Гиалуронансинтаза
| Гиалуронансинтаза | |||
|---|---|---|---|
 Гиалуронансинтаза вируса хлореллы с ионом марганца, показанным в виде голубой сферы. [ 1 ] | |||
| Идентификаторы | |||
| Номер ЕС. | 2.4.1.212 | ||
| Номер CAS. | 39346-43-5 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| |||
Гиалуронансинтазы ( HAS ) представляют собой мембраносвязанные ферменты , которые используют УДФ-α- N -ацетил -D -глюкозамин и УДФ-α- D -глюкуронат в качестве субстратов для производства гликозаминогликана гиалуронана на поверхности клетки и вытеснения его через мембрану в клетки. внеклеточное пространство.
Изоформы
[ редактировать ]На сегодняшний день описаны три гиалуронансинтазы млекопитающих — HAS1 , HAS2 и HAS3 . Каждая из этих изоформ находится в разных участках хромосомы. [ 2 ] и был клонирован . [ 3 ] Двумя основными различиями между изоформами являются длина цепи молекул гиалуронана , которые они производят, и легкость, с которой они могут высвобождаться с поверхности клетки. [ 4 ] [ 5 ] Когда клетки млекопитающих стимулируются изменениями в их непосредственном окружении ( цитокины , близость внеклеточного матрикса ), изоформы HAS реагируют по-разному и, по-видимому, находятся под разными механизмами контроля.
Во время развития эмбриона каждая изоформа уникально экспрессируется как в пространстве, так и во времени.
- HAS2, вероятно, является наиболее важной синтазой на данный момент, поскольку мыши, лишенные способности экспрессировать HAS2 ( мыши с нокаутом ), умирают в середине беременности . [ 6 ]
- Мыши, нокаутные по HAS1 или HAS3, не оказывают влияния на развитие плода. [ 7 ]
Изоформы HAS также обладают различными физиологическими эффектами и терапевтическим потенциалом. HAS2 сверхэкспрессируется в клеточных линиях рака молочной железы и связан с в лимфатические узлы метастазированием , тогда как HAS1 и HAS3 не имеют какой-либо корреляции с развитием рака или метастазированием. [ 8 ] HAS-2 также был предложен в качестве нанотерапевтического средства для борьбы с остеоартритом суставов, проявляющимся синовиальным воспалением в результате повышенной деполимеризации гиалуронана. [ 9 ] Напротив, стимуляция HAS3 связана с усилением воспаления и атеропрогрессии за счет увеличения высвобождения интерлейкинов и активации макрофагов . [ 10 ]
Структура
[ редактировать ]HAS1 имеет единственный каталитический домен и встроен в трансмембранный канал для образования гликозидных связей при выработке гиалуронана (HA). Он содержит пять трансмембранных спиралей и три интерфейсные спирали , общая архитектура которых напоминает вигвам. [ 11 ] Изоформа бактерии Pasteurella multocida (pmHAS) содержит 972 остатка, в которых делеция остатков 1–117 не влияет на активность фермента, а С-конец активного центра располагается вокруг остатков 686–703. [ 12 ] В каталитическом домене существуют два каталитических остатка; мутация аспарагиновой кислоты в аспарагин в положении 196 (D196N) приводит к потере активности GlcUA- трансферазы , а мутация аспарагиновой кислоты в лизин в положении 477 (D477K) приводит к потере активности GlcNAc -трансферазы. Комбинация обоих мутантов приводит к аналогичной активности по сравнению с диким типом. pmHAS дополнительно содержит последовательность Asp-Gly-Ser, консервативную среди бета- гликозилтрансфераз . [ 13 ]
HAS2 регулируется путем димеризации и убиквитинирования . В клетках почек обезьян COS-1 , трансфицированных мышиными плазмидами HAS2 и HAS3 , один сайт убиквитинирования виден на лизине в остатке 190. Мутант K190R образовал димер HAS2, а меченные флагом и 6myc-меченные HAS2 и HAS3 показали образование как гомо-, так и гетеродимеров друг с другом. [ 14 ] HAS вируса хлореллы (Cv-HAS) имеет примерно 45% сходства последовательности с человеческим HAS2. [ 11 ] [ 13 ]
HAS3 регулируется посредством усечения конца 3'UTR , что подавляет NUDT21 , регулятор полиаденилирования . Потеря этого регуляторного сайта на моделях грызунов приводит к неопластическим , способствующих ремоделированию процессам, увеличению фенотипов , и повышению синтеза ГК, но также увеличивает вероятность легочной гипертензии . [ 15 ] HAS3 имеет более высокую идентичность последовательности с HAS2 (71%) по сравнению с HAS1 (57%). [ 16 ] HAS1, HAS2 и HAS3 имеют тринадцать, четырнадцать и четырнадцать остатков цистеина соответственно по сравнению с S. pyogenes (spHAS), который содержит шесть остатков цистеина. Цистеин сульфгидрилом, , отравленный в положении 225 в spHAS ингибирует активность фермента, но неясно, оказывают ли дисульфидные связи существенное влияние на активность ферментов HAS. Графики гидропатии трех изоформ HAS1, HAS2 и HAS3 позволяют предположить, что каждый белок организован в мембране одинаковым образом. [ 17 ]
Механизм
[ редактировать ]Инициация биосинтеза гиалуронана гиалуронансинтазами класса I включает генерацию праймера N-ацетилглюкозамина (GlcNAc) in situ посредством гидролиза UDP - GlcNAc с последующей диффузией праймера в активный центр. [ 11 ] Активный сайт содержит два отдельных домена, каждый из которых способен связывать либо возникающую цепь УДФ-гиалуронан, либо мономер УДФ-сахар . [ 18 ] Удлинение цепи, начиная с праймера GlcNAc, происходит при последовательном добавлении чередующихся единиц UDP-GlcA и UDP-GlcNAc к восстанавливающему концу растущей цепи. [ 19 ]
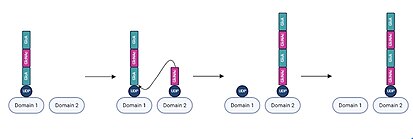
На каждой итерации удлинения цепи один домен активного сайта занят существующей цепью УДФ-гиалуронана. Мономер UDP-сахара, соответствующий следующей единице, затем связывается с незанятым доменом активного сайта. [ 20 ] Впоследствии гидроксильная группа на связанном мономере УДФ-сахара осуществляет нуклеофильную атаку на аномерный углерод восстанавливающего концевого мономера цепи УДФ-гиалуронана, вытесняя УДФ из цепи гиалуронана и сдвигая удлиненную цепь в домен, ранее занимаемый УДФ-мономер сахара. После этого процесса смещенный UDP отделяется от другого домена активного сайта. [ 17 ] Затем процесс связывания и удлинения мономера повторяется с чередованием добавления единиц GlcA и GlcNAc по мере того, как цепь UDP-гиалуронана сдвигается от одного домена активного сайта к другому. [ 21 ]
HAS1, HAS2 и HAS3 осуществляют функционально эквивалентный биосинтез гиалуронана, но демонстрируют различия в кинетических характеристиках, в том числе HAS1 демонстрирует более высокую константу Михаэлиса, чем HAS2 или HAS3. [ 22 ] На кинетику HAS и транспорт белка также влияют посттрансляционные и эпигенетические модификации . [ 23 ] [ 24 ]
Роль в метастазировании рака
[ редактировать ]HAS может играть роль на всех стадиях метастазирования рака. Вырабатывая антиадгезивную HA, HAS может позволить опухолевым клеткам высвободиться из первичной опухолевой массы, а если HA связывается с рецепторами, такими как CD44, активация Rho GTPases может способствовать EMT раковых клеток. Во время процессов интравазации или экстравазации взаимодействие HAS, продуцирующего HA, с рецепторами, такими как CD44 или RHAMM, способствует клеточным изменениям, которые позволяют раковым клеткам проникать в сосудистую или лимфатическую систему. Путешествуя по этим системам, ГК, вырабатываемая HAS, защищает раковую клетку от физического повреждения. Наконец, при формировании метастатического поражения HAS продуцирует HA, позволяя раковой клетке взаимодействовать с нативными клетками во вторичном участке и производить опухоль самостоятельно. [ 25 ]
Увеличение производства ГК раковыми клетками увеличивает инвазивную способность. Взаимодействие HA с CD44 активирует киназу фокальной адгезии (FAK), важную молекулу в процессе подвижности клеток, путем координации растворения фокальных спаек на переднем крае клетки и образования на отстающем крае. [ 26 ] Другим сигнальным путем, активируемым взаимодействием HA с CD44, является путь Akt, который приводит к экспрессии остеопонтина, молекулы, которая может стимулировать миграцию клеток. [ 27 ] Предполагается, что ГК, вырабатываемая HAS, защищает раковую клетку от физического повреждения в кровеносной или лимфатической системах. Эта роль ГК была показана на других типах клеток, но еще не исследована на раковых клетках. [ 28 ] ГК, продуцируемая HAS, усиливает секрецию различных ММП, протеолитических ферментов, которые участвуют во многих стадиях метастатического каскада. [ 29 ] Исследования показали, что различные HAS могут по-разному влиять на стадии метастазирования в зависимости от молекулярной массы и количества ГК, которую они производят.
Ссылки
[ редактировать ]- ^ Мэлони, Финн П.; Куклевич, Джереми; Кори, Робин А.; Би, Юньчэнь; Хо, Руоя; Матеушяк, Лукаш; Простите, Элс; Стейерт, Ян; Стэнсфельд, Филип Дж.; Циммер, Йохен (7 апреля 2022 г.). «Структура, распознавание субстрата и инициация гиалуронансинтазы» . Природа . 604 (7904): 195–201. Бибкод : 2022Natur.604..195M . дои : 10.1038/s41586-022-04534-2 . ПМЦ 9358715 . ПМИД 35355017 .
- ^ Спайсер А.П., Селдин М.Ф., Олсен А.С., Браун Н., Уэллс Д.Е., Доггетт Н.А., Итано Н., Кимата К., Инадзава Дж., Макдональд Дж.А. (1997). «Хромосомная локализация генов гиалуронансинтазы человека и мыши». Геномика . 41 (3): 493–7. дои : 10.1006/geno.1997.4696 . ПМИД 9169154 .
- ^ Итано Н., Кимата К. (2002). «Гиалуронансинтазы млекопитающих» . ИУБМБ Жизнь . 54 (4): 195–9. дои : 10.1080/15216540214929 . ПМИД 12512858 .
- ^ Итано Н., Савай Т., Ёсида М., Ленас П., Ямада Ю., Имагава М., Шиномура Т., Хамагути М., Ёсида Ю., Онуки Ю., Мияучи С., Спайсер А.П., Макдональд Дж.А., Кимата К. (1999). «Три изоформы гиалуронансинтазы млекопитающих обладают различными ферментативными свойствами» . Журнал биологической химии . 274 (35): 25085–92. дои : 10.1074/jbc.274.35.25085 . ПМИД 10455188 .
- ^ Стерн Р., Асари А.А., Сугахара К.Н. (2006). «Фрагменты гиалуроновой кислоты: информационная система». Европейский журнал клеточной биологии . 85 (8): 699–715. дои : 10.1016/j.ejcb.2006.05.009 . ПМИД 16822580 .
- ^ Камениш Т.Д., Спайсер А.П., Брем-Гибсон Т., Бистерфельдт Дж., Августин М.Л., Калабро А., Кубалак С., Клевер С.Е., Макдональд Дж.А. (2000). «Нарушение гиалуронансинтазы-2 отменяет нормальный сердечный морфогенез и опосредованную гиалуронатом трансформацию эпителия в мезенхиму» . Журнал клинических исследований . 106 (3): 349–60. дои : 10.1172/JCI10272 . ПМК 314332 . ПМИД 10930438 .
- ^ Бай К.Дж., Спайсер А.П., Маскаренхас М.М., Ю.Л., Очоа К.Д., Гарг Х.Г., Куинн Д.А. (2005). «Роль гиалуронансинтазы 3 в повреждении легких, вызванном аппаратом искусственной вентиляции легких» . Американский журнал респираторной медицины и медицины интенсивной терапии . 172 (1): 92–8. дои : 10.1164/rccm.200405-652OC . ПМК 2718450 . ПМИД 15790861 .
- ^ Ли, Пэн; Сян, Тинсю; Ли, Хунчжун; Ли, Цяньцянь; Ян, Бинг; Хуан, Цзин; Чжан, Сян; Ши, Юань; Тан, Цзиньсян; Жэнь, Гошэн (1 октября 2015 г.). «Сверхэкспрессия гиалуронансинтазы 2 коррелирует с онкогенезом и метастазами рака молочной железы человека» . Международный журнал клинической и экспериментальной патологии . 8 (10): 12101–12114. ПМК 4680340 . ПМИД 26722395 .
- ^ Ли, Хуйминь, Лэй, Ли, Лицинь; Фэн, Кэ, Вэй; Сюй, Чунь; Лун, Син (ноябрь 2019 г.). эндогенной гиалуроновой кислоты за счет доставки гиалуронансинтазы 2» / Бибкод . «Нанотерапия суставов : 2019AdM .... . doi : adma.201904535 . 10.1002 .S2CID 3104535L : увеличение производства 202745517 .
- ^ Хоманн, Сюзанна; Грандох, Мария; Киене, Лена С.; Подсвязек, Янина; Фельдманн, Катрин; Рабауш, Берит; Надь, Надин; Лер, Стефан; Кречмер, Инга; Оберхубер, Александр; Боллики, Пол; Фишер, Йенс В. (март 2018 г.). «Гиалуронансинтаза 3 способствует воспалению бляшек и атеропрогрессии» . Матричная биология . 66 :67-80. дои : 10.1016/j.matbio.2017.09.005 . ПМК 6023397 . ПМИД 28987865 .
- ^ Перейти обратно: а б с Мэлони, Финн П.; Куклевич, Джереми; Кори, Робин А.; Би, Юньчэнь; Хо, Руоя; Матеушяк, Лукаш; Простите, Элс; Стейерт, Ян; Стэнсфельд, Филип Дж.; Циммер, Йохен (7 апреля 2022 г.). «Структура, распознавание субстрата и инициация гиалуронансинтазы» . Природа . 604 (7904): 195–201. Бибкод : 2022Natur.604..195M . дои : 10.1038/s41586-022-04534-2 . ПМЦ 9358715 . ПМИД 35355017 .
- ^ Цзин, В. (10 июня 2003 г.). «Анализ двух активных центров гиалуронансинтазы и хондроитинсинтазы Pasteurella multocida» . Гликобиология . 13 (10): 661–671. дои : 10.1093/гликоб/cwg085 . ПМИД 12799342 .
- ^ Перейти обратно: а б Цзин, В.; ДеАнджелис, Польша (1 сентября 2000 г.). «Рассечение двух трансферазных активностей гиалуронансинтазы Pasteurella multocida: в одном полипептиде существуют два активных центра» . Гликобиология . 10 (9): 883–889. дои : 10.1093/гликоб/10.9.883 . ПМИД 10988250 .
- ^ Карусу, Евгения; Камирё, Масару; Скандалис, Спирос С.; Руусала, Айно; Астериу, Триас; Пасси, Альберто; Ямасита, Хидетоши; Хеллман, Ульф; Хелдин, Карл-Хенрик; Хелдин, Параскеви (июль 2010 г.). «Активность гиалуронансинтазы 2 регулируется путем димеризации и убиквитинирования» . Журнал биологической химии . 285 (31): 23647–23654. дои : 10.1074/jbc.M110.127050 . ПМЦ 2911313 . ПМИД 20507985 .
- ^ Ценг, Виктор; Коллум, Скотт Д.; Аллаузи, Айед; Кротти, Кэтрин; Елигар, Саманта; Траммелл, Аарон; Райан Смит, М.; Канг, Бум-Ён; Сатлифф, Рой Л.; Ингрэм, Дженнифер Л.; Джьотула, Сома ССК; Тандавараян, Раджараджан А.; Хуанг, Ховард Дж.; Нозик, Ева С.; Вагнер, Эрик Дж.; Майкл Харт, К.; Кармути-Кинтана, Гарри (август 2022 г.). «Укорочение 3'UTR HAS2 способствует гиперсинтезу гиалуронана и биоэнергетической дисфункции при легочной гипертензии» . Матричная биология . 111 : 53–75. дои : 10.1016/j.matbio.2022.06.001 . ПМЦ 9676077 . ПМИД 35671866 .
- ^ Спайсер, Эндрю П.; Олсон, Джеффри С.; Макдональд, Джон А. (апрель 1997 г.). «Молекулярное клонирование и характеристика кДНК, кодирующей третью предполагаемую гиалуронансинтазу млекопитающих» . Журнал биологической химии . 272 (14): 8957–8961. дои : 10.1074/jbc.272.14.8957 . ПМИД 9083017 .
- ^ Перейти обратно: а б Вайгель, Пол Х.; Хаскалл, Винсент К.; Тамми, Маркку (май 1997 г.). «Гиалуронансинтазы» . Журнал биологической химии . 272 (22): 13997–14000. дои : 10.1074/jbc.272.22.13997 . ПМИД 9206724 .
- ^ Хаббард, Кейтлин; Макнамара, Джошуа Т.; Адзумая, Кэли; Патель, Мехул С.; Циммер, Йохен (апрель 2012 г.). «Гиалуронансинтаза катализирует синтез и мембранную транслокацию гиалуронана». Журнал молекулярной биологии . 418 (1–2): 21–31. дои : 10.1016/j.jmb.2012.01.053 . ПМИД 22343360 .
- ^ ДеАнджелис, Польша (ноябрь 1999 г.). «Гиалуронансинтазы: захватывающие гликозилтрансферазы позвоночных, бактериальных патогенов и вирусов водорослей» . Клеточные и молекулярные науки о жизни . 56 (7): 670–682. дои : 10.1007/s000180050461 . ПМЦ 11147114 . ПМИД 11212314 . S2CID 21344879 .
- ^ Прем, П. (1 апреля 1983 г.). «Синтез гиалуроната в дифференцированных клетках тератокарциномы. Механизм роста цепи» . Биохимический журнал . 211 (1): 191–198. дои : 10.1042/bj2110191 . ПМЦ 1154343 . ПМИД 6870820 .
- ^ Вайгель, Пол Х. (1 октября 2002 г.). «Функциональные характеристики и каталитические механизмы бактериальных гиалуронансинтаз» . IUBMB Life (Международный союз биохимии и молекулярной биологии: Жизнь) . 54 (4): 201–211. дои : 10.1080/15216540214931 . ПМИД 12512859 . S2CID 25537143 .
- ^ Итано, Такахиро, Мамору; Ямада, Имагава, Мичико; Хамагути, Мичинари; Онуки, Ёдзи; , Кодзи (август 1999 г.). Макдональд, Джон А.; Кимата Гиалуронансинтазы млекопитающих обладают отличительными ферментативными свойствами» . Журнал биологической химии . 274 (35): 25085–25092. doi : 10.1074/jbc.274.35.25085 . PMID 10455188 .
- ^ Каон, Илария; Парнигони, Арианна; Виола, Мануэла; Карусова, Евгения; Пасси, Альберто; Вигетти, Давиде (январь 2021 г.). «Метаболизм клеточной энергии и синтез гиалуронана» . Журнал гистохимии и цитохимии . 69 (1): 35–47. дои : 10.1369/0022155420929772 . ПМК 7780193 . ПМИД 32623953 .
- ^ Мелеро-Фернандес де Мера, РМ; Арасу, Юта; Карня, Р.; Оикари, С.; Рилла, К.; Вигетти, Д.; Пасси, А.; Хелдин, П.; Тамми, Мичиган; Дин, Эй Джей (июль 2019 г.). «Влияние мутаций в сайтах посттрансляционной модификации на транспорт гиалуронансинтазы 2 (HAS2)». Матричная биология . 80 : 85–103. дои : 10.1016/j.matbio.2018.10.004 . ПМИД 30394292 . S2CID 53217005 .
- ^ Бхарадвадж А.Г., Ковар Дж.Л., Лафман Э., Еловски С., Окли Г.Г., Симпсон М.А. (2009). «Спонтанному метастазированию рака простаты способствует избыточный синтез и переработка гиалуронана» . Американский журнал патологии . 174 (3): 1027–36. дои : 10.2353/ajpath.2009.080501 . ПМЦ 2665762 . ПМИД 19218337 .
- ^ Фудзита И., Китагава М., Накамура С., Адзума К., Исии Г., Хигаси М., Киши Х., Хиваса Т., Кода К., Накадзима Н., Харигая К. (2002). «Передача сигналов CD44 через киназу фокальной адгезии и ее антиапоптотический эффект» . Письма ФЭБС . 528 (1–3): 101–8. дои : 10.1016/S0014-5793(02)03262-3 . ПМИД 12297287 .
- ^ Пак Джей Би, Квак Х.Дж., Ли Ш. (2008). «Роль гиалуронана в инвазии глиомы» . Адгезия и миграция клеток . 2 (3): 202–7. дои : 10.4161/cam.2.3.6320 . ПМК 2634087 . ПМИД 19262113 .
- ^ Цзян Д., Лян Дж., Фань Дж., Ю С., Чен С., Луо Ю., Прествич Г.Д., Маскаренхас М.М., Гарг Х.Г., Куинн Д.А., Гомер Р.Дж., Гольдштейн Д.Р., Букала Р., Ли П.Дж., Меджитов Р., Нобл П.В. (2005) . «Регуляция повреждения и восстановления легких с помощью Toll-подобных рецепторов и гиалуронана». Природная медицина . 11 (11): 1173–9. дои : 10.1038/nm1315 . ПМИД 16244651 . S2CID 11765495 .
- ^ Данн К.М., Ли ПК, Уилсон СМ, Иида Дж., Василук К.Р., Хаггер М., Маккарти Дж.Б. (2009). «Ингибирование гиалуронансинтазы снижает экспрессию и активность матриксной металлопротеиназы-7 (MMP-7)». Операция . 145 (3): 322–9. дои : 10.1016/j.surg.2008.11.008 . ПМИД 19231585 .
Внешние ссылки
[ редактировать ]- ЕС 2.4.1.212
- гиалуронан + синтаза Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)