Физиология динозавров
Физиология динозавров исторически была спорным предметом, особенно их терморегуляцией . В последнее время было представлено много новых доказательств, которые обычно приводят к физиологии динозавров , включая не только метаболические системы и терморегуляцию, но и в респираторных и сердечно -сосудистых системах.
В первые годы палеонтологии динозавров широко считалось, что они были вялыми, громоздкими и растягивающимися хладнокровными ящерицами . [ сомнительно - обсудить ] Тем не менее, с обнаружением гораздо более полных скелетов в западных Соединенных Штатах, начиная с 1870 -х годов, ученые могут сделать более осознанные интерпретации биологии и физиологии динозавров. Эдвард Дрибер Коуп , противник Отниэля Чарльза Марша в костяных войнах , выдвинул, по крайней мере, некоторых динозавров как активных и мудрых, как видно на картине двух боевых лапоретов, произведенных под его руководством Чарльзом Р. Найтом . [ 1 ]
Параллельно, развитие дарвиновской эволюции и открытия Archeopteryx и Compsognathus заставили Томаса Генри Хаксли , чтобы предложить, чтобы динозавры тесно связаны с птицами. [ 2 ] Несмотря на эти соображения, образ динозавров как большие рептилии уже укоренились, [ 1 ] и большинство аспектов их палеобиологии были истолкованы как обычно рептилий в первой половине двадцатого века. [ 3 ] Начиная с 1960 -х годов и с появлением эпохи эпохи динозавров , виды динозавров и их физиологии резко изменились, в том числе открытие пернатых динозавров в ранних меловых возрастных месторождениях в Китае, что указывает на то, что птицы эволюционировали из очень гибких манирапторанских динозавров.
История
[ редактировать ]Ранние интерпретации
[ редактировать ]

Изучение динозавров началось в 1820 -х годах в Англии. Пионеры в этой области, такие как Уильям Бакленд , Гидеон Мантелл и Ричард Оуэн , интерпретировали первые, очень фрагментарные останки как принадлежащие к крупным квадрупедным зверям. [ 4 ] Их ранняя работа можно увидеть сегодня в динозаврах Crystal Palace , построенных в 1850 -х годах, которые представляют известных динозавров в качестве Elephantine . рептилий, подобных [ 5 ] Несмотря на эти выступления рептилий, Оуэн предположил, что сердце и респираторные системы динозавров были более похожи на схему млекопитающего, чем рептилия. [ 4 ]
Изменение видов и эпохи динозавров
[ редактировать ]В конце 1960 -х годов появились подобные идеи, начиная с Джона Острома работы над Deinonychus и Evolution . [ 6 ] Его ученик, Боб Баккер , популяризировал меняющуюся мысль в серии бумаг, начиная с превосходства динозавров в 1968 году. [ 7 ] В этих публикациях он сильно утверждал, что динозавры были теплыми и активными животными, способными к устойчивым периодам высокой активности. В большинстве своих произведений Баккер создал свои аргументы как новые доказательства, ведущие к возрождению идей, популярных в конце 19 -го века, часто ссылаясь на продолжающийся ренессанс динозавров . Он использовал множество анатомических и статистических аргументов для защиты своего дела, [ 8 ] [ 9 ] Методология которого была яростно обсуждалась среди ученых. [ 10 ]
Эти дебаты вызвали интерес к новым методам определения палеобиологии вымерших животных, таких как гистология кости , которые были успешно применены для определения скоростей роста многих динозавров.
Сегодня обычно считается, что многие или, возможно, у всех динозавров были более высокие показатели метаболизма, чем живые рептилии, но также и то, что ситуация более сложная и разнообразная, чем первоначально предложил Баккер. Например, в то время как меньшие динозавры могли быть истинными эндотермами , более крупными формами могли быть инерционные гомеотермы , [ 11 ] [ 12 ] Или что многие динозавры могли иметь промежуточные показатели метаболизма. [ 13 ]
Кормление и пищеварение
[ редактировать ]Самыми ранними динозаврами почти наверняка были хищники и поделились несколькими хищническими особенностями с их ближайшими родственниками, не связанными с динозаврами, такими как Лагосутус , в том числе: относительно большие изогнутые лезвия зубы в больших, широко раскрывающихся челюстях, которые закрываются как ножницы; Относительно небольшие животы , поскольку плотоядные не требуют больших пищеварительных систем . Позже динозавры считались хищниками, иногда росли намного больше, но сохраняли один и тот же набор функций. Вместо того, чтобы жевать пищу, эти хищники проглотили ее целесообразно. [ 14 ]
Привычки кормления орнитомимозавров и овираптозавров являются загадкой: хотя они развивались из теропод хищной Полем [ 14 ]
Особенности других групп динозавров показывают, что они были травоядными. Эти функции включают в себя:
- Челюсти, которые лишь слегка открылись и закрылись, так что все зубы встречались одновременно
- Большие животы, которые могут вместить большие растительности и хранить ее в течение долгого времени, чтобы переваривать растительность
- Кишки, которые, вероятно, содержали эндосимбиотические микроорганизмы, которые переваривают целлюлозу , поскольку ни одно известное животное не может переваривать этот жесткий материал напрямую [ 14 ]
Сауроподы , которые были травоядными, не жевали их пищу, так как их зубы и челюсти кажутся подходящими только для снятия листьев с растений. Орнитишки , также травоядные, показывают различные подходы. У бронированных анкилозавров и стегозавров были маленькие головы, слабые челюсти и зубы, и, как полагают, питались почти так же, как и Sauropods. были У пахицефалозавров небольшие головы, слабые челюсти и зубы, но их отсутствие больших пищеварительных систем предполагает другую диету, возможно, фрукты, семена или молодые побеги, которые были бы более питательными для них, чем листья. [ 14 ]
С другой стороны, орнитоподы, такие как Hypsilophodon , Iguanodon и различные гадрозавры, имели возбужденные клювы для обдумывания растительности, челюстей и зубов, которые были хорошо адаптированы для жевания. У рогатых цератопсийцев были похожие механизмы. [ 14 ]
Часто предполагалось, что, по крайней мере, некоторые динозавры использовали проглатываемые камни, известные как гастролиты , чтобы помочь пищеварению, размолов пищу в мускулистых желумце , и что это была особенность, которую они делили с птицами. В 2007 году Оливер Вингс рассмотрел ссылки на гастролиты в научной литературе и обнаружил значительную путаницу, начиная с отсутствия согласованного и объективного определения «гастролита». Он обнаружил, что проглатываемые твердые камни или пескарь могут помочь пищеварению у птиц, которые в основном питаются зерном, но могут быть не важны - и что птицы, которые едят насекомых летом, и зерно зимой обычно избавляются от камней и песка летом. Гастролиты часто описывались как важные для динозавров Sauropod , чья диета растительности требовала очень тщательного пищеварения, но крылья пришли к выводу, что эта идея была неверной: гастролиты обнаруживаются только с небольшим процентом окаменелостей Sauropod; Там, где они были обнаружены, количество слишком малы, и во многих случаях камни слишком мягкие, чтобы быть эффективными в шлифовании пищи; Большинство из этих гастролитов высоко отполированы, но гастролиты, используемые современными животными для измельчения пищи, шерохованы износом и коррозируются желудочными кислотами; Следовательно, гастролиты Sauropod, вероятно, были проглочены случайно. С другой стороны, он пришел к выводу, что гастролиты, найденные с ископаемыми продвинутыми Динозавры, такие как Sinornithomus и Caudipteryx, напоминают динозавры, и что использование гастролитов для измельчения пищи могло появиться в начале группы динозавров, из которых эти динозавры и птицы развивались. [ 15 ] [ 16 ]
Репродуктивная биология
[ редактировать ]При откладывании яиц женские птицы выращивают особый тип кости в конечностях между твердой внешней костью и костным мозгом . [ 17 ] Эта медуллярная кость, которая богата кальцием , используется для изготовления яичных скорлупы, а птицы, которые вырабатывали его, поглощают его, когда они закончили откладывать яйца. [ 18 ] Медуллярная кость была обнаружена у окаменелостей Theropods Tyrannosaurus и Allosaurus и Ornithopod Tenontosaurus . [ 18 ] [ 19 ]
Поскольку линия динозавров, которая включает в себя аллозавр и тиранозавр, расходилась от линии, которая привела к тому, что Tenontosaurus очень рано в эволюции динозавров, наличие медуллярной кости в обеих группах предполагает, что динозавры в целом продуцируют медуллельную ткань. С другой стороны, крокодильцы родственниками динозавров , которые являются вторыми ближайшими динамичными после птиц, не производят медуллярную кость. Эта ткань, возможно, впервые появилась в Ornithodires , триасовой группе архозавров , из которой, как считается, эволюционировали динозавры. [ 18 ]
Медуллярная кость была обнаружена в образцах размера суб-адольта, что говорит о том, что динозавры достигли сексуальной зрелости, прежде чем они стали полноценными. Сексуальная зрелость в размере суб-адульта также встречается у рептилий и у млекопитающих среднего и крупного размера, но птицы и мелкие млекопитающие достигают сексуальной зрелости только после того, как они взрослые, что происходит в течение их первого года. Ранняя сексуальная зрелость также связана со специфическими особенностями жизненных циклов животных: молодые рождаются относительно хорошо развитыми, а не беспомощными; и скорость смерти среди взрослых высока. [ 18 ]
Респираторная система
[ редактировать ]Воздушные мешки
[ редактировать ]
Примерно с 1870 года ученые в целом согласились с тем, что посткраниальные скелеты многих динозавров содержали много полостей, заполненных воздухом ( посткраниальная скелетная пневматичность , особенно в позвонках. Пневматизация черепа (такие как параназальные пазухи ) обнаруживается в обоих синапсах и архозаврах , но посткраниальная пневматизация встречается только у птиц, неавийских сауришских динозавров и птерозавров .
В течение долгого времени эти полости рассматривались просто как устройства, способствующие весам, но Баккер предположил, что они были связаны с воздушными мешками, птиц такими как те, которые делают дыхательные системы наиболее эффективными из всех животных ». [ 9 ]
Джон Рубен и соавт. внутри и выезд), прикрепленной к крокодилоподобному печенообразному механизму-мышцы, прикрепленные в основном лобкому к (1997, 1999, 2003, 2004) оспаривали это и предположили, что динозавры имели «приливную» дыхательную систему ( легкие расширяются до вдыхания; Когда эти мышцы расслабляются, легкие возвращаются к своему предыдущему размеру и форме, а животное выдыхает. Они также представили это как причину сомнения, что птицы произошли от динозавров . [ 20 ] [ 21 ] [ 22 ] [ 23 ] [ 24 ]
Критики утверждают, что без птичьих воздушных мешков скромные улучшения в нескольких аспектах систем кровообращения современной рептилии позволили бы рептилию достичь от 50% до 70% потока кислорода млекопитающего схожного размера, [ 25 ] И что отсутствие птичьих воздушных мешков не предотвратит развитие эндотермии. [ 26 ] Очень немногие официальные опровержения были опубликованы в научных журналах Ruben et al. Утверждение о том, что динозавры не могли иметь воздушные мешки в стиле птиц; Но один указывает, что окаменелости Sinosauropteryx , на котором они основывали большую часть своих аргументов, было серьезно сплющено, и поэтому было невозможно сказать, была ли печень правильной формой, чтобы действовать как часть печеночного поршневого механизма. [ 27 ] Некоторые недавние документы просто отмечают без дальнейших комментариев, что Ruben et al. спорил против присутствия воздушных мешков у динозавров. [ 28 ]
Исследователи представили доказательства и аргументы в пользу воздушных мешочков в сауроподах , « Просауропод », коелурозавра , цератозавры , а также Теропод аэростон и колофиз .
В продвинутых сауроподах («Neosauropods») позвонки областей нижней части спины и бедра показывают признаки воздушных мешочков. В ранних сауроподах только позвонки шейки матки (шея) показывают эти особенности. Если последовательность развития, обнаруженная у эмбрионов птиц, является руководством , воздушные мешки фактически развивались перед каналами в скелете, которые размещают их в более поздних формах. [ 29 ] [ 30 ]

Свидетельство о воздушных мешках также было обнаружено в Тероподах. Исследования показывают, что окаменелости коелурозавров , [ 31 ] цератозавры , [ 28 ] и теропод Coelophysis и Aerosteon демонстрируют свидетельство воздушных мешков. Коелофиз , из позднего триаса , является одним из первых динозавров, у которых окаменелости свидетельствуют каналы для воздушных мешочков. [ 30 ] Аэростеон , поздний меловой аллозавра , имел наиболее похожие на птицы воздушные мешки. [ 32 ]
Ранние сауроподоморфы , в том числе группа, традиционно называемая «просауроподами», также могли иметь воздушные мешки. Несмотря на возможные пневматические углубления в Plateosaurus и Thecodontosaurus , в отступлениях очень малы. В одном исследовании в 2007 году пришло выяснение, что у просауропод, вероятно, были брюшные и шейные воздушные мешки, основанные на доказательствах для их сестринских таксонов (Theropods и Sauropods). Исследование пришло к выводу, что было невозможно определить, имели ли у прозауроподс легкие, похожие на птицу легкое, но воздушные мешки почти наверняка присутствовали. [ 33 ] Дальнейшее указание на наличие воздушных мешков и их использование в вентиляции легких связано с реконструкцией объема воздушного обмена (объем воздуха, обменяемый с каждым дыханием) Plateosaurus , который при выраженном соотношении объема воздуха на массу тела при 29 мл/кг аналогичны значениям гусей и других птиц и намного выше, чем типичные значения млекопитающих. [ 34 ]
До сих пор никаких доказательств воздушных мешочков не было обнаружено у орнитишских динозавров. Но это не подразумевает, что у орнитишхов не было показателей метаболизма, сопоставимых с показателями млекопитающих, поскольку у млекопитающих также нет воздушных мешочков. [ 35 ]
Было предложено три объяснения для развития воздушных мешков в динозаврах: [ 32 ]
- Увеличение дыхательной способности. Это, вероятно, самая распространенная гипотеза, и она хорошо соответствует идее, что многие динозавры имели довольно высокие показатели метаболизма .
- Улучшение баланса и маневренности путем снижения центра тяжести и снижения вращательной инерции . Однако это не объясняет расширение воздушных мешков в четвероногих сауроподах.
- Как механизм охлаждения. Кажется, что воздушные мешки и перья развивались примерно в одно и то же время в колурозаврах . Если перья сохраняют тепло, их владельцы потребовали бы средства рассеивания избыточного тепла. Эта идея правдоподобна, но нуждается в дальнейшей эмпирической поддержке.
Расчеты объемов различных частей респираторной системы Sauropod подтверждают Apatosaurus доказательства воздушных мешков, подобных птицам, у Sauropods:
- Предполагая, что апатозавра , как и ближайшие выжившие родственники динозавров, крокодильцы и птицы, не имели диафрагмы , объем мертвого пространства 30-тонного образца будет около 184 литров. Это общий объем рта, трахеи и воздушных трубок. Если животное выдыхает меньше этого, устаревший воздух не исключен и высасывается обратно в легкие при следующем вдыхании.
- Оценки его приливного объема - количество воздуха, перемещаемого в легкие или из легких за одним дыханием - зависят от типа дыхательной системы, которую у животного имело: 904 литров, если я птицы; 225 литров, если млекопитающий; 19 литров, если рептил.
Исходя из этого, у Apatosaurus не было респираторной системы рептилий, так как его приливный объем был бы меньше, чем его объем мертвого пространства, так что устаревший воздух не был исключен, но был отсосан в легкие. Аналогичным образом, система млекопитающих предоставила бы легким только около 225 - 184 = 41 литра свежего, кислородного воздуха на каждом дыхании. Поэтому апатозавра , должно быть, имела либо систему, неизвестную в современном мире, либо такую, как птицы , с множественными воздушными мешками и протекающим легким. Кроме того, птичья система потребуется только объем легких в размере около 600 литров, в то время как млекопитающее потребовалось бы около 2950 литров, что превысило бы около 1700 литров места, доступного в 30-тонном сундуке апатозавра . [ 36 ]
Респираторные системы динозавров с птицами, похожими на воздушные мешки, могли быть способны поддерживать более высокие уровни активности, чем млекопитающие с одинаковым размером и сборкой. В дополнение к обеспечению очень эффективного запаса кислорода, быстрый поток воздуха был бы эффективным механизмом охлаждения, который необходим для животных, которые активны, но слишком велики, чтобы избавиться от избыточного тепла через их шкуры. [ 35 ]
Палеонтолог Питер Уорд утверждал, что эволюция системы воздушного мешка, которая впервые появляется в самых ранних динозаврах, могла быть в ответ на очень низкий (11%) атмосферный кислород карнского и норьянного периода триаса. [ 37 ]

Неушинные процессы на ребрах
[ редактировать ]У птиц есть шпоры, называемые « неконтролируемыми процессами » на задней краях их ребер, и они придают мышцам груди больше рычагов при перекачке груди для улучшения подачи кислорода. Размер незрелых процессов связан с потребностями в образе жизни птиц и кислороде: они самые короткие с птицами для ходьбы и самые длинные в дайвинговых птицах, которые должны быстро пополнить свои запасы кислорода, когда они поверхностно. Неавийские манирапторанские динозавры также имели эти неконтролируемые процессы, и они были пропорционально до тех пор, пока у современных птиц, что указывает на то, что манирапторам нуждались в подаче кислорода с высокой емкостью. [ 38 ] [ 39 ]
Пластины, которые, возможно, функционировали так же, как и неведанизированные процессы, наблюдались у окаменелостей орнитишского динозавра Thescelosaurus , и были интерпретированы как доказательство высокого потребления кислорода и, следовательно, высокой скорости метаболизма. [ 40 ]
Носовые турбинаты
[ редактировать ]Носовые турбинаты представляют собой страшные структуры тонкой кости в полости носа . У большинства млекопитающих и птиц они присутствуют и выкладываются слизистыми мембранами , которые выполняют две функции. Они улучшают обоняние, увеличивая область, доступную для поглощения воздушных химических веществ, и они тепло и увлажняют вдыхаемый воздух, а также извлекают тепло и влагу из выдоха от воздуха, чтобы предотвратить высыхание легких.
Джон Рубен и другие утверждают, что у динозавров не было обнаружено никаких доказательств носовых турбинатов. Все исследованные ими динозавры, имели носовые отрывки, которые были слишком узкими и короткими, чтобы вместить носовые турбинаты, поэтому динозавры не могли бы выдержать частоту дыхания, необходимую для млекопитающего или птичьего метаболизма, пока в состоянии покоя, потому что их легкие высыхали бы вне. [ 20 ] [ 21 ] [ 41 ] [ 42 ] Тем не менее, против этого аргумента были выдвинуты возражения. Носовые турбинаты отсутствуют или очень маленькие у некоторых птиц (например, катисты , прокеллариформные и соколиформные ) и млекопитающих (например, киты, муравьеды , летучие мыши , слоны и большинство приматов ), хотя эти животные полностью эндотермические и в некоторых случаях очень активны. [ 43 ] [ 44 ] [ 45 ] [ 46 ] Другие исследования приходят к выводу, что носовые турбинаты хрупкие и редко встречаются у окаменелостей. В частности, никто не был найден в ископаемых птицах. [ 47 ]
В 2014 году Джейсон Бурк и другие в анатомическом отчете сообщили о обнаружении носовых турбинатов у пахицефалозавров. [ 48 ]
Сердечно -сосудистая система
[ редактировать ]
В принципе можно было бы ожидать, что у динозавров будет двух частей циркуляций, управляемых четырех камерными сердцами, так как многим потребовалось бы высокое кровяное давление, чтобы доставить кровь к их головам, которые были высокими от земли, но легкие позвоночных могут только переносить довольно низкие артериальное давление. [ 35 ] В 2000 году был описан скелет Thescelosaurus , который в настоящее время выставлен в музее естественных наук Северной Каролины , был описан как включающие остатки четырех камерного сердца и аорту . Авторы интерпретировали структуру сердца как указывающая на повышенную скорость метаболизма для TheScelosaurus , а не рептилия хладнокровного . [ 49 ] Их выводы были оспорены; Другие исследователи опубликовали статью, в которой они утверждают, что сердце действительно является конкрецией полностью минерального «цемента». Как они отмечают: анатомия, приведенная для объекта, неверна, например, предполагаемая «аорта» является самой узкой, где она встречается с «сердцем» и не хватает артерий, разветвленных от нее; «Сердце» частично охватывает одно из ребер и в некоторых местах имеет внутреннюю структуру концентрических слоев; и еще одна конкреция сохраняется за правой ногой. [ 50 ] Оригинальные авторы защищали свою позицию; Они согласились с тем, что грудь содержит тип конкреции, но тот, который сформировался вокруг и частично сохранил более мышечные части сердца и аорты. [ 51 ]
Независимо от личности объекта, он может иметь небольшую степень для внутренней анатомии динозавров и скорости метаболизма. Как современные крокодильцы , так и птицы, самые близкие живые родственники динозавров, имеют четырехкамерные сердца, хотя и модифицированные у крокодилов, и поэтому динозавры, вероятно, также имели их. Однако такие сердца не обязательно связаны с скоростью метаболизма. [ 52 ]
Рост и жизненный цикл
[ редактировать ]Яйцо динозавра не было обнаружено, которое больше, чем баскетбол, и эмбрионы крупных динозавров были обнаружены в относительно маленьких яйцах, например, Маайасаура . [ 53 ] Как и млекопитающие, динозавры перестали расти, когда они достигли типичного размера взрослых своих видов, в то время как зрелые рептилии продолжали расти медленно, если у них достаточно пищи. Динозавры всех размеров росли быстрее, чем современные рептилии аналогичного размера; Но результаты сравнений с «теплыми» современными животными аналогичного размера зависят от их размеров: [ 54 ] [ 55 ]
| Вес (кг) | Сравнительные темпы роста динозавров | Современные животные в таком размере |
|---|---|---|
| 0.22 | Медленнее, чем сумчатые | Крыса |
| 1 – 20 | Аналогично сумчатым, медленнее, чем прекосоциальные птицы (те, которые рождаются, способны к бегу) | От морской свинки до Андина Кондора |
| 100 – 1000 | Быстрее, чем сумчатые, похожие на предварительные птицы, медленнее, чем плацентарные млекопитающие | От красного кенгуру до белого медведя |
| 1500 – 3500 | Похоже на большинство плацентарных млекопитающих | От американского бизона до носорога |
| 25000 и более | Очень быстро, похоже на современные киты ; Но примерно вдвое меньше, чем у уменьшенной альтрической птицы (той, которая родилась беспомощной)-если бы можно было увеличить птицу до 25 000 килограммов (25 длинных тонн; 28 коротких тонн) | Киты |

Tyrannosaurus rex показал «подростковый всплеск роста»: [ 56 ] [ 57 ]
- ½ тонны в возрасте 10 лет
- Очень быстрый рост до 2 тонн в середине подросткового возраста (около ½ тонны в год).
- незначительный рост после второго десятилетия.
Исследование, проведенное в 2008 году одного скелета HADROSAUR Hypacrosaurus, пришло к выводу, что этот динозавр вырос еще быстрее, достигая полноценного размера в возрасте около 15 лет; Основным доказательством было количество и расстояние кольца роста в его костях. Авторы обнаружили, что это согласуется с теорией жизненного цикла, что виды добычи должны расти быстрее, чем их хищники, если они теряют много несовершеннолетних из-за хищников, а местная среда обеспечивает достаточно ресурсов для быстрого роста. [ 58 ]
Похоже, что отдельные динозавры были довольно недолгими, например, самым старым (после смерти), обнаруженного Тираннозавром до сих пор, был 28, а самым старым зауроподом было 38 лет. [ 56 ] Хищничество, вероятно, было ответственным за высокий уровень смертности очень молодых динозавров и сексуальную конкуренцию за высокий уровень смертности от сексуально зрелых динозавров. [ 59 ]
Метаболизм
[ редактировать ]Научное мнение о стиле жизни, метаболизме и температурной регуляции динозавров с течением времени варьировалось с момента открытия динозавров в середине 19-го века. Активность метаболических ферментов варьируется в зависимости от температуры, поэтому контроль температуры жизненно важен для любого организма, будь то эндотермический или эктотермический. Организмы могут быть классифицированы как poikilotherms (poikilo - изменение), которые терпимы к внутренним колебаниям температуры, и гомеотермы (гомео - то же самое), которые должны поддерживать постоянную температуру ядра. Животные могут быть дополнительно классифицированы как эндотермы , которые регулируют их температуру внутренне, и эктотермы , которые регулируют температуру с помощью внешних источников тепла.
« Теплая кровяность »-это сложный и довольно неоднозначный термин, потому что он включает в себя некоторые или все:
- Гомеотермия , т.е. поддержание довольно постоянной температуры тела. Современные эндотермы поддерживают различные температуры: от 28 ° C (от 82 ° F) до 30 ° C (86 ° F) в монотриях и лени ; 33 ° C (91 ° F) до 36 ° C (97 ° F) в сумчатых ; 36 ° C (97 ° F) до 38 ° C (100 ° F) в большинстве плацент ; и около 41 ° C (106 ° F) у птиц. [ 35 ]
- Тахиметаболизм , т.е. поддержание высокой скорости метаболизма , особенно в состоянии покоя. Это требует довольно высокой и стабильной температуры тела, поскольку биохимические процессы работают примерно вдвое меньше, если температура животного падает на 10 ° C; Большинство ферментов имеют оптимальную рабочую температуру , и их эффективность быстро падает за пределы предпочтительного диапазона. [ 60 ]
- Endothermy , т.е. способность генерировать тепло внутри страны, например, «сжигая» жир, а не через поведение, такое как суть или мышечная активность. Хотя эндотермия в принципе является наиболее надежным способом поддержания довольно постоянной температуры, она дорого; Например, современным млекопитающим в 10-13 раз больше еды, чем современные рептилии. [ 35 ]
Крупные динозавры также могли сохранять свою температуру инерционной гомеотермией, также известной как «объемная гомеотермия» или «массовая гомеотермия». Другими словами, тепловая способность таких крупных животных была настолько высокой, что потребуется два дня или более, чтобы их температура значительно изменилась, и это сглажило бы вариации, вызванные ежедневными температурными циклами. Этот сглаживающий эффект наблюдался у крупных черепах и крокодилов , но Plateosaurus , который весил около 700 килограммов (1500 фунтов), мог быть самым маленьким динозавром, в котором он был бы эффективным. Инерционная гомеотермия не была бы возможна для маленьких видов, ни для молодых из более крупных видов. [ 35 ] Ферментация растительности в кишках больших травоядных животных также может производить значительное тепло, но этот метод поддержания высокой и стабильной температуры не был бы возможен для плотоядных животных или для небольших травоядных животных или молодых более крупных травоядных животных. [ 61 ]
Поскольку внутренние механизмы вымерших существ не являются непостижимыми, большинство дискуссий фокусируются на гомеотермии и тахиметаболизме.
Оценка показателей метаболизма осложняется различием между скоростями во время отдыха и активным. Во всех современных рептилиях и большинстве млекопитающих и птиц максимальные показатели во время всеобъемлющей деятельности в 10-20 раз выше, чем минимальные скорости, находясь в состоянии покоя. Тем не менее, у нескольких млекопитающих эти показатели различаются в течение 70. Теоретически, для земельного позвоночного позвоночника можно было бы иметь скорость метаболизма рептилий в состоянии покоя и, похожие на птицу при работе. Тем не менее, животное с такой низкой скоростью покоя не сможет быстро расти. Огромные травоядные сауроподы , возможно, были настолько постоянно в поисках пищи, что их расходы на энергию были бы почти одинаковыми, независимо от того, были ли их скорости метаболизма высокими или низкими. [ 62 ]
Теории
[ редактировать ]Основные возможности - это: [ 35 ]
- Динозавры были хладнокровными, как современные рептилии, за исключением того, что большой размер многих стабилизировал бы температуру их тела.
- Они были теплыми, больше похожими на современных млекопитающих или птиц, чем на современные рептилии.
- Они не были ни хладнокровными, ни теплыми в современных терминах, но имели метаболизмы, которые отличались от и в некотором роде промежуточные между тем современными холодными и теплыми животными.
- Они включали животных с двумя или тремя из этих типов метаболизма.
Динозавры были около 150 миллионов лет, поэтому вполне вероятно, что разные группы развивали различные метаболизмы и терморегуляторные режимы, и что у некоторых развились различные физиологии от первых динозавров.
Если у всех или некоторых динозавров были промежуточные метаболизмы, у них могли быть следующие функции: [ 35 ]
- Низкие скорости метаболизма в состоянии покоя, что уменьшило бы количество пищи, которая им нужна, и позволит им использовать больше этой пищи для роста, чем животные с высокими показателями метаболизма в состоянии покоя.
- Инерционная гомеотермия
- Способность контролировать потерю тепла, расширяя и сокращая кровеносные сосуды, находящиеся под кожей, как это делают многие современные рептилии.
- Клубы с двумя частями, движимые четырехкамерованными сердцами.
- Высокая аэробная способность , позволяющая устойчивая активность.
Роберт Рейд предположил, что такие животные можно рассматривать как «неудачные эндотермы». Он предусматривал как динозавров, так и триасовых предков млекопитающих, проходящих через сцену с этими чертами. Млекопитающие были вынуждены стать меньше, так как архизавры стали доминировать в экологических нишах для средних и крупных животных. Их уменьшение размера сделало их более уязвимыми для потери тепла, потому что это увеличивало их соотношение площади поверхности к массе и, таким образом, заставило их увеличить внутреннюю генерирование тепла и, таким образом, стать полными эндотермами. С другой стороны, динозавры стали средними и очень крупными животными и, таким образом, смогли сохранить метаболизм «промежуточного». [ 35 ]
Структура кости
[ редактировать ]Арманд де Рикклес обнаружил хавердийские каналы в костях динозавров и утверждал, что у динозавров было свидетельство эндотермии. Эти каналы распространены у «теплокровных» животных и связаны с быстрым ростом и активным стилем жизни, потому что они помогают переработать кость, чтобы облегчить быстрый рост и восстановить повреждения, вызванные стрессом или травмами. [ 63 ] Плотная вторичная хавержийская кость, которая образуется во время ремоделирования, встречается во многих живых эндотермах, а также в динозаврах, птерозаврах и терапидах . Вторичные хаверсские каналы коррелируют с размером и возрастом, механическим стрессом и оборотом питательных веществ. Наличие вторичных каналов Хейверсии предполагает сопоставимый рост костей и продолжительность жизни у млекопитающих и динозавров. [ 64 ] Баккер утверждал, что присутствие фиброламеллярной кости (быстро производится и имела волокнистый, тканый внешний вид) у окаменелостей динозавров было свидетельством эндотермии. [ 9 ]
Однако в результате других, в основном более поздних исследований структура кости не считается надежным показателем метаболизма у динозавров, млекопитающих или рептилий:
- Кости динозавров часто содержат линии арестованного роста (лаги), образованные переменными периодами медленного и быстрого роста; На самом деле многие исследования подсчитывают кольца роста, чтобы оценить возраст динозавров. [ 55 ] [ 56 ] Образование колец роста обычно обусловлено сезонными изменениями температуры, и это сезонное влияние иногда рассматривается как признак медленного метаболизма и эктотермии. Но кольца роста встречаются у белых медведей и у млекопитающих, которые спят. [ 65 ] [ 66 ] Взаимосвязь между лагами и сезонной зависимостью роста остается нерешенной. [ 67 ]
- Фиброламеллярная кость довольно распространена у молодых крокодилов и иногда встречается у взрослых. [ 68 ] [ 69 ]
- Гавийская кость была найдена у черепах, крокодилов и черепах, [ 70 ] но часто отсутствует в маленьких птицах, летучих мышах, строги и грызунов. [ 69 ]
Тем не менее, De Ricqlès продолжал исследования кости структуры динозавров и архозавров . В середине 2008 года он стал соавтором статьи, в которой изучались образцы костей из широкого спектра архозавров, включая ранних динозавров, и пришел к выводу, что: [ 71 ]
- Даже самые ранние архозавриформные средства , возможно, были способны к очень быстрому росту, что говорит о том, что у них был довольно высокий уровень метаболизма. Хотя делать выводы о самых ранних архозавриформных из более поздних форм является сложным, поскольку видоспецифические различия в структуре кости и скорости роста очень вероятно, существуют исследовательские стратегии, которые могут минимизировать риск того, что такие факторы приведут к ошибкам в анализе.
- Архозавры разделились на три основные группы в триасе : орнитодираны , из которых развивались динозавры, оставались приверженными быстрому росту; Предки крокодилов приняли более типичные «медленные темпы роста»; И большинство других триасовых архозавров имели промежуточные темпы роста.
Остеогистологический анализ плотности и плотности сосудов, формы и площади остеоцитов завершил неавийские динозавры, а большинство архозавриформ (кроме протеросухуса , крокодилов и фитозавров ) сохранили тепло и содержали метаболические показатели, сходные с таковыми у территории и птиц. [ 72 ]
Скорость метаболизма, артериальное давление и поток
[ редактировать ]Эндотермы сильно полагаются на аэробный метаболизм и имеют высокие показатели потребления кислорода во время активности и отдыха. Кислород, требуемый тканями, переносится кровью, и, следовательно, скорость крови и кровяное давление, лежащее в основе теплокровных эндотерм, значительно выше, чем у хладнокровных эктотерм. [ 73 ] Можно измерить минимальное кровяное давление динозавров, оценив вертикальное расстояние между сердцем и верхней частью головы, потому что этот столб крови должен иметь давление на дне, равное гидростатическому давлению, полученному из плотности крови и гравитация. К этому давлению добавлено, что для перемещения крови через систему кровообращения. В 1976 году было отмечено, что из-за их роста у многих динозавров было минимальное кровяное давление в эндотермическом диапазоне, и что у них, должно быть легкие. [ 74 ] Не ясно, имели ли эти динозавры высокое кровяное давление просто для поддержки колонны крови или для поддержки высоких скоростей кровотока, требуемых эндотермией или обоими.
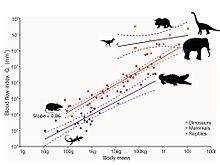
Тем не менее, недавний анализ крошечных отверстий в костях ископаемых ног динозавров обеспечивает датчик для скорости кровотока и, следовательно, скорость метаболизма. [ 75 ] Отверстия называются питательными отверстиями, а питательную артерию - это основной кровеносный сосуд, проходящий через внутреннюю часть кости, где она разветвляется в крошечные сосуды системы Хэверсийского канала . Эта система отвечает за замену старой кости новой костью, тем самым восстанавливая микробороды, которые встречаются естественным образом во время передвижения. Без этого восстановления микробористы накапливались, что приводит к переломам стресса и, в конечном счете, катастрофической недостаточностью кости. Размер отверстия питательных веществ обеспечивает индекс кровотока через него, в соответствии с уравнением Хаген-Пойзель . Конечно, размер также связан с размером тела животного, поэтому этот эффект удаляется путем анализа аллометрии . Индекс кровотока в питательном отверстии бедренных костей у живых млекопитающих увеличивается прямо пропорционально максимальной скорости метаболизма животных, измеряемых во время максимальной устойчивой локомоции. Индекс кровотока млекопитающих примерно в 10 раз больше, чем у эктотермических рептилий. Десять видов ископаемых динозавров из пяти таксономических групп выявляют индексы, даже выше, чем у млекопитающих, когда учитывается размер тела, что указывает на то, что они были очень активными, аэробными животными. Таким образом, высокая скорость кровотока, высокое кровяное давление, четырехкамерное сердце и устойчивый аэробный метаболизм соответствуют Эндотермия .
Темпы роста
[ редактировать ]Динозавры выросли от небольших яиц до нескольких тонн в весе относительно быстро. Естественная интерпретация этого заключается в том, что динозавры очень быстро превращали пищу в вес тела, что требует довольно быстрого метаболизма как для активного корма, так и для быстрого ассимиляции пищи. [ 76 ] Развивающиеся кости, обнаруженные у несовершеннолетних, явно пористые, что было связано с васкуляризацией и скоростью осаждения кости, все это предполагает, что скорости роста, близкие к темам, наблюдаемым у современных птиц.
Но предварительное исследование взаимосвязи между размером взрослого, скоростью роста и температурой тела пришло к выводу, что более крупные динозавры имели более высокую температуру тела, чем меньшие; По оценкам, апатозавр , самый большой динозавр в образце, имел температуру тела, превышающая 41 ° C (106 ° F), тогда как оценивались меньшие динозавры, имеют температуру тела около 25 ° C (77 ° F) [ 77 ] - Для сравнения нормальная температура тела человека составляет около 37 ° C (99 ° F). [ 78 ] [ 79 ] Основываясь на этих оценках, исследование пришло к выводу, что крупные динозавры были инерционными гомеотермами (их температура была стабилизирована их огромной массой), и что динозавры были эктотермическими (в разговорных словах «хладнокровные», потому что они не генерировали столько тепла, сколько млекопитающих млекопитающих когда не двигаться и не переваривать пищу). [ 77 ] Эти результаты согласуются с взаимосвязи между размерами динозавров и темпами роста (описано выше). [ 80 ] [ 81 ] Исследования Sauropodomorph Massospondylus и раннего Theropod Syntarsus ( Megapnosaurus ) выявляют темпы роста 3 кг/год и 17 кг/год, соответственно, намного медленнее, чем оцениваемые из Maiasaura и наблюдаемые у современных птиц. [ 82 ]
Соотношения изотопа кислорода в кости
[ редактировать ]Соотношение изотопов 16 O и 18 O в кости зависит от температуры, что кость образовалась при: чем выше температура, тем больше 16 O. Barrick and Sowers (1999) проанализировали коэффициенты изотопа в двух тероподах , которые жили в умеренных областях с сезонными изменениями температуры, тиранозавра (США) и Giganotosaurus (Аргентина): [ 83 ]
- Спинные позвонки от обоих динозавров не показали признаков сезонных изменений, что указывает на то, что оба сохраняли постоянную температуру ядра, несмотря на сезонные изменения температуры воздуха.
- Ребра и кости ног от обоих динозавров показали большую изменчивость температуры и более низкую среднюю температуру по мере увеличения расстояния от позвонков.
Баррик и ливни пришли к выводу, что оба динозавра были эндотермическими, но на более низких уровнях метаболика, чем современные млекопитающие, и что инерционная гомеотермия была важной частью их регуляции температуры в качестве взрослых. Их аналогичный анализ некоторых поздних меловых орнитишхов в 1996 году пришел к выводу, что эти животные показали сходную схему. [ 84 ]
Однако эта точка зрения была оспорена. Данные указывают на гомеотермию, но сами по себе не могут доказать эндотермию. Во -вторых, производство кости, возможно, не было непрерывным в областях, близких конечностях конечностей - в скелетных линиях аллозавра арестованного роста («лаги»; довольно похожие на кольца роста ) редки или отсутствуют в больших костях конечностей, но распространены в пальцах и пальцы ног. Хотя нет абсолютного доказательства того, что лаги связаны с температурой, они могут отметить время, когда конечности были настолько крутыми, что кости перестали расти. Если это так, то данные об соотношениях кислорода были бы неполными, особенно в моменты, когда конечности были самыми крутыми. Соотношения изотопа кислорода могут быть ненадежным методом оценки температур, если нельзя показать, что рост костей был одинаково непрерывным во всех частях животного. [ 35 ]
Коэффициенты хищника - PREY
[ редактировать ]- Хладнокровные хищники нуждаются в гораздо меньшем количестве пищи, чем теплокровные, поэтому данная масса добычи может поддерживать гораздо более холодные хищники, чем теплокровные.
- Соотношение общей массы хищников к охоте в общинах динозавров было гораздо больше похоже на коэффициент современных и недавних теплокровных сообществ, чем в недавних или ископаемых холоднокровных сообществах.
- Следовательно, хищные динозавры были теплыми. И поскольку самыми ранними динозаврами (например, Staurikosaurus , Herrerasaurus ) были хищники, все динозавры должны были быть теплыми.
Этот аргумент подвергся критике по нескольким основаниям и больше не воспринимается всерьез (следующий список критики далеко не исчерпывающий): [ 86 ] [ 87 ]
- Оценки весов динозавров сильно различаются, и даже небольшое изменение может иметь большое значение для рассчитанного соотношения хищника и писков.
- Его образец , возможно, не был представителем. Баккер получил свои цифры, считая образцы музея, но они имеют предвзятость к редким или особенно хорошо сохранившимся образцам, и не представляют то, что существует в ископаемых кроватях. Даже ископаемые кровати могут не точно представлять фактические популяции, например, меньшие и молодые животные имеют менее надежные кости и, следовательно, с меньшей вероятностью будут сохранены.
- Не существует опубликованных соотношений хищников и пищи для больших эктотермических хищников, потому что такие хищники очень редки и в основном встречаются только на довольно небольших островах. Большие эктотермические травоядные животные одинаково редки. Таким образом, Баккер был вынужден сравнивать соотношения хищников и перораторов млекопитающих с соотношениями рыб и беспозвоночных, где ожидания жизни намного короче, а другие различия также искажают сравнение.
- Концепция предполагает, что популяции хищников ограничены только доступностью добычи. Однако другие факторы, такие как нехватка участков гнездования, каннибализм или хищничество одного хищника на другом, могут удерживать популяции хищников ниже предела, налагаемого биомассой добычи, и это вводящее в заблуждение уменьшит отношение хищника и пищи.
- Например, экологические факторы могут ввести в заблуждение соотношение хищников и перо: хищник может охотиться только за некоторыми из присутствующих видов «добычи»; Болезнь, паразиты и голод могут убить некоторых добычных животных, прежде чем хищники получит шанс охотиться на них.
- Очень трудно точно заявить о том, что охотятся на что. Например, молодых травоядных могут быть охотятся ящерицами и змеями, в то время как взрослые охотятся млекопитающими. И наоборот, молодые из многих хищников живут в основном на беспозвоночных и переключаются на позвоночных по мере их роста.
Осанка и походка
[ редактировать ]В этом разделе нужны дополнительные цитаты для проверки . ( Сентябрь 2008 г. ) |

Термы динозавров были прямыми и удерживались под их телом, а не растягивались по бокам, как у ящериц и Ньютонов. Доказательством для этого являются углы поверхностей сустава и расположение прикреплений мышц и сухожилий на костях. Попытки представлять динозавров с растягивающимися конечностями приводят к существам с вывихнутыми бедрами, коленями, плечами и локтями. [ 88 ]
Ограничение перевозчика гласит, что воздушные дышащие позвоночные с двумя легкими, которые сгибают свои тела вбок во время передвижения, трудно двигаться и дышать одновременно. Это сильно ограничивает выносливость и заставляет их тратить больше времени на отдых, чем на перемещение. [ 89 ]
Растягивающиеся конечности требуют сгибания боком во время передвижения (за исключением черепах и черепах, которые очень медленные и чьи броне сохраняют их тела довольно жесткими). Однако, несмотря на ограничения переноса, растянутые конечности эффективны для существ, которые тратят большую часть своего времени, опираясь на свои животы и перемещаются только на несколько секунд за раз - потому что это соглашение сводит к минимуму затраты на энергию вставать и лежать.
Эпряженные конечности увеличивают затраты на вставание и лежа, но избегают ограничений перевозчика. Это указывает на то, что динозавры были активными животными, потому что естественный отбор предположил бы сохранение растягивающихся конечностей, если бы динозавры были вялыми и провели большую часть своего бодрствующего времени. Активный образ жизни требует метаболизма, который быстро восстанавливает энергетические принадлежности и разрушает отходы, которые вызывают усталость, то есть, это требует довольно быстрого метаболизма и значительной степени гомеотермии.
Кроме того, прямое осанка требует точного баланса, результата быстро функционирующей нервно -мышечной системы. Это предполагает эндотермический метаболизм, потому что эктотермическое животное будет неспособным ходить или бежать и, следовательно, уклоняться от хищников, когда его температура ядра была снижена. Другие доказательства эндотермии включают длину конечности (многие динозавры обладали сравнительно длинными конечностями) и двуногих, оба найденные сегодня только в эндотермах. [ 90 ] Многие двуногих динозавров обладали грейковыми костями с коротким бедром по сравнению с длиной теленка. Как правило, это является адаптацией к частому устойчивому бегу, характерна для эндотерм, которые, в отличие от эктотерм, способны производить достаточную энергию, чтобы предотвратить начало анаэробного метаболизма в мышцах. [ 91 ]
Баккер и Остром отметили, что у всех динозавров были прямые задних конечностей и что у всех четвероночных динозавров были прямые передние конечности; и что среди живых животных только эндотермические («теплые») млекопитающие и птицы имеют прямые конечности (острем признал, что случайная «высокая прогулка» крокодилов была частичным исключением). Баккер утверждал, что это было явным доказательством эндотермии в динозаврах, в то время как Остром считал ее убедительным, но не окончательным. [ 9 ] [ 92 ]
Исследование 2009 года подтвердило гипотезу о том, что эндотермия была широко распространена, по крайней мере, у более крупных неавийских динозавров, и что она была правдоподобно наследственной для всех динозавриформ, основанных на биомеханике бега, [ 93 ] Хотя также было высказано предположение, что эндотермия появилась гораздо раньше в эволюции архозавраморфа , возможно, даже предшествуя происхождению архозавриформ . [ 72 ]

Перья и нити
[ редактировать ]В настоящее время нет сомнений в том, что у многих видов динозавров были перья, в том числе Shuvuuia , Sinosauropteryx и Dilong (ранний тираннозавр ). [ 94 ] [ 27 ] [ 95 ] Они были истолкованы как изоляция и, следовательно, доказательства теплоты.
Но прямые, однозначные впечатления о перьях были обнаружены только у коелурозавров (которые включают птиц и тиранозавров, среди прочего), поэтому в настоящее время перья не дают нам информации о метаболизмах других основных групп динозавров, например, колофизиды , цератозавры , карнозавры , или сауроподы . Несмотря на нити, также присутствовал по крайней мере у некоторых орнитишхов, таких как Tianyulong , Kulindadromeus и Psittacosaurus , не только указывая на эндотермию в этой группе, но и то, что перья уже присутствовали в первом орниттодиране (последний общий предок динозавров и птероза). Их отсутствие в определенных группах, таких как Ankylosauria, может быть результатом подавления перьев. [ 96 ] Хотя нити впервые появились только в Coelurosauria в соответствии с реконструкциями максимального правдоподобия и что покровные структуры Psittacosaurus , Tianyulong и Kulindadromeus , независимо отлученных от филам, но это было за счет предположения примитивных предков птерозавра. [ 97 ]
Окаменечная кожа Carnotaurus ( абелисаврид и, следовательно, не коелурозавр) показывает непоколебимая, похожая на рептилий кожа с рядами ударов, [ 98 ] Но вывод о том, что Carnotaurus был обязательно без перья, подвергся критике, поскольку впечатления не покрывают все тело, находясь только в боковой области, но не в спине. [ 99 ] Взрослый карнотавр весил около 2 тонн, [ 100 ] И у млекопитающих такого размера и большего есть очень короткие, разреженные волосы или голые шкуры, поэтому, возможно, кожа Carnotaurus ничего не говорит нам о том, имели ли меньшие неэлурозаврские тероподы перья. тиранозавра Yutyrannus есть перья и весил 1,1 тонны. Известно, что у [ 100 ]
Управления кожи пелорозавра и других сауропод (динозавры с слоновыми телами и длинными шеями) показывают большие гексагональные чешуйки, а в некоторых сауроподах, таких как Saltasaurus , в коже были костяные тарелки. [ 101 ] Кожа цератопсийцев состояла из больших полигональных чешуйков, иногда с рассеянными круглыми пластинами. [ 102 ] «Мумифицируемые» остаются, а кожные впечатления от гасрозавридов показывают галечные весы. Маловероятно, что у анкилозавридов, таких как Euoplocephalus , имели изоляцию, поскольку большая часть их площади поверхности была покрыта коснированными ручками и тарелками. [ 103 ] Точно так же нет никаких доказательств изоляции в стегозаврах . Таким образом, изоляция и повышенная скорость метаболизма, стоящая за их развитием, могла быть ограничена тероподами или даже только подмножества теропод. [ Цитация необходима ] Отсутствие перьев или другого вида изоляции не указывает на эктотермию или низкие метаболизмы, как это наблюдалось в относительной безволосной мегафауне, свиньи, свиньи, человеческих детям и безволосной летучей мыши, совместимой с эндотермией. [ 100 ]
Полярные динозавры
[ редактировать ]Окаменелости динозавров были обнаружены в регионах, которые были близки к полюсам в соответствующие времена, особенно в юго -восточной Австралии , Антарктиде и на склоне Аляски северной . Нет никаких доказательств серьезных изменений в угле оси Земли , поэтому полярные динозавры и остальные эти экосистемы должны были бы справиться с теми же крайним изменением длины дня в течение года, который происходит в аналогичных широтах (вплоть до Полный день без тьмы летом и целый день без солнечного света зимой). [ 104 ]
Исследования ископаемой растительности показывают, что северный уклон Аляски имел максимальную температуру 13 ° C (55 ° F) и минимальную температуру 2 ° C (36 ° F) до 8 ° C (46 ° F) за последние 35 миллионов Годы мелового ( немного прохладнее Портленда , штат Орегон, но немного теплее, чем Калгари , Альберта). Несмотря на это, на северном склоне Аляски нет окаменелостей крупных холодных животных, таких как ящерицы и крокодильцы, которые были распространены в одно и то же время в Альберте, Монтане и Вайоминге . Это говорит о том, что, по крайней мере, некоторые неавийские динозавры были теплыми. [ 104 ] Было предложено, чтобы североамериканские полярные динозавры могли мигрировать в более теплые регионы, когда приближалась зима, что позволило бы им обитать Аляску летом, даже если они были холодными. [ 105 ] Но путешествие туда и Монтана, вероятно, использовала бы больше энергии, чем хладнокровные земельные позвоночные, которые производят через год; Другими словами, аляскинские динозавры должны были быть теплыми, независимо от того, мигрировали или остались на зиму. [ 106 ] Документ 2008 года о миграции динозавров Фила Р. Белла и Эрика Снайти предложила, чтобы большинство полярных динозавров, включая сауропод , анкилозавры и гипсилофодоны , вероятно , , хотя Hadrosaurid переписывались , тероподы Полем [ 107 ] [ 108 ]

Труднее определить климат Юго -Восточной Австралии, когда ископаемые кровати динозавров были установлены с 115 до 105 миллионов лет назад , к концу раннего мела: эти месторождения содержат доказательства вечной мерзлоты , ледяных клиньев и кожух Движение подземного льда, которое предполагает средние годовые температуры, варьировавшись от -6 ° C (21 ° F) до 5 ° C (41 ° F); Исследования изотопа кислорода этих отложений дают среднегодовую температуру 1,5 ° C (от 34,7 ° F) до 2,5 ° C (36,5 ° F). Однако разнообразие ископаемой растительности и большой размер некоторых ископаемых деревьев превышают то, что встречается в таких холодных условиях сегодня, и никто не объяснил, как такая растительность могла бы выжить при холодных температурах, предложенных физическими показателями-для сравнения Fairbanks , Аляска в настоящее время имеет среднюю годовую температуру 2,9 ° C (37,2 ° F). [ 104 ] Ежегодная миграция из юго -восточной Австралии было бы очень сложно для довольно мелких динозавров в таких, как Лиелинасавра , травоядное, около 60 сантиметров (2,0 фута) до 90 сантиметров (3,0 фута), потому что морские пути на север заблокировали проход к прогреве широты. [ 104 ] Образцы костей от Leellynasaura и Timimus , орнитомимида около 3,5 метров (11 футов) длиной и 1,5 метра (4,9 фута) в бедре, предположили, что у этих двух динозавров были разные способы выжить в холодных, темных зим: у образца Тмимуса были линии линий линий арестованный рост (лаги для короткого; аналогично кольцам роста ), и он мог спятить ; Но образец Leelynasaura не показал признаков лагов, поэтому он, возможно, оставался активным в течение зимы. [ 109 ] Исследование 2011 года, посвященное гипсолофодонту и костям Theropod, также пришло к выводу, что эти динозавры не спят в течение зимы, но оставались активными. [ 110 ]
Доказательства поведенческой терморегуляции
[ редактировать ]Некоторые динозавры, например, Спинозавр и Уранозавр , были на спине «паруса», поддерживаемых шипами, растущими со позвоночных . (Это также было правдой, кстати, для синапсид Dimetrodon .) Такие динозавры могли бы использовать эти паруса для:
- Возьмите тепло, греясь с «парусами» под прямым углом до солнечных лучей.
- Потерять тепло, используя «паруса» в качестве радиаторов, стоя в тени или, обращаясь непосредственно в сторону или от солнца.
Но это было очень маленькое меньшинство известных видов динозавров. Одной из распространенных интерпретаций пластин на спинах стегозавров является как теплообменники для терморегуляции, так как пластины заполнены кровеносными сосудами, которые теоретически могут поглощать и рассеивать тепло. [ 111 ]
Это могло бы работать для стегозавра с большими пластинами, такими как стегозавр , но другие стегозавры, такие как Wuerhosaurus , Tuojiangosaurus и Kentrosaurus, обладали гораздо меньшими пластинами с поверхностной площадью сомнительной ценности для терморегуляции. Тем не менее, идея стегозаврских пластин в качестве теплообменников была недавно подвергнута сомнению. [ 112 ]
Другие доказательства
[ редактировать ]Endothermy требует частого дыхания, что может привести к потере воды. У живых птиц и млекопитающих потеря воды ограничена вытягиванием влаги из выдыхаемого воздуха с покрываемыми слизи респираторными турбинатами, покрываемыми тканью костными листами в носовой полости. Несколько динозавров имеют обонятельные турбинаты, используемые для запаха, но ни один из них еще не был идентифицирован с респираторными турбинатами. [ 113 ]
Поскольку эндотермия допускает утонченный нервно -мышечный контроль, и поскольку мозговая материя требует большого количества энергии для поддержания, некоторые предполагают, что увеличение размера мозга указывает на повышение активности и, таким образом, эндотермию. Коэффициент энцефализации (уравнения) динозавров, показатель размер мозга, рассчитанная с использованием эндокастов мозга, варьируется на спектре от птиц, похожей на рептилий. Используя только эквалайзер, коелурозавры, по -видимому, были такими же активными, как живые млекопитающие, в то время как тероподы и орнитоподы падают где -то между млекопитающими и рептилиями, а другие динозавры напоминают рептилии. [ 113 ]
Исследование, опубликованное Роджером Сеймуром в 2013 году, добавило большую поддержку идеи, что динозавры были эндотермическими. После изучения крокодилов соленой воды Seymour обнаружил, что даже если их большие размеры могут обеспечить стабильные и высокие температуры тела, во время активности эктоболизм крокодила обеспечивал меньше аэробных способностей и генерировать только 14% от общей мышечной мощности аналогичного эндотермического млекопитающего перед полной усталостью Полем Сеймур полагал, что динозаврам нужно было бы быть эндотермическими, поскольку им потребовались бы лучшие аэробные способности и более высокая выработка электроэнергии, чтобы конкурировать и доминировать над млекопитающими как активными земельными животными в мезозойскую эпоху. [ 114 ]
Ранний метаболизм архозавра
[ редактировать ]В этом разделе нужны дополнительные цитаты для проверки . ( Сентябрь 2008 г. ) |
Похоже, что у самых ранних динозавров были особенности, которые составляют основу для аргументов для теплокровных динозавров, особенно возвышенных конечностей. Это поднимает вопрос: «Как динозавры стали теплыми?» Наиболее очевидными возможными ответами являются:
- «Их непосредственные предки ( архозавры ) были хладнокровными, а динозавры начали развивать теплокровные очень рано в своей эволюции». Это подразумевает, что динозавры развили значительную степень теплости за очень короткое время, возможно, менее 20 млн лет. теплокровной у предков млекопитающих эволюция Но [ 115 ] и, возможно, до появления волос около 164 метров года назад в середине юрского периода [ 116 ] ).
- «Непосредственные предки динозавров ( архозавры ) были, по крайней мере, довольно теплыми, и динозавры развивались дальше в этом направлении». ответ поднимает 2 проблемы: ( ) начала триасового и видов а Этот эволюция архозавров по -прежнему очень плохо изучена. Большое количество встречается с людей Ранняя speneri ); ( Б ) Крокодильцы развивались незадолго до динозавров и тесно связаны с ними, но хладнокровны (см. Ниже).
Крокодильцы представляют некоторые головоломки, если кто -то считает динозавров как активных животных с довольно постоянной температурой тела. Крокодильцы развивались незадолго до динозавров и, второго, являются птицами, являются самыми близкими живыми родственниками динозавров-но современные крокодильцы хладнокровны. Это поднимает несколько вопросов:
- Если динозавры в значительной степени были «теплыми», когда и как быстро развивалась теплая кровяность в их происхождении?
- Современные крокодильцы хладнокровны, но имеют несколько особенностей, связанных с теплыми крови. Как они приобрели эти функции?
Современные крокодилы хладнокровны, но могут двигаться с прямыми конечностями и имеют несколько функций, обычно связанных с теплыми клетками, потому что они улучшают кислородное снабжение животного: [ 117 ]
- 4-камерные сердца. У млекопитающих и птиц есть четырехкамерные сердца. Неконтрокодильские рептилии имеют трехчастотные сердца, которые менее эффективны, поскольку они позволяют оксигенированной и деоксигенированной крови смешиваться и, следовательно, посылают некоторую деоксигенированную кровь в организм, а не в легкие. Сердца современных крокодилов четыре камер, но меньше по сравнению с размером тела и работают при более низком давлении, чем у современных млекопитающих и птиц. У них также есть обход, который делает их функционально трехквартирными, когда они находятся под водой, сохранение кислорода. [ 118 ]
- диафрагма , которая помогает дыханию.
- Вторичное небо , которое позволяет животному есть и дышать одновременно.
- Печеночный поршневой механизм для перекачки легких. Это отличается от механизмов накапливания легких млекопитающих и птиц, но похоже на то, что некоторые исследователи утверждают, что обнаружили у некоторых динозавров. [ 20 ] [ 22 ]
Так почему же естественный отбор предпочитал эти особенности, которые важны для активных теплокровных существ, но мало очевидного применения для хладнокровных хищников засада, которые проводят большую часть своего времени, плавающего в воде или лежа на берегах рек?

В конце 1980-х годов было высказано предположение, что крокодильцы были первоначально активными, теплыми хищниками и что их предки архозавра были теплыми. [ 89 ] [ 117 ] Совсем недавно исследования развития показывают, что у крокодильских эмбрионов сначала развиваются полностью четырехкамерованные сердца, а затем развивают модификации, которые заставляют их сердца функционировать как трехкамерные под водой. Используя принцип, который онтогенез повторяет филогения , исследователи пришли к выводу, что у первоначальных крокодилов были полностью 4-камерные сердца и, следовательно, были теплыми, и что более поздние у крокодилов развивались шунтирование, когда они возвращались к хладнокровным хищникам для водных засад. [ 119 ] [ 120 ]
Более поздние исследования по структурам костей архозавра и их последствиях для темпов роста также свидетельствуют о том, что ранние архозавры имели довольно высокие показатели метаболизма и что триасовые предки крокодилов вернулись к более обычному «рептилированным» метаболическим показателям. [ 71 ]
Если эта точка зрения верна, развитие теплокровной архизавры (достигая своего пика у динозавров) и у млекопитающих потребовалось бы больше времени. Это также соответствовало бы ископаемым доказательствам:
- Самые ранние крокодиломорфы, например, Terrestrisuchus , были тонкими, длинноногими наземными хищниками.
- Эпряженные конечности появились довольно рано в эволюции архозавров , а у рауисучианцев очень плохо адаптированы для любой другой осанки. [ 121 ]
Смотрите также
[ редактировать ]- Классификация динозавров
- Ренессанс динозавров
- Эволюция динозавров
- Эволюционная физиология
- Список динозавров
- Происхождение птиц
- Аргентинское черно-белое тегу#теплуя кровь
Ссылки
[ редактировать ]- ^ Jump up to: а беременный Лукас, Спенсер Г. (1999). Динозавры: учебник (3 -е изд.). McGraw-Hill Companies, Inc. с. 3–9 . ISBN 978-0-07-303642-7 .
- ^ Фасковский DE, Weishampel DB (2005). «Теропода I: природа красная в зубах и когтях». В Фантовском де, Weishampel DB (ред.). Эволюция и вымирание динозавров (2 -е изд.). Издательство Кембриджского университета. С. 265–299. ISBN 978-0-521-81172-9 .
- ^ Колберт, эх; Cowles, RB; Богирт, CM (июнь 1947 г.). «Температурные допуски в американском аллигаторе и их привычки, эволюция и вымирание динозавров». Ежеквартальный обзор биологии . 22 (2): 145–146. doi : 10.1086/395712 .
- ^ Jump up to: а беременный Лукас, Спенсер Г. (2000). Динозавры: учебник (3 -е изд.). McGraw-Hill Companies, Inc. с. 1–3 . ISBN 978-0-07-303642-7 .
- ^ Торренс, Хью (1997). «Политика и палеонтология». В Фарлоу, Джеймс О.; Бретт-Сурман, Майкл К. (ред.). Полный динозавр . Блумингтон: издательство Университета Индианы. С. 175–190 . ISBN 978-0-253-33349-0 .
- ^ Бентон, Майкл Дж. (2000). «Краткая история палеонтологии динозавров». В Пол, Грегори С. (ред.). Научная американская книга динозавров . Нью -Йорк: Пресса Святого Мартина. С. 10–44. ISBN 978-0-312-26226-6 .
- ^ Баккер, RT (1968). «Превосходство динозавров». Открытие . 3 (2): 11–22.
- ^ Bakker, RT (1987). «Возвращение танцующих динозавров» . В Черкасе, SJ; Олсон, Е.К. (ред.). Динозавры прошлого и настоящего, вып. Я Университет Вашингтона Пресс. ISBN 978-0-295-96541-3 .
- ^ Jump up to: а беременный в дюймовый Баккер, Роберт Т. (1972). «Анатомические и экологические доказательства эндотермии у динозавров». Природа . 238 (5359): 81–85. Bibcode : 1972natur.238 ... 81b . doi : 10.1038/238081a0 . S2CID 4176132 .
- ^ Томас, RDK; Олсон, ЕС (1980). Холодный взгляд на теплых динозавров . Westview Press. ISBN 978-0-89158-464-3 .
- ^ Бентон, MJ (2005). Палеонтология позвоночных (3 -е изд.). Wiley-Blackwell. С. 221–223. ISBN 978-0-632-05637-8 .
- ^ Паладино, FV; О'Коннор, депутат; Spotila, JR (1990). «Метаболизм черепах из кожи, гигантотермии и терморегуляции динозавров». Природа . 344 (6269): 858–860. Bibcode : 1990natur.344..858p . doi : 10.1038/344858a0 . S2CID 4321764 .
- ^ Баррик, Re; Душ, WJ; Fischer, AG (1996). «Сравнение терморегуляции четырех орнитишских динозавров и варанидной ящерицы из двух мела двух лекарств: данные из изотопов кислорода». Палаис . 11 (4): 295–305. Bibcode : 1996palai..11..295b . doi : 10.2307/3515240 . JSTOR 3515240 .
- ^ Jump up to: а беременный в дюймовый и Норман, DB (апрель 2001 г.). «Кормление динозавра» . Энциклопедия наук о жизни . Джон Уайли и сыновья. doi : 10.1038/npg.els.0003321 . ISBN 978-0470016176 Полем Получено 10 сентября 2009 года .
- ^ Wings, O. (2007). «Обзор функции гастролита с последствиями для ископаемых позвоночных и пересмотренной классификации» (PDF) . Acta Palaeontologica Polonica . 52 (1): 1–16 . Получено 10 сентября 2008 года .
- ^ Крылья, О. и Сандер. PM (март 2007 г.). «Никакой желудочной мельницы в динозаврах Sauropod: новые данные из анализа массы и функции гастролита в страусах» . Труды Королевского общества б . 274 (1610): 635–640. doi : 10.1098/rspb.2006.3763 . PMC 2197205 . PMID 17254987 .
- ^ http://jeb.biologists.org/content/184/1/63.full.pdf [ только URL PDF ]
- ^ Jump up to: а беременный в дюймовый Ли, Эндрю Х.; Вернинг, Сара (2008). «Сексуальная зрелость в растущих динозаврах не соответствует моделям роста рептилии» . Труды Национальной академии наук . 105 (2): 582–587. Bibcode : 2008pnas..105..582L . doi : 10.1073/pnas.0708903105 . PMC 2206579 . PMID 18195356 .
- ^ Schweitzer, MH; Wittmeyer, JL; Хорнер, младший (2005). «Гендерная специфическая репродуктивная ткань в RATITES и Tyrannosaurus rex » (PDF) . Наука . 308 (5727): 1456–1460. Bibcode : 2005sci ... 308.1456s . doi : 10.1126/science.1112158 . PMID 15933198 . S2CID 30264554 .
- ^ Jump up to: а беременный в Рубен, JA; Джонс, ТД; Geist, NR; Хиллуениус, WJ (ноябрь 1997). «Структура легких и вентиляция у динозавров и ранних птиц». Наука . 278 (5341): 1267–1270. Bibcode : 1997sci ... 278.1267r . doi : 10.1126/science.278.5341.1267 .
- ^ Jump up to: а беременный Рубен, JA; Джонс, ТД; Geist, NR; Лейтч, А.; Hillenius, WJ (1997). «Вентиляция легких и газообмен у динозавров теропод». Наука . 278 (5341): 1267–1270. Bibcode : 1997sci ... 278.1267r . doi : 10.1126/science.278.5341.1267 .
- ^ Jump up to: а беременный Рубен, JA; Dal Sasso, C.; Geist, NR; Хиллуениус, WJ; Джонс, ТД; Signore, M. (январь 1999). «Функция легких и метаболическая физиология динозавров теропод» (PDF) . Наука . 283 (5401): 514–516. Bibcode : 1999sci ... 283..514r . doi : 10.1126/science.283.5401.514 . PMID 9915693 .
- ^ Рубен, JA; Джонс, ТД; Geist, NR (2003). «Дыхание и репродуктивная палеофизиология динозавров и ранних птиц» (PDF) . Физиол. Биохимия. Zool . 76 (2): 141–164. doi : 10.1086/375425 . HDL : 10211.1/1472 . PMID 12794669 . S2CID 222819 .
- ^ Hillenius, WJ & Ruben, JA (ноябрь -декабрь 2004 г.). «Эволюция эндотермии в наземных позвоночных: кто? Когда? Почему?». Физиологическая и биохимическая зоология . 77 (6): 1019–1042. doi : 10.1086/425185 . PMID 15674773 . S2CID 29300018 .
- ^ Хикс, JW & Farmer, CG (ноябрь 1997 г.). «Вентиляция легких и газообмен у динозавров теропод» . Наука . 278 (5341): 1267–1270. Bibcode : 1997sci ... 278.1267r . doi : 10.1126/science.278.5341.1267 .
- ^ Хикс, JW & Farmer, CG (сентябрь 1999 г.). «Потенциал газового обмена в легких рептилий: последствия для соединения динозавров и авиев». Физиология дыхания . 117 (2–3): 73–83. doi : 10.1016/s0034-5687 (99) 00060-2 . PMID 10563436 .
- ^ Jump up to: а беременный Currie, PJ & Chen, PJ. (Декабрь 2001 г.). «Анатомия sinosauropteryx prima из Liaoning, северо -восточный Китай» (PDF) . Канадский журнал наук о Земле . 38 (12): 1705–1727. Bibcode : 2001cajes..38.1705c . doi : 10.1139/cjes-38-12-1705 .
- ^ Jump up to: а беременный O'Connor, P. & Claessens, L. (июль 2005 г.). «Основная конструкция легочной птицы и вентиляция протекания у неавийских динозавров с теропод». Природа . 436 (7048): 253–256. Bibcode : 2005natur.436..253o . doi : 10.1038/nature03716 . PMID 16015329 . S2CID 4390587 .
- ^ Ведель, MJ (2003). «Позвоночная пневматичность, воздушные мешки и физиология динозавров Sauropod» . Палеобиология . 29 (2): 243–255. doi : 10.1666/0094-8373 (2003) 029 <0243: vpasat> 2,0.co; 2 . S2CID 46619244 . Полный текст в настоящее время онлайн на «Findarticles.com: позвоночная пневматичность, воздушные мешки и физиология динозавров Sauropod» . Палеобиология . 2003. и «Позвоночная пневматичность, воздушные мешки и физиология динозавров Sauropod» (PDF) . Архивировано из оригинала (PDF) 17 декабря 2008 года. Подробный анатомический анализ можно найти в Ведель, MJ (2003). сговор с Vertbral PDF . А 23 (2): 344–3 doi : 2003) 023 [0344: TOVPI] 10.1671/0272-4634 ( S2CID 58884062 .
- ^ Jump up to: а беременный Ведель, MJ (июнь 2006 г.). «Происхождение посткраниальной скелетной пневматичности у динозавров» (PDF) . Интегративная зоология . 1 (2): 80–85. doi : 10.1111/j.1749-4877.2006.00019.x . PMID 21395998 .
- ^ Найш, Д.; Martill, DM; Фрей, Э. (июнь 2004 г.). «Экология, систематика и биогеографические отношения динозавров, в том числе новый теропод, из формирования Сантаны (? Альбиан, ранний меховой) Бразилия». Историческая биология . 16 (2–4): 57–70. Citeseerx 10.1.1.394.9219 . doi : 10.1080/08912960410001674200 . S2CID 18592288 . Это также одна из нескольких тем, представленных в посте в блоге Naish, «Базальные динозавры тирана и мой питомец Мирамия » . - Примечание Мирисхия была коелурозаврой , который, по мнению Найш, был тесно связан с Compsognathus .
- ^ Jump up to: а беременный Sereno PC, Martinez RN, Wilson JA, Varricchio DJ, Alcober OA, et al. (Сентябрь 2008 г.). Кемп Т (ред.). «Свидетельство о птичье внутритарационные воздушные мешки в новом хищническом динозавре из Аргентины» . Plos один . 3 (9): E3303. Bibcode : 2008ploso ... 3.3303S . doi : 10.1371/journal.pone.0003303 . PMC 2553519 . PMID 18825273 .
- ^ Ведель, Мэтью (2007). «Что пневматичность говорит нам о« просауроподах »и наоборот» (PDF) . Специальные статьи в палеонтологии . 77 : 207–222. Архивировано из оригинала (PDF) 5 июля 2008 года . Получено 31 октября 2007 года .
- ^ Mallison, H. (2010). «Digital Plateosaurus II: оценка диапазона движения конечностей и позвоночного столба и предыдущих реконструкций с использованием цифрового скелетного крепления» . Acta Palaeontologica Polonica . 55 (3): 433–458. doi : 10.4202/app.2009.0075 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Рейд, Рех (1997). «Динозаврская физиология: случай для« промежуточных »динозавров». В Фарлоу, Джо; Бретт-Сурман, MK (Eds.). Полный динозавр . Блумингтон: издательство Университета Индианы. С. 449–473. ISBN 978-0-253-33349-0 Полем Получено 12 сентября 2008 года .
- ^ Паладино, FV; Spotila, Jr; Додсон, П. (1997). «План гиганта: моделирование физиологии крупных динозавров». В Фарлоу, Джо; Бретт-Сурман, MK (Eds.). Полный динозавр . Блумингтон, Индиана: издательство Университета Индианы. С. 491–504. ISBN 978-0-253-21313-6 .
- ^ Ward, Peter (2006) Из воздуха: динозавры, птицы и древние страницы атмосферы Земли 159–198, Национальная академическая пресса. ISBN 9780309141239 .
- ^ Codd, Jr; Мэннинг, PL; Норелл, Массачусетс; Перри, SF (январь 2008 г.). «Птиеподобная механика для дыхания в динозаврах манирапторов» . Труды Королевского общества б . 275 (1631): 157–161. doi : 10.1098/rspb.2007.1233 . PMC 2596187 . PMID 17986432 . Краткое описание новостей в «Почему у динозавров было« дыхание птицы » . 7 ноября 2007 г.
- ^ Щекотка, pg; Ennos, Ar; Леннокс, Ле; Перри, SF; Codd, Jr (ноябрь 2007 г.). «Функциональная значимость невидимых процессов у птиц» . Журнал экспериментальной биологии . 210 (Pt 22): 3955–3961. doi : 10.1242/jeb.008953 . PMID 17981863 .
- ^ Фишер, PE; Рассел, да; Stoskopf, Mk; Баррик, Re; Молоток, м.; Kuzmitz AA (апрель 2000 г.). «Сердечно -сосудистые доказательства среднего или более высокого уровня метаболизма у орнитишского динозавра». Наука . 288 (5465): 503–505. Bibcode : 2000sci ... 288..503f . doi : 10.1126/science.288.5465.503 . PMID 10775107 . Но обратите внимание, что главный предмет этой статьи заключается в том, что ископаемое предоставило убедительные доказательства 4-камерного сердца, которое не является широко распространенным.
- ^ Рубен, JA; Хиллуениус, WJ; Geist, NR; Лейтч, А.; Джонс, ТД; Керри, PJ; Хорнер, младший; ESPE, G. (август 1996 г.). «Метаболический статус некоторых поздних меловых динозавров» (PDF) . Наука . 273 (5279): 1204–1207. Bibcode : 1996sci ... 273.1204R . doi : 10.1126/science.273.5279.1204 . S2CID 84693210 .
- ^ Рубен, JA & Jones, TD (2000). «Селективные факторы, связанные с происхождением меха и перьев» . Американский зоолог . 40 (4): 585–596. doi : 10.1093/icb/40.4.585 .
- ^ Банг, BG (1966). "Обонятельный аппарат procellariforms" Acta Anatomica 65 (1): 391–4 Doi : 10.1159/ 000142884 5965973PMID
- ^ Банг, BG (1971). «Функциональная анатомия обонятельной системы в 23 порядка птиц». Acta Anatomica . 79. 79 : 1–76. doi : 10.1159/isbn.978-3-318-01866-0 . ISBN 978-3-8055-1193-3 Полем PMID 5133493 .
- ^ Скотт, Дж. Х. (1954). «Регулирующая тепло функция слизистой оболочки назала». Журнал ларинологии и отологии . 68 (5): 308–317. doi : 10.1017/s0022215100049707 . PMID 13163588 . S2CID 32082759 .
- ^ Coulombe, Hn; Сэм Х. Риджуэй; Sh; Эванс, мы (1965). «Респираторный обмен водой у двух видов свинья». Наука . 149 (3679): 86–88. Bibcode : 1965sci ... 149 ... 86c . doi : 10.1126/science.149.3679.86 . PMID 17737801 . S2CID 38947951 .
- ^ Witmer, LM (август 2001 г.). «Положение ноздри у динозавров и других позвоночных и ее значение для носовой функции». Наука . 293 (5531): 850–853. Citeseerx 10.1.1.629.1744 . doi : 10.1126/science.1062681 . PMID 11486085 . S2CID 7328047 .
- ^ «Исследование дыхания динозавров показывает, что носы усиливают пахнущий и охлажденный мозг» .
- ^ Фишер, Пол Э.; Рассел, Дейл А.; Stoskopf, Michael K.; Баррик, Риз Э.; Хаммер, Майкл; Кузмитц, Эндрю А. (апрель 2000 г.). «Сердечно -сосудистые доказательства среднего или более высокого уровня метаболизма у орнитишского динозавра». Наука . 288 (5465): 503–505. Bibcode : 2000sci ... 288..503f . doi : 10.1126/science.288.5465.503 . PMID 10775107 .
- ^ Роу, Тимоти; McBride, Earle F.; Серено, Пол С. (февраль 2001 г.). «Технический комментарий: динозавр с камнем сердца» . Наука . 291 (5505): 783a. doi : 10.1126/science.291.5505.783a . PMID 11157158 .
- ^ Рассел, Дейл А .; Фишер, Пол Э.; Баррик, Риз Э.; Стоскопф, Майкл К. (февраль 2001 г.). «Ответ: динозавр с камнем» . Наука . 291 (5505): 783a. doi : 10.1126/science.291.5505.783a . PMID 11157158 .
- ^ Chinsamy, Anusuya; и Hillenius, Willem J. (2004). «Физиология неавийских динозавров». Динозаврия , 2 -й. 643–659.
- ^ Карпентер, К.; Хирш, KF; Хорнер, младший (1994). "Введение". В Карпентере, К.; Хирш, KF; Хорнер, младший (ред.). Яйца динозавров и дети . Издательство Кембриджского университета. ISBN 978-0-521-56723-7 .
- ^ Эриксон, GM; Карри Роджерс, К.; Еерби, СА (июль 2001 г.). «Модели роста динозавров и быстрые темпы роста птиц». Природа . 412 (6845): 429–433. Bibcode : 2001natur.412..429e . doi : 10.1038/35086558 . PMID 11473315 . S2CID 4319534 . (Ошибка: doi : 10.1038/nature16488 , PMID 26675731 , реверкция Watch ) Примечание Кристина Роджерс также опубликовала статьи под ее девичьей фамилией Кристиной Карри.
- ^ Jump up to: а беременный Карри, К.А. (1999). «Онтогенетическая гистология апатозавра (Dinosauria: Sauropoda): новое понимание темпов роста и долголетия» . Журнал палеонтологии позвоночных . 19 (4): 654–665. Bibcode : 1999jvpal..19..654c . doi : 10.1080/02724634.1999.10011179 . Архивировано из оригинала 15 июля 2010 года.
- ^ Jump up to: а беременный в Эриксон, GM; Маковицки, PJ; Керри, PJ; Норелл, Массачусетс; Еерби, СА; Брочу, Калифорния (август 2004 г.). «Гигантизм и сравнительные параметры жизни динозавров тиранозавридов» (PDF) . Природа . 430 (7001): 772–775. Bibcode : 2004natur.430..772e . doi : 10.1038/nature02699 . PMID 15306807 . S2CID 4404887 . (Ошибка: doi : 10.1038/nature16487 , PMID 26675726 , реверкция Watch )
- ^ Хорнер, младший; Падиан, К. (сентябрь 2004 г.). «Динамика возраста и роста тиранозавра Рекса» . Труды Королевского общества б . 271 (1551): 1875–1880. doi : 10.1098/rspb.2004.2829 . PMC 1691809 . PMID 15347508 .
- ^ Купер, Ln; Ли, ах; Конус, ML; Хорнер, младший (август 2008 г.). «Относительные темпы роста динозавров хищников и добычи отражают эффекты хищничества» . Труды Королевского общества б . 275 (1651): 2609–2615. doi : 10.1098/rspb.2008.0912 . PMC 2605812 . PMID 18682367 .
- ^ Эриксон, GM; Керри, PJ; Inouye, Bd; Винн, А.А. (июль 2006 г.). «Жизненные таблицы тиранозавров: пример биологии неавийской биологии населения динозавров». Наука . 313 (5784): 213–217. Bibcode : 2006sci ... 313..213e . doi : 10.1126/science.1125721 . PMID 16840697 . S2CID 34191607 .
- ^ McGowan, C. (1991). Динозавры, плеск и морские драконы . Гарвардский университет издательство. С. 133–135 . ISBN 978-0-674-20769-1 .
- ^ Mackie, RI (2002). «Мутационное ферментативное расщепление в желудочно -кишечном тракте: разнообразие и эволюция» . Интегративная и сравнительная биология . 42 (2): 319–326. doi : 10.1093/ICB/42.2.319 . PMID 21708724 .
- ^ Пол, Г.С. (1998). «Правило Коупа» . В Карпентере, К.; Fraser, N.; Chure, D.; Киркленд, Джи (ред.). Моррисон Симпозиум Симпозиум Симпозиум: специальный выпуск журнала Modern Geology . Тейлор и Фрэнсис. п. 211. ISBN 978-90-5699-183-8 Полем Получено 12 сентября 2008 года .
- ^ Ricqles, AJ de. (1974). Эволюция эндотермии: гистологические данные . Эволюционная теория 1: 51–80
- ^ Fastovsky & Weishampel 2009, с.258.
- ^ Чинсми, а.; Rich, T.; Викерс-Рич, П. (1998). «Полярная гистология кости динозавров» . Журнал палеонтологии позвоночных . 18 (2): 385–390. Bibcode : 1998jvpal..18..385c . doi : 10.1080/02724634.1998.10011066 . Архивировано из оригинала 15 июля 2010 года.
- ^ Klevezal, GA ; Мина, MV; Oreshkin, Av (1996). Записывающие структуры млекопитающих. Определение возраста и реконструкция истории жизни . CRC Press. ISBN 978-90-5410-621-0 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Fastovsky & Weishampel 2009, с.260.
- ^ Enlow, DH (1963). Принципы ремоделирования костей. Отчет о процессах роста и реконструкции пост-повторного роста в длинных костях и нижней челюсти . Спрингфилд, Иллинойс: CC Thomas.
- ^ Jump up to: а беременный Рейд, Рех (1984). «Первичная физиология костей и динозавров». Геологический журнал . 121 (6): 589–598. Bibcode : 1984geom..121..589r . doi : 10.1017/s0016756800030739 . S2CID 128981835 .
- ^ Рейд, Рех (1997). «Как росли динозавры». В Фарлоу, Джо; Бретт-Сурман, MK (Eds.). Полный динозавр . Блумингтон: издательство Университета Индианы. С. 403–413 . ISBN 978-0-253-33349-0 .
- ^ Jump up to: а беременный De Ricqlès, A.; Падиан, К . ; Knoll, F.; Хорнер, младший (апрель -июнь 2008 г.). «О происхождении высоких темпов роста у архозавров и их древних родственников: комплементарные гистологические исследования триасовых архозавриформ и проблему« филогенетического сигнала »в гистологии кости». Анналес де палеонтологии . 94 (2): 57–76. Bibcode : 2008anpal..94 ... 57d . doi : 10.1016/j.annpal.2008.03.002 . Аннотация также онлайн на «Происхождение высоких темпов роста у архозавров» . 31 мая 2008 г. Получено 3 июня 2008 года .
- ^ Jump up to: а беременный Legendre, Lucas J.; Генард, Гийом; Бота-Бринк, Дженнифер; Кубо, Хорхе (1 ноября 2016 г.). «Палеоистологические доказательства приколов высокой метаболизма у архозавров» . Систематическая биология . 65 (6): 989–996. doi : 10.1093/sysbio/syw033 . ISSN 1063-5157 . PMID 27073251 .
- ^ Сеймур, Роджер С.; Bennett-Stamper, Christina L.; Джонстон, Соня Д.; Перевозчик, Дэвид Р.; Григг, Гордон С. (2004). «Свидетельство для эндотермических предков крокодилов в стволе эволюции архозавра» (PDF) . Физиологическая и биохимическая зоология . 77 (6): 1051–1067. doi : 10.1086/422766 . HDL : 2440/1933 . PMID 15674775 . S2CID 10111065 .
- ^ Сеймур, Роджер С. (1976). «Динозавры, эндотермия и кровяное давление». Природа . 262 (5565): 207–208. Bibcode : 1976natur.262..207s . doi : 10.1038/262207a0 . S2CID 4157084 .
- ^ Сеймур, Роджер С.; Смит, Сара Л; Белый, Крейг Р.; Хендерсон, Дональд М.; Schwarz Wings, Daniela (2012). «Кровоток к длинным костям указывает на метаболизм активности у млекопитающих, рептилий и динозавров» . Труды Королевского общества б . 279 (1728): 451–456. doi : 10.1098/rspb.2011.0968 . PMC 3234558 . PMID 21733896 .
- ^ Пол, Г.С. (1988). Хищные динозавры мира . Нью -Йорк: Саймон и Шустер. п. 155 ISBN 978-0-671-61946-6 .
- ^ Jump up to: а беременный Gillooly, JF; Аллен, AP; Чарнова, Эл (август 2006 г.). «Окаменелости динозавров предсказывают температуру тела» . PLOS Биология . 4 (8): E248. doi : 10.1371/journal.pbio.0040248 . PMC 1489189 . PMID 16817695 . Есть менее техническое резюме в Гросс, Л. (август 2006 г.). «Математика и окаменелости разрешают дебаты по метаболизму динозавров» . PLOS Биология . 4 (8): E255. doi : 10.1371/journal.pbio.0040255 . PMC 1489188 . PMID 20076616 .
- ^ Mackowiak, PA; СС Вассерман; ММ Левин (23 сентября 1992 г.). «Критическая оценка 98,6 ° F (37,0 ° C), верхний предел нормальной температуры тела и другое наследие Карла Рейнхольда Августа Вундерлиха». Джама . 268 (12): 1578–1580. doi : 10.1001/Jama.1992.03490120092034 . PMID 1302471 .
- ^ Sund-Levander, Märtha; Форсберг, Кристина; Варен, Лис Карин (2002). «Нормальная пероральная, ректальная, барабанная и подмышечная температура тела у взрослых мужчин и женщин: систематический обзор литературы». Скандинавский журнал заботливых наук . 16 (2): 122–128. doi : 10.1046/j.1471-6712.2002.00069.x . PMID 12000664 .
- ^ Эриксон, Грегори М.; Роджерс, Кристина Карри; Йерби, Скотт А. (2001). «Модели роста динозавров и быстрые темпы роста птиц». Природа . 412 (6845): 429–433. Bibcode : 2001natur.412..429e . doi : 10.1038/35086558 . PMID 11473315 . S2CID 4319534 . (Ошибка: doi : 10.1038/nature16488 , PMID 26675731 , реверкция Watch )
- ^ Карри, Кристина А. (1999). «Онтогенетическая гистология апатозавра (Dinosauria: Sauropoda): новое понимание темпов роста и долголетия». Журнал палеонтологии позвоночных . 19 (4): 654–665. Bibcode : 1999jvpal..19..654c . doi : 10.1080/02724634.1999.10011179 .
- ^ Фасковский, Дэвид Э.; Weishampel, Dvid B. (2009). Динозавры: краткая естественная история (PDF) . п. 258
- ^ Barrick, Re & Sowers, WJ (1999). «Термофизиология и биология гиганотозавра : сравнение с тиранозавром » . Palaeontologia Electronica . 2 (2). Архивировано из оригинала 17 мая 2011 года . Получено 10 сентября 2008 года .
- ^ Баррик, Re; Душ, WJ; Фишер, А.Г. (август 1996 г.). «Сравнение терморегуляции четырех орнитишских динозавров и варанидной ящерицы из двух мела двух лекарств: данные из изотопов кислорода». Палаис . 11 (4): 295–305. Bibcode : 1996palai..11..295b . doi : 10.2307/3515240 . JSTOR 3515240 .
- ^ Баккер, RT (сентябрь 1974 г.). «Биоэнергетика динозавра - ответ Беннетту и Далзеллу, а также Feduccia». Эволюция 28 (3): 497–503. doi : 10.2307/2407178 . JSTOR 2407178 . PMID 28564852 .
- ^ Фасковский, де; Weishampel, DB; Sibbick, J. (2005). Эволюция и вымирание динозавров . Издательство Кембриджского университета. ISBN 978-0-521-81172-9 .
- ^ Фарлоу, Джо (1980). «Коэффициенты биомассы хищников/добычи, общественные пищевые сети и физиология динозавров» (PDF) . В Томасе, RDK; Олсон, Е.К. (ред.). Холодный взгляд на теплых динозавров . Boulder, CO: Американская ассоциация развития науки. С. 55–83. Архивировано из оригинала (PDF) 17 декабря 2008 года.
- ^ Это было признано не позднее 1909 года: «Доктор Холланд и обширные сауропод» . Архивировано из оригинала 12 июня 2011 года. Аргументы и многие изображения также представлены в Десмонд А. (1976). Горячие кровные динозавры . Doubleday. ISBN 978-0-385-27063-2 .
- ^ Jump up to: а беременный Carrier, DR (1987). «Эволюция локомоторной выносливости в тетраподах: обход механического ограничения». Палеобиология . 13 (3): 326–341. Bibcode : 1987pbio ... 13..326c . doi : 10.1017/s0094837300008903 . JSTOR 2400734 . S2CID 83722453 .
- ^ Fastovsky & Weishampel 2009, с.251.
- ^ Fastovsky & Weishampel 2009, с.252.
- ^ Ostrom, JH (1980). «Доказательства эндотермии у динозавров» (PDF) . В Томасе, RDK; Олсон, Е.К. (ред.). Холодный взгляд на теплых динозавров . Boulder, CO: Американская ассоциация развития науки. С. 82–105. Архивировано из оригинала (PDF) 27 мая 2008 года.
- ^ Понцер, Х. .; Аллен, В.; Хатчинсон, Дж. .; Фарке, А.А. (2009). Фарке, Эндрю Аллен (ред.). «Биомеханика бега указывает на эндотермию в двуногих динозаврах» . Plos один . 4 (11): E7783. Bibcode : 2009ploso ... 4.7783p . doi : 10.1371/journal.pone.0007783 . PMC 2772121 . PMID 19911059 .
- ^ Schweitzer, MH; Ja watt; Р. Авчи; Л. Кнапп; Л. Чиаппе; М. Норелл; М. Маршалл. (Июль 1999). «Бета-кератин специфичная иммунологическая реакционная способность в перьев, подобных структурам мелового Альварессаврида, Shuvuuia deserti » . Журнал экспериментальной зоологии . 285 (2): 146–157. doi : 10.1002/(SICI) 1097-010X (19990815) 285: 2 <146 :: AID-JEZ7> 3.0.CO; 2-A . PMID 10440726 . Архивировано с оригинала 10 декабря 2012 года.
- ^ Xu x.; Норелл, Массачусетс; Куанг х.; Ван Х.; Zhao Q. & Jia C. (2004). «Базальные тиранозавра из Китая и доказательства протофиков у тиранозавра» (PDF) . Природа . 431 (7009): 680–684. Bibcode : 2004natur.431..680x . doi : 10.1038/nature02855 . PMID 15470426 . S2CID 4381777 .
- ^ Бентон, Майкл Дж.; Dhouailly, Danielle; Цзян, Баою; Макнамара, Мария (1 сентября 2019 г.). «Раннее происхождение перьев» . Тенденции в экологии и эволюции . 34 (9): 856–869. doi : 10.1016/j.tree.2019.04.018 . HDL : 10468/8068 . ISSN 0169-5347 . PMID 31164250 . S2CID 174811556 .
- ^ Барретт, Пол М.; Эванс, Дэвид С.; Кампионе, Николас Э. (30 июня 2015 г.). «Эволюция эпидермальных структур динозавров» . Биологические письма . 11 (6): 20150229. DOI : 10.1098/rsbl.2015.0229 . PMC 4528472 . PMID 26041865 .
- ^ Бонапарт, JF; Novas, Ee; Coria, RA (1990). « Carnotaurus Sastrei Bonaparte, рогатый, слегка построенный карнозавр из среднего мела Патагонии» . Музей естественной истории вклад округа Лос -Анджелес в науке . 416 : 1–42. Архивировано из оригинала 10 мая 2008 года.
- ^ Доктор философии, Андреа Кау (1 марта 2016 г.). «Тероподида: Мити и постмодерн » . Therpreada . Получено 25 ешь 2020 .
- ^ Jump up to: а беременный в Пол, Грегори С. (2016). Полевой гид Принстона по динозаврам - 2 -е издание . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. ISBN 978-0-691-16766-4 .
- ^ Czerkas, SA (1994). Локли, мг; Dos Santos, VF; Мейер, Калифорния; Охота, AP (Eds.). «История и интерпретация впечатлений от кожи Sauropod». Гайя . 10 Лиссабон, Португалия.
- ^ Dodson, P. & Barlowe, WD (1996). Рогатые динозавры: естественная история . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. ISBN 978-0-691-05900-6 Полем Смотрите также изображение в «Цератопсская кожа» . Архивировано из оригинала 3 февраля 2014 года.
- ^ Gosline, A. (16 ноября 2004 г.). «Bulletproof -Bulletpronation Armor" Dinosaurs ' . New Scientist.com Service.
- ^ Jump up to: а беременный в дюймовый Рич, Т.Г.; Викерс-Рич, П.; Gangloff, RA (февраль 2002 г.). «Полярные динозавры» . Наука . 295 (5557): 979–980. doi : 10.1126/science.1068920 . PMID 11834803 . S2CID 28065814 . Смотрите также Vickers-Rich, P. & Rich, TH (2004). «Динозавры Антарктики» (PDF) . Scientific American. Архивировано из оригинала (PDF) 17 мая 2008 г. и Фиорильо, АР (2004). «Динозавры арктической Аляски» (PDF) . Scientific American . 291 (6): 84–91. Bibcode : 2004sciam.291f..84f . doi : 10.1038/Scientificamerican1204-84 . PMID 15597984 . Архивировано из оригинала (PDF) 17 мая 2008 года.
- ^ Хоттон, Н. (1980). «Альтернатива эндотермии динозавров: счастливые странники». В Томасе, RDK; Олсон, Е.К. (ред.). Холодный взгляд на теплых динозавров . Boulder, CO: Американская ассоциация развития науки. С. 311–350. ISBN 978-0-89158-464-3 .
- ^ Пол, Г.С. (1988). Хищные динозавры мира . Нью -Йорк: Саймон и Шустер. С. 160–161 . ISBN 978-0-671-61946-6 .
- ^ Белл, Фил Р.; Snively, E. (2008). «Полярные динозавры на параде: обзор миграции динозавров» . Алхринге . 32 (3): 271–284. doi : 10.1080/03115510802096101 .
- ^ Ллойд, Робин (4 декабря 2008 г.). «Полярные динозавры пережили холодные темные зимы» . LivesCience.com . Imaginova . Получено 11 декабря 2008 года .
- ^ Чинсми, а.; Рич, Т.Г.; Викерс-Рич, П. (1998). «Полярная гистология кости динозавров». Журнал палеонтологии позвоночных . 18 (2): 385–390. Bibcode : 1998jvpal..18..385c . doi : 10.1080/02724634.1998.10011066 . JSTOR 4523908 . Смотрите также Лесли, М. (декабрь 2007 г.). «Принимая температуру динозавра» . Смитсоновский журнал. [ Постоянная мертвая ссылка ]
- ^ Вудворд, Холли Н.; Рич, Томас Х.; Чинсами, Анусуя; Викерс-Рич, Патриция (2011). «Динамика роста полярных динозавров Австралии» . Plos один . 6 (8): E23339. BIBCODE : 2011PLOSO ... 6233339W . doi : 10.1371/journal.pone.00233339 . PMC 3149654 . PMID 21826250 .
- ^ de Bufrenil, v.; Фарлоу, Джо; Де Риклс, А. (1986). «Рост и функция пластин стегозавра : доказательства гистологии кости». Палеобиология . 12 (4): 459–473. Bibcode : 1986pbio ... 12..459d . doi : 10.1017/s0094837300003171 . JSTOR 2400518 . S2CID 89139852 .
- ^ «Стегозавры пластины и шипы только для внешнего вида» . 18 мая 2005 г. Архивировано с оригинала 27 сентября 2011 года.
- ^ Jump up to: а беременный Fastovsky & Weishampel 2009, с.255.
- ^ Сеймур Роджер С. (5 июля 2013 г.). «Максимальная аэробная и анаэробная выработка электроэнергии в крупных крокодилах по сравнению с млекопитающими: последствия для гигантотермии динозавров» . Plos один . 8 (7): E69361. BIBCODE : 2013PLOSO ... 869361S . doi : 10.1371/journal.pone.0069361 . PMC 3702618 . PMID 23861968 .
- ^ Kermack, DM & Kermack, KA (1984). Эволюция персонажей млекопитающих Лондон: Croom Helm Captain Szabo Publishers. П. 149. ISBN 978-0-7099-1534-8 .
- ^ Самое раннее явное свидетельство о волосах или мехе - ископаемые из Касторокауда , с 164 метра лет назад в середине юрского периода , Ji, Q.; Luo, ZX; Юань, CX; Tabrum, AR (февраль 2006 г.). «Плавательная млекопитающая из средней юры и экоморфологическая диверсификация ранних млекопитающих» . Наука . 311 (5764): 1123–7. Bibcode : 2006sci ... 311.1123j . doi : 10.1126/science.1123026 . PMID 16497926 . S2CID 46067702 . Смотрите также новостной пункт на «Юрский» Бивер »найден; переписывает историю млекопитающих» . Архивировано из оригинала 3 марта 2006 года. -х годах можно было быть свидетельство о волосах в ранних С 1950-х годов в 1950 цинодонах, таких как Тринаксодон : Бринк, как (1955). «Исследование скелета Diademodon ». Palaeontologia Africana . 3 : 3–39. и Кемп, Т.С. (1982). Млекопитающие рептилии и происхождение млекопитающих . Лондон: академическая пресса. п. 363 . ISBN 978-0-12-404120-2 Полем Но отверстие (маленькие туннели) в костях Cynodont Snout в лучшем случае является неоднозначным доказательством, поскольку аналогичные отверстия находятся в нескольких живых рептилиях: Estes, R. (1961). «Анатомия черепа рептилий Cynodont thrinaxodon liorhinus ». Бюллетень Музея сравнительной зоологии (1253): 165–180. и Рубен, JA & Jones, TD (2000). «Селективные факторы, связанные с происхождением меха и перьев» . Американский зоолог . 40 (4): 585–596. doi : 10.1093/icb/40.4.585 .
- ^ Jump up to: а беременный McGowan, C. (1991). Динозавры, плеск и морские драконы . Гарвардский университет издательство. С. 151–152 . ISBN 978-0-674-20769-1 .
- ^ «Секрет крокодильного сердца» . Австралийская вещательная корпорация .
- ^ Саммерс, AP (2005). «Эволюция: сердечные крокодилы» . Природа . 434 (7035): 833–834. Bibcode : 2005natur.434..833s . doi : 10.1038/434833a . PMID 15829945 . S2CID 4399224 .
- ^ Seymour, Rs; Bennett-Stamper, Cl; Джонстон, SD; Перевозчик, доктор; Григг, GC (2004). «Свидетельство для эндотермических предков крокодилов в стволе эволюции архозавра» (PDF) . Физиологическая и биохимическая зоология . 77 (6): 1051–1067. doi : 10.1086/422766 . HDL : 2440/1933 . PMID 15674775 . S2CID 10111065 . См. Также объяснение этого, с полезными иллюстрациями в Майерс, PZ (19 апреля 2005 г.). "Горячие крокодилы?" Полем Архивировано из оригинала 16 февраля 2008 года.
- ^ Хатчинсон, младший (март -апрель 2006 г.). PDF ( ) Рендус учетные записи Palevol 5 (3–4): 519–530. Bibcode : 2006crpal . два 10.1016/j.crpv.2005.09.002:
Внешние ссылки
[ редактировать ]- Термофизиология и биология Giganotosaurus: сравнение с тиранозавром, архивным 17 мая 2011 года на Wayback Machine от Re Barrick and WJ Sowers (1999)
- Сердце динозавра обнаружено
- Эволюция крокодила без сердечного пласта
физиологии Типы | ||
|---|---|---|
| Животные |  | |
| Растения | ||
| Ячейки | ||
| Связанные темы | ||