Антисмысловая РНК

Антисмысловая РНК ( асРНК ), также называемая антисмысловым транскриптом, [ 1 ] природный антисмысловой транскрипт (NAT) [ 2 ] [ 3 ] [ 4 ] или антисмысловой олигонуклеотид , [ 5 ] представляет собой одноцепочечную РНК , которая комплементарна белку, кодирующему информационную РНК (мРНК), с которой она гибридизуется, и тем самым блокирует ее трансляцию в белок . АсРНК (которые встречаются в природе) были обнаружены как у прокариот , так и у эукариот . [ 1 ] Их можно разделить на короткие (<200 нуклеотидов) и длинные (>200 нуклеотидов) некодирующие РНК (нкРНК). [ 4 ] Основная функция asRNA – регуляция экспрессии генов . asRNA также могут быть получены синтетическим путем и нашли широкое применение в качестве исследовательских инструментов для нокдауна генов . Они также могут иметь терапевтическое применение. [ 6 ] [ 1 ] [ 4 ]
Открытие и история разработки лекарств
[ редактировать ]Некоторые из самых ранних asRNA были обнаружены при исследовании функциональных белков. Примером была micF asRNA . внешней мембраны При характеристике порина ompC в E.coli некоторые из наблюдаемых клонов промотора ompC были способны подавлять экспрессию другого мембранного порина, такого как ompF. Было обнаружено, что область, ответственная за эту функцию репрессии, представляет собой локус длиной 300 пар оснований выше промотора ompC. Эта область из 300 пар оснований на 70% гомологична по последовательности 5'-концу мРНК ompF и, таким образом, транскрипт этого локуса из 300 пар оснований комплементарен мРНК ompF. Позже было обнаружено, что этот транскрипт, обозначенный micF, представляет собой asRNA ompF и способен подавлять экспрессию ompF в условиях стресса путем образования дуплекса с мРНК ompF. Это вызывает деградацию мРНК ompF. [ 2 ]
В отличие от РНК micF, обнаруженной случайно, большинство asRNA были обнаружены в результате полногеномного поиска малых регуляторных РНК и анализа транскриптома . Традиционно первый шаг включает в себя вычислительные прогнозы, основанные на некоторых известных характеристиках асРНК. Во время вычислительного поиска области кодирования исключаются. регионам, которые, по прогнозам, имеют консервативные структуры РНК и действуют как сиротские промоторы и Rho-независимые терминаторы Во время анализа предпочтение отдается . Поскольку компьютерный поиск сосредоточен на межгенной области , asRNA, которые транскрибируются с противоположной цепи кодирующего гена, скорее всего, будут пропущены с помощью этого метода. Для обнаружения asRNA, транскрибируемой из кодирующей области, микрочипы олигонуклеотидов можно использовать . В этом методе в качестве зондов можно использовать одну или обе цепи кодирующих генов. Помимо компьютерного поиска и микрочипов, некоторые asRNA были обнаружены путем секвенирования клонов кДНК, а также картирования промоторных элементов. [ 7 ] Хотя многие результаты упомянутых выше подходов привели к появлению множества возможных asRNA, лишь немногие из них оказались настоящими asRNA с помощью дальнейших функциональных тестов. Чтобы свести к минимуму количество ложноположительных результатов, новые подходы последних лет были сосредоточены на специфической транскрипции, связывании хроматина некодирующих РНК и исследованиях отдельных клеток. [ 1 ]
Идея использования asRNA в качестве мишени для лекарств возникла в 1978 году, когда Замечник и Стивенсон обнаружили антисмысловой олигонуклеотид по отношению к вирусной РНК вируса скаркомы Рауса, который был способен ингибировать репликацию вируса и синтез белка. С тех пор много усилий было посвящено разработке asRNA в качестве потенциальных лекарств. первый препарат asRNA — фомивирсен В 1998 году FDA одобрило . Фомивирсен, олигонуклеотид из 21 пары оснований, был разработан для лечения цитомегаловирусного ретинита у пациентов со СПИДом. Он действует путем воздействия на транскрибируемую мРНК вируса и, следовательно, ингибирования репликации цитомегаловируса. Несмотря на то, что фомивирсен был снят с производства в 2004 году из-за потери рынка, он послужил успешным и вдохновляющим примером использования асРНК в качестве мишеней для лекарств или кандидатов на лекарства. [ 5 ]
Другим примером использования асРНК в качестве терапевтического средства является мипомерсен , который был одобрен FDA в 2013 году. Мипомерсен был разработан для контроля уровня холестерина липопротеинов низкой плотности (ЛПНП) у пациентов с гомозиготной семейной гиперхолестеринемией (HoFH), которая является редкое аутосомно-доминантное генетическое заболевание. Из-за высокого уровня общего холестерина (650–1000 мг/дл) и рецепторов ЛПНП (более 600 мг/дл) при ГСГ пациенты с ГСГ имеют высокий риск развития ишемической болезни сердца. Поскольку было обнаружено, что белок апо-B-100 необходим для производства липопротеинов очень низкой плотности (ЛПОНП) и ЛПНП, мипомерсен дополняет мРНК апо-B-100 и нацеливает ее на РНКазы H. деградацию, зависимую от В конечном итоге мипомерсен способен снижать уровень ЛПНП. [ 8 ]
Примеры разных видов
[ редактировать ]Первые обнаруженные asRNA были у прокариот, включая плазмиды , бактериофаги и бактерии . Например, в плазмиде ColE1 asRNA, называемая РНК I, играет важную роль в определении количества копий плазмиды путем контроля репликации. Репликация ColE1 основана на транскрипции праймерной РНК, называемой РНК II. После транскрипции РНК II она гибридизуется со своей матрицей ДНК и позже расщепляется РНКазой H. В присутствии РНК I, РНК I и РНК II образуют дуплекс, который приводит к конформационному изменению РНК II. Следовательно, РНК II не может гибридизоваться со своей матрицей ДНК, что приводит к низкому числу копий ColE1. В бактериофаге P22 asRNA sar помогает регулировать литический и лизогенный циклы, контролируя экспрессию Ant. [ 9 ] АсРНК были обнаружены не только у прокариот, но и у растений. Наиболее хорошо описанный пример регуляции asRNA у растений - ген цветущего локуса C (FLC) . Ген FLC у Arabidopsis thaliana кодирует фактор транскрипции, который предотвращает экспрессию ряда генов, индуцирующих переход цветка. В холодной среде asRNA гена FLC, обозначенная COOLAIR, экспрессируется и ингибирует экспрессию FLC посредством модификации хроматина, что, следовательно, обеспечивает цветение. [ 10 ] Другим хорошо изученным примером является ген DOG1 (задержка прорастания 1). Уровень его экспрессии негативно регулируется антисмысловым транскриптом (asDOG1 или 1GOD), действующим в цис. [ 11 ] В клетках млекопитающих типичным примером регуляции asRNA является инактивация Х-хромосомы. Xist , asRNA, может рекрутировать полисотовый репрессивный комплекс 2 (PRC2), что приводит к гетерохроматинизации Х-хромосомы. [ 3 ]
Классификация
[ редактировать ]Антисмысловые РНК можно классифицировать по-разному. С точки зрения регуляторных механизмов некоторые авторы группируют asRNA на взаимодействия РНК-ДНК, взаимодействия РНК-РНК в ядре или цитоплазме и взаимодействия РНК-белок ( эпигенетические ). [ 3 ] Антисмысловые РНК можно классифицировать по типу промоторов, которые инициируют экспрессию asРНК: независимые промоторы, общие двунаправленные промоторы или загадочные промоторы. Что касается длины, хотя asRNA в целом классифицируется как lncRNA , существуют короткие asRNA длиной менее 200 пар оснований. Поскольку установлено, что регуляторный механизм asRNA является видоспецифичным, asRNA также можно классифицировать по видам. [ 1 ] Один из распространенных способов классификации asRNA заключается в том, где asRNA транскрибируются относительно их генов-мишеней: цис-действующие и транс-действующие . [ нужна ссылка ]
Цис-действующий
[ редактировать ]Цис-действующие asRNA транскрибируются с противоположной цепи целевого гена в локусе целевого гена. Они часто демонстрируют высокую степень или полную комплементарность целевому гену. Если цис-действующая асРНК регулирует экспрессию генов, нацеливаясь на мРНК, она может нацеливаться только на индивидуальную мРНК. При взаимодействии с нацеливающими мРНК цис-действующие асРНК могут либо блокировать связывание рибосом, либо рекрутировать РНКазу для разрушения нацеливающих мРНК. Следовательно, функция этих цис-действующих asRNA заключается в подавлении трансляции нацеливающих мРНК. [ 2 ] Помимо цис-действующих асРНК, которые нацелены на мРНК, существуют цис-действующие эпигенетические сайленсеры и активаторы . Было показано, что антисмысловая РНК подавляет трансляцию домена LINE1-ORF2 Entamoeba histolytica . Однако еще не подтверждено, является ли он цис-действующим или транс-действующим. [ 12 ]
С точки зрения эпигенетической модификации, цис-действие относится к природе этих asRNA, которые регулируют эпигенетические изменения вокруг локусов, где они транскрибируются. Вместо того, чтобы нацеливаться на отдельные мРНК, эти цис-действующие эпигенетические регуляторы могут рекрутировать ферменты, модифицирующие хроматин, которые могут оказывать влияние как на локусы транскрипции, так и на соседние гены. [ 3 ]
Транзакции
[ редактировать ]Транс-действующие asRNA транскрибируются из локусов, которые расположены дистальнее от направляющих генов. В отличие от цис-действующих asRNA, они обладают низкой степенью комплементарности с целевым геном, но могут быть более длинными, чем цис-действующие asRNA. Они также могут быть нацелены на несколько локусов. Из-за этих свойств транс-действующих asRNA они образуют менее стабильные комплексы со своими нацеливающими транскриптами и иногда требуют помощи со стороны белка-шаперона РНК , такого как Hfq, для выполнения своих функций. Из-за сложности транс-действующих асРНК в настоящее время они считаются менее пригодными для применения лекарствами мишенями. [ 2 ]
Функция
[ редактировать ]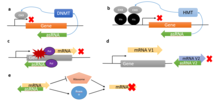
Эпигенетическая регуляция
[ редактировать ]Многие примеры asRNA демонстрируют ингибирующее действие на инициацию транскрипции посредством эпигенетических модификаций.
Метилирование ДНК
[ редактировать ]Метилирование ДНК может привести к долгосрочному подавлению определенных генов. Репрессия функциональных белков посредством метилирования ДНК, индуцированного asRNA, была обнаружена при ряде заболеваний человека. В классе альфа-талассемии — типе заболевания крови, при котором снижается уровень гемоглобина, что приводит к недостаточности кислорода в тканях. [ 13 ] Ген гемоглобина альфа1 (HBA1) подавляется аномальным транскриптом предполагаемого РНК-связывающего белка Luc7-подобного (LUC71), который служит asRNA для HBA1 и индуцирует метилирование промотора HBA1. [ 1 ] Другим примером является подавление гена-супрессора опухоли p15INK4b, также называемого CDKN2B, при остром лимфобластном лейкозе и остром миелоидном лейкозе. АсРНК, которая отвечает за этот эффект молчания, представляет собой антисмысловую некодирующую РНК в локусе INK ( ANRIL ), которая экспрессируется в том же локусе, который кодирует p15INK4b. [ 3 ]
Модификация гистонов
[ редактировать ]В эукариотических клетках ДНК плотно упакована гистонами . Модификация гистонов может изменить взаимодействие с ДНК, что в дальнейшем может вызвать изменения в экспрессии генов . Биологические последствия метилирования гистонов зависят от контекста. В общем, метилирование гистонов приводит к репрессии генов, но также может быть достигнута активация генов. [ 14 ] Данные показали, что метилирование гистонов может быть индуцировано asRNA. Например, ANRIL, помимо способности индуцировать метилирование ДНК, может также репрессировать соседний ген CDKN2B , CDKN2A , путем рекрутирования полисотового репрессивного комплекса 2 (PRC2), который приводит к метилированию гистонов (H3K27me). Другой классический пример — инактивация Х-хромосомы с помощью XIST . [ 1 ]
Эпигенетическая модификация, индуцированная ANRIL, является примером цис-действующей эпигенетической регуляции. [ 3 ] Кроме того, модификация хроматина, индуцированная антисмысловой РНК, может быть как транс-действующей. Например, у млекопитающих asRNA HOTAIR транскрибируется из локуса гомеобокса C (HOXC), но она рекрутирует PRC2 в HOXD, который откладывает H3K27 и подавляет HOXD. HOTAIR высоко экспрессируется в первичных опухолях молочной железы. [ 1 ]
Котранскрипционная регуляция
[ редактировать ]Эпигенетические регуляции, такие как метилирование ДНК и метилирование гистонов, могут подавлять экспрессию генов путем ингибирования инициации транскрипции. Однако иногда репрессия генов может быть достигнута путем преждевременного прекращения или замедления процесса транскрипции. АсРНК могут участвовать в этом уровне регуляции генов. Например, в бактериальных или эукариотических клетках, где присутствуют сложные РНК-полимеразы, двунаправленная транскрипция в одном и том же локусе может привести к столкновению полимераз и привести к прекращению транскрипции. Даже когда столкновение полимеразы маловероятно во время слабой транскрипции, также может произойти пауза полимеразы, которая блокирует элонгацию и приводит к репрессии генов. Одним из примеров является репрессия гена IME4 его asRNA RME2 . Другой способ котранскрипционного воздействия на транскрипцию - блокирование сплайсинга. Одним из классических примеров у человека является с цинковым пальцем ген гомеобокса 2, связывающий E-бокс ( ZEB2 ), который кодирует E-кадгерин, репрессор транскрипции. Эффективная трансляция мРНК ZEB2 требует присутствия внутренний сайт входа рибосомы (IRES) в интроне мРНК на 5'-конце . Экспрессируя asRNA ZEB2, она может маскировать сайт сплайсинга и поддерживать IRES в мРНК, что приводит к эффективному синтезу E-кадгерина. Наконец, в зависимости от уровня экспрессии asRNA могут образовываться разные изоформы смыслового транскрипта. Следовательно, asRNA-зависимая регуляция не ограничивается механизмом включения/выключения; скорее, он представляет собой тонкую систему управления тембром. [ 1 ]
Посттранскрипционная регуляция
[ редактировать ]Прямая посттранскрипционная модуляция с помощью asRNA относится к мРНК, на которые непосредственно нацелены asRNA; таким образом, это влияет на перевод. Некоторые характеристики этого типа асРНК описаны в цис- и транс-действующих асРНК. Этот механизм относительно быстрый, поскольку и направляющая мРНК, и ее асРНК должны одновременно присутствовать в одной и той же клетке. Как описано в цис-действующих asRNA, спаривание мРНК-asRNA может привести к блокировке входа в рибосому и зависимой от РНКазы H деградации. В целом, asRNA, нацеленные на мРНК, могут либо активировать, либо ингибировать трансляцию смысловых мРНК, причем ингибирующий эффект является наиболее распространенным. [ 1 ]
Терапевтический потенциал
[ редактировать ]АсРНК как регуляторный элемент имеют множество преимуществ, поэтому их можно рассматривать в качестве мишени для лекарств. Прежде всего, asRNA регулируют экспрессию генов на нескольких уровнях, включая транскрипцию, посттранскрипцию и эпигенетическую модификацию. Во-вторых, цис-действующие asRNA специфичны к последовательности и демонстрируют высокую степень комплементарности с нацеливающими генами. [ 1 ] В-третьих, уровень экспрессии asRNA очень мал по сравнению с уровнем экспрессии нацеливающих мРНК; следовательно, для достижения эффекта требуется лишь небольшое количество asRNA. С точки зрения целевых показателей лекарств это представляет собой огромное преимущество, поскольку для достижения эффективности требуется лишь низкая дозировка. [ 4 ]
В последние годы большое внимание привлекла идея нацеливания asRNA на усиление экспрессии генов локус-специфическим способом. Из-за характера разработки лекарств всегда легче иметь лекарства, действующие как понижающие регуляторы или ингибиторы. Однако существует потребность в разработке лекарств, которые могут активировать или усиливать экспрессию генов, таких как гены-супрессоры опухолей, нейропротекторные факторы роста и гены, молчание которых обнаруживается при некоторых менделевских расстройствах. В настоящее время подход к восстановлению недостаточной экспрессии генов или функции белка включает заместительную ферментную терапию, терапию микроРНК и доставку функциональной кДНК. Однако каждый из них имеет некоторые недостатки. Например, синтезированный белок, используемый в заместительной ферментной терапии, часто не может имитировать всю функцию эндогенного белка. Кроме того, заместительная ферментная терапия требует пожизненного лечения и несет большое финансовое бремя для пациента. Из-за локус-специфической природы asRNA и доказательств изменений в экспрессии asRNA при многих заболеваниях предпринимались попытки создать одноцепочечные олигонуклеотиды, называемые антагоNAT, для ингибирования asRNA и, в конечном итоге, для увеличения специфической экспрессии генов. [ 4 ]
Несмотря на обещания использовать asRNA в качестве мишеней или кандидатов в лекарственные препараты, остаются некоторые проблемы, требующие решения. [ 15 ] Прежде всего, asRNA и антагоNAT могут легко разрушаться РНКазой или другими деградирующими ферментами. Чтобы предотвратить деградацию терапевтических олиогонуклеотидов, обычно требуется химическая модификация. Наиболее распространенной химической модификацией олигонуклеотидов является добавление тиофосфатной связи к основной цепи. [ 5 ] Однако фосфротиоатная модификация может быть провоспалительной. Побочные эффекты, включая лихорадку, озноб или тошноту, наблюдались после местного введения олигонуклеотидов, модифицированных фосфротиоатом. Во-вторых, большую проблему представляет и нецелевая токсичность. Несмотря на локус-специфическую природу эндогенных asRNA, только 10–50% синтезированных олигонуклеотидов показали ожидаемый таргетный эффект. Одной из возможных причин этой проблемы являются высокие требования к распознаванию структуры asRNA целевой последовательностью и РНКазой H. Единственное несовпадение может привести к искажению вторичной структуры и привести к нецелевым эффектам. [ 4 ] Наконец, было показано, что искусственные asRNA имеют ограниченное внутриклеточное поглощение. [ 5 ] Хотя было показано, что нейроны и глия обладают способностью свободно поглощать голые антисмысловые олигонуклеотиды, прослеживаемые носители, такие как вирусы и липидные везикулы, по-прежнему будут идеальными для контроля и мониторинга внутриклеточной концентрации и метаболизма. [ 4 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к Пелечано В., Штайнмец Л.М. (декабрь 2013 г.). «Регуляция генов посредством антисмысловой транскрипции». Обзоры природы. Генетика . 14 (12): 880–893. дои : 10.1038/nrg3594 . ПМИД 24217315 . S2CID 2152962 .
- ^ Перейти обратно: а б с д Сабери Ф, Камали М, Наджафи А, Язданпараст А, Могаддам ММ (28 июля 2016 г.). «Природные антисмысловые РНК как регуляторные элементы мРНК у бактерий: обзор функций и приложений» . Письма по клеточной и молекулярной биологии . 21 :6. дои : 10.1186/s11658-016-0007-z . ПМЦ 5415839 . ПМИД 28536609 .
- ^ Перейти обратно: а б с д и ж Магистри М., Фагихи М.А., Сен-Лоран Г., Валестедт К. (август 2012 г.). «Регуляция структуры хроматина длинными некодирующими РНК: фокус на природных антисмысловых транскриптах» . Тенденции в генетике . 28 (8): 389–396. дои : 10.1016/j.tig.2012.03.013 . ПМЦ 3768148 . ПМИД 22541732 .
- ^ Перейти обратно: а б с д и ж г Валестедт С (июнь 2013 г.). «Нацеливание на длинную некодирующую РНК для терапевтического усиления экспрессии генов». Обзоры природы. Открытие наркотиков . 12 (6): 433–446. дои : 10.1038/nrd4018 . ПМИД 23722346 . S2CID 288163 .
- ^ Перейти обратно: а б с д Коле Р., Крайнер А.Р., Альтман С. (январь 2012 г.). «РНК-терапия: помимо РНК-интерференции и антисмысловых олигонуклеотидов» . Обзоры природы. Открытие наркотиков . 11 (2): 125–140. дои : 10.1038/nrd3625 . ПМЦ 4743652 . ПМИД 22262036 .
- ^ Вайс Б., Давидкова Г., Чжоу Л.В. (март 1999 г.). «Генная терапия антисмысловыми РНК для изучения и модуляции биологических процессов» . Клеточные и молекулярные науки о жизни . 55 (3): 334–358. дои : 10.1007/s000180050296 . ПМЦ 11146801 . ПМИД 10228554 . S2CID 9448271 .
- ^ Томасон МК, Шторц Г (2010). «Бактериальные антисмысловые РНК: сколько их и что они делают?» . Ежегодный обзор генетики . 44 (1): 167–188. doi : 10.1146/annurev-genet-102209-163523 . ПМК 3030471 . ПМИД 20707673 .
- ^ Вонг Э., Голдберг Т. (февраль 2014 г.). «Мипомерсен (кинамро): новый антисмысловой олигонуклеотидный ингибитор для лечения гомозиготной семейной гиперхолестеринемии» . П&Т . 39 (2): 119–122. ПМЦ 3956393 . ПМИД 24669178 .
- ^ Саймонс Р.В. (декабрь 1988 г.). «Естественный контроль антисмысловой РНК - краткий обзор». Джин . 72 (1–2): 35–44. дои : 10.1016/0378-1119(88)90125-4 . ПМИД 2468573 .
- ^ Итсварт Р., Ву З, Дин С. (сентябрь 2012 г.). «Контроль времени цветения: еще одно окно в связь между антисмысловой РНК и хроматином». Тенденции в генетике . 28 (9): 445–453. дои : 10.1016/j.tig.2012.06.002 . ПМИД 22785023 .
- ^ Федак, Галина; Палусинска, Малгожата; Кшичмоник, Катажина; Бжезняк, Лиен; Ятусевич, Руслан; Петрас, Збигнев; Качановский, Шимон; Свежевский, Шимон (2016). «Контроль покоя семян арабидопсиса с помощью цис-действующего некодирующего антисмыслового транскрипта» . Proc Natl Acad Sci США . 113 (48):E7846–E7855. дои : 10.1073/pnas.1608827113 . ПМК 5137729 . ПМИД 27856735 .
- ^ Каур, Девиндер; Аграхари, Мридула; Сингх, Шаши Шекхар; Мандал, Прабхат Кумар; Бхаттачарья, Алок; Бхаттачарья, Судха (2021). «Транскриптомный анализ Entamoeba histolytica обнаруживает специфичную для домена экспрессию смысловой цепи ORF, кодируемых LINE, с массивной антисмысловой экспрессией домена RT». Плазмида . 114 : 102560. doi : 10.1016/j.plasmid.2021.102560 . ПМИД 33482228 .
- ^ «альфа-талассемия» . Домашний справочник по генетике . Национальная медицинская библиотека НИЗ США. 14 ноября 2017 г.
- ^ Точечный младший (2010). «Метилирование гистонов». Справочник по клеточной сигнализации (второе изд.). стр. 2389–2397. дои : 10.1016/b978-0-12-374145-5.00287-4 . ISBN 978-0-12-374148-6 .
- ^ Вайс, Б. (редактор): Антисмысловые олигодезоксинуклеотиды и антисмысловая РНК: новые фармакологические и терапевтические агенты, CRC Press, Бока-Ратон, Флорида, 1997.
Виды нуклеиновых кислот |
|---|