжают реагировать на первоначальный образец подкрепления, хотя они теперь его наказывают за настойчивость в этом. Роллс и др. [83] отметили, что такая модель поведения особенно необычна, учитывая, что пациенты сообщили, что поняли правило.
Второй компонент теста — «вымирание». И снова участники учатся нажимать кнопку для изображения А, но не для изображения Б. Однако на этот раз вместо изменения правил правило меняется вообще. Теперь участник будет наказан за нажатие кнопки в ответ на любую картинку. Правильный ответ — вообще не нажимать кнопку, но людям с дисфункцией OFC трудно устоять перед искушением нажать кнопку, несмотря на то, что их за это наказывают.
Задача по азартным играм в Айове
[ редактировать ]Имитация принятия решений в реальной жизни , игра в азартные игры в Айове широко используется в исследованиях познания и эмоций. [84] Участникам на экране компьютера представлены четыре виртуальные колоды карт. Им говорят, что каждый раз, когда они выбирают карту, они могут выиграть игровые деньги. Им говорят, что цель игры — выиграть как можно больше денег. Однако время от времени, выбирая карту, они теряют немного денег. Задача должна быть непрозрачной, то есть участники не должны сознательно вырабатывать правило, и они должны выбирать карты на основе своей « интуитивной реакции ». Две колоды являются «плохими», что означает, что в течение достаточно длительного времени они принесут чистый убыток; две другие колоды являются «хорошими колодами» и со временем принесут чистую прибыль.
Большинство здоровых участников пробуют карты из каждой колоды и после 40–50 выборов довольно хорошо ориентируются в хороших колодах. Однако пациенты с дисфункцией OFC продолжают упорно использовать плохие колоды, иногда даже несмотря на то, что в целом они теряют деньги. Параллельное измерение кожно-гальванической реакции показывает, что здоровые участники демонстрируют «стрессовую» реакцию на зависание над плохими колодами уже после 10 попыток, задолго до того, как они осознают, что колоды плохие. Напротив, у пациентов с дисфункцией OFC никогда не развивается эта физиологическая реакция на предстоящее наказание. Бечара и его коллеги объясняют это с точки зрения гипотезы соматических маркеров . Задача об азартных играх в Айове в настоящее время используется рядом исследовательских групп с помощью фМРТ для изучения того, какие области мозга активируются этой задачей у здоровых добровольцев, а также в клинических группах с такими заболеваниями, как шизофрения и обсессивно-компульсивное расстройство .
Тест на оплошность — это серия эпизодов, рассказывающих о социальном событии, во время которого кто-то сказал что-то, чего не следовало говорить, или о неловком происшествии. Задача участника — определить, что было сказано неловко, почему это было неловко, как люди почувствовали бы себя в ответ на оплошность и на контрольный вопрос. Хотя впервые он был разработан для использования у людей с аутистическим спектром , [85] тест также чувствителен к пациентам с дисфункцией OFC, которые не могут судить о том, что произошло что-то социально неловкое, несмотря на то, что они, кажется, прекрасно понимают историю.
См. также
[ редактировать ]Дополнительные изображения
[ редактировать ]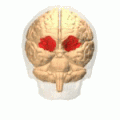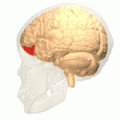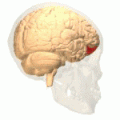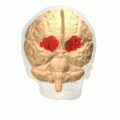 Орбитальная извилина показана красным.
Орбитальная извилина показана красным. Медиальная поверхность коры головного мозга – извилины.
Медиальная поверхность коры головного мозга – извилины. Базальная поверхность головного мозга. Орбитальная извилина показана красным.
Базальная поверхность головного мозга. Орбитальная извилина показана красным. Латеральная орбитофронтальная кора
Латеральная орбитофронтальная кора Медиальная орбитофронтальная кора, вид внутреннего среза.
Медиальная орбитофронтальная кора, вид внутреннего среза.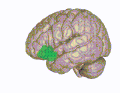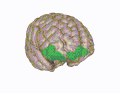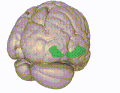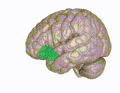 3D-визуализация орбитофронтальной коры головного мозга среднего человека
3D-визуализация орбитофронтальной коры головного мозга среднего человека Орбитофронтальная кора выделена зеленым на корональных изображениях Т1 МРТ.
Орбитофронтальная кора выделена зеленым на корональных изображениях Т1 МРТ. Орбитофронтальная кора выделена зеленым на сагиттальных изображениях МРТ Т1.
Орбитофронтальная кора выделена зеленым на сагиттальных изображениях МРТ Т1. Орбитофронтальная кора выделена зеленым на поперечных изображениях Т1 МРТ.
Орбитофронтальная кора выделена зеленым на поперечных изображениях Т1 МРТ.









Ссылки
[ редактировать ]- ^ Перейти обратно: а б Крингельбах М.Л. (2005). «Орбитофронтальная кора: связь вознаграждения с гедонистическим опытом». Обзоры природы Неврология . 6 (9): 691–702. дои : 10.1038/nrn1747 . ПМИД 16136173 . S2CID 205500365 .
- ^ Филлипс, Л.Х., Макферсон, ЮВ. И Делла Сала, С. (2002). «Возраст, познание и эмоции: роль анатомической сегрегации в лобных долях: роль анатомической сегрегации в лобных долях». в Дж. Графмане (ред.), Справочнике нейропсихологии: лобные доли. Elsevier Science, Амстердам, стр. 73–98.
- ^ Барбас Х., Гашгаи Х., Ремпель-Клоуэр Н., Сяо Д. (2002)Анатомические основы функциональной специализации префронтальной коры у приматов. В: Справочник по нейропсихологии (Графман Дж., изд.), стр. 1–27. Амстердам: Elsevier Science BV
- ^ Готфрид, Джей А.; Залд, Дэвид Х. (декабрь 2005 г.). «О запахе обонятельной орбитофронтальной коры головного мозга человека: метаанализ и сравнение с приматами» . Обзоры исследований мозга . 50 (2): 287–304. дои : 10.1016/j.brainresrev.2005.08.004 . ПМИД 16213593 .
- ^ Роллс, Эдмунд Т. (июнь 2004 г.). «Функции орбитофронтальной коры» . Мозг и познание . 55 (1): 11–29. дои : 10.1016/S0278-2626(03)00277-X . ПМИД 15134840 .
- ^ Крингельбах, Мортен Л. (сентябрь 2005 г.). «Орбитофронтальная кора человека: связь вознаграждения с гедонистическим опытом» . Обзоры природы Неврология . 6 (9): 691–702. дои : 10.1038/nrn1747 . ISSN 1471-003X . ПМИД 16136173 .
- ^ Рашворт, MFS; Беренс, TEJ; Рудебек, PH; Уолтон, Мэн (апрель 2007 г.). «Контрастные роли поясной извилины и орбитофронтальной коры в принятии решений и социальном поведении» . Тенденции в когнитивных науках . 11 (4): 168–176. дои : 10.1016/j.tics.2007.01.004 . ПМИД 17337237 .
- ^ Диксон, Мэтью Л.; Тиручсельвам, Рави; Тодд, Ребекка; Кристофф, Калина (октябрь 2017 г.). «Эмоции и префронтальная кора: интегративный обзор» . Психологический вестник . 143 (10): 1033–1081. дои : 10.1037/bul0000096 . ISSN 1939-1455 . ПМИД 28616997 .
- ^ Крингельбах, Мортен Л.; Роллс, Эдмунд Т. (апрель 2004 г.). «Функциональная нейроанатомия орбитофронтальной коры человека: данные нейровизуализации и нейропсихологии» . Прогресс нейробиологии . 72 (5): 341–372. doi : 10.1016/j.pneurobio.2004.03.006 . ISSN 0301-0082 . ПМИД 15157726 .
- ^ Дамасио, Антонио Р. (1994). Ошибка Декарта: эмоции, разум и человеческий мозг . Нью-Йорк: Патнэм. ISBN 978-0-399-13894-2 .
- ^ Фустер, Дж. М. Префронтальная кора головного мозга (Raven Press, Нью-Йорк, 1997).
- ^ Исама Н., Фэйсон В., Пейн М.Э., Макфолл Дж., Стеффенс Д.С., Бейер Дж.Л., Кришнан Р., Тейлор В.Д. (2010). «Изменчивость структуры лобно-височного мозга: важность привлечения афроамериканцев к нейробиологическим исследованиям» . ПЛОС ОДИН . 5 (10): е13642. Бибкод : 2010PLoSO...513642I . дои : 10.1371/journal.pone.0013642 . ПМЦ 2964318 . ПМИД 21049028 .
- ^ Уйлингс Х.Б., Гроеневеген Х.Дж., Колб Б. (2003). «Есть ли у крыс префронтальная кора?». Поведение мозга Res . 146 (1–2): 3–17. дои : 10.1016/j.bbr.2003.09.028 . ПМИД 14643455 . S2CID 32136463 .
- ^ Перейти обратно: а б Макки, Сотт; Петридес, Майкл (2006). «Глава 2: Орбитофронтальная кора: морфология и архитектура борозд и извилин». В Зальде, Дэвид Х.; Раух, Скотт (ред.). Орбитофронтальная кора . Нью-Йорк: Издательство Оксфордского университета. п. 34. ISBN 9780198565741 .
- ^ Макки, Сотт; Петридес, Майкл (2006). «Глава 2: Орбитофронтальная кора: морфология и архитектура борозд и извилин». В Зальде, Дэвид Х.; Раух, Скотт (ред.). Орбитофронтальная кора . Нью-Йорк: Издательство Оксфордского университета. п. 24. ISBN 9780198565741 .
- ^ Перейти обратно: а б с Пассингем, Ричард Э.; Уайз, Стивен П. (1012). «Глава 4. Орбитальная префронтальная кора: выбор объектов на основе результатов». Нейробиология префронтальной коры: анатомия, эволюция и происхождение понимания . Грейт-Кларендон-стрит, Оксфорд: Издательство Оксфордского университета. п. 97. ИСБН 9780199552917 .
- ^ Перейти обратно: а б Хабер, С.Н.; Беренс, TE (3 сентября 2014 г.). «Нейронная сеть, лежащая в основе обучения на основе стимулов: значение для интерпретации нарушений цепи при психических расстройствах» . Нейрон . 83 (5): 1019–39. дои : 10.1016/j.neuron.2014.08.031 . ПМК 4255982 . ПМИД 25189208 .
- ^ Перейти обратно: а б Барбас, Хелен; Зикопулос, Василис (2006). «Глава 4: Последовательные и параллельные схемы обработки эмоций в орбитофронтальной коре приматов». В Раухе, Скотт Л.; Зальд, Дэвид Х. (ред.). Орбитофронтальная кора . Нью-Йорк: Издательство Оксфордского университета. п. 67.
- ^ Перейти обратно: а б с Прайс, Джозеф Л. (2006). «Глава 3: Соединения орбитальной коры». В Раухе, Скотт Л.; Зальд, Дэвид Х. (ред.). Орбитофронтальная кора . Нью-Йорк: Издательство Оксфордского университета. п. 42.
- ^ Рудебек, PH; Мюррей, Е.А. (декабрь 2011 г.). «Балканизация орбитофронтальной коры приматов: отдельные субрегионы для сравнения и противопоставления значений» . Анналы Нью-Йоркской академии наук . 1239 (1): 1–13. Бибкод : 2011NYASA1239....1R . дои : 10.1111/j.1749-6632.2011.06267.x . ПМЦ 3951748 . ПМИД 22145870 .
- ^ Перейти обратно: а б с Роллс, ET (март 2000 г.). «Орбитофронтальная кора и вознаграждение» . Кора головного мозга . 10 (3): 284–94. дои : 10.1093/cercor/10.3.284 . ПМИД 10731223 .
- ^ Роллс, ET (ноябрь 2004 г.). «Конвергенция сенсорных систем орбитофронтальной коры у приматов и дизайн мозга для эмоций» . Анатомические записи, часть A: открытия в молекулярной, клеточной и эволюционной биологии . 281 (1): 1212–25. дои : 10.1002/ar.a.20126 . ПМИД 15470678 .
- ^ Перейти обратно: а б Ремпель-Клоуэр, Нидерланды (декабрь 2007 г.). «Роль связей орбитофронтальной коры в эмоциях». Анналы Нью-Йоркской академии наук . 1121 (1): 72–86. Бибкод : 2007NYASA1121...72R . дои : 10.1196/анналы.1401.026 . ПМИД 17846152 . S2CID 21317263 .
- ^ Прайс, Джозеф Л. (2006). «Глава 3: Соединения орбитальной коры». В Раухе, Скотт Л.; Зальд, Дэвид Х. (ред.). Орбитофронтальная кора . Нью-Йорк: Издательство Оксфордского университета. п. 45.
- ^ Перейти обратно: а б с Викенхайзер, AM; Шенбаум, Г. (август 2016 г.). «Над рекой, через лес: когнитивные карты в гиппокампе и орбитофронтальной коре» . Обзоры природы. Нейронаука . 17 (8): 513–23. дои : 10.1038/nrn.2016.56 . ПМЦ 5541258 . ПМИД 27256552 .
- ^ Перейти обратно: а б Феттс, П; Шульце, Л; Даунар, Дж (2017). «Кортико-стриатально-таламическая петля орбитофронтальной коры: многообещающие терапевтические цели при психических заболеваниях» . Границы системной нейронауки . 11:25 . дои : 10.3389/fnsys.2017.00025 . ПМК 5406748 . ПМИД 28496402 .
- ^ Уилсон, Роберт С.; Такахаси, Юджи К.; Шенбаум, Джеффри; Нив, Яэль (январь 2014 г.). «Орбитофронтальная кора как когнитивная карта пространства задач» . Нейрон . 81 (2): 267–279. дои : 10.1016/j.neuron.2013.11.005 . ISSN 0896-6273 . ПМК 4001869 . ПМИД 24462094 .
- ^ Перейти обратно: а б с Садакка, БФ; Викенхайзер, AM; Шенбаум, Г. (14 марта 2017 г.). «К теоретической роли тонического норадреналина в орбитофронтальной коре головного мозга в обеспечении гибкого обучения» . Нейронаука . 345 : 124–129. doi : 10.1016/j.neuroscience.2016.04.017 . ПМК 5461826 . ПМИД 27102419 .
- ^ Сталнакер, Т.А.; Куч, Северная Каролина; Шенбаум, Г. (май 2015 г.). «Чего не делает орбитофронтальная кора» . Природная неврология . 18 (5): 620–7. дои : 10.1038/nn.3982 . ПМЦ 5541252 . ПМИД 25919962 .
- ^ Тобиа, MJ; Го, Р.; Шварце, У.; Бемер, В.; Глешер, Дж.; Финк, Б.; Маршнер, А.; Бюхель, К.; Обермайер, К.; Соммер, Т. (1 апреля 2014 г.). «Нейронные системы выбора и оценки с контрфактическими обучающими сигналами». НейроИмидж . 89 : 57–69. doi : 10.1016/j.neuroimage.2013.11.051 . ISSN 1053-8119 . ПМИД 24321554 . S2CID 35280557 .
- ^ Искьердо, А. (1 ноября 2017 г.). «Функциональная неоднородность орбитофронтальной коры головного мозга крыс при обучении с вознаграждением и принятии решений» . Журнал неврологии . 37 (44): 10529–10540. doi : 10.1523/JNEUROSCI.1678-17.2017 . ПМК 6596524 . ПМИД 29093055 .
- ^ Рудебек, PH; Мюррей, Е.А. (декабрь 2011 г.). «Балканизация орбитофронтальной коры приматов: отдельные субрегионы для сравнения и противопоставления значений» . Анналы Нью-Йоркской академии наук . 1239 (1): 1–13. Бибкод : 2011NYASA1239....1R . дои : 10.1111/j.1749-6632.2011.06267.x . ПМЦ 3951748 . ПМИД 22145870 .
- ^ Крингельбах, МЛ; Роллс, ET (апрель 2004 г.). «Функциональная нейроанатомия орбитофронтальной коры человека: данные нейровизуализации и нейропсихологии». Прогресс нейробиологии . 72 (5): 341–72. doi : 10.1016/j.pneurobio.2004.03.006 . ПМИД 15157726 . S2CID 13624163 .
- ^ Сескусс, Г; Кальду, X; Сегура, Б; Дреер, Дж. К. (май 2013 г.). «Обработка первичных и вторичных вознаграждений: количественный метаанализ и обзор исследований функциональной нейровизуализации человека». Неврологические и биоповеденческие обзоры . 37 (4): 681–96. doi : 10.1016/j.neubiorev.2013.02.002 . hdl : 2066/117487 . ПМИД 23415703 . S2CID 8335094 .
- ^ Падоа-Скиоппа, К; Конен, Кентукки (15 ноября 2017 г.). «Орбитофронтальная кора: нейронная цепь принятия экономических решений» . Нейрон . 96 (4): 736–754. дои : 10.1016/j.neuron.2017.09.031 . ПМЦ 5726577 . ПМИД 29144973 .
- ^ Шарп, MJ; Шенбаум, Г. (май 2016 г.). «Назад к основам: предсказания в орбитофронтально-амигдальном контуре» . Нейробиология обучения и памяти . 131 : 201–6. дои : 10.1016/j.nlm.2016.04.009 . ПМК 5541254 . ПМИД 27112314 .
- ^ Барбас, Х. (август 2007 г.). «Поток информации об эмоциях через височные и орбитофронтальные пути» . Журнал анатомии . 211 (2): 237–49. дои : 10.1111/j.1469-7580.2007.00777.x . ПМЦ 2375774 . ПМИД 17635630 .
Задняя орбитофронтальная кора воздействует на двойные системы миндалевидного тела, которые оказывают противоположное воздействие на центральные вегетативные структуры. Оба пути берут начало в задней орбитофронтальной коре, но один из них в значительной степени нацелен на тормозные вставочные массы, активация которых может в конечном итоге растормозить центральные вегетативные структуры во время эмоционального возбуждения.
- ^ Зикопулос, Б; Хойстад, М; Джон, Ю; Барбас, Х. (17 мая 2017 г.). «Задние орбитофронтальные и передние поясные пути к целевой тормозной и возбуждающей системам миндалевидного тела с противоположными функциями» . Журнал неврологии . 37 (20): 5051–5064. doi : 10.1523/JNEUROSCI.3940-16.2017 . ПМЦ 5444191 . ПМИД 28411274 .
Обнаруженная здесь специфическая иннервация тормозных систем миндалевидного тела, а также дифференциальное воздействие на них дофамина позволяют предположить, как могут достигаться различные вегетативные состояния. Сильное влияние pOFC на IM, которое активирует нейроны DARPP-32+ и CB+, может помочь модулировать вегетативную функцию путем снижения регуляции CeM и тем самым облегчать социальные взаимодействия у приматов... С другой стороны, в состоянии паники, когда выживание воспринимается как под угрозой, уровень дофамина заметно возрастает. Таким образом, нейроны DARPP-32+ при ИМ могут быть преимущественно ингибированы, что делает путь pOFC неэффективным.
- ^ Уолтон МЭ; Беренс Т.Э.; Бакли М.Дж.; Рудебек PH; Рашворт МФ (2010). «Отделимые системы обучения в мозге макак и роль орбитофронтальной коры в условном обучении» . Нейрон . 65 (6): 927–939. дои : 10.1016/j.neuron.2010.02.027 . ПМЦ 3566584 . ПМИД 20346766 .
- ^ Кэмпбелл-Мейкледжон Д.К.; Канаи Р.; Бахрами Б.; Бах ДР; Долан Р.Дж.; Ропсторф А.; Компакт-диск Фрита (2012). «Структура орбитофронтальной коры предсказывает социальное влияние» . Современная биология . 22 (4): Р123–Р124. Бибкод : 2012CBio...22.R123C . дои : 10.1016/j.cub.2012.01.012 . ПМК 3315000 . ПМИД 22361146 .
- ^ Танферна А.; Лопес-Хименес Л.; Блас Дж.; Хиральдо Ф.; Серджио Ф. (2012). «Как советы экспертов влияют на принятие решений» . ПЛОС ОДИН . 7 (11): e49748. Бибкод : 2012PLoSO...749748M . дои : 10.1371/journal.pone.0049748 . ПМК 3504100 . ПМИД 23185425 .
- ^ Берридж, КК; Крингельбах, МЛ (6 мая 2015 г.). «Системы удовольствия в мозге» . Нейрон . 86 (3): 646–64. дои : 10.1016/j.neuron.2015.02.018 . ПМЦ 4425246 . ПМИД 25950633 .
- ^ Нуман, Майкл (2015). Нейробиология социального поведения: к пониманию просоциального и антисоциального мозга . Лондон, Великобритания; Уолтем, Массачусетс: Эльзевир. п. 85. ИСБН 978-0-12-416040-8 . OCLC 879584151 .
- ^ Роллс, Эдмунд Т. (2006). «Глава 5 Нейрофизиология и функции орбитофронтальной коры». В Зальде, Дэвид Х.; Раух, Скотт Л. (ред.). Орбитофронтальная кора . Нью-Йорк: Издательство Оксфордского университета.
- ^ Хант LT; Маласекера WMN; де Беркер АО; Миранда Б; Фермер С; Беренс Т.Д.; Кеннерли SW (26 сентября 2018 г.). «Тройная диссоциация внимания и принятия решений в префронтальной коре» . Природная неврология . 21 (9): 1471–1481. дои : 10.1038/s41593-018-0239-5 . ПМК 6331040 . ПМИД 30258238 .
- ^ Шульц, Вольфрам; Трамбле, Леон (2006). «Глава 7: Участие орбитофронтальных нейронов приматов в вознаграждении, неопределенности и обучении 173 Вольфрам Шульц и Леон Трембле». В Зальде, Дэвид Х.; Раух, Скотт: Л. (ред.). Орбитофронтальная кора . Нью-Йорк: Издательство Оксфордского университета.
- ^ Шенбаум Г., Такахаши Ю., Лю Т., МакДанналд М. (2011). «Значит ли орбитофронтальная кора значение?» . Анналы Нью-Йоркской академии наук . 1239 (1): 87–99. Бибкод : 2011NYASA1239...87S . дои : 10.1111/j.1749-6632.2011.06210.x . ПМК 3530400 . ПМИД 22145878 .
- ^ Берлин, Хизер А.; Роллс, Эдмунд Т.; Иверсен, Сьюзен Д. (декабрь 2005 г.). «Пограничное расстройство личности, импульсивность и орбитофронтальная кора» . Американский журнал психиатрии . 162 (12): 2360–2373. дои : 10.1176/appi.ajp.162.12.2360 . ISSN 0002-953X . ПМИД 16330602 .
- ^ Ха, Сонджи; Сон, Ин-Юнг; Ким, Намук; Сим, Хён Чжон; Чхон, Гын-А (декабрь 2015 г.). «Характеристики мозга при расстройствах аутистического спектра: структура, функции и связь на протяжении всей жизни» . Экспериментальная нейробиология . 24 (4): 273–284. дои : 10.5607/en.2015.24.4.273 . ISSN 1226-2560 . ПМЦ 4688328 . ПМИД 26713076 .
- ^ Джековский, АП; Араужо Фильо, генеральный менеджер; Алмейда, АГ; Араужо, СМ; Рейс, М; Нери, Ф; Батиста, ИК; Сильва, я; Ласерда, Алабама (июнь 2012 г.). «Вовлечение орбитофронтальной коры в психические расстройства: обновленные данные нейровизуализации» . Бразильский журнал психиатрии . 34 (2): 207–12. дои : 10.1590/S1516-44462012000200014 . ПМИД 22729418 .
- ^ Милад, MR; Раух, С.Л. (декабрь 2007 г.). «Роль орбитофронтальной коры при тревожных расстройствах». Анналы Нью-Йоркской академии наук . 1121 (1): 546–61. Бибкод : 2007NYASA1121..546M . дои : 10.1196/анналы.1401.006 . ПМИД 17698998 . S2CID 34467365 .
- ^ Накао, Т; Окада, К; Канба, С. (август 2014 г.). «Нейробиологическая модель обсессивно-компульсивного расстройства: данные недавних нейропсихологических исследований и нейровизуализации» . Психиатрия и клинические нейронауки . 68 (8): 587–605. дои : 10.1111/pcn.12195 . ПМИД 24762196 . S2CID 5528241 .
- ^ Файнберг, Н.А.; Потенца, Миннесота; Чемберлен, СР; Берлин, штат Гавайи; Мензис, Л; Бечара, А; Саакян, Б.Дж.; Роббинс, ТВ; Буллмор, ET; Холландер, Э. (февраль 2010 г.). «Исследование компульсивного и импульсивного поведения, от моделей животных до эндофенотипов: обзор повествования» . Нейропсихофармакология . 35 (3): 591–604. дои : 10.1038/нпп.2009.185 . ПМК 3055606 . ПМИД 19940844 .
- ^ Милад, MR; Раух, С.Л. (январь 2012 г.). «Обсессивно-компульсивное расстройство: за пределами отдельных кортико-стриарных путей» . Тенденции в когнитивных науках . 16 (1): 43–51. дои : 10.1016/j.tics.2011.11.003 . ПМЦ 4955838 . ПМИД 22138231 .
- ^ Ваги, М.М; Роббинс, TW (октябрь 2017 г.). «Функциональные нейровизуализационные исследования обсессивно-компульсивного расстройства на основе задач: обзор, основанный на гипотезах». В Питтенджере, Кристофер (ред.). Обсессивно-компульсивное расстройство: феноменология, патофизиология и лечение Обсессивно-компульсивное расстройство: феноменология, патофизиология и лечение . Том. 1. Издательство Оксфордского университета. стр. 239–240. дои : 10.1093/med/9780190228163.003.0022 .
- ^ Шенбаум, Г; Чанг, CY; Лукантонио, Ф; Такахаши, Ю.К. (декабрь 2016 г.). «Нестандартное мышление: орбитофронтальная кора, воображение и как мы можем лечить зависимость» . Нейропсихофармакология . 41 (13): 2966–2976. дои : 10.1038/нпп.2016.147 . ПМК 5101562 . ПМИД 27510424 .
- ^ Кооб, Г.Ф.; Волков, Н.Д. (январь 2010 г.). «Нейросхема зависимости» . Нейропсихофармакология . 35 (1): 217–38. дои : 10.1038/нпп.2009.110 . ПМК 2805560 . ПМИД 19710631 .
- ^ Мурман, Делавэр (2 февраля 2018 г.). «Роль орбитофронтальной коры головного мозга в употреблении алкоголя, злоупотреблении им и зависимости» . Прогресс нейропсихофармакологии и биологической психиатрии . 87 (Часть А): 85–107. дои : 10.1016/j.pnpbp.2018.01.010 . ПМК 6072631 . ПМИД 29355587 .
- ^ Говин, Дж.Л.; Макки, С; Паулюс, член парламента (1 сентября 2013 г.). «Измененная обработка данных, связанная с риском, у потребителей психоактивных веществ: дисбаланс боли и выгоды» . Наркотическая и алкогольная зависимость . 132 (1–2): 13–21. doi : 10.1016/j.drugalcdep.2013.03.019 . ПМЦ 3748224 . ПМИД 23623507 .
У людей с SUD наблюдаются некоторые нарушения обработки во время принятия рискованных решений, которые включают измененную оценку вариантов (VMPFC) и результатов (OFC и полосатое тело), плохую оценку неопределенности (ACC и островковая кора), снижение исполнительного контроля (DLPFC), ослабленное влияние эмоциональной значимости (миндалевидное тело) и снижение чувствительности к соматическим маркерам (соматосенсорная кора). Эти различия в нейронной обработке во время принятия рискованных решений среди людей с СНС были связаны с более низкими поведенческими показателями при выполнении рискованных задач и более обширной историей употребления психоактивных веществ.
- ^ Чейз, HW; Эйкхофф, С.Б.; Лэрд, Арканзас; Хогарт, Л. (15 октября 2011 г.). «Нейронная основа обработки стимулов к наркотикам и тяги к ним: метаанализ оценки вероятности активации» . Биологическая психиатрия . 70 (8): 785–93. doi : 10.1016/j.biopsych.2011.05.025 . ПМЦ 4827617 . ПМИД 21757184 .
Медиальная область OFC демонстрировала большую активацию сигналами препарата по сравнению с контрольными сигналами и последовательно активировалась в подгруппе, не обращавшейся за лечением. Имеются убедительные доказательства того, что эта область играет роль в аппетитном поведении и принятии решений (86,87), особенно в отношении ожидания вознаграждения (88), предсказанного условными стимулами (89–94), которые могут контролировать выбор инструментальных действий.
- ^ Лукантонио, Ф; Каприоли, Д; Шенбаум, Г. (январь 2014 г.). «Переход от «модельного» к «безмодельному» поведенческому контролю при зависимости: вовлечение орбитофронтальной коры и дорсолатерального полосатого тела» . Нейрофармакология . 76 Часть Б: 407–15. doi : 10.1016/j.neuropharm.2013.05.033 . ПМК 3809026 . ПМИД 23752095 .
- ^ Рубия, К. (15 июня 2011 г.). « «Холодная» нижняя лобно-стриатальная дисфункция при синдроме дефицита внимания и гиперактивности по сравнению с «горячей» вентромедиальной орбитофронтально-лимбической дисфункцией при расстройстве поведения: обзор». Биологическая психиатрия . 69 (12): с69–87. doi : 10.1016/j.biopsych.2010.09.023 . ПМИД 21094938 . S2CID 14987165 .
- ^ Роселл, ДР; Сивер, ЖЖ (июнь 2015 г.). «Нейробиология агрессии и насилия» . Спектры ЦНС . 20 (3): 254–79. дои : 10.1017/S109285291500019X . ПМИД 25936249 .
- ^ Перейти обратно: а б Роллс, ET (сентябрь 2016 г.). «Теория депрессии, не связанная с вознаграждением» (PDF) . Неврологические и биоповеденческие обзоры . 68 : 47–58. doi : 10.1016/j.neubiorev.2016.05.007 . ПМИД 27181908 . S2CID 8145667 .
- ^ Дж. Уилсон; М. Дженкинсон; ИЭПП де Араужо; Мортен Л. Крингельбах ; ET Rolls и Питер Джеззард (октябрь 2002 г.). «Быстрая, полностью автоматизированная глобальная и локальная оптимизация магнитного поля для фМРТ человеческого мозга». НейроИмидж . 17 (2): 967–976. дои : 10.1016/S1053-8119(02)91172-9 . ПМИД 12377170 .
- ^ Ахмари, ЮВ; Догерти, Д.Д. (август 2015 г.). «Рассечение цепей OCD: от моделей на животных до целенаправленного лечения» . Депрессия и тревога . 32 (8): 550–62. дои : 10.1002/da.22367 . ПМЦ 4515165 . ПМИД 25952989 .
- ^ Вертечи, Пьетро; Лоттем, Эран; Сарра, Дарио; Годиньо, Беатрис; Тревес, Исаак; Кендера, Тьяго; Лохуис, Маттейс Николаи Уде; Майнен, Закари Ф. (08 апреля 2020 г.). «Решения на основе умозаключений в задаче поиска пищи в скрытом состоянии: дифференциальный вклад префронтальных областей коры» . Нейрон . 106 (1): 166–176.e6. дои : 10.1016/j.neuron.2020.01.017 . ISSN 0896-6273 . ПМК 7146546 . ПМИД 32048995 .
- ^ Сноуден Дж.С.; Батгейт Д.; Варма А.; Блэкшоу А.; Гиббонс ZC; Нири Д. (2001). «Отличные поведенческие профили при лобно-височной деменции и семантической деменции» . J Neurol Нейрохирургия Психиатрия . 70 (3): 323–332. дои : 10.1136/jnnp.70.3.323 . ПМЦ 1737271 . ПМИД 11181853 .
- ^ Сяо, Сюн; Дэн, Ханфэй; Вэй, Лей; Хуан, Яньван; Ван, Цзожэнь (сентябрь 2016 г.). «Нейральная активность орбитофронтальной коры способствует управлению ожиданием» . Европейский журнал неврологии . 44 (6): 2300–2313. дои : 10.1111/ejn.13320 . ISSN 1460-9568 . ПМИД 27336203 . S2CID 205105682 .
- ^ Перейти обратно: а б Паулюс член парламента; Хозак Н.Е.; Заушер Б.Е.; Фрэнк Л.; Браун Г.Г.; Брафф Д.Л.; Шукит М.А. (2002). «Поведенческие и функциональные данные нейровизуализации префронтальной дисфункции у субъектов, зависимых от метамфетамина» . Нейропсихофармакология . 26 (1): 53–63. дои : 10.1016/s0893-133x(01)00334-7 . ПМИД 11751032 .
- ^ Перейти обратно: а б Топлак МЕ; Джайн У.; Таннок Р. (2005). «Исполнительные и мотивационные процессы у подростков с синдромом дефицита внимания и гиперактивности (СДВГ)» . Поведенческие и функции мозга . 1 (1): 8–20. дои : 10.1186/1744-9081-1-8 . ПМЦ 1183187 . ПМИД 15982413 .
- ^ Перейти обратно: а б Вердехо-Гарсия А.; Бечара А.; Рекнор ЕС; Перес-Гарсия М. (2006). «Исполнительная дисфункция у лиц, зависимых от психоактивных веществ во время употребления наркотиков и воздержания: исследование поведенческих, когнитивных и эмоциональных коррелятов зависимости». Журнал Международного нейропсихологического общества . 12 (3): 405–415. дои : 10.1017/s1355617706060486 . ПМИД 16903133 . S2CID 15939155 .
- ^ Ча, Джиук; Гринберг, Цафрир; Карлсон, Джошуа М.; ДеДора, Дэниел Дж.; Хайчак, Грег; Мухика-Пароди, Лилианна Р. (12 марта 2014 г.). «Общеструктурные и функциональные показатели прогнозируют генерализацию страха в вентромедиальной префронтальной коре: последствия для генерализованного тревожного расстройства» . Журнал неврологии . 34 (11): 4043–4053. doi : 10.1523/JNEUROSCI.3372-13.2014 . ISSN 0270-6474 . ПМК 6705282 . ПМИД 24623781 .
- ^ Волков Н.Д.; Фаулер Дж.С. (2000). «Наркомания – болезнь навязчивости и влечений: поражение орбитофронтальной коры» . Кора головного мозга . 10 (3): 318–325. дои : 10.1093/cercor/10.3.318 . ПМИД 10731226 .
- ^ Чемберлен-старший; Одлауг БЛ; Булугури В.; Файнберг Н.А.; Грант Дж. Э. (2009). «Трихотилломания: нейробиология и лечение». Неврологические и биоповеденческие обзоры . 33 (6): 831–842. doi : 10.1016/j.neubiorev.2009.02.002 . ПМИД 19428495 . S2CID 6956143 .
- ^ Мензис Л. (2008). «Интеграция данных нейровизуализации и нейропсихологических исследований обсессивно-компульсивного расстройства: пересмотр орбитофронто-стриарной модели» . Неврологические и биоповеденческие обзоры . 32 (3): 525–549. doi : 10.1016/j.neubiorev.2007.09.005 . ПМЦ 2889493 . ПМИД 18061263 .
- ^ Перейти обратно: а б Текин С.; Каммингс Дж.Л. (2002). «Фронтально-подкорковые нейрональные цепи и клиническая нейропсихиатрия: обновление». Журнал психосоматических исследований . 53 (2): 647–654. дои : 10.1016/s0022-3999(02)00428-2 . ПМИД 12169339 .
- ^ Рахман С.; Саакян Б.Дж.; Ходжес-младший; Роджерс Р.Д.; Роббинс Т.В. (1999). «Специфические когнитивные нарушения при раннем поведенческом варианте лобно-височной деменции» . Мозг . 122 (8): 1469–1493. дои : 10.1093/мозг/122.8.1469 . ПМИД 10430832 .
- ^ Сили WW; Кроуфорд Р.; Расковский К.; Крамер Дж. Х.; Вайнер М.; Миллер Б.Л.; Горно-Темпини Л. (2008). «Атрофия лобной паралимбической сети при очень легком поведенческом варианте лобно-височной деменции» . Архив неврологии . 65 (2): 249–255. дои : 10.1001/archneurol.2007.38 . ПМЦ 2544627 . ПМИД 18268196 .
- ^ Чибане, Исландия; Баучер, О; Дюбо, Ф; Тран, TPY; Мохамед, я; Маклахлан, Р.; Сэдлер, РМ; Дебьен, Р; Кармант, Л; Нгуен, Дания (ноябрь 2017 г.). «Орбитофронтальная эпилепсия: серия случаев и обзор литературы». Эпилепсия и поведение . 76 : 32–38. дои : 10.1016/j.yebeh.2017.08.038 . ПМИД 28928072 . S2CID 13656956 .
- ^ Голд, Дж.А.; Шер, Ю; Мальдонадо, младший (2016). «Лобная долевая эпилепсия: учебник для психиатров и систематический обзор психиатрических проявлений». Психосоматика . 57 (5): 445–64. дои : 10.1016/j.psym.2016.05.005 . ПМИД 27494984 .
- ^ Смит, младший; Силлай, К; Винклер, П; Кинг, Д.В.; Лоринг, Д.В. (2004). «Орбитофронтальная эпилепсия: электроклинический анализ хирургических случаев и обзор литературы» . Стереотаксическая и функциональная нейрохирургия . 82 (1): 20–5. дои : 10.1159/000076656 . ПМИД 15007215 . S2CID 18811550 .
- ^ Роллс ЕТ; Хорнак Дж.; Уэйд Д.; МакГрат Дж. (1994). «Обучение, связанное с эмоциями, у пациентов с социальными и эмоциональными изменениями, связанными с повреждением лобных долей» . J Neurol Нейрохирургия Психиатрия . 57 (12): 1518–1524. дои : 10.1136/jnnp.57.12.1518 . ПМЦ 1073235 . ПМИД 7798983 .
- ^ Бечара А.; Дамасио А.Р.; Дамасио Х.; Андерсон С.В. (1994). «Нечувствительность к будущим последствиям повреждения префронтальной коры человека». Познание . 50 (1–3): 7–15. дои : 10.1016/0010-0277(94)90018-3 . ПМИД 8039375 . S2CID 204981454 .
- ^ Камень ВЭ; Барон-Коэн С.; Найт RT (1998а). «Вклад лобной доли в теорию разума». Журнал медицинских расследований . 10 (5): 640–656. CiteSeerX 10.1.1.330.1488 . дои : 10.1162/089892998562942 . ПМИД 9802997 . S2CID 207724498 .