Strepsirrhini
| Strepsirrhini | |
|---|---|

| |
| Образец разнообразия стрепсиргина; Восемь биологических родов изображены (сверху, слева направо): Lemur , Propithecus , Daubentonia , Varecia , Microcebus , † Darwinius , Loris , Otolemur | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Сорт: | Млекопитающая |
| Заказ: | Приматы |
| Подотряд: | Strepsirrhini E. Джеффрой , 1812 |
| Инфрадарты | |
|
† Адапориформ | |

| |
| ● Эоцен-миоценовые окаменелости Диапазон существующих приматов стрепсиргина | |
Strepsirrhini или Strepsirhini ( / s t r ɛ p s ə r aɪ n i / ; Strep -sə -ry -nee ) -это подборка приматов из , который включает в себя лимуриформные приматы, которые состоят из Мадагаскара , из Галаго ( «Бушбаби») и Потто лемуров Африки , а также Лорисов из Индии и Юго -Восточной Азии . [ А ] В совокупности они называются Strepsirrhines. Также принадлежат приматы приматов , которые процветали во время эоцена в Европе, Северной Америке и Азии, но исчезли из большей части северного полушария, когда климат охлаждается. Адапариформ иногда называют «лемур-подобными», хотя разнообразие как лемуров, так и адапиформ не подтверждает это сравнение.
Стрепсиррины определяются их «влажным» (влажным) ринарием (кончиком морды)-отсюда и разговорным, но неточным термином «мокрой носовой»-аналогично носорогам собак и кошек. У них также есть меньший мозг, чем симии сравнительно размера , большие обонятельные доли для запаха, рватный орган для обнаружения феромонов и двукратный матку с эпителиохариальной плацентой . Их глаза содержат отражающий слой, чтобы улучшить их ночное зрение , и их глазные розетки включают кольцо кости вокруг глаза, но им не хватает стены тонкой кости за ним. Приматы Strepsirrhine производят свой собственный витамин С , тогда как гаплориновые приматы должны получить его из своих рационов. Лемуриформные приматы характеризуются зубцами , специализированным набором зубов спереди, нижняя часть рта, в основном используемая для расчесывания меха во время груминга .
Многие из сегодняшних живых стрепсирринов находятся под угрозой исчезновения из -за разрушения среды обитания , охоты на бусмит и живого захвата для экзотической торговли домашними животными . Как живые, так и вымершие стрепсиррины поведенчески разнообразны, хотя все в основном являются древесными (Деревцы). Большинство живых лемуроформных явлений являются ночными , в то время как большинство адапиформ были суточными . Как живые, так и вымершие группы в основном питались фруктами , листьями и насекомыми .
Этимология
[ редактировать ]Таксономическое ), Rhinos), Rhinos), Rhinos ) название Стрепсиррини происходит от греческого στρέέέές Стрепсис «поворот» и ῥῥς rhis «нос, морда, (в пл.) Ноздри» ( Gen ῥινός hroinos , [ 5 ] что относится к появлению извилистых (в форме запятой) ноздрей на ринариуме или влажном носу. [ 6 ] [ 7 ] Название было впервые использовано французским натуралистом Этиенной Джеффрой Сен-Хилером в 1812 году в качестве подчиненного звания, сравнимого с Платримини ( обезьяны Нового Света ) и Катархини ( обезьяны Старого Света ). [ 8 ] В своем описании он упомянул « Les Narines Terminales et Sinueuses » («Терминал и обмотки ноздри»). [ 9 ]
Когда британский зоолог Реджинальд Иннес Покок возродил Стрепсиррини и определил Хаплорини в 1918 году, он опустил второй [ 10 ] Это « Haplo of этого», преподает это » и« Haplo rr этого »). [ 11 ] [ 12 ] Хотя он не удалил второй «R» из Platyrrhini или Catarrhini, оба из которых также были названы E. Джеффрой в 1812 году. После Покока многие исследователи продолжали писать Strepsirrhini с одним "R" до тех пор, пока приматологи Полина Дженкинс и Прю Нейпир не указали на ошибку в 1987 году. [ 13 ]
Эволюционная история
[ редактировать ]Стрепсиррины включают в себя вымершие адапариформы и лимуриформные приматы, которые включают лемуры и лоризоиды ( Lorises , Pottos и Galagos ). [ 14 ] Стрепсиррины расходились от приматов гаплорина в начале излучения приматов между 55 и 90 млн лет. Старые даты дивергенции основаны на оценках генетического анализа , в то время как более молодые даты основаны на дефицитной записи ископаемых . Лемуриформные приматы могли развиваться из Cercamoniines или Sivaladapids , которые были адапариформи, которые могли возникнуть в Азии. Когда -то считалось, что они эволюционировали из адапидов , более специализированной и более молодой ветви адапиформ, прежде всего из Европы.
| Примат филогения | |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| Strepsirrhines и Haplorhines расходились вскоре после появления первых настоящих приматов (эуприматов). Взаимосвязь между эуприматами, трэшами , колугосом и plesiadapiforms менее определена. Иногда Plesiadapiforms сгруппированы с эуприматами в соответствии с приматами порядка , Colugos сгруппированы с приматами при Primatomorpha , и все четыре сгруппированы в соответствии с Еваархонтой . [ 15 ] |
Лемуры попали из Африки в Мадагаскар между 47 и 54 млн лет, тогда как Лорисы отделялись от африканских галаго, около 40 млн лет, а затем колонизировали Азию. Лемуриформные и, в частности, лемуры Мадагаскара, часто изображаются ненадлежащим образом как « живые окаменелости » или в качестве примеров « базальных » или «низших» приматов. [ 16 ] [ 17 ] [ 18 ] Эти взгляды исторически препятствовали пониманию эволюции млекопитающих и эволюции стрипсиргиновых черт, [ 16 ] такие как их зависимость от запаха ( обоняния ), характеристики их скелетной анатомии и размер их мозга, что относительно невелико. В случае лемуров естественный отбор заставила эту изолированную популяцию приматов значительно диверсифицировать и заполнить богатое разнообразие экологических ниш , несмотря на их меньший и менее сложный мозг по сравнению с симианами. [ 17 ] [ 18 ]
Неясное происхождение
[ редактировать ]Дивергенция между стрепсирринами, симианами и тарси, вероятно, последовало почти сразу после того, как приматы впервые развивались. [ 19 ] Хотя несколько окаменелостей живых приматских групп - лемуроформных, тарсеров и симиан - известны с раннего и среднего эоцена, [ 20 ] [ 21 ] [ 22 ] Данные из генетики и недавних ископаемых обнаруживают, что оба предполагают, что они могли присутствовать во время раннего адаптивного излучения . [ 23 ]
Происхождение самых ранних приматов, из которых эволюционировали оба симии и ласки, является загадкой. И их место происхождения, и группа, из которой они появились, неясны. Хотя ископаемые записи , демонстрирующие их первоначальное излучение через северное полушарие, очень подробно,, [ 24 ] Окаменелостные записи от тропиков (где приматы, скорее всего, впервые развивались) очень редки, особенно в то время, когда приматы и другие основные клады [ B ] эврианских млекопитающих впервые появились.
Отсутствие подробных тропических окаменелостей, генетики и приматологи использовали генетический анализ для определения родства между линиями приматов и количеством времени с момента их расходящегося . Используя эти молекулярные часы , даты дивергенции для основных линий приматов предположили, что приматы развивались более 80–90 млн лет, почти 40 миллионов лет до того, как первые примеры появляются в записи ископаемых. [ 25 ]
Ранние приматы включают как ночные , так и суточные виды с мелким телом, [ 26 ] и все были древесными, с руками и ногами, специально адаптированными для маневрирования на маленьких ветвях. [ 27 ] Плезиадапариформ из раннего палеоцена иногда считаются «архаичными приматами», потому что их зубы напоминали зубы ранних приматов и потому, что они обладали адаптацией к жизни на деревьях, таких как расходящийся большой носок ( Hallux ). Хотя Plesiadapiforms были тесно связаны с приматами, они могут представлять собой парафилетическую группу, из которой приматы могут или не могут быть напрямую развиваться, [ 28 ] и некоторые роды , возможно, были более тесно связаны с Колугосом , [ C ] которые считаются более тесно связанными с приматами. [ 29 ]

Первые настоящие приматы (эуприматы) не появляются в ископаемом отчете до раннего эоцена (~ 55 млн лет), после чего они излучались через северное полушарие в течение короткого периода быстрого глобального потепления, известного как палеоцен -эоценовый термический максимум . [ 23 ] Эти первые приматы включали Канюс , Донрусселли , Алтания и Тейлхардина на северных континентах, [ 30 ] а также более сомнительные (и фрагментарные) ископаемые Альтиатлазия из палеоцена Африки. [ 19 ] Эти самые ранние ископаемые приматы часто делятся на две группы, адапариформ [ D ] и омбомиформ [ E ] Оба появились внезапно в ископаемом записи без переходных форм, чтобы указать происхождение, [ 31 ] и обе группы были богаты разнообразием и были широко распространены на протяжении всего эоцена.
Последним филиалом, который развивался, были адапариформа, разнообразная и широко распространенная группа, которая процветала во время эоцена (от 56 до 34 миллионов лет назад [ MYA ]) в Европе, Северной Америке и Азии. Они исчезли из большей части северного полушария , когда климат охлаждал: последняя из адапаформ вымерла в конце миоцена ( ~ 7 млн лет).
Адапариформная эволюция
[ редактировать ]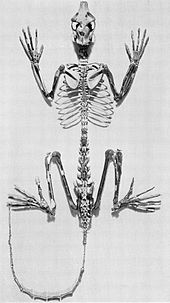
Приматы адапариформ - это вымершие стрепсиррины, которые разделяют много анатомических сходств с лемурами. [ 32 ] Иногда их называют приматами, похожими на лемур, хотя разнообразие как лемуров, так и адапиформ не поддерживает эту аналогию. [ 33 ]
Как и живые стрепсиррины, адапариформ были чрезвычайно разнообразными, [ 16 ] с по меньшей мере 30 родами и 80 видами, известными по ископаемым записям с начала 2000 -х годов. [ 34 ] [ 35 ] Они диверсифицировались по всей Лорасии во время эоцена, [ 32 ] Некоторые достигают Северной Америки через сухопутный мост . [ 36 ] Они были одними из самых распространенных млекопитающих, встречающихся в ископаемых кроватях с того времени. Несколько редких видов также были найдены в Северной Африке. [ 32 ] Наиболее базальными из адапариформ являются роды Кантий из Северной Америки и Европы и Донрусселли из Европы. Последний носит самые наследственные признаки , поэтому его часто считают родственной группой или группой STEM другой адапариформы. [ 37 ]
Адапариформ часто делятся на три основные группы:
- Адапиды чаще всего встречаются в Европе, хотя самые старые образцы ( адапоиды из среднего эоцена Китая) указывают на то, что они, скорее всего, развивались в Азии и иммигрировали. Они вымерли в Европе во время грандиозного купе , часть значительного события вымирания в конце эоцена. [ 38 ]
- Notharctids , которые наиболее близко напоминали некоторые из лемуров Мадагаскара, приезжают из Европы и Северной Америки. Европейское отделение часто называют Cercamoniines . [ 39 ] Североамериканская филиал процветала во время эоцена, но не выжила в олигоцене . [ 40 ] [ 41 ] Как и адапиды, европейская филиала также вымерли к концу эоцена. [ 36 ]
- Сивадапиды Южной и Восточной Азии наиболее известны из миоцена , и единственные адапариформ, чтобы выжить после границы эоцена/олигоцена (~ 34 млн лет). Их отношение к другим адапиформ остается неясным. [ 42 ] Они исчезли до окончания миоцена (~ 7 млн лет). [ 32 ]
Взаимосвязь между адапариформой и лимуриформной приматами не была четко продемонстрирована, поэтому положение адапариформ в качестве парафилетической группы STEM сомнительно. Как данные молекулярных часов, так и новые ископаемые обнаруживают, что лимуриформная дивергенция от других приматов и последующее разделение лемур-лоризоидов оба предшествуют появлению адапариформ в раннем эоцене. [ 31 ] Новые методы калибровки могут согласовать расхождения между молекулярными часами и ископаемыми записями, в пользу более поздних дат дивергенции. [ 43 ] Окаменечная запись предполагает, что адапиформ стрепсиргина и гаплориновые омомиформы развивались независимо до раннего эоцена, хотя их наиболее базальные члены имеют достаточное количество стоматологических сходств, чтобы предположить, что они расходились во время палеоцена (66–55 MYA). [ 32 ]
Лемуриформная эволюция
[ редактировать ]Lemurifors Origins неясно и обсуждается. Американский палеонтолог Филипп Гингерх предположил, что лемуриформные приматы эволюционировали из одного из нескольких родов европейских адапидов, основанных на сходстве между передними нижними зубами адапидов и зубцами существующей лемуроформной формы; Тем не менее, эта точка зрения не сильно поддерживается из -за отсутствия явных переходных окаменелостей. [ 44 ] Вместо этого, лемуриформные средства могут быть выпущены от очень ранней ветви азиатских Cercamoniines или Sivaladapids, которые мигрировали в Северную Африку. [ 22 ] [ 45 ]
трех 40-миллионных ископаемых лоризоидов ( Каранизия , Сахарагалаго и Вадилемер ) в лемуриформные отложениях Египта Эль-Фаюм в период между 1997 и 2005 годами, самые старые известные Пока открытия Уганда . Эти новые находки демонстрируют, что во время среднего эоцена присутствовали лемуроформные приматы и что лимуриформная линия и все другие таксоны стрепсиргина расходились до этого. [ 46 ] [ 47 ] [ 48 ] Jebelemur из Туниса датируется поздним ранним или ранним средним эоценом (от 52 до 46 млн лет) и считался Cercamoniine, [ 49 ] но также, возможно, был стеблем лемуроформной. [ 22 ] Азибииды из Алжира датируются примерно в то же время и могут быть родственной группой джебелемеридов . Вместе с Plesiopithecus из позднего Эйгипта Эйгипта эти три могут квалифицироваться как стволовые лимуриформ из Африки. [ 45 ]
Оценки молекулярных часов показывают, что лемуры и лоризоиды расходились в Африке во время палеоцена, приблизительно 62 млн лет. Между 47 и 54 млн лет лемуры рассеялись на Мадагаскар с помощью рафтинга . [ 47 ] В изоляции лемуры диверсифицировали и заполняли ниши, часто заполненные обезьянами и обезьянами сегодня. [ 50 ] В Африке лорисы и галаго отдались во время эоцена, примерно 40 млн. М. [ 47 ] В отличие от лемуров на Мадагаскаре, им пришлось конкурировать с обезьянами и обезьянами, а также с другими млекопитающими. [ 17 ]
История классификации
[ редактировать ]
Таксономия Strepsirrhines является спорной и имеет сложную историю. Смущенные таксономические терминологии и упрощенные анатомические сравнения создали заблуждения о приматах и филогении стрепсиргина , иллюстрируемые вниманием средств массовой информации, окружающей единственную ископаемость "IDA" в 2009 году.
Приматы стрепсиррина были впервые сгруппированы под родом Lemur шведским таксономистом Карлом Линнеусом в 10 -м издании Systema Naturae, опубликованном в 1758 году. В то время было признано только три вида, один из которых (Colugo) больше не признается приматом. [ 51 ] В 1785 году голландский натуралист Питер Боддарт разделил род Lemur на два рода: Prosimia для Lemurs, Colugos, Tarsiers и Tardigradus для Lorises. [ 52 ] Десять лет спустя, é. Джеффрой и Жорж Кювье сгруппировали таперов и галаго из -за сходства в их морфологии задних конечностей , которая поддерживала немецкий зоолог Йоханн Карл Вильгельм Иллигер , который поместил их в семейный макротарси во время размещения лемуров и Тарсие в семье просимии (просами) в 1811 году в 1811 году в 1811 году в 1811 году в 1811 году в 1811 году в 1811 году в 1811 году. . [ 53 ]
Английский сравнительный анатомист Уильям Генри Флауэр создал подготовку Lemroide в 1883 году, чтобы отличить эти примыты от симиан, которые были сгруппированы под английским биологом ул. Подряд Джордж Джексон Миварт Антропоэда (= SIMFIMS). Согласно цвету, подготовка Лемруайда содержала семейство семей Лемурды (лемуры, лориз и галаго), херомида ( да-эй ) и стратегическое. Lemroidea была позже заменена подготовкой Иллигера Prosimi. [ 53 ] Много лет назад, в 1812 году, é. Джеффрой впервые назвал Suborde Strepsirrhini, в который он включил Tarsiers. [ 54 ] Эта таксономия оставалась незамеченной до 1918 года, когда Покок сравнивал структуру носа и восстановил использование подряд стрепсиррини, а также перемещала копетки и симии в новый подряд, Хаплорини. [ 11 ] [ 53 ] Только в 1953 году, когда британский анатомист Уильям Чарльз Осман Хилл написал целый объем об анатомии стрепсиррина, таксономическое предложение Покока стало заметным и более широко использоваться. С тех пор таксономия приматов переключилась между стрепсррини-газлолорхини и пропими-антропоидом. [ 53 ]
Большая часть академической литературы обеспечивает базовую основу для таксономии приматов, обычно включающих несколько потенциальных таксономических схем. [ 55 ] Хотя большинство экспертов согласны с филогениями , [ 56 ] Многие не согласны со всеми уровнями классификации приматов. [ 55 ]
| 2 инфрадерк [ 3 ] | 3 инфрадерк [ 4 ] |
|---|---|
|
|
Противоречия
[ редактировать ]Наиболее часто повторяющиеся дебаты в приматологии в 1970 -х, 1980 -х годах и начале 2000 -х годов касались филогенетического положения копченов по сравнению как с симионами, так и с другими пронимицами. [ 57 ] [ 58 ] Тарси чаще всего помещаются либо в подряд, Хаплорини, с симианами, либо в подходе Просимии со стрепсирринами. [ 50 ] Prosimii является одним из двух традиционных подчиков приматов и основан на эволюционных оценках (группах, объединенных анатомическими признаками), а не на филогенетических кладах, в то время как таксономия Strepsirrhini-Haplorrhini была основана на эволюционных отношениях. [ 7 ] Тем не менее, обе системы сохраняются, потому что таксономия Prosimii-Antropoidea знакома и часто встречается в исследовательской литературе и учебниках. [ 25 ]
Стрепсиррины традиционно характеризуются несколькими симплсуоморфными (наследственными) признаками, не разделенными симианами, особенно ринарием. [ f ] [ 50 ] [ 60 ] Другие симплсуоморфии включают длинные морды , запутанные верхнеропинги , относительно большие обонятельные луковицы и меньшие мозги. The ToothComb - это синапоморфия (общая, полученная черта), наблюдаемая среди лемуроформных [ 50 ] Хотя это часто и неправильно используется для определения клады стрепсриргина. [ G ] Приматы Strepsirrhine также объединены в обладание эпителиохариальной плацентой . [ 50 ] В отличие от Tarsiers и Semians, Strepsirrhines способны производить свой собственный витамин С и не нуждаются в нем в своем рационе. [ 62 ] Дальнейшие генетические доказательства взаимосвязи между ласкими и симионами как гаплориновой клады - это общее владение тремя синусоидальными маркерами . [ 63 ]
Из -за их исторически смешанных собраний, которые включали в себя таперов и близких родственников приматов, как Prosimii, так и Strepsirrhini считались таксонами для мусора для «нижних приматов». [ 64 ] [ 65 ] Несмотря на это, клады Strepsirrrhine и Haplorrhine обычно принимаются и рассматриваются как предпочтительное таксономическое разделение. Тем не менее, Tarseriers по -разному все еще очень похожи на Strepsirrhines и Semians, [ 25 ] А так как ранний раскол между стрепсарринами, Tarsiers и Semians древние и трудно решить, [ 66 ] Иногда используется третья таксономическая договоренность с тремя подчинениями: Prosimi, Strategic и Anthropoidea. [ 14 ] Чаще всего термин «пронимиан» больше не используется в официальной таксономии, [ 67 ] но все еще используется, чтобы проиллюстрировать поведенческую экологию каперов по сравнению с другими приматами. [ 60 ]
В дополнение к противоречиям по поводу Tarsiers, дебаты о происхождении симиан, когда -то ставшие под сомнение кладу стрепсиррина. Аргументы для эволюционной связи между адапирицей и симианами, сделанными палеонтологами Гинджерихом, Элвином Л. Саймонсом , Таб Расмуссеном и другими, могли бы потенциально исключить адапариформу из Стрепсиррини. [ 68 ] [ 69 ] В 1975 году Джингерих предложил новый подборка, Symiolemuriformes, [ 70 ] предположить, что стрепсиррины более тесно связаны с симианами, чем тарси. [ 71 ] Однако никакие четкие отношения между ними не были продемонстрированы в начале 2000 -х годов. [ 69 ] Идея вскоре восстановилась в 2009 году во время внимания средств массовой информации, окружающей Дарвиний Масилле (дублированного «Ида»), Cercamoniine из Германии, который рекламировался как « недостающее звено между людьми и более ранними приматами» («Симианы и адапариформа»). [ 72 ] Однако кладистический анализ был ошибочным, а филогенетические выводы и терминология были расплывчатыми. [ 72 ] Хотя авторы отметили, что Дарвиний не был «ископаемым лемуром», они подчеркнули отсутствие зуба, [ 73 ] которые не обладали адапариформой. [ 37 ]
Инфраординальная классификация и терминология клады
[ редактировать ]
| Стрепсиррини Филогения [ 45 ] |
В пределах стрепсиррини две общие классификации включают в себя либо два инфраордации (Adapiformes и Lemuriformes) [ 74 ] или три инфраордации (Adapiformes, Lemuriformes, Lorisiformes). [ 75 ] Обнаруженные из числа язвов 20-х годов проводятся (Do Odoraion) в Trebrarer, наше использование его для поиска, Midian. [ 76 ] В некоторых случаях в приматах приматов включаются плезиадапариформ, и в этом случае эуприматы иногда рассматриваются как подряд, причем стрепсиррини становятся недостатком, а лемуроформные и другие становятся парвотелями. [ 14 ] Независимо от инфраординальной таксономии, Strepsirrhini состоит из трех ранжевых суперсемей и 14 семей, семь из которых вымерли. [ 74 ] Три из этих вымерших семей включали недавно вымерших гигантских лемуров Мадагаскара, [ 77 ] Многие из которых вымерли в течение последних 1000 лет после прибытия человека на остров. [ 78 ]
Когда Strepsirrhini делится на два инфрадарта, кладу, содержащую все приматы с зубцами, можно назвать «Lemuriforms». [ 74 ] Когда он делится на три инфрадарта, термин «Lemuriforms» относится только к лемурам Мадагаскара, [ 50 ] и приматы с зубцами называются либо «коронными стрепсирринами» [ 42 ] или «существующие стрепсиррины». [ 7 ] Путаница этой конкретной терминологии с общим термином «стрепсиргин», наряду с упрощенными анатомическими сравнениями и расплывчатыми филогенетическими выводами, может привести к неправильным представлениям о филогениях приматов и недоразумениях о приматах из эоцина, как видно из освещения в СМИ в горы . [ 72 ] Поскольку скелеты адапариформ имеют сильное сходство со скелями лемуров и лорисов, исследователи часто называют их «примитивными» стрепсирринами, [ 79 ] Лемурские предки, или сестринская группа для живых стрепсарринов. Они включены в Strepsirrhini, [ 7 ] и считаются базальными членами клады. [ 80 ] Хотя их статус как истинных приматов не подвергается сомнению, сомнительная связь между адапирицей и другими живыми и ископаемыми приматами приводит к множественным классификациям в стрепсррини. Часто адапариформ помещается в их собственное подряд из -за анатомических различий с лемуриформными и их неясными отношениями. Когда подчеркиваются общие признаки с лемурориформ (которые могут быть или не быть синапоморфными), они иногда сводятся к семействам в рамках инфракоретических Lemuriformes (или суперсемейства Lemuroidea). [ 32 ]
Первым описанным ископаемым приматом был адапирический адапис Parrisiensis французским натуралистом Жоржа Кювье в 1821 году, [ 49 ] который сравнил его с Hyrax (« Le Daman »), затем считался членом ныне устаревшей группы под названием Pachyderms . [ 81 ] Он не был признан приматом, пока он не был переоценен в начале 1870 -х годов. [ 34 ] [ 49 ] Первоначально, адапариформ были включены в семейство Adapidae, которые были разделены на две или три подсемейства: Adapinae, Notharctinae, а иногда и Sivaladapinae. Все адапиформ в Северной Америке были сосредоточены под notharctinae, в то время как формы Старого Света обычно были назначены в Adapinae. Примерно в 1990 -х годах появились две различные группы европейских «адапидов», основанных на различиях в посткраниальном скелете и зубах. Одна из этих двух европейских форм была идентифицирована как Cercamoniines, которые были связаны с нехарктсами, которые были обнаружены в основном в Северной Америке, в то время как другая группа попадает в традиционную адапидную классификацию. [ 37 ] Три основных адапиформных подразделения в настоящее время обычно рассматриваются как три семейства в адапариформ (notharctidae, adapidae и sivaladapidae), но также используются другие подразделения от одной до пяти семей. [ 34 ]
Анатомия и физиология
[ редактировать ]Управляющий аппарат
[ редактировать ]
Все лемурописы обладают специализированной стоматологической структурой, называемой «Зубком», [ 7 ] [ 82 ] за исключением Aye-Aaye, в котором структура была модифицирована в два постоянно растущих (гипсо-гиптодонтских) резцов (или собачьих зубов ), аналогично структуру грызунов . Часто ToothComb неправильно используется для характеристики всех стрепсрирринов. Вместо этого он уникален для Lemuriforms и не виден среди адапиформ.
Lemuriforms жених перорально, а также обладает когтями на втором ноге для каждой ноги для царапин в областях, которые недоступны для рта и языка. [ 7 ] [ 82 ] [ 83 ] Адапариформ, возможно, имел когти для ухода за собой, [ 37 ] Но это мало доказательств этого. [ 84 ] ToothComb состоит из двух или четырех затруднительных нижних резцов и перкатлентных нижних собак, за которыми следуют премоляра в форме собак . [ 85 ] Он используется для расчесывания меха во время перорального ухода. Пролистые волосы, которые накапливаются между зубами зубов, удаляются подмолковой или «нижней частью». [ 83 ] Адапариформ не обладал зубцем. Вместо этого их более низкие резцы варьировались по ориентации - от несколько складывания до несколько вертикали - и нижние собаки проецировались вверх и часто были заметными.
Глаза
[ редактировать ]Как и все приматы, орбиты стрепсиррина (глазные розетки) имеют посторбитальный стержень , защитное кольцо кости, созданное соединением между лобными и зигоматическими костями . [ 86 ] У живых и вымерших стрепсирринов не хватает тонкой стены кости за глаза, называемой посторбитальным закрытием, которое наблюдается только у приматов гаплорина. [ 86 ] [ 87 ] Хотя глаза Strepsirrhines указывают вперед, давая стереоскопическое зрение , [ 86 ] Орбиты не сталкиваются полностью вперед. [ 87 ] Считается, что среди живых стрепсирринов большинство или все виды обладают отражающим слоем за сетчаткой глаз, называемой таптумом Lucidum (состоящий из рибофлавина кристаллов [ 88 ] ), который улучшает зрение при слабом освещении, [ 89 ] [ 82 ] Но им не хватает фовеа , которая улучшает дневное видение. Это отличается от Tarsiers, в которых отсутствует таптум Lucidum, но обладает фовеа. [ 90 ]
Череп
[ редактировать ]
Приматы Strepsirrhine имеют мозг, относительно сравнимый с или немного больше по размеру, чем большинство млекопитающих. [ 16 ] [ 50 ] Однако по сравнению с симианами они имеют относительно небольшое соотношение размера мозга к телу. [ 87 ] Strepsirrhines также традиционно отмечается своим неисключенным симфизом нижней челюсти (две половинки нижней челюсти), [ 87 ] Однако слияние симфиза нижней челюсти было распространено в адапариформе, [ 91 ] Примечательно notharctus . [ 92 ] Кроме того, несколько вымерших гигантских лемуров демонстрировали слитый симфиз нижней челюсти. [ 93 ]
Уши
[ редактировать ]Многие ночные виды имеют большие, независимо подвижные уши, [ 94 ] [ 95 ] Хотя существуют значительные различия в размерах и формах уха между видами. [ 96 ] Структура среднего и внутреннего уха стрепсирринов отличается между лемурами и лоризоидами. У лемуров расширяется барабанная полость , которая окружает среднее ухо. Это оставляет эктотимпановое кольцо , которое поддерживает барабанную перепонку , свободную в слуховой булле . [ 97 ] Эта черта также виден в адапариформе. [ 91 ] Однако у лоризоидов барабанная полость меньше, а эктотимпановое кольцо прикрепляется к краю слуховой булла. Барабанная полость в лоризоидах также имеет два вспомогательных воздушных пространства, которых нет в лемурах.
Артерии шеи
[ редактировать ]Как лоризоиды, так и лемуры Chirogaleid заменили внутреннюю сонную артерию увеличенной восходящей глоточной артерией . [ 97 ]
Кости лодыжки
[ редактировать ]Strepsirrhines также обладают отличительными чертами в их лапке (кости лодыжки), которые отличают их от гаплоринов, таких как наклонная фасфель тало-физика (лицо, где встречаются кость и малоберцовая кость ) и разница в положении сгибателя Fibularis сухожилие на талус. [ 98 ] Эти различия дают стрепсарринам способность делать более сложные вращения лодыжки и указывать, что их ноги обычно переворачиваются или поворачиваются внутрь, адаптация для захвата вертикальных опоров. [ 99 ]
Половые характеристики
[ редактировать ]Сексуальное дихроматизм (различные паттерны окраски между мужчинами и женщинами) можно увидеть у большинства видов коричневого лемура, [ 100 ] Но в остальном лемуры показывают очень мало, если какая -либо разница в размере тела или веса между полами. Отсутствие сексуального диморфизма не характерно для всех стрепсирринов. [ 101 ] Некоторые адапариформ были сексуально диморфическими, с мужчинами с большим сагиттальным гребнем (гребень кости на вершине черепа, к которому прикрепляются мышцы челюсти) и собачьи зубы. [ 102 ] Лоризоиды демонстрируют некоторый сексуальный диморфизм, [ 101 ] Но мужчины, как правило, не более чем на 20 процентов больше, чем женщины. [ 103 ]
Ринарий и обоняние
[ редактировать ]
- (А) Дварф Лемур
- (Б) Большой Галаго
- (В) Малый Галаго
- (Г) звуковая жизнь
- (Д) Гарсир
Стрепсиррины имеют длинную морду, которая заканчивается влажным и чувствительным к сенсорным ринарием , похожим на то, что у собак и многих других млекопитающих. Ринарий окружен вибрациями , которые также чувствительны к прикосновению. Спуганные верхнечеристорбиналы на внутренней стороне их фильтра носа, тепло и увлажняют входящий воздух, в то время как обонятельные рецепторы основной обонятельной системы, выстилающие этимотурбиналы, обнаруживают запахи с воздухом. [ 50 ] [ 104 ] Обонятельные луковицы лемуров сопоставимы по размеру с другими у других древесных млекопитающих. [ 50 ]
Поверхность ринария не имеет обонятельных рецепторов, поэтому он не используется для запаха с точки зрения обнаружения летучих веществ. Вместо этого он имеет чувствительные сенсорные рецепторы ( клетки Меркель ). Ринарий, верхняя губа и десны плотно соединены складкой слизистой оболочки, называемой Philtrum , которая проходит от кончика носа ко рту. [ 105 ] Верхняя губа ограничена этим соединением и имеет меньше нервов для контроля движения, что оставляет ее менее подвижным, чем верхние губы симиан. [ 106 ] [ 107 ] Philtrum создает разрыв ( диастема ) между корнями первых двух верхних резцов. [ 105 ] [ 108 ]
Стрепсиррин ринарий может собирать относительно нелетучивые химические вещества на основе жидкости (традиционно классифицируемые как феромоны ) и передавать их в орган Вомероназального (VNO), [ 109 ] который расположен внизу и перед полостью носа , над ртом. [ 110 ] VNO -это конструкция, похожая на канал , изготовленную из хряща [ 111 ] и изолирован от воздуха, проходящего через носовую полость. [ 112 ] VNO связан с ртом через носопалатиновые протоки (которые общаются через резкое отверстие ), которые проходят через твердый небо на вершине, передняя часть рта. [ 110 ] Флюиды, перемещающиеся от ринария ко рту, а затем вверх по насопалатиновым протоках в VNO обнаруживаются, и информация передается на аксессуальную обонятельную луковицу , которая относительно велика в стрепсирринах. [ 113 ] Из аксессуальной обонятельной луковицы информация отправляется в миндалину , которая обрабатывает эмоции, а затем к гипоталамусу , который обрабатывает основные функции организма и метаболические процессы. Этот нервный путь отличается от того, который используется основной обонятельной системой. [ 114 ]
Все лемурообразные имеют vno, как и глюди и некоторые обезьяны Нового Света. [ 115 ] Адапариформа демонстрирует разрыв между верхними резцами, который указывает на наличие VNO, но существует некоторое разногласия по поводу того, обладали ли они носорогом. [ 116 ]
Репродуктивная физиология
[ редактировать ]Уплотняя стрепсиррины имеют эпителиохариальную плаценту, [ 50 ] где материнская кровь не входит в прямой контакт с хорионом плода , как в гемохориальной плаценте гаплоринов. Стрепсиррин матка имеет две отдельные камеры (двукратные). [ 106 ] Несмотря на сходные периоды беременности с гаплоринами сравнительно размера, темпы роста плода, как правило, медленнее в стрепсирринах, что приводит к новорожденному потомству, которые составляют всего треть новорожденных гаплорина. [ 106 ] [ 117 ] Существующие Strepsirrhines также имеют более низкую базальную скорость метаболизма , которая возвышается у женщин во время беременности, что удовлетворяет большие требования к матери. [ 118 ]
Большинство приматов имеют две молочные железы , [ 119 ] Но число и позиции варьируются между видами в стрипсирринах. [ 120 ] У лорисов есть две пары, [ 121 ] В то время как другие, как и кольцевой лемур, имеют одну пару на груди (грузоподъемность). [ 122 ] Ай-Ай также имеет две молочные железы, но они расположены рядом с пахом (паховым). [ 123 ] У женщин клитор иногда увеличивается и брюшна, напоминая мужской пенис , что может затруднить идентификацию пола для человеческих наблюдателей. [ 124 ] У клитора также может быть костная структура, похожая на бакулум (кость полового члена) у мужчин. [ 125 ] Большинство мужчин -приматов имеют бакулум, но он обычно больше в стрепсарринах и обычно раздвоился на кончике. [ 126 ]
Поведение
[ редактировать ]
Приблизительно три четверти всех существующих видов стрепсиргина ночной, спят в гнездах, сделанных из мертвых листьев или пустых деревьев в течение дня. [ 127 ] Все лоризоиды из континентальной Африки и Азии являются ночными, что сводит к минимуму их конкуренцию с приматами -симианами региона, которые являются суточными. Лемуры Мадагаскара, живущие в отсутствие обезьян, более изменчивы в своих циклах активности. Ай-эй, мышиные лемуры , шерстяные лемуры и спортивные лемуры являются ночными, в то время как лемуры с кольцевым хвостом и большинство их родственников, сифаков и Индри являются суточными. [ 80 ] Тем не менее, некоторые или все коричневые лемуры ( Eulemur ) являются кафеморальными , что означает, что они могут быть активными днем или ночью, в зависимости от таких факторов, как температура и хищничество . [ 128 ] Многие сохранившиеся стрепсиррины хорошо адаптированы для ночной активности из -за их относительно больших глаз; большие, подвижные уши; чувствительные тактильные волосы; сильное обоняние; и таптум Lucidum за сетчаткой. [ 95 ] Среди адапариформ большинство считаются суточными, за исключением Pronycticebus и Godinotia из средней эоцена Европы, оба из которых имели большие орбиты, которые предполагают ночную. [ 80 ] [ 91 ]
Репродукция у большинства видов стрепсиргина имеет тенденцию быть сезонным, особенно у лемуров. Ключевые факторы, которые влияют на сезонное воспроизведение, включают длину сезона влажных сил, последующую доступность пищи и время созревания вида. [ 129 ] Как и другие приматы, Strepsirrhines являются относительно медленными заводчиками по сравнению с другими млекопитающими. Их период беременности и интервалы межбита обычно длинные, и молодые развиваются медленно, как у приматов гаплорина. [ 106 ] [ 117 ] В отличие от симиан, некоторые стрепсиррины производят два или три потомства, хотя некоторые производят только одно потомство. Те, кто производит несколько потомков, имеют тенденцию строить гнезда для своих молодых. Считается, что эти две признаки являются плезиоморфными (наследственными) для приматов. [ 106 ] Молодые являются предварительными (относительно зрелыми и подвижными) при рождении, но не так координируются, как копытные (копытные млекопитающие). [ 50 ] Уход за младенцами мать относительно продлена по сравнению со многими другими млекопитающими, а в некоторых случаях младенцы цепляются за мех матери своими руками и ногами. [ 106 ]
Несмотря на их относительно меньший мозг по сравнению с другими приматами, лемуры продемонстрировали уровни технического интеллекта в решении проблем, которые сопоставимы с тем, которые наблюдаются у симиан. Тем не менее, их социальный интеллект отличается, часто подчеркивая внутреннюю конкуренцию над сотрудничеством, что может быть связано с адаптацией для их непредсказуемой среды. [ 130 ] Хотя лемуры не наблюдались с использованием объектов в качестве инструментов в дикой природе , их можно научить использовать объекты в качестве инструментов в неволе и продемонстрировать базовое понимание функциональных свойств используемых объектов. [ 131 ]
Социальные системы и общение
[ редактировать ]Ночные стрепсриррины традиционно описываются как «одинокие», хотя этот термин больше не пользуется предпочтениями исследователей, которые их изучают. [ 132 ] Многие считаются «одинокими фургерами», но многие демонстрируют сложную и разнообразную социальную организацию, часто перекрывая домашние диапазоны , инициируя социальные контакты ночью и делятся спящими местами в течение дня. Даже системы спаривания различны, как видно из шерстяных лемуров, которые живут в моногамных парах размножения . [ 133 ] Из -за этого социального разнообразия среди этих одиночных, но социальных приматов, уровень социального взаимодействия которого сопоставим с суточными симионами, [ 132 ] Были предложены альтернативные классификации, чтобы подчеркнуть их общительный, рассеянный или одиночный характер. [ 134 ]
Среди существующих стрепсирринов только суточные и кафтемральные лемуры развивались, чтобы жить в многояменных/многофемторных группах , сравнимых с большинством живых маминов. [ 133 ] [ 135 ] эта социальная черта, наблюдаемая в двух существующих семействах лемуров ( Indriidae и Lemuridae ), развивалась независимо. Считается, что [ 136 ] Размеры групп меньше в социальных лемурах, чем у обезьян, и, несмотря на сходство, структуры сообщества различаются. [ 137 ] Женское доминирование, которое редко встречается в обезьянах, довольно распространено у лемуров. [ 138 ] Strepsirrhines тратят значительное количество времени на уходу друг за другом ( Allogroming ). [ 139 ] Когда лемуриформные приматывают жениха, они облизывают мех, а затем расчесывают его со своими зубами. Они также используют свой когти для поцарапания, чтобы царапать места, которые они не могут достать своим ртом. [ 83 ]
Как и обезьяны Нового Света, Strepsirrhines полагаются на маркировку запаха для большей части их общения. Это включает в себя размазывание выделения из эпидермальных ароматических желез на ветвях деревьев, а также мочу и фекалии . В некоторых случаях Strepsirrhines может помазать себя мочой ( промывание мочи ). Позы тела и жесты могут использоваться, хотя длинная морда, не-мобильные губы и снижение лицевой энворки ограничивают использование выражений лица в стрепсирринах. вызовы с коротким диапазоном, дальние вызовы и звонки с сигнализацией . Также используются [ 140 ] Ночные виды более ограничены отсутствием света, поэтому их системы связи отличаются от систем суточных видов, часто используя на большие расстояния призывы к претенденту на их территорию . [ 141 ]
Локомоция
[ редактировать ]

Живые стрепсиррины преимущественно древесины, причем только кольцевые хвост тратит значительно время на землю. [ 142 ] Большинство видов перемещаются в четвероногих (на четырех ногах) на деревьях, в том числе пять родов более мелких, ночных лемуров. [ 127 ] [ 142 ] Галаго, индриды, спортивные лемуры и бамбуковые лемуры прыгают с вертикальных поверхностей, [ 142 ] и индриды очень специализированы для вертикального цепляния и прыжка . [ 127 ] Лорисы медленные, преднамеренные альпинисты. [ 142 ]
Анализ вымерших адапариформ посткраниальные скелеты предполагают различное локомоторное поведение. [ 142 ] Европейские адапиды адапи , палеолемер и лептадапис обменивались адаптацией для медленного скалолазания, как лорисы, хотя они, возможно, были четвероногими бегунами, такими как маленькие обезьяны нового мира. И Notharctus , и Smidodectes из Северной Америки и Europolemur из Европы демонстрируют пропорции конечности и совместные поверхности, сопоставимые с вертикальными липкими и подпрыгивающими лемурами, но не были такими специализированными, как инвентариды для вертикального цепляния, что предполагало, что они бежали по ветвям и не прыгали так сильно. [ 91 ] [ 142 ] Notharctids Cantius и Pronycticebus, по -видимому, были как мугильные дендрарийские четвероноги, с адаптацией, сопоставимыми с коричневыми лемурами. [ 142 ]
Диета
[ редактировать ]Приматы в основном питаются фруктами (включая семена), листья (включая цветы) и добычу животных ( членистоногие , мелкие позвоночные и яйца). Диеты заметно различаются между видами стрепсиргина. Как и другие листовые ( фоливовые ) приматы, некоторые стрепсиррины могут переваривать целлюлозу и гемицеллюлозу . [ 143 ] Некоторые стрепсиррины, такие как галаго, стройные лорисы и angwantibos , в первую очередь насекомые . Другие виды, такие как лемуры, отмеченные вилкой , и кустарники, когтя , специализируются на деревьях, в то время как индуриды, спортивные лемуры и бамбуковые лемуры-фоливоры. Многие стрепсиррины-это фругиваторы (едоки фруктов), а другие, такие как лемуры с кольцевым хвостом и мыши, являются всеядными , есть смесь фруктов, листьев и животных. [ 144 ]
Среди адапариформ Frugivory, по-видимому, была наиболее распространенной диетой, особенно для среднего размера до крупных видов, таких как Cantius , Pelycodus и Cercamonius . [ 144 ] Фоливори также была распространена среди адапиформ среднего и большого размера, включая Smilodectes , Notharctus , Adapis и Leptadapis . Острые острые острова на зубах некоторых из меньших адапатиформ, таких как анхомоми , и Donrussellia , указывают на то, что они были частично или в первую очередь насекомые. [ 91 ] [ 144 ]
Распределение и среда обитания
[ редактировать ]В настоящее время вымершие адапиформные приматы были в основном обнаружены в Северной Америке, Азии и Европе, с несколькими видами в Африке. Они процветали во время эоцена, когда эти регионы носили более тропический характер, и они исчезли, когда климат стал прохладным и сухой. [ 36 ] Сегодня лемуриформны ограничены в тропиках, [ 145 ] в диапазоне от 28 до 26 ° С. [ 16 ] Лорисы встречаются как в экваториальной Африке , так и в Юго-Восточной Азии , в то время как галаго ограничены лесами и лесами Африки к югу от Сахары . [ 17 ] [ 97 ] Лемуры эндемичны для Мадагаскара, хотя большая часть их разнообразия и среды обитания была потеряна из -за недавней человеческой деятельности. [ 17 ] [ 50 ]
Как и почти со всеми приматами, Strepsirrhines обычно проживают в тропических тропических лесах . Эти среды обитания позволяют стрепсамрринам и другим приматам развивать разнообразные сообщества симпатрических видов . В восточных тропических лесах Мадагаскара, до 11 или 12 видов имеют одни и те же леса, и до прибытия человека в некоторых лесах почти вдвое превзошло это разнообразие. [ 145 ] Несколько видов лемура встречаются в более сухих, сезонных лесах, включая колючий лес на южной оконечности острова, хотя общины лемуров в этих регионах не так богаты. [ 146 ]
Сохранение
[ редактировать ]
Как и все другие нечеловеческие приматы, Strepsirrhines сталкиваются с повышенным риском вымирания из-за человеческой деятельности, в частности, вырубки лесов в тропических регионах. Большая часть их среды обитания была преобразована для использования человеком, такого как сельское хозяйство и пастбища. [ 27 ] Угрозы, с которыми сталкиваются приматы стрепсиргина, делятся на три основные категории: разрушение среды обитания , охота (для бушковой или традиционной медицины ) и живой захват экспорта или местной экзотической торговли домашним животным. Хотя охота часто запрещена, законы, защищающие их, редко применяются. На Мадагаскаре местные табу, известные как Фади, иногда помогают защитить виды лемуров, хотя некоторые все еще охотятся за традиционной медициной. [ 147 ]
В 2012 году Международный союз сохранения природы (МСОП) объявил, что лемуры были «самыми находящимися под угрозой исчезновения млекопитающих», в значительной степени из -за повышения незаконных логотиков и охоты после политического кризиса в 2009 году . [ 148 ] В Юго -Восточной Азии медленные лорисы угрожают экзотическую торговлю домашними животными и традиционную медицину, в дополнение к разрушению среды обитания. [ 149 ] [ 150 ] Как лемуры, так и медленные лорисы защищены от коммерческой международной торговли в соответствии с приложением CITES I. [ 151 ]
Пояснительные заметки
[ редактировать ]- ^ Jump up to: а беременный Хотя монофилетические отношения между лемурами и лоризоидами широко распространены, их имя клады не является. Термин Lemuriform используется здесь, потому что он вытекает из одной популярной таксономии, которая превращает кладу приматов зубьев в один инфратор и вымершие, не зубчатые адапариформы в другую, как в подряд Стрепсиррини. [ 2 ] [ 3 ] Тем не менее, другая популярная альтернативная таксономия помещает лоризоиды в свой собственный недостаток, Lorisiformes. [ 4 ]
- ^ « Клада » - это группа, состоящая из предка и всех его потомков.
- ^ Колуго также называют дермоптеранами и летающими лемурами .
- ^ Адапариформ иногда называют адапидами или адапоидами .
- ^ Омомиформ как или омомииды, предполагается, были тесно связаны с держиками .
- ^ Поскольку считается, что они являются близкими родственниками копечков, омомиформ классифицируется как гаплорины. Тем не менее, расстояние расстояния корней их верхних резцов предполагает, что у них, возможно, был ринарий, такой как стрепсиррины. [ 37 ] [ 59 ]
- ^ Частые упоминания о «стрипсирринском зубном матче» или ссылках на Strepsirrhini как «зубные приматы» могут быть найдены в литературе. [ 7 ] [ 50 ] Тем не менее, одной группе стрепсирринов не хватает зубов - адапариформ. Таким образом, зуб, является основным отличительным признаком лемурообразных, [ 37 ] Хотя по крайней мере одна семья сохраняет его только в модифицированной форме. [ 61 ]
Ссылки
[ редактировать ]Цитаты
[ редактировать ]- ^ «Стрепсиррини» . paleobiodb.org . Получено 2021-07-22 .
- ^ Szalay & Delson 1980 , p. 149
- ^ Jump up to: а беременный Cartmill 2010 , с. 15
- ^ Jump up to: а беременный Hartwig 2011 , с. 20–21.
- ^ στρέέές , ῥῥς . Лидделл, Генри Джордж ; Скотт, Роберт ; Грек -английский лексикон в проекте Персея .
- ^ Angle-Simons 2007 , с.
- ^ Jump up to: а беременный в дюймовый и фон глин Vaughan, Ryan & Czaplewski 2011 , с.
- ^ Осман Хилл 1953 , с. 39
- ^ Джеффрой Сен-Хилер 1812 , с. 156
- ^ Греческий rho ⟨ρ⟩ r написан с Spiritus asper , когда слово инициал, то есть rh , и часто удваивается до ⟨ῤῥ⟩ rrh (также написано ⟨ρρ⟩), когда такое слово представляет собой вторую часть составное слово . См. Древней греческой фонологии #жидкости , #побужденных согласных и #Консонанта .
- ^ Jump up to: а беременный Groves 2008 , с. 166
- ^ Pocock 1918 , p. 51
- ^ Jenkins & Napier 1987 , p. 1
- ^ Jump up to: а беременный в Роуз 2006 , с. 166
- ^ Rose 2006 , с. 157 и 165–167.
- ^ Jump up to: а беременный в дюймовый и Whitten & Brockman 2001 , p. 322.
- ^ Jump up to: а беременный в дюймовый и Hartwig 2011 , с. 29
- ^ Jump up to: а беременный Fichtel & Kappeler 2009 , p. 397.
- ^ Jump up to: а беременный Hartwig 2011 , с. 28–29.
- ^ Борода 2002 , с. 133.
- ^ Саймонс 2003 , с. 15–17.
- ^ Jump up to: а беременный в Год 2006 , с. 446.
- ^ Jump up to: а беременный Роуз 2006 , с. 343.
- ^ Rose 2006 , с. 168–169.
- ^ Jump up to: а беременный в Hartwig 2011 , с. 22
- ^ Hartwig 2011 , с. 24
- ^ Jump up to: а беременный Cartmill & Smith 2011 , с. 84
- ^ Роза 2006 , с. 169
- ^ Роза 2006 , с. 165.
- ^ Miller, Gunnell & Martin 2005 , p.
- ^ Jump up to: а беременный Роуз 2006 , с. 178–179.
- ^ Jump up to: а беременный в дюймовый и фон Роуз 2006 , с. 179
- ^ Covert 2002 , с. 15–16.
- ^ Jump up to: а беременный в Gebo 2002 , p. 21
- ^ Covert 2002 , с. 14
- ^ Jump up to: а беременный в Gebo 2002 , p. 35
- ^ Jump up to: а беременный в дюймовый и фон Роуз 2006 , с. 182.
- ^ Rose 2006 , с. 185–186.
- ^ Rose 2006 , с. 182–185.
- ^ В Yearinot 1998 , p. 239
- ^ Gunnell, Rose & Rasmussen 2008 , с.
- ^ Jump up to: а беременный Роуз 2006 , с. 186
- ^ Steiper & Seiffert 2012 , с. 6006.
- ^ Роза 2006 , с. 182 и 186.
- ^ Jump up to: а беременный в Tabuce et al. 2009 , с. 4091–4092.
- ^ Hartwig 2011 , с. 24–25.
- ^ Jump up to: а беременный в Vaughan, Ryan & Czaplewski 2011 , с. 170–171.
- ^ Роза 2006 , с. 187.
- ^ Jump up to: а беременный в Роуз 2006 , с. 185.
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м Cartmill & Smith 2011 , с. 89
- ^ Groves 2008 , с. 89–90 и 96.
- ^ Groves 2008 , p. 103
- ^ Jump up to: а беременный в дюймовый Schwartz 2003 , с. 53–54.
- ^ Groves 2008 , с. 113–114.
- ^ Jump up to: а беременный Hartwig 2011 , с. 20 и 22.
- ^ Sussman 2003 , p. 45
- ^ Schwartz 2003 , p. 54
- ^ Angle-Simons 2007 , с.
- ^ Борода 1988 , с. 92
- ^ Jump up to: а беременный Hartwig 2011 , с. 28
- ^ Angle-Simons 2007 , p.
- ^ Schwartz 2003 , p. 71
- ^ Angle-Simons 2007 , p.
- ^ Cartmill 2010 , с. 16–17.
- ^ Rasmussen & Nekaris 1998 , p.
- ^ Ancle-Simons 2007 , с. 31–32.
- ^ Groves 1998 , p. 13
- ^ Роза 2006 , с. 167
- ^ Jump up to: а беременный Beard 2002 , с. 146–147.
- ^ Gingerich 1975 , p. 164.
- ^ Yoder 2003 , с. 161–162.
- ^ Jump up to: а беременный в Williams et al. 2010 , с. 567.
- ^ Franzen et al. 2009 , с. E5723.
- ^ Jump up to: а беременный в Cartmill & Smith 2011 , с. 90
- ^ Hartwig 2011 , с. 20
- ^ Groves 2005 , p. 121.
- ^ Godfrey & Jungers 2002 , P. 106
- ^ Godfrey & Jungers 2002 , P. 97
- ^ Роза 2006 , с. 181.
- ^ Jump up to: а беременный в Covert 2002 , p. 18
- Dunkel , Zijlstra & Groves 2012 , с. 68
- ^ Jump up to: а беременный в Mittermeier, Rylands & Constant 1999 , p. 4
- ^ Jump up to: а беременный в Осман Хилл 1953 , с. 96
- ^ Gebo 2002 , p. 39
- ^ Cuozzo & Yamashita 2006 , p. 73.
- ^ Jump up to: а беременный в Роуз 2006 , с. 166–167.
- ^ Jump up to: а беременный в дюймовый Tattersall 2006 , p. 7
- ^ Ollivier et al. 2004 .
- ^ Angle-Simons 2007 , p.
- ^ Mittermeier, Rylands & Constant 1999 , p. 6
- ^ Jump up to: а беременный в дюймовый и Роуз 2006 , с. 180.
- ^ Роза 2006 , с. 184.
- ^ Godfrey & Jungers 2002 , стр. 106 и 112.
- ^ Angle-Simons 2007 , p.
- ^ Jump up to: а беременный Mittermeier, Rylands & Constant 1999 , с. 4–5.
- ^ Angle-Simons 2007 , p.
- ^ Jump up to: а беременный в Cartmill & Smith 2011 , с. 91
- ^ Дагосто 1988 , с. 47–48.
- ^ Дагосто 1988 , с. 49
- ^ Angle-Simons 2007 , с.
- ^ Jump up to: а беременный Whitten & Brockman 2001 , p. 323.
- ^ Rose 2006 , с. 168 и 180.
- ^ Plavcan 2004 , p.
- ^ Angle-Simons 2007 , с.
- ^ Jump up to: а беременный Angle-Simons 2007 , с. 392–394.
- ^ Jump up to: а беременный в дюймовый и фон Mittermeier, Rylands & Constant 1999 , с. 24–25.
- ^ Angle-Simons 2007 , с.
- ^ Борода 1988 , с. 85
- ^ Ankel-Simons 2007 , с. 392–393 и 400–401.
- ^ Jump up to: а беременный Angle-Simons 2007 , с. 392–393.
- ^ Angle-Simons 2007 , с.
- ^ Angle-Simons 2007 , с.
- ^ Angle-Simons 2007 , с.
- ^ Ancle-Simons 2007 , с. 410–411.
- ^ Mittermeier, Rylands & Constant 1999 , p. 26
- ^ Rose 2006 , с. 181–182.
- ^ Jump up to: а беременный Whitten & Brockman 2001 , с. 325–326.
- ^ Whitten & Brockman 2001 , с. 330–331.
- ^ Cartmill & Smith 2011 , с. 88
- ^ Осман Хилл 1953 , с. 81.
- ^ Осман Хилл 1953 , с. 93.
- ^ Осман Хилл 1953 , с. 391.
- ^ Осман Хилл 1953 , с. 672.
- ^ Angle-Simons 2007 , с.
- ^ Angle-Simons 2007 , с.
- ^ Angle-Simons 2007 , p.
- ^ Jump up to: а беременный в Mittermeier, Rylands & Constant 1999 , с. 4–6.
- ^ Mittermeier, Rylands & Constant 1999 , p. 5
- ^ Whitten & Brockman 2001 , с. 325 и 335.
- ^ Faell & Helperer 2009 , стр. 395-396.
- ^ Faell & Helperer 2009 , стр. 401-402.
- ^ Jump up to: а беременный Nekaris & Bearder 2011 , с. 51
- ^ Jump up to: а беременный Mittermeier, Rylands & Constant 1999 , с. 26–27.
- ^ Gould, Sauther & Cameron 2011 , с. 74
- ^ Fichtel & Kappeler 2009 , с. 395 и 397.
- ^ Fichtel & Kappeler 2009 , p. 398.
- ^ Mittermeier, Rylands & Constant 1999 , с. 15–16.
- ^ Overdorff & Tecot 2006 , p. 247
- ^ Cloninger & Kedia 2011 , с. 86
- ^ Mittermeier, Rylands & Constant 1999 , с. 25–26.
- ^ Mittermeier, Rylands & Constant 1999 , с. 5 и 26.
- ^ Jump up to: а беременный в дюймовый и фон глин Covert 2002 , с. 16–17.
- ^ Mittermeier, Rylands & Constant 1999 , с. 22–24.
- ^ Jump up to: а беременный в Covert 2002 , p. 16
- ^ Jump up to: а беременный Mittermeier, Rylands & Constant 1999 , p. 14
- ^ Mittermeier, Rylands & Constant 1999 , p. 19
- ^ Mittermeier, Rylands & Constant 1999 , с. 32–34.
- ^ «Лемуры обнаружили, что являются« самыми угрожающими млекопитающими »в мире» . CNN. 13 июля 2012 года. Архивировано с оригинала 21 августа 2012 года . Получено 28 сентября 2012 года .
- ^ Fitch-Snyder & Livingstone 2008 .
- ^ Nekaris et al. 2010 , с. 878.
- ^ «Приложения I, II и III» . Конвенция о международной торговле исчезающими видами дикой фауны и флоры. 25 сентября 2012 года. Архивировано с оригинала 9 октября 2012 года . Получено 1 октября 2012 года .
Литература цитируется
[ редактировать ]- Ankel-Simons, F. (2007). Анатомия приматов (3 -е изд.). Академическая пресса. ISBN 978-0-12-372576-9 .
- Beard, KC (1988). «Филогенетическая значимость стрепсаргинизма у палеогеновых приматов». Международный журнал приматологии . 9 (2): 83–96. doi : 10.1007/bf02735730 . S2CID 32127974 .
- Кэмпбелл, CJ; Fuentes, A.; Mackinnon, KC; Bearder, SK; Stumpf, R. M, Eds. (2011). Приматы в перспективе (2 -е изд.). Издательство Оксфордского университета. ISBN 978-0-19-539043-8 .
- Gould, L.; Sauther, M.; Кэмерон, А. "Глава 5: Lemuriformes". В Campbell et al. (2011) , с. 55–79.
- Хартвиг В. "Глава 3: Эволюция приматов". В Campbell et al. (2011) , с. 19–31.
- Nekaris, Nai; Берег, С.К. "Глава 4: Лоризиформные приматы Азии и материковой Африки: разнообразие, окутанное темнотой". В Campbell et al. (2011) , с. 34–54.
- Cartmill, M. (2010). «Классификация приматов и разнообразие» . В Платт, М.; Газанфар, А. (ред.). Примат нейроэтология . Издательство Оксфордского университета. С. 10–30. ISBN 978-0-19-532659-8 .
- Cartmill, M.; Смит, Ф.Х. (2011). Человеческая родословная . Джон Уайли и сыновья. ISBN 978-1-118-21145-8 .
- Клонингер, CR; Kedia, S. (2011). «Глава 5: Филогенез человеческой личности: выявление предшественников сотрудничества, альтруизма и благополучия» . В Суссмане, RW; Клонингер, кр (ред.). Происхождение альтруизма и сотрудничества . Спрингер. ISBN 978-1-4419-9519-3 .
- Дагосто М. (1988). «Последствия посткраниальных доказательств для происхождения эуприматов» . Журнал человеческой эволюции . 17 (1–2): 35–77. doi : 10.1016/0047-2484 (88) 90048-6 .
- Дункель, Ар; Zijlstra, JS; Groves, CP (2012). «Гигантские кролики, мармозты и британские комедии: этимология имен лемуров, часть 1» (PDF) . Lemur News . 16 : 64–70. ISSN 1608-1439 . Архивировано из оригинала (PDF) 2016-11-06 . Получено 2012-09-28 .
- Fitch-Snyder, H.; Ливингстон, К. (2008). «Лориз: неожиданный примат». Zoonooz . С. 10–14. ISSN 0044-5282 .
- Fichtel, C.; Каппелер, PM (2009). «Глава 19: Человеческие универсалии и приматы симппсуморфии: установление базовой линии лемура» . В Каппелере, PM; Шелк, JB (ред.). Имейте в виду разрыв: отслеживание происхождения человеческих универсалий . Спрингер. С. 395–426. ISBN 978-3-642-02724-6 .
- Franzen, JL; Gingerich, PD; Habersetzer, J.; Хурум, JH; фон Кенигсвальд, W.; Смит, BH (2009). «Полный скелет приматов из среднего эоцена Месселя в Германии: морфология и палеобиология» . Plos один . 4 (5): E5723. Bibcode : 2009ploso ... 4.5723f . doi : 10.1371/journal.pone.0005723 . PMC 2683573 . PMID 19492084 .
- Джеффрой Сен-Хилер, é. (1812). «Следуя за таблицей четырехлетних. Вторая семья. Лемурийцы. Стрепсиррини» . Анналы музея естественной истории (на французском языке). 19 : 156–170.
- Gingerich, PD (1975). «Новый род адапид (млекопитающие, приматы) из позднего эоцена Южной Франции и его значение для происхождения более высоких приматов». Вклад музея палеонтологии . 24 (15). Мичиганский университет: 163–170. HDL : 2027.42/48482 .
- Годино, М. (1998). «Резюме адапиформной систематики и филогения». Folia Primatologica . 69 (Suppl. 1): 218–249. doi : 10.1159/000052715 . S2CID 84469830 .
- Годино, М. (2006). «Lemuriform Origins, как рассматривалось из ископаемой записи». Folia Primatologica . 77 (6): 446–464. doi : 10.1159/000095391 . PMID 17053330 . S2CID 24163044 .
- Gould, L.; Sauther, ML, eds. (2006). Лемуры: экология и адаптация . Спрингер. ISBN 978-0-387-34585-7 .
- Cuozzo, FP; Yamashita, N. « Глава 4: Влияние экологии на зубы существующих лемуров: обзор адаптации зубов, функции и истории жизни ». В Gould & Sauther (2006) , стр. 67–96.
- Tattersall, I. " Глава 1: Происхождение малагасийских стрепсирхин приматов ". В Gould & Sauther (2006) , с. 3–18.
- Groves, CP (1998). «Систематика тарси и лорисов». Приматы . 39 (1): 13–27. doi : 10.1007/bf025557740 . S2CID 10869981 .
- Groves, CP (2005). «Стрепсиррини» . В Уилсоне, де ; Ридер, DM (ред.). Виды млекопитающих мира: таксономическая и географическая ссылка (3 -е изд.). Балтимор: издательство Джона Хопкинса. п. 111. ISBN 0-801-88221-4 Полем OCLC 62265494 .
- Groves, CP (2008). Расширенная семья: давно потерянные кузены: личный взгляд на историю приматологии . Сохранение International. ISBN 978-1-934151-25-9 .
- Ганнелл, GF; Роуз, КД; Расмуссен, Д.Т. (2008). "Euprimates". В Janis, CM; Ганнелл, GF; Ухен, MD (ред.). Маленькие млекопитающие, ксенартраны и морские млекопитающие . Эволюция третичных млекопитающих Северной Америки. Тол. 2. Кембридж, Англия; Нью -Йорк: издательство Кембриджского университета. С. 239–262. ISBN 978-0-521-78117-6 .
- Hartwig, WC, ed. (2002). Запись окаменелости приматов . Издательство Кембриджского университета. ISBN 978-0-521-66315-1 .
- Борода, KC « Глава 9: базальные антропоиды ». В Hartwig (2002) , с. 133–149.
- Covert, HH " Глава 3: Самые ранние окаменелости и эволюция проницистов: введение ". В Хартвиге (2002) , с. 13–20.
- Gebo, DL « Глава 4: Адапориформа: филогения и адаптация ». В Hartwig (2002) , с. 21–44.
- Годфри, LR; Jungers, WL « Глава 7: Четвертичные ископаемые лемуры ». В Hartwig (2002) , с. 97–121.
- Дженкинс, PD; Нейпир, PH (1987). Стрепсиррини, в том числе подконтрольные мадагасканские лемуры и семейные Tarsiidae Каталог приматов в Британском музее (естественная история) и в других местах Британских островов. Тол. Часть 4. Британский музей (естественная история). ISBN 978-0-565-01008-9 .
- Миллер, эр; Ганнелл, GF; Martin, RD (2005). «Глубокое время и поиск антропоидных происхождений» (PDF) . Американский журнал физической антропологии . 128 : 60–95. doi : 10.1002/ajpa.20352 . PMID 16369958 .
- Mittermeier, ra; Rylands, AB; Konstant, WR (1999). «Приматы мира: введение» . В Новаке, Р.М. (ред.). Уокерские млекопитающие мира (6 -е изд.). Johns Hopkins University Press. С. 1–52. ISBN 978-0-8018-6251-9 .
- Некарис, Кай; Шепард, Кр; Старр, Кр; Nijman, V. (2010). «Изучение культурных драйверов для торговли дикой природой с помощью этноприматологического подхода: тематическое исследование стройных и медленных лорисов ( Loris и Nycticebus ) в Южной и Юго -Восточной Азии». Американский журнал приматологии . 72 (10): 877–886. doi : 10.1002/ajp.20842 . PMID 20806336 . S2CID 21711250 .
- Ollivier, FJ; Самуэльсон, да; Брукс, де; Льюис, Пенсильвания; Каллберг, я; Комароми, А.М. (2004). «Сравнительная морфология таптума Lucidum (среди отдельных видов)». Ветеринарная офтальмология . 7 (1): 11–22. doi : 10.1111/j.1463-5224.2004.00318.x . PMID 14738502 . S2CID 15419778 .
- Осман Хилл, WC (1953). "Я - Стрепсирхини". Приматы сравнительная анатомия и таксономия . Edinburgh Univ. Пабы. Наука и математика. Тол. 3. Edinburgh University Press. OCLC 500576914 .
- Overdorff, DJ; Tecot, SR (2006). «Глава 11: Социальные пары и защита ресурсов в диких красных лемурах ( Eulemur Rubriventer )» . В Gould, L.; Sauther, ML (ред.). Лемуры: экология и адаптация . Спрингер. С. 235–254. doi : 10.1007/978-0-387-34586-4_11 . ISBN 978-0-387-34585-7 Полем S2CID 85613386 .
- Plavcan, JM (2004). «Глава 13: Сексуальный отбор, меры сексуального отбора и половой диморфизм у приматов» . В Каппелере, PM; Ван Шайк, CP (ред.). Сексуальный отбор у приматов: новые и сравнительные перспективы . Издательство Кембриджского университета. С. 230–252. ISBN 978-0-521-53738-4 .
- Pocock, RI (1918). «На внешних символах лемуров и тарзия » . Труды зоологического общества Лондона . 1918 (1–2): 19–53. doi : 10.1111/j.1096-3642.1918.tb02076.x .
- Расмуссен, DT; Nekaris, Kai (1998). «Эволюционная история лоризиформных приматов» (PDF) . Folia Primatologica . 69 (Suppl. 1) (7): 250–285. doi : 10.1159/000052716 . PMID 9595692 . S2CID 14113728 .
- Роуз, К.Д. (2006). Начало эпохи млекопитающих . Johns Hopkins University Press. ISBN 978-0-8018-8472-6 .
- Steiper, я; Seiffert, ER (2012). «Доказательства конвергентного замедления молекулярных скоростей приматов и его последствий для времени ранней эволюции приматов» . Труды Национальной академии наук . 109 (16): 6006–6011. Bibcode : 2012pnas..109.6006S . doi : 10.1073/pnas.1119506109 . PMC 3341044 . PMID 22474376 .
- Sussman, RW (2003). Экология приматов и социальная структура . Pearson Custom Publishing. ISBN 978-0-536-74363-3 .
- Szalay, FS; Delson, E. (1980). Эволюционная история приматов . Академическая пресса . ISBN 978-0-12-680150-7 Полем OCLC 893740473 .
- Tabuce, R.; Marivaux, L.; Lebrun, R.; Adaci, M.; Бенсала, М.; Fabre, P.-H.; и др. (2009). «Статус антропоида против стрепсиргина африканских приматов эоцена Algeripithecus и Azibius : краниодиентационные доказательства» . Труды Королевского общества B: Биологические науки . 276 (1676): 4087–4094. doi : 10.1098/rspb.2009.1339 . PMC 2821352 . PMID 19740889 .
- «Африканское происхождение антропоидных приматов ставит под сомнение с новым обнаружением ископаемых» . Science Daily (пресс -релиз). 17 сентября 2009 г.
- Воган, Т.; Райан, Дж.; Czaplewski, N. (2011). «Глава 12: Приматы» . МАММАЛОГИ (5 -е изд.). Jones & Bartlett Learning. ISBN 978-0-7637-6299-5 .
- Уиттен, PL; Брокман, Д.К. (2001). «Глава 14: репродуктивная экология Strepsirrhine» . В Эллисон, Pt (ed.). Репродуктивная экология и эволюция человека . ПРИБОРЫ Издатели. С. 321–350. ISBN 978-0-202-30658-2 .
- Уильямс, Б.А.; Кей, RF; Кристофер Кирк, E.; Росс, CF (2010). « Darwinius Masillae - это Strepsrrhine - ответ Franzen et al. (2009)» (PDF) . Журнал человеческой эволюции . 59 (5): 567–573, обсуждение 573–579. doi : 10.1016/j.jhevol.2010.01.003 . PMID 20188396 . Архивировано из оригинала (PDF) 17 мая 2013 года.
- Райт, Патриция С.; Саймонс, Элвин Л., ред. (2003). Tarsiers: прошлое, настоящее и будущее . Рутгерс Университет Пресс. ISBN 978-0-8135-3236-3 .
- Schwartz, JH « Глава 3: Насколько близко сходства между Tarsius и другими приматами? ». В Wright & Simons (2003) , с. 50–96.
- Саймонс, Эль " Глава 1: ископаемое запись о цирсие эволюции ". В Wright & Simons (2003) , стр. 9–34.
- Йодер, ad « Глава 7: Филогенетическое положение рода Tarsius : на чьи стороны вы? ». В Wright & Simons (2003) , с. 161–175.
Внешние ссылки
[ редактировать ] СМИ, связанные со Стрепсрихини в Wikimedia Commons
СМИ, связанные со Стрепсрихини в Wikimedia Commons  Данные, связанные со Strepsirrhini в Wikispecies
Данные, связанные со Strepsirrhini в Wikispecies


| Базы данных управления авторитетом : национальный |
|---|