Рибонуклеаза H
| рибонуклеаза H | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Номер ЕС. | 3.1.26.4 | ||
| Номер CAS. | 9050-76-4 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
| ретровирусная рибонуклеаза H | |||
|---|---|---|---|
| Идентификаторы | |||
| Номер ЕС. | 3.1.26.13 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| |||
Рибонуклеаза H (сокращенно РНКаза H или RNH ) представляет собой семейство неспецифичных последовательности эндонуклеазных ферментов , которые катализируют расщепление РНК в РНК/ ДНК субстрате посредством гидролитического механизма . Члены семейства РНКазы H встречаются практически во всех организмах: от бактерий до архей и эукариот .
Семейство разделено на эволюционно родственные группы с несколько разными предпочтениями в отношении субстратов , широко называемые рибонуклеазами H1 и H2. [ 2 ] Геном человека кодирует как H1, так и H2. Человеческая рибонуклеаза H2 представляет собой гетеротримерный комплекс, состоящий из трех субъединиц, мутации в любой из которых являются одной из генетических причин редкого заболевания , известного как синдром Айкарди-Гутьера . [ 3 ] Третий тип, тесно связанный с Н2, встречается лишь у немногих прокариот . [ 4 ] тогда как H1 и H2 встречаются во всех сферах жизни . [ 4 ] , подобные РНКазе H1, Кроме того, домены H ретровирусной рибонуклеазы H встречаются в многодоменных белках обратной транскриптазы , которые кодируются ретровирусами , такими как ВИЧ, и необходимы для репликации вируса. [ 5 ] [ 6 ]
У эукариот рибонуклеаза H1 участвует в репликации ДНК митохондриального генома . И H1, и H2 участвуют в задачах поддержания генома, таких как обработка структур R-петли . [ 2 ] [ 7 ]
Классификация и номенклатура
[ редактировать ]Рибонуклеаза H представляет собой семейство эндонуклеазных ферментов с общей субстратной специфичностью для цепи РНК РНК - ДНК дуплексов . По определению, РНКазы H расщепляют фосфодиэфирные связи основной цепи РНК, оставляя 3'- гидроксильную и 5'- фосфатную группы. [ 7 ] РНКазы H были предложены как члены эволюционно родственного суперсемейства, включающего другие нуклеазы и ферменты, обрабатывающие нуклеиновые кислоты, такие как ретровирусные интегразы , ДНК- транспозазы , резольвазы соединения Холлидея , белки Piwi и Argonaute , различные экзонуклеазы и сплайсосомный белок Prp8 . [ 8 ] [ 9 ]
РНКазы H можно в общих чертах разделить на два подтипа, H1 и H2, которым по историческим причинам присвоены обозначения арабскими цифрами у эукариот и обозначениями римскими цифрами у прокариотов . Таким образом, РНКаза HI Escherichia coli является гомологом РНКазы H1 Homo sapiens . [ 2 ] [ 7 ] У E. coli и многих других прокариот ген rnhA кодирует HI, а ген rnhB кодирует HII. Третий родственный класс, названный HIII, встречается у некоторых бактерий и архей ; он тесно связан с прокариотическими ферментами HII. [ 4 ]
Структура
[ редактировать ]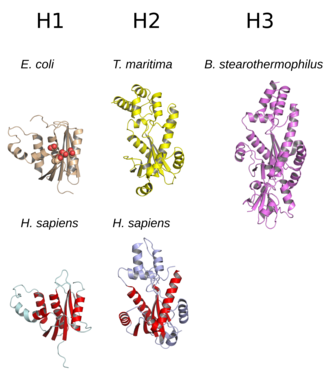
Структура окруженного РНКазы H обычно состоит из 5-нитевого β-листа, набором α-спиралей . [ 10 ] Все РНКазы H имеют активный сайт , сосредоточенный на мотиве консервативной последовательности, состоящем из остатков аспартата и глутамата , часто называемом мотивом DEDD. Эти остатки взаимодействуют с каталитически необходимыми магния . ионами [ 7 ] [ 5 ]
РНКазы H2 крупнее H1 и обычно имеют дополнительные спирали. Доменная ; организация ферментов варьируется некоторые прокариотические и большинство эукариотических членов группы H1 имеют дополнительный небольшой домен на N-конце, известный как «гибридный связывающий домен», который облегчает связывание с гибридными дуплексами РНК: ДНК и иногда обеспечивает повышенную процессивность . [ 2 ] [ 7 ] [ 11 ] Хотя все члены группы H1 и прокариотические члены группы H2 функционируют как мономеры, эукариотические ферменты H2 являются облигатными гетеротримерами . [ 2 ] [ 7 ] Прокариотические ферменты HIII являются членами более широкой группы H2 и имеют большинство общих структурных особенностей с H2, с добавлением N-концевого TATA-бокс-связывающего домена . [ 7 ] Домены ретровирусной РНКазы H, встречающиеся в многодоменных белках обратной транскриптазы, имеют структуру, очень напоминающую группу H1. [ 5 ]
РНКазы H1 были тщательно изучены с целью изучения взаимосвязи между структурой и ферментативной активностью. Они также используются, особенно гомолог E. coli , в качестве модельных систем для изучения сворачивания белков . [ 12 ] [ 13 ] [ 14 ] Внутри группы H1 была выявлена взаимосвязь между более высоким сродством к связыванию субстрата и наличием структурных элементов, состоящих из спирали и гибкой петли, обеспечивающих более крупную и базовую поверхность связывания субстрата. С-спираль имеет разбросанное таксономическое распространение; он присутствует в гомологах E. coli и РНКазы H1 человека и отсутствует в домене H РНКазы ВИЧ, но примеры ретровирусных доменов с C-спиралями действительно существуют. [ 15 ] [ 16 ]
Функция
[ редактировать ]Ферменты рибонуклеазы H расщепляют фосфодиэфирные связи РНК в двухцепочечном гибриде РНК:ДНК, оставляя 3'- гидроксильную и 5'- фосфатную группы на обоих концах сайта разреза с помощью механизма катализа с помощью ионов двух металлов, при котором два двухвалентные катионы, такие как Mg2+ и Mn2+, непосредственно участвуют в каталитической функции. [ 17 ] В зависимости от различий в аминокислотных последовательностях эти РНКазы H подразделяются на РНКазы H 1-го и 2-го типа. [ 7 ] [ 18 ] РНКазы H типа 1 имеют прокариотические и эукариотические РНКазы H1 и ретровирусную РНКазу H. РНКазы H типа 2 имеют прокариотические и эукариотические РНКазы H2 и бактериальную РНКазу H3. Эти РНКазы H существуют в мономерной форме, за исключением эукариотических РНКаз H2, которые существуют в гетеротримерной форме. [ 19 ] [ 20 ] РНКазы H1 и H2 имеют разные предпочтения в отношении субстратов и разные, но перекрывающиеся функции в клетке. У прокариот и низших эукариот ни один из ферментов не является незаменимым , тогда как считается, что оба фермента необходимы для высших эукариот. [ 2 ] Совместная активность ферментов H1 и H2 связана с поддержанием стабильности генома за счет деградации ферментами РНК-компонента R-петлей . [ 21 ] [ 22 ]
Рибонуклеаза H1
[ редактировать ]| Идентификаторы | |||
|---|---|---|---|
| Символ | РНКаза H | ||
| Пфам | PF00075 | ||
| Пфам Клан | CL0219 | ||
| ИнтерПро | ИПР002156 | ||
| PROSITE | ПС50879 | ||
| |||
Ферментам рибонуклеазы H1 требуется как минимум четыре рибонуклеотиды , содержащих пары оснований , в субстрате, и они не могут удалить один рибонуклеотид из цепи, которая в противном случае состоит из дезоксирибонуклеотидов. По этой причине считается маловероятным, что ферменты РНКазы H1 участвуют в процессинге РНК-праймеров из фрагментов Оказаки во время репликации ДНК . [ 2 ] РНКаза H1 не является существенной для одноклеточных организмов, где она была исследована; у E. coli РНКазы H1 нокаут придает температурно-чувствительный фенотип, [ 7 ] а у S. cerevisiae они вызывают нарушения реакции на стресс. [ 23 ]
У многих эукариот, включая млекопитающих , гены РНКазы H1 включают последовательность, нацеленную на митохондрии , что приводит к экспрессии изоформ с присутствующим MTS или без него. В результате РНКаза H1 локализуется как в митохондриях , так и в ядре . В моделях мышей с нокаутом мутанты с нулевым содержанием РНКазы H1 летальны во время эмбриогенеза из-за дефектов репликации митохондриальной ДНК . [ 2 ] [ 24 ] [ 25 ] Дефекты репликации митохондриальной ДНК, вызванные потерей РНКазы H1, вероятно, связаны с дефектами процессинга R-петли . [ 22 ]
Рибонуклеаза H2
[ редактировать ]| Идентификаторы | |||
|---|---|---|---|
| Символ | РНКаза HII | ||
| Пфам | PF01351 | ||
| Пфам Клан | CL0219 | ||
| ИнтерПро | ИПР024567 | ||
| |||
У прокариот РНКаза H2 ферментативно активна в виде мономерного белка. У эукариот это облигатный гетеротример, состоящий из каталитической субъединицы A и структурных субъединиц B и C. Хотя субъединица A близко гомологична прокариотической РНКазе H2, субъединицы B и C не имеют явных гомологов у прокариот и плохо консервативны при уровень последовательности даже среди эукариот. [ 26 ] [ 27 ] Субъединица B опосредует межбелковые взаимодействия между комплексом H2 и PCNA , что локализует H2 в фокусах репликации . [ 28 ]
Как прокариотические, так и эукариотические ферменты H2 могут расщеплять отдельные рибонуклеотиды в цепи. [ 2 ] однако они имеют несколько разные модели расщепления и предпочтения в отношении субстратов: прокариотические ферменты имеют более низкую процессивность и гидролизуют последовательные рибонуклеотиды более эффективно, чем рибонуклеотиды с 5'- дезоксирибонуклеотидом, тогда как эукариотические ферменты более процессивны и гидролизуют оба типа субстрата с одинаковой эффективностью. [ 2 ] [ 27 ] Субстратная специфичность РНКазы H2 позволяет ей участвовать в эксцизионной репарации рибонуклеотидов , удаляя неправильно включенные рибонуклеотиды из ДНК, в дополнение к процессингу R-петли . [ 29 ] [ 30 ] [ 28 ] Хотя и H1, и H2 присутствуют в ядре клеток млекопитающих , H2 является там доминирующим источником активности РНКазы H и важен для поддержания стабильности генома. [ 28 ]
Некоторые прокариоты обладают дополнительным геном типа H2, обозначенным РНКазой HIII в номенклатуре римских цифр, используемой для прокариотических генов. Белки HIII более тесно связаны с группой H2 по идентичности последовательностей и структурному сходству, но имеют предпочтения в отношении субстратов, которые больше напоминают H1. [ 7 ] [ 31 ] В отличие от HI и HII, которые широко распространены среди прокариот, HIII обнаружен лишь у немногих организмов с разбросанным таксономическим распространением; он несколько чаще встречается у архей и редко или никогда не встречается в том же геноме прокариот, что и HI. [ 32 ]
Механизм
[ редактировать ]
Активный центр почти всех РНКаз H содержит четыре отрицательно заряженных аминокислотных остатка, известных как мотив DEDD; часто гистидин , например, в ВИЧ-1, человеке или E. coli. также присутствует [ 2 ] [ 7 ]
Заряженные остатки связывают два иона металлов, необходимых для катализа; в физиологических условиях это ионы магния , но марганец обычно поддерживает ферментативную активность, [ 2 ] [ 7 ] в то время как кальций или высокая концентрация Mg2+ ингибируют активность. [ 11 ] [ 33 ] [ 34 ]
Основываясь на экспериментальных данных и компьютерном моделировании, фермент активирует молекулу воды, связанную с одним из ионов металлов с консервативным гистидином. [ 33 ] [ 35 ] Переходное состояние носит ассоциативный характер. [ 17 ] и образует промежуточное соединение с протонированной фосфатной и депротонированной уходящей группой алкоксида. [ 35 ] Уходящая группа протонируется через глутамат, который имеет повышенное значение рКа и, вероятно, будет протонирован. Механизм аналогичен механизму РНКазы Т и субъединицы RuvC фермента Cas9 , которые также используют гистидиновый и двухметаллический ионный механизм.
Механизм высвобождения продукта расщепления до сих пор не выяснен. Экспериментальные данные кристаллографии с временным разрешением и подобных нуклеаз указывают на роль третьего иона в реакции, рекрутируемой в активный центр. [ 36 ] [ 37 ]
В биологии человека
[ редактировать ]Геном человека содержит четыре гена, кодирующие РНКазу H:
- RNASEH1 , пример подтипа H1 (мономерного).
- RNASEH2A , каталитическая субъединица тримерного комплекса H2.
- RNASEH2B , структурная субъединица тримерного комплекса H2.
- RNASEH2C , структурная субъединица тримерного комплекса H2.
Кроме того, в геноме часто появляется генетический материал ретровирусного происхождения, что отражает интеграцию геномов эндогенных ретровирусов человека . Такие события интеграции приводят к присутствию генов, кодирующих обратную транскриптазу ретровируса , которая включает домен РНКазы H. Примером является ERVK6 . [ 38 ] с длинными терминальными повторами (LTR) и недлинными концевыми повторами (non-LTR) Ретротранспозоны также распространены в геноме и часто включают свои собственные домены РНКазы H со сложной эволюционной историей. [ 39 ] [ 40 ] [ 41 ]
Роль в болезни
[ редактировать ]
В небольших исследованиях мутации в человеческой РНКазе H1 были связаны с хронической прогрессирующей внешней офтальмоплегией , распространенным признаком митохондриальных заболеваний . [ 25 ]
Мутации в любой из трех субъединиц РНКазы H2 хорошо известны как причины редкого генетического заболевания, известного как синдром Айкарди-Гутьера (AGS). [ 3 ] который проявляется неврологическими и дерматологическими симптомами в раннем возрасте. [ 43 ] Симптомы АГС очень напоминают симптомы врожденной вирусной инфекции и связаны с неадекватной активацией интерферона I типа . АГС также может быть вызван мутациями в других генах: TREX1 , SAMHD1 , ADAR и MDA5 /IFIH1, каждый из которых участвует в процессинге нуклеиновых кислот. [ 44 ] Характеристика распределения мутаций в популяции пациентов с AGS выявила 5% всех мутаций AGS в RNASEH2A, 36% в 2B и 12% в 2C. [ 45 ] Мутации в 2B связаны с несколько более легкими неврологическими нарушениями. [ 46 ] и с отсутствием интерферон-индуцированной активации генов, которая может быть обнаружена у пациентов с другими генотипами, связанными с АГС. [ 44 ]
В вирусах
[ редактировать ]
Две группы вирусов используют обратную транскрипцию как часть своего жизненного цикла: ретровирусы , которые кодируют свои геномы в одноцепочечной РНК и реплицируются через промежуточную двухцепочечную ДНК; и вирусы дцДНК-RT , которые реплицируют свои геномы двухцепочечной ДНК через промежуточный «прегеном» РНК. Патогенные примеры включают вирус иммунодефицита человека и вирус гепатита В соответственно. Оба кодируют большие многофункциональные белки обратной транскриптазы (RT), содержащие домены РНКазы H. [ 48 ] [ 49 ]
Ретровирусные белки RT из ВИЧ-1 и вируса мышиного лейкоза являются наиболее изученными членами этого семейства. [ 50 ] [ 51 ] Ретровирусная RT отвечает за преобразование одноцепочечной РНК генома вируса в двухцепочечную ДНК. Этот процесс требует трех этапов: во-первых, РНК-зависимая активность ДНК-полимеразы производит минус-цепь ДНК из матрицы плюс-цепи РНК, образуя гибридный промежуточный продукт РНК:ДНК; во-вторых, цепь РНК разрушается; и в-третьих, ДНК-зависимая активность ДНК-полимеразы синтезирует плюс-цепочечную ДНК, образуя двухцепочечную ДНК в качестве конечного продукта. Второй этап этого процесса осуществляется доменом РНКазы H, расположенным на С-конце белка RT. [ 5 ] [ 6 ] [ 52 ] [ 53 ]
РНКаза H выполняет три типа расщепляющих действий: неспецифическую деградацию генома плюс-цепи РНК, специфическое удаление праймера минус-цепи тРНК и удаление праймера плюс-цепи полипуринового тракта (PPT). [ 54 ] РНКаза H играет роль в праймировании плюс-цепи, но не в обычном методе синтеза новой последовательности праймера. Скорее, РНКаза H создает «праймер» из PPT, который устойчив к расщеплению РНКазой H. При удалении всех оснований, кроме PPT, PPT используется в качестве маркера конца области U3 ее длинного концевого повтора . [ 53 ]
Поскольку активность РНКазы H необходима для пролиферации вируса, этот домен считается мишенью для разработки антиретровирусных препаратов, используемых при лечении ВИЧ / СПИДа и других состояний, вызванных ретровирусами. ингибиторы ретровирусной РНКазы H нескольких различных хемотипов Идентифицированы , механизм действия многих из которых основан на хелатировании катионов активного центра. [ 55 ] Ингибиторы обратной транскриптазы , которые специфически ингибируют полимеразную функцию RT, широко используются в клинической практике, но не ингибиторы функции РНКазы H; это единственная ферментативная функция, кодируемая ВИЧ, на которую еще не воздействуют лекарства, используемые в клинической практике. [ 52 ] [ 56 ]
Эволюция
[ редактировать ]РНКазы H широко распространены и встречаются во всех сферах жизни . Это семейство принадлежит к более крупному суперсемейству нуклеазных ферментов. [ 8 ] [ 9 ] и считается эволюционно древним. [ 57 ] В геномах прокариот часто присутствуют множественные гены РНКазы H, но существует небольшая корреляция между появлением генов HI, HII и HIII и общими филогенетическими отношениями , что позволяет предположить, что горизонтальный перенос генов мог играть роль в установлении распределения этих ферментов. РНКаза HI и HIII редко или никогда не встречаются в одном и том же геноме прокариот. Когда геном организма содержит более одного гена РНКазы H, они иногда имеют значительные различия в уровне активности. Было высказано предположение, что эти наблюдения отражают эволюционную закономерность, которая сводит к минимуму функциональную избыточность генов РНКазы H. [ 7 ] [ 32 ] РНКаза HIII, уникальная для прокариот, имеет разбросанное таксономическое распространение и обнаруживается как у бактерий , так и у архей ; [ 32 ] Считается, что он довольно рано отошел от HII. [ 58 ]
Эволюционная траектория РНКазы H2 у эукариот, особенно механизм, с помощью которого эукариотические гомологи стали облигатными гетеротримерами, неясен; субъединицы B и C не имеют очевидных гомологов у прокариот. [ 2 ] [ 27 ]
Приложения
[ редактировать ]Поскольку РНКаза H специфически разрушает только РНК в гибридах двухцепочечной РНК:ДНК, ее обычно используют в качестве лабораторного реагента в молекулярной биологии . Очищенные препараты РНКазы HI и HII E.coli коммерчески доступны. РНКаза HI часто используется для разрушения матрицы РНК после синтеза первой цепи комплементарной ДНК (кДНК) путем обратной транскрипции . Его также можно использовать для расщепления определенных последовательностей РНК в присутствии коротких комплементарных сегментов ДНК. [ 59 ] высокочувствительные методы, такие как поверхностный плазмонный резонанс . Для обнаружения можно использовать [ 60 ] [ 61 ] РНКаза HII может использоваться для разрушения компонента РНК-праймера фрагмента Оказаки или для введения одноцепочечных разрывов в положениях, содержащих рибонуклеотид. [ 59 ] Вариант ПЦР с горячим стартом , известный как РНКаза H-зависимая ПЦР или рчПЦР, был описан с использованием термостабильной РНКазы HII из гипертермофильной археи Pyrococcus abyssi . [ 62 ] Следует отметить, что белок- ингибитор рибонуклеазы, обычно используемый в качестве реагента, не эффективен для ингибирования активности ни HI, ни HII. [ 59 ]
История
[ редактировать ]Рибонуклеазы H были впервые обнаружены в лаборатории Питера Хаузена РНК:ДНК , когда в 1969 году исследователи обнаружили гибридную эндонуклеазную активность теленка в тимусе и дали ей название «рибонуклеаза H », чтобы обозначить ее гибридную специфичность. [ 26 ] [ 63 ] [ 64 ] Впоследствии активность РНКазы H была обнаружена в E. coli. [ 65 ] и в образце онковирусов с РНК-геномами вирусов во время ранних исследований обратной транскрипции . [ 66 ] [ 67 ] Позже выяснилось, что экстракт тимуса теленка содержит более одного белка с активностью РНКазы H. [ 68 ] и что E. coli содержала два гена РНКазы H. [ 69 ] [ 70 ] Первоначально фермент, ныне известный как РНКаза H2 у эукариот, имел обозначение H1 и наоборот, но названия эукариотических ферментов были заменены так, чтобы они соответствовали названиям в E. coli , чтобы облегчить сравнительный анализ, в результате чего появилась современная номенклатура, в которой прокариотические ферменты обозначаются как римскими цифрами, а эукариотические ферменты арабскими цифрами. [ 2 ] [ 26 ] [ 31 ] [ 71 ] Прокариотическая РНКаза HIII, о которой сообщалось в 1999 году, была последним идентифицированным подтипом РНКазы H. [ 31 ]
Характеристика эукариотической РНКазы H2 исторически была сложной задачей, отчасти из-за ее низкой распространенности. [ 2 ] Тщательные усилия по очистке фермента позволили предположить, что, в отличие от РНКазы H2 E. coli , эукариотический фермент состоит из нескольких субъединиц. [ 72 ] Гомолог S. cerevisiae белка E. coli (то есть субъединица H2A) был легко идентифицирован с помощью биоинформатики дрожжей . генома при секвенировании [ 73 ] но было обнаружено, что соответствующий белок не обладает ферментативной активностью изолированно. [ 2 ] [ 23 ] В конце концов, субъединицы B и C дрожжей были выделены путем совместной очистки и обнаружено, что они необходимы для ферментативной активности. [ 74 ] Однако субъединицы B и C дрожжей имеют очень низкую идентичность последовательностей со своими гомологами в других организмах, и соответствующие человеческие белки были окончательно идентифицированы только после того, как было обнаружено, что мутации во всех трех вызывают синдром Айкарди-Гутьера . [ 2 ] [ 3 ]
Ссылки
[ редактировать ]- ^ ВВП : 1JL1 ; Гёдкен Э.Р., Маркизи С. (декабрь 2001 г.). «Энергетика нативного состояния термостабилизированного варианта рибонуклеазы HI». Журнал молекулярной биологии . 314 (4): 863–71. дои : 10.1006/jmbi.2001.5184 . ПМИД 11734003 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д Серрителли С.М., Крауч Р.Дж. (март 2009 г.). «Рибонуклеаза H: ферменты эукариот» . Журнал ФЭБС . 276 (6): 1494–505. дои : 10.1111/j.1742-4658.2009.06908.x . ПМК 2746905 . ПМИД 19228196 .
- ^ Jump up to: а б с Кроу Ю.Дж., Лейтч А., Хейворд Б.Е., Гарнер А., Пармар Р., Гриффит Е. и др. (август 2006 г.). «Мутации в генах, кодирующих субъединицы рибонуклеазы H2, вызывают синдром Айкарди-Гутьера и имитируют врожденную вирусную инфекцию головного мозга». Природная генетика . 38 (8): 910–6. дои : 10.1038/ng1842 . ПМИД 16845400 . S2CID 8076225 .
- ^ Jump up to: а б с Фигель М., Новотны М. (август 2014 г.). «Кристаллическая структура комплекса РНКаза H3-субстрат демонстрирует параллельную эволюцию гибридного распознавания РНК/ДНК» . Исследования нуклеиновых кислот . 42 (14): 9285–94. дои : 10.1093/nar/gku615 . ПМЦ 4132731 . ПМИД 25016521 .
- ^ Jump up to: а б с д Дэвис Дж. Ф., Гостомска З., Гостомский З., Джордан С.Р., Мэтьюз Д.А. (апрель 1991 г.). «Кристаллическая структура домена рибонуклеазы H обратной транскриптазы ВИЧ-1». Наука . 252 (5002): 88–95. Бибкод : 1991Sci...252...88D . дои : 10.1126/science.1707186 . ПМИД 1707186 .
- ^ Jump up to: а б Хансен Дж., Шульце Т., Меллерт В., Мёллинг К. (январь 1988 г.). «Идентификация и характеристика ВИЧ-специфической РНКазы H с помощью моноклональных антител» . Журнал ЭМБО . 7 (1): 239–43. дои : 10.1002/j.1460-2075.1988.tb02805.x . ПМК 454263 . ПМИД 2452083 .
- ^ Jump up to: а б с д и ж г час я дж к л м Тадокоро Т., Канайя С. (март 2009 г.). «Рибонуклеаза H: молекулярное разнообразие, домены связывания субстрата и каталитический механизм прокариотических ферментов». Журнал ФЭБС . 276 (6): 1482–93. дои : 10.1111/j.1742-4658.2009.06907.x . ПМИД 19228197 . S2CID 29008571 .
- ^ Jump up to: а б Майорек К.А., Дунин-Горкавич С., Стечкевич К., Мушевска А., Новотны М., Гинальски К., Буйницкий Ю.М. (апрель 2014 г.). «Суперсемейство РНКазы H: новые члены, сравнительный структурный анализ и эволюционная классификация» . Исследования нуклеиновых кислот . 42 (7): 4160–79. дои : 10.1093/nar/gkt1414 . ПМЦ 3985635 . ПМИД 24464998 .
- ^ Jump up to: а б Райс П., Крейги Р., Дэвис Д.Р. (февраль 1996 г.). «Ретровирусные интегразы и их родственники» . Современное мнение в области структурной биологии . 6 (1): 76–83. дои : 10.1016/s0959-440x(96)80098-4 . ПМИД 8696976 .
- ^ Шмитт Т.Дж., Кларк Дж.Э., Ноттс Т.А. (декабрь 2009 г.). «Термическое и механическое многосостоятельное сворачивание рибонуклеазы H». Журнал химической физики . 131 (23): 235101. Бибкод : 2009JChPh.131w5101S . дои : 10.1063/1.3270167 . ПМИД 20025349 .
- ^ Jump up to: а б Новотны М., Серрителли С.М., Гирландо Р., Гайдамаков С.А., Крауч Р.Дж., Ян В. (апрель 2008 г.). «Специфическое распознавание гибрида РНК/ДНК и усиление активности РНКазы H1 человека с помощью HBD» . Журнал ЭМБО . 27 (7): 1172–81. дои : 10.1038/emboj.2008.44 . ПМЦ 2323259 . ПМИД 18337749 .
- ^ Чеккони С., Шанк Э.А., Бустаманте С., Маркизи С. (сентябрь 2005 г.). «Прямое наблюдение сворачивания трех состояний одной белковой молекулы». Наука . 309 (5743): 2057–60. Бибкод : 2005Sci...309.2057C . дои : 10.1126/science.1116702 . ПМИД 16179479 . S2CID 43823877 .
- ^ Холлиен Дж., Маркизи С. (март 1999 г.). «Термодинамическое сравнение мезофильных и термофильных рибонуклеаз H». Биохимия . 38 (12): 3831–6. дои : 10.1021/bi982684h . ПМИД 10090773 .
- ^ Рашке Т.М., Маркизи С. (апрель 1997 г.). «Кинетический промежуточный продукт сворачивания рибонуклеазы H напоминает расплавленную кислоту глобулу и частично развернутые молекулы, обнаруженные в нативных условиях». Структурная биология природы . 4 (4): 298–304. дои : 10.1038/nsb0497-298 . ПМИД 9095198 . S2CID 33673059 .
- ^ Шульц С.Дж., Шампу Ж.Дж. (июнь 2008 г.). «Активность РНКазы H: структура, специфичность и функция в обратной транскрипции» . Вирусные исследования . 134 (1–2): 86–103. doi : 10.1016/j.virusres.2007.12.007 . ПМЦ 2464458 . ПМИД 18261820 .
- ^ Шампу Дж.Дж., Шульц С.Дж. (март 2009 г.). «Рибонуклеаза H: свойства, субстратная специфичность и роль в обратной транскрипции ретровирусов» . Журнал ФЭБС . 276 (6): 1506–16. дои : 10.1111/j.1742-4658.2009.06909.x . ПМЦ 2742777 . ПМИД 19228195 .
- ^ Jump up to: а б Ян В., Ли Дж.Й., Новотны М. (апрель 2006 г.). «Создание и разрушение нуклеиновых кислот: двухионный катализ Mg2+ и субстратная специфичность» . Молекулярная клетка . 22 (1): 5–13. doi : 10.1016/j.molcel.2006.03.013 . ПМИД 16600865 .
- ^ Отани Н., Харуки М., Морикава М., Канайя С. (январь 1999 г.). «Молекулярное разнообразие РНКаз H». Журнал бионауки и биоинженерии . 88 (1): 12–9. дои : 10.1016/s1389-1723(99)80168-6 . ПМИД 16232566 .
- ^ Бубек Д., Рейнс М.А., Грэм С.С., Астелл К.Р., Джонс Э.Ю., Джексон А.П. (май 2011 г.). «PCNA направляет активность РНКазы H типа 2 на субстраты репликации и репарации ДНК» . Исследования нуклеиновых кислот . 39 (9): 3652–66. дои : 10.1093/nar/gkq980 . ПМК 3089482 . ПМИД 21245041 .
- ^ Фигил М., Чон Х., Серрителли С.М., Цибульска М., Крауч Р.Дж., Новотны М. (март 2011 г.). «Структурная и биохимическая характеристика комплекса РНКазы H2 человека раскрывает молекулярную основу распознавания субстрата и дефектов синдрома Айкарди-Гутьера» . Журнал биологической химии . 286 (12): 10540–50. дои : 10.1074/jbc.M110.181974 . ПМК 3060507 . ПМИД 21177858 .
- ^ Амон Дж. Д., Кошланд Д. (декабрь 2016 г.). «РНКаза H обеспечивает эффективное восстановление повреждений ДНК, вызванных R-петлей» . электронная жизнь . 5 : e20533. doi : 10.7554/eLife.20533 . ПМК 5215079 . ПМИД 27938663 .
- ^ Jump up to: а б Лима В.Ф., Мюррей Х.М., Дамл С.С., Харт К.Э., Хунг Г., Де Ойос С.Л. и др. (июнь 2016 г.). «Жизнеспособные мыши с нокаутом РНКазы H1 показывают, что РНКаза H1 необходима для обработки R-петли, функции митохондрий и печени» . Исследования нуклеиновых кислот . 44 (11): 5299–312. дои : 10.1093/nar/gkw350 . ПМЦ 4914116 . ПМИД 27131367 .
- ^ Jump up to: а б Арудчандран А., Серрителли С., Наримацу С., Итая М., Шин Д.Ю., Симада Ю., Крауч Р.Дж. (октябрь 2000 г.). «Отсутствие рибонуклеазы H1 или H2 изменяет чувствительность Saccharomyces cerevisiae к гидроксимочевине, кофеину и этилметансульфонату: значение роли РНКазы H в репликации и репарации ДНК» . Гены в клетки . 5 (10): 789–802. дои : 10.1046/j.1365-2443.2000.00373.x . ПМИД 11029655 .
- ^ Серрителли С.М., Фролова Е.Г., Фенг С., Гринберг А., Лав П.Е., Крауч Р.Дж. (март 2003 г.). «Неспособность производить митохондриальную ДНК приводит к эмбриональной смертности у мышей с нулевым Rnaseh1» . Молекулярная клетка . 11 (3): 807–15. дои : 10.1016/s1097-2765(03)00088-1 . ПМИД 12667461 .
- ^ Jump up to: а б Рейес А., Мельчионда Л., Наска А., Каррара Ф., Ламантеа Е., Занолини А. и др. (июль 2015 г.). «Мутации RNASEH1 нарушают репликацию мтДНК и вызывают митохондриальную энцефаломиопатию у взрослых» . Американский журнал генетики человека . 97 (1): 186–93. дои : 10.1016/j.ajhg.2015.05.013 . ПМЦ 4572567 . ПМИД 26094573 .
- ^ Jump up to: а б с Холлис Т., Шабан Н.М. (1 января 2011 г.). «Структура и функции ферментов РНКазы H». В Николсоне А.В. (ред.). Рибонуклеазы . Нуклеиновые кислоты и молекулярная биология. Шпрингер Берлин Гейдельберг. стр. 299–317 . дои : 10.1007/978-3-642-21078-5_12 . ISBN 978-3-642-21077-8 .
- ^ Jump up to: а б с Чон Х., Василев А., ДеПамфилис М.Л., Чжао Ю., Чжан Дж., Бургерс П.М. и др. (январь 2009 г.). «Вклад двух дополнительных субъединиц, RNASEH2B и RNASEH2C, в активность и свойства комплекса РНКазы H2 человека» . Исследования нуклеиновых кислот . 37 (1): 96–110. дои : 10.1093/нар/gkn913 . ПМК 2615623 . ПМИД 19015152 .
- ^ Jump up to: а б с Рейнс М.А., Джексон А.П. (август 2014 г.). «Рибонуклеаза H2 в здоровье и болезни». Труды Биохимического общества . 42 (4): 717–25. дои : 10.1042/BST20140079 . ПМИД 25109948 .
- ^ Вахба Л., Амон Дж. Д., Кошланд Д., Вика-Росс М. (декабрь 2011 г.). «РНКаза H и многочисленные факторы биогенеза РНК взаимодействуют, предотвращая возникновение нестабильности генома гибридами РНК:ДНК» . Молекулярная клетка . 44 (6): 978–88. дои : 10.1016/j.molcel.2011.10.017 . ПМЦ 3271842 . ПМИД 22195970 .
- ^ Ким Н., Хуанг С.Н., Уильямс Дж.С., Ли Ю.К., Кларк А.Б., Чо Дж.Е. и др. (июнь 2011 г.). «Мутагенный процессинг рибонуклеотидов в ДНК дрожжевой топоизомеразой I» . Наука . 332 (6037): 1561–4. Бибкод : 2011Sci...332.1561K . дои : 10.1126/science.1205016 . ПМК 3380281 . ПМИД 21700875 .
- ^ Jump up to: а б с Отани Н., Харуки М., Морикава М., Крауч Р.Дж., Итая М., Канайя С. (январь 1999 г.). «Идентификация генов, кодирующих Mn2+-зависимую РНКазу HII и Mg2+-зависимую РНКазу HIII из Bacillus subtilis: классификация РНКаз H на три семейства». Биохимия . 38 (2): 605–18. дои : 10.1021/bi982207z . ПМИД 9888800 .
- ^ Jump up to: а б с Кочива Х., Томита М., Канаи А. (июль 2007 г.). «Эволюция генов рибонуклеазы H у прокариот во избежание наследования избыточных генов» . Эволюционная биология BMC . 7 (1): 128. Бибкод : 2007BMCEE...7..128K . дои : 10.1186/1471-2148-7-128 . ПМК 1950709 . ПМИД 17663799 .
- ^ Jump up to: а б Алла Н.Р., Николсон А.В. (декабрь 2012 г.). «Доказательства двойной функциональной роли консервативного гистидина в гетеродуплексном расщеплении РНК·ДНК человеческой РНКазой H1» . Журнал ФЭБС . 279 (24): 4492–500. дои : 10.1111/февраль 12035 . ПМЦ 3515698 . ПМИД 23078533 .
- ^ Роста Э., Ян В., Хаммер Дж. (февраль 2014 г.). «Ингибирование кальцием катализа двухметаллическими ионами рибонуклеазы H1» . Журнал Американского химического общества . 136 (8): 3137–44. дои : 10.1021/ja411408x . ПМЦ 3985467 . ПМИД 24499076 .
- ^ Jump up to: а б Дюрр С., Богусевич О, Берта Д., Суардиас Р., Питер С., Джамбрина П.Г., Питер С., Шао Ю., Роста Е (16 июня 2021 г.). «Роль консервативных остатков в мотиве DEDDh: механизм переноса протона РНКазы H ВИЧ-1» . АКС-катализ . 11 (13): 7915–7927. дои : 10.1021/acscatal.1c01493 . S2CID 236285134 .
- ^ Ган Дж., Шоу Дж., Тропеа Дж.Э., Во Д.С., Корт Д.Л., Джи Икс (январь 2008 г.). «Пошаговая модель процессинга двухцепочечной РНК рибонуклеазой III» . Мол Микробиол . 67 (1): 143–54. дои : 10.1111/j.1365-2958.2007.06032.x . ПМИД 18047582 .
- ^ Самара Н.Л., Ян В. (август 2019 г.). «Торговля катионами ускоряет гидролиз РНК» . Структурная и молекулярная биология природы . 25 (8): 715–721. дои : 10.1038/s41594-018-0099-4 . ПМК 6110950 . ПМИД 30076410 .
- ^ Реус К., Майер Дж., Заутер М., Шерер Д., Мюллер-Ланцш Н., Миз Э. (март 2001 г.). «Геномная организация эндогенного ретровируса человека HERV-K (HML-2.HOM) (ERVK6) на хромосоме 7». Геномика . 72 (3): 314–20. дои : 10.1006/geno.2000.6488 . ПМИД 11401447 .
- ^ Ustyantsev K, Blinov A, Smyshlyaev G (14 March 2017). "Convergence of retrotransposons in oomycetes and plants" . Mobile DNA . 8 (1): 4. doi : 10.1186/s13100-017-0087-y . PMC 5348765 . PMID 28293305 .
- ^ Устьянцев К, Новикова О, Блинов А, Смышляев Г (май 2015). «Конвергентная эволюция рибонуклеазы h в ретротранспозонах LTR и ретровирусах» . Молекулярная биология и эволюция . 32 (5): 1197–207. дои : 10.1093/molbev/msv008 . ПМК 4408406 . ПМИД 25605791 .
- ^ Малик Х.С. (2005). «Эволюция рибонуклеазы H в ретромобильных элементах». Цитогенетические и геномные исследования . 110 (1–4): 392–401. дои : 10.1159/000084971 . ПМИД 16093691 . S2CID 7481781 .
- ^ Фигил М., Чон Х., Серрителли С.М., Цибульска М., Крауч Р.Дж., Новотны М. (март 2011 г.). «Структурная и биохимическая характеристика комплекса РНКазы H2 человека раскрывает молекулярную основу распознавания субстрата и дефектов синдрома Айкарди-Гутьера» . Журнал биологической химии . 286 (12): 10540–50. дои : 10.1074/jbc.M110.181974 . ПМК 3060507 . ПМИД 21177858 .
- ^ Орчези С., Ла Пиана Р., Фацци Э. (2009). «Синдром Айкарди-Гутьера» . Британский медицинский бюллетень . 89 : 183–201. дои : 10.1093/bmb/ldn049 . ПМИД 19129251 .
- ^ Jump up to: а б Кроу Ю.Дж., Манель Н. (июль 2015 г.). «Синдром Айкарди-Гутьера и интерферонопатии I типа». Обзоры природы. Иммунология . 15 (7): 429–40. дои : 10.1038/nri3850 . ПМИД 26052098 . S2CID 34259643 .
- ^ Кроу Ю.Дж., Чейз Д.С., Ловенштейн Шмидт Дж., Шинкевич М., Форте Г.М., Горналл Х.Л. и др. (февраль 2015 г.). «Характеристика фенотипов заболеваний человека, связанных с мутациями TREX1, RNASEH2A, RNASEH2B, RNASEH2C, SAMHD1, ADAR и IFIH1» . Американский журнал медицинской генетики. Часть А. 167А (2): 296–312. дои : 10.1002/ajmg.a.36887 . ПМЦ 4382202 . ПМИД 25604658 .
- ^ Райс Дж., Патрик Т., Пармар Р., Тейлор К.Ф., Эби А., Айкарди Дж. и др. (октябрь 2007 г.). «Клинический и молекулярный фенотип синдрома Айкарди-Гутьера» . Американский журнал генетики человека . 81 (4): 713–25. дои : 10.1086/521373 . ПМК 2227922 . ПМИД 17846997 .
- ^ Сарафианос С.Г., Дас К., Тантилло С., Кларк А.Д., Дин Дж., Уиткомб Дж.М. и др. (март 2001 г.). «Кристаллическая структура обратной транскриптазы ВИЧ-1 в комплексе с полипуриновым трактом РНК:ДНК» . Журнал ЭМБО . 20 (6): 1449–61. дои : 10.1093/emboj/20.6.1449 . ПМК 145536 . ПМИД 11250910 .
- ^ Сигер С., Мейсон В.С. (май 2015 г.). «Молекулярная биология инфекции вирусом гепатита В» . Вирусология . 479–480: 672–86. дои : 10.1016/j.virol.2015.02.031 . ПМК 4424072 . ПМИД 25759099 .
- ^ Моеллинг К., Брокер Ф., Керриган Дж. Э. (01 января 2014 г.). «РНКаза H: специфичность, механизмы действия и противовирусная мишень». В Виченци Э., Поли Дж. (ред.). Ретровирусы человека . Методы молекулярной биологии. Том. 1087. Хумана Пресс. стр. 71–84. дои : 10.1007/978-1-62703-670-2_7 . ISBN 978-1-62703-669-6 . ПМИД 24158815 .
- ^ Мизуно М., Ясукава К., Иноуэ К. (февраль 2010 г.). «Понимание механизма стабилизации обратной транскриптазы вируса мышиного лейкоза Молони путем устранения активности РНКазы H». Бионауки, биотехнологии и биохимия . 74 (2): 440–2. дои : 10.1271/bbb.90777 . ПМИД 20139597 . S2CID 28110533 .
- ^ Коте М.Л., Рот М.Дж. (июнь 2008 г.). «Обратная транскриптаза вируса мышиного лейкоза: структурное сравнение с обратной транскриптазой ВИЧ-1» . Вирусные исследования . 134 (1–2): 186–202. doi : 10.1016/j.virusres.2008.01.001 . ПМЦ 2443788 . ПМИД 18294720 .
- ^ Jump up to: а б Новотны М., Фигель М. (1 января 2013 г.). «Домен РНКазы H: структура, функция и механизм». В LeGrice S, Gotte M (ред.). Обратная транскриптаза вируса иммунодефицита человека . Спрингер Нью-Йорк. стр. 53–75. дои : 10.1007/978-1-4614-7291-9_3 . ISBN 978-1-4614-7290-2 .
- ^ Jump up to: а б Бейльхарц Г.Л., Гётте М. (апрель 2010 г.). «Рибонуклеаза H ВИЧ-1: структура, каталитический механизм и ингибиторы» . Вирусы . 2 (4): 900–26. дои : 10.3390/v2040900 . ПМК 3185654 . ПМИД 21994660 .
- ^ Кларманн Г.Дж., Хокинс М.Е., Ле Грайс С.Ф. (2002). «Раскрытие сложностей ретровирусной рибонуклеазы H раскрывает ее потенциал как терапевтической мишени». Обзоры по СПИДу . 4 (4): 183–94. ПМИД 12555693 .
- ^ Трамонтано Э., Ди Санто Р. (2010). «Ингибиторы функции РНКазы H, ассоциированные с RT ВИЧ-1: последние достижения в разработке лекарств». Современная медицинская химия . 17 (26): 2837–53. дои : 10.2174/092986710792065045 . ПМИД 20858167 .
- ^ Цао Л, Сун В, Де Клерк Э, Чжан П, Лю Икс (июнь 2014 г.). «Последние успехи в исследовании низкомолекулярных ингибиторов РНКазы H ВИЧ-1». Современная медицинская химия . 21 (17): 1956–67. дои : 10.2174/0929867321666140120121158 . ПМИД 24438523 .
- ^ Ма Б.Г., Чен Л., Цзи Х.Ф., Чен Ч.Ж., Ян Ф.Р., Ван Л. и др. (февраль 2008 г.). «Характеристики очень древних белков». Связь с биохимическими и биофизическими исследованиями . 366 (3): 607–11. дои : 10.1016/j.bbrc.2007.12.014 . ПМИД 18073136 .
- ^ Бриндефальк Б., Дессайи Б.Х., Йейтс С., Оренго С., Вернер Ф., Пул А.М. (март 2013 г.). «Эволюционная история суперсемейства TBP-доменов» . Исследования нуклеиновых кислот . 41 (5): 2832–45. дои : 10.1093/нар/gkt045 . ПМЦ 3597702 . ПМИД 23376926 .
- ^ Jump up to: а б с Николс Н.М., Юэ Д. (1 января 2001 г.). Рибонуклеазы . Том. Глава 3. John Wiley & Sons, Inc., стр. Раздел 3.13. дои : 10.1002/0471142727.mb0313s84 . ISBN 978-0-471-14272-0 . ПМИД 18972385 . S2CID 221604377 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Лоо Дж.Ф., Ван С.С., Пэн Ф., Хэ Дж.А., Хэ Л., Го Ю.К. и др. (июль 2015 г.). «Платформа SPR без ПЦР, использующая РНКазу H для обнаружения микроРНК 29a-3p в мазках из горла людей, зараженных вирусом гриппа A H1N1». Аналитик . 140 (13): 4566–75. Бибкод : 2015Ана...140.4566L . дои : 10.1039/C5AN00679A . ПМИД 26000345 . S2CID 28974459 .
- ^ Гудрич Т.Т., Ли Х.Дж., Корн Р.М. (апрель 2004 г.). «Прямое обнаружение геномной ДНК с помощью измерений SPR-визуализации с помощью ферментативной амплификации микрочипов РНК». Журнал Американского химического общества . 126 (13): 4086–7. CiteSeerX 10.1.1.475.1922 . дои : 10.1021/ja039823p . ПМИД 15053580 .
- ^ Добоси-младший, Роуз С.Д., Бельц К.Р., Рупп С.М., Пауэрс К.М., Белке М.А., Уолдер Дж.А. (август 2011 г.). «РНКаза H-зависимая ПЦР (рчПЦР): улучшенная специфичность и обнаружение полиморфизма отдельных нуклеотидов с использованием блокированных расщепляемых праймеров» . БМК Биотехнология . 11:80 . дои : 10.1186/1472-6750-11-80 . ПМЦ 3224242 . ПМИД 21831278 .
- ^ Штейн Х., Хаузен П. (октябрь 1969 г.). «Фермент из тимуса теленка, разрушающий фрагмент РНК гибридов ДНК-РНК: влияние на ДНК-зависимую РНК-полимеразу». Наука . 166 (3903): 393–5. Бибкод : 1969Sci...166..393S . дои : 10.1126/science.166.3903.393 . ПМИД 5812039 . S2CID 43683241 .
- ^ Хаузен П., Штейн Х. (июнь 1970 г.). «Рибонуклеаза H. Фермент, расщепляющий фрагмент РНК гибридов ДНК-РНК» . Европейский журнал биохимии . 14 (2): 278–83. дои : 10.1111/j.1432-1033.1970.tb00287.x . ПМИД 5506170 .
- ^ Миллер Х.И., Риггс А.Д., Гилл Г.Н. (апрель 1973 г.). «Рибонуклеаза H (гибрид) в Escherichia coli. Идентификация и характеристика» . Журнал биологической химии . 248 (7): 2621–4. дои : 10.1016/S0021-9258(19)44152-5 . ПМИД 4572736 .
- ^ Мёллинг К., Болоньези Д.П., Бауэр Х., Бюзен В., Плассманн Х.В., Хаузен П. (декабрь 1971 г.). «Ассоциация вирусной обратной транскриптазы с ферментом, разрушающим фрагмент РНК гибридов РНК-ДНК». Природа . 234 (51): 240–3. дои : 10.1038/newbio234240a0 . ПМИД 4331605 .
- ^ Грандженетт Д.П., Джерард Г.Ф., Грин М. (декабрь 1972 г.). «Рибонуклеаза H: повсеместная активность в вирионах опухолевых вирусов, содержащих рибонуклеиновую кислоту» . Журнал вирусологии . 10 (6): 1136–42. дои : 10.1128/jvi.10.6.1136-1142.1972 . ПМК 356594 . ПМИД 4118867 .
- ^ Бюзен В., Хаузен П. (март 1975 г.). «Особая активность рибонуклеазы H в тимусе теленка» . Европейский журнал биохимии . 52 (1): 179–90. дои : 10.1111/j.1432-1033.1975.tb03985.x . ПМИД 51794 .
- ^ Канайя С., Крауч Р.Дж. (январь 1983 г.). «Последовательность ДНК гена, кодирующего рибонуклеазу H Escherichia coli» . Журнал биологической химии . 258 (2): 1276–81. дои : 10.1016/S0021-9258(18)33189-2 . ПМИД 6296074 .
- ^ Итая М. (ноябрь 1990 г.). «Выделение и характеристика второй РНКазы H (РНКазы HII) Escherichia coli K-12, кодируемой геном rnhB» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (21): 8587–91. Бибкод : 1990PNAS...87.8587I . дои : 10.1073/pnas.87.21.8587 . ПМК 55002 . ПМИД 2172991 .
- ^ Крауч Р.Дж., Арудчандран А., Серрителли С.М. (1 января 2001 г.). «РНКаза H1 Saccharomyces cerevisiae: методы и номенклатура». Рибонуклеазы – Часть А. Методы энзимологии. Том. 341. С. 395–413. дои : 10.1016/s0076-6879(01)41166-9 . ISBN 978-0-12-182242-2 . ПМИД 11582793 .
- ^ Франк П., Браунсхофер-Райтер К., Винтерсбергер У., Гримм Р., Бюзен В. (октябрь 1998 г.). «Клонирование кДНК, кодирующей большую субъединицу человеческой РНКазы HI, гомолога прокариотической РНКазы HII» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (22): 12872–7. Бибкод : 1998PNAS...9512872F . дои : 10.1073/pnas.95.22.12872 . ПМК 23637 . ПМИД 9789007 .
- ^ Франк П., Брауншофер-Рейтер К., Винтерсбергер Ю. (январь 1998 г.). «Дрожжевая РНКаза H(35) является аналогом РНКазы HI млекопитающих и эволюционно связана с прокариотической РНКазой HII». Письма ФЭБС . 421 (1): 23–6. дои : 10.1016/s0014-5793(97)01528-7 . ПМИД 9462832 .
- ^ Чон Х.С., Баклунд П.С., Чен Х.К., Караванов А.А., Крауч Р.Дж. (01 января 2004 г.). «РНКаза H2 Saccharomyces cerevisiae представляет собой комплекс из трех белков» . Исследования нуклеиновых кислот . 32 (2): 407–14. дои : 10.1093/nar/gkh209 . ПМЦ 373335 . ПМИД 14734815 .
Внешние ссылки
[ редактировать ]- Запись GeneReviews/NCBI/NIH/UW о синдроме Айкарди-Гутьера
- РНКаза + H в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)