Fc-рецептор
| Иммуноглобулиноподобный рецептор | |
|---|---|
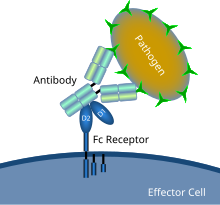 Схематическая диаграмма, показывающая взаимодействие рецептора Fc с микробным патогеном, покрытым антителами. | |
| Идентификаторы | |
| Символ | Fc-рецептор |
| Мембраном | 10 |
В иммунологии рецептор Fc — это белок, обнаруженный на поверхности определенных клеток , включая, среди прочего, B-лимфоциты , фолликулярные дендритные клетки , естественные клетки-киллеры , макрофаги , нейтрофилы , эозинофилы , базофилы человека , тромбоциты и тучные клетки , которые способствуют на защитные функции иммунной системы .Его название происходит от его специфичности связывания с частью антитела, известной как область Fc (кристаллизуемый фрагмент) . Рецепторы Fc связываются с антителами, которые прикрепляются к инфицированным клеткам или вторгающимся патогенам . Их активность стимулирует фагоцитарные или цитотоксические клетки уничтожать микробы или инфицированные клетки посредством антитело-опосредованного фагоцитоза или антитело-зависимой клеточно-опосредованной цитотоксичности . Некоторые вирусы , такие как флавивирусы, используют рецепторы Fc, чтобы помочь им инфицировать клетки с помощью механизма, известного как антителозависимое усиление инфекции. [1]
Классы
[ редактировать ]Существует несколько различных типов рецепторов Fc (сокращенно FcR), которые классифицируются в зависимости от типа антител , которые они распознают. Латинская буква, используемая для обозначения типа антитела, преобразуется в соответствующую греческую букву, которая ставится после части названия «Fc». Например, те, которые связывают наиболее распространенный класс антител, IgG , называются Fc-гамма-рецепторами (FcγR), те, которые связывают IgA, называются Fc-альфа-рецепторами (FcαR), а те, которые связывают IgE, называются Fc-эпсилон-рецепторами (Fc-epsilon рецепторы). FcεR). Классы FcR также различаются по клеткам, которые их экспрессируют (макрофаги, гранулоциты, естественные клетки-киллеры, Т- и В-клетки) и сигнальным свойствам каждого рецептора. [2]
Fc-гамма-рецепторы
[ редактировать ]Все Fcγ-рецепторы (FcγR) относятся к суперсемейству иммуноглобулинов и являются наиболее важными Fc-рецепторами для индукции ( меченых фагоцитоза опсонизированных ) микробов. [3] В это семейство входят несколько членов: FcγRI (CD64), FcγRIIA ( CD32 ), FcγRIIB (CD32), FcγRIIIA (CD16a), FcγRIIIB (CD16b), которые различаются по сродству к антителам из-за разной молекулярной структуры . [4] Например, FcγRI связывается с IgG сильнее, чем FcγRII или FcγRIII. FcγRI также имеет внеклеточную часть, состоящую из трех иммуноглобулиновых (Ig)-подобных доменов , что на один домен больше, чем у FcγRII или FcγRIII. Это свойство позволяет FcγRI связывать одну молекулу IgG (или мономер все рецепторы Fcγ должны связывать несколько молекул IgG в составе иммунного комплекса . ), но для активации [5]
Рецепторы Fc-гамма различаются по своему сродству к IgG, и аналогичным образом разные подклассы IgG обладают уникальным сродством к каждому из гамма-рецепторов Fc. [6] Эти взаимодействия дополнительно регулируются гликаном (олигосахаридом) в положении CH2-84.4 IgG. [6] Например, фукоза , содержащая гликаны CH2-84.4, создавая стерические препятствия, снижает сродство IgG к FcγRIIIA. [6] Напротив, гликаны G0, которые лишены галактозы и вместо этого оканчиваются фрагментами GlcNAc, обладают повышенным сродством к FcγRIIIA. [6]
Неонатальный Fc-рецептор
[ редактировать ]Другой FcR экспрессируется на нескольких типах клеток и по структуре подобен MHC класса I. Этот рецептор также связывает IgG и участвует в сохранении этого антитела. [7] Однако, поскольку этот рецептор Fc также участвует в передаче IgG от матери либо через плаценту к плоду , либо с молоком к грудному ребенку , его называют неонатальным рецептором Fc ( FcRn ). [8] [9] Недавно исследования показали, что этот рецептор играет роль в гомеостазе уровней IgG в сыворотке.
Fc-альфа-рецепторы
[ редактировать ]Только один рецептор Fc принадлежит к подгруппе FcαR, которая называется FcαRI (или CD89). [10] FcαRI обнаруживается на поверхности нейтрофилов , эозинофилов, моноцитов, некоторых макрофагов (включая клетки Купфера ) и некоторых дендритных клеток . [10] Он состоит из двух внеклеточных Ig-подобных доменов и является членом как суперсемейства иммуноглобулинов , так и семейства многоцепочечных рецепторов иммунного распознавания (MIRR). [3] Он передает сигналы, связываясь с двумя сигнальными цепями FcRγ. [10] Другой рецептор также может связывать IgA, хотя он имеет более высокое сродство к другому антителу, называемому IgM . [11] Этот рецептор называется рецептором Fc-альфа/мю (Fcα/μR) и представляет собой трансмембранный белок I типа . Имея один Ig-подобный домен во внеклеточной части, этот Fc-рецептор также является членом суперсемейства иммуноглобулинов. [12]
Fc-эпсилон-рецепторы
[ редактировать ]Известны два типа FcεR: [3]
- высокоаффинный рецептор FcεRI является членом суперсемейства иммуноглобулинов (имеет два Ig-подобных домена). FcεRI обнаруживается на эпидермальных клетках Лангерганса , эозинофилах, тучных клетках и базофилах. [13] [14] В результате своего клеточного распределения этот рецептор играет важную роль в контроле аллергических реакций . FcεRI также экспрессируется на антигенпрезентирующих клетках и контролирует выработку важных иммунных медиаторов, называемых цитокинами , которые способствуют воспалению . [15]
- рецептор с низким сродством FcεRII ( CD23 ) представляет собой лектин C-типа . FcεRII выполняет множество функций мембраносвязанного или растворимого рецептора; он контролирует рост и дифференцировку В-клеток и блокирует связывание IgE эозинофилов, моноцитов и базофилов. [16]
Сводная таблица
[ редактировать ]| Название рецептора | Основной лиганд антитела | Сродство к лиганду | Распределение клеток | Эффект после связывания с антителом |
| FcγRI ( CD64 ) | IgG1 и IgG3 | Высокий (Кд ~ 10 −9 М) | Макрофаги Нейтрофилы Эозинофилы Дендритные клетки | Фагоцитоз Активация клеток Активация дыхательного взрыва Индукция уничтожения микробов |
| FcγRIIA ( CD32 ) | IgG | Низкий (Kd > 10 −7 М) | Макрофаги Нейтрофилы Эозинофилы Тромбоциты клетки Лангерганса | Фагоцитоз Дегрануляция (эозинофилы) |
| FcγRIIB1 (CD32) | IgG | Низкий (Kd > 10 −7 М) | В-клетки Тучные клетки | Нет фагоцитоза Подавление активности клеток |
| FcγRIIB2 (CD32) | IgG | Низкий (Kd > 10 −7 М) | Макрофаги Нейтрофилы Эозинофилы | Фагоцитоз Подавление активности клеток |
| FcγRIIIA ( CD16a ) | IgG | Низкий (Kd > 10 −6 М) | NK-клетки Макрофаги (некоторые ткани) | Индукция антителозависимой клеточно-опосредованной цитотоксичности (ADCC) Индукция высвобождения цитокинов макрофагами |
| FcγRIIIB ( CD16b ) | IgG | Низкий (Kd > 10 −6 М) | Эозинофилы Макрофаги Нейтрофилы Тучные клетки Фолликулярные дендритные клетки | Индукция уничтожения микробов |
| FcεRI | IgE | Высокий (Кд ~ 10 −10 М) | Тучные клетки Эозинофилы Базофилы клетки Лангерганса Моноциты | Дегрануляция Фагоцитоз |
| FcεRII ( CD23 ) | IgE | Низкий (Kd > 10 −7 М) | В-клетки Эозинофилы клетки Лангерганса | Возможная молекула адгезии Транспорт IgE через эпителий кишечника человека Механизм положительной обратной связи для повышения аллергической сенсибилизации (В-клетки) |
| FcαRI ( CD89 ) | IgA | Низкий (Kd > 10 −6 М) | Моноциты Макрофаги Нейтрофилы Эозинофилы | Фагоцитоз Индукция уничтожения микробов |
| Fcα/мкР (CD351) | IgA и IgM | Высокий для IgM, средний для IgA | В-клетки Мезангиальные клетки Макрофаги | Эндоцитоз Индукция уничтожения микробов |
| FcμR [17] | IgM | (неизвестный) | Человеческий FcμR преимущественно экспрессируется лимфоцитами, но не фагоцитами. [18] | функция не полностью выяснена/разнообразна [19] |
| FcRn | IgG | с высоким содержанием кислых клеточных эндосом низкий уровень pH в нейтральной внеклеточной среде | Моноциты Макрофаги Дендритные клетки Эпителиальные клетки Эндотелиальные клетки Гепатоциты | Переносит IgG от матери к плоду через плаценту. Переносит IgG от матери к ребенку с молоком. Защищает IgG от деградации Переносит IgG через эндотелиальные/эпителиальные слои. |
Функции
[ редактировать ]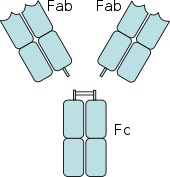
Рецепторы Fc обнаружены на ряде клеток иммунной системы, включая фагоциты , такие как макрофаги и моноциты , гранулоциты, такие как нейтрофилы и эозинофилы , а также лимфоциты врожденной иммунной системы ( естественные клетки-киллеры ) или адаптивной иммунной системы (например, В-клетки ). [20] [21] [22] Они позволяют этим клеткам связываться с антителами, которые прикрепляются к поверхности микробов или инфицированных микробами клеток, помогая этим клеткам идентифицировать и устранять микробные патогены . Рецепторы Fc связывают антитела в их Fc-участке (или хвосте), взаимодействие, которое активирует клетку, обладающую рецептором Fc. [23] Активация фагоцитов является наиболее распространенной функцией, приписываемой Fc-рецепторам. Например, макрофаги начинают поглощать и убивать IgG покрытый патоген путем фагоцитоза после задействования их Fcγ-рецепторов. [24] Другой процесс, в котором участвуют рецепторы Fc, называется антителозависимой клеточно-опосредованной цитотоксичностью (ADCC). Во время ADCC рецепторы FcγRIII на поверхности естественных клеток-киллеров (NK) стимулируют NK-клетки высвобождать цитотоксические молекулы из их гранул для уничтожения покрытых антителами клеток-мишеней. [25] FcεRI имеет другую функцию. FcεRI представляет собой рецептор Fc на гранулоцитах , который участвует в аллергических реакциях и защите от паразитарных инфекций . Когда присутствует соответствующий аллергический антиген или паразит, перекрестное связывание по крайней мере двух молекул IgE и их Fc-рецепторов на поверхности гранулоцита заставит клетку быстро высвободить предварительно сформированные медиаторы из своих гранул. [3]
Сигнальные механизмы – Fc-гамма-рецепторы
[ редактировать ]Активация
[ редактировать ]Fc-гамма-рецепторы относятся к группе некаталитических тирозин-фосфорилированных рецепторов , которые имеют сходный сигнальный путь, включающий фосфорилирование остатков тирозина. [26] Рецепторы генерируют сигналы внутри своих клеток посредством важного мотива активации, известного как мотив активации на основе тирозина иммунорецепторов (ITAM). [27] ITAM представляет собой специфическую последовательность аминокислот (YXXL), встречающуюся дважды в тесной последовательности во внутриклеточном хвосте рецептора. Когда фосфатные группы добавляются к остатку тирозина (Y) ITAM с помощью мембраносвязанных ферментов семейства киназ Src , сигнальный каскад внутри клетки генерируется . Эта реакция фосфорилирования обычно следует за взаимодействием рецептора Fc с его лигандом . ITAM присутствует во внутриклеточном хвосте FcγRIIA, и его фосфорилирование индуцирует фагоцитоз в макрофагах. FcγRI и FcγRIIIA не имеют ITAM, но могут передавать активирующий сигнал своим фагоцитам путем взаимодействия с другим белком, который имеет его. Этот адаптерный белок называется субъединицей Fcγ и, как и FcγRIIA, содержит две последовательности YXXL, характерные для ITAM.
Торможение
[ редактировать ]Присутствия только одного мотива YXXL недостаточно для активации клеток и представляет собой мотив (I/VXXYXXL), известный как ингибирующий мотив на основе иммунорецептора тирозина (ITIM). FcγRIIB1 и FcγRIIB2 имеют последовательность ITIM и представляют собой ингибирующие Fc-рецепторы; они не индуцируют фагоцитоз. Тормозное действие этих рецепторов контролируется ферментами, удаляющими фосфатные группы из остатков тирозина; фосфатазы ингибируют передачу сигналов Fcγ - SHP-1 и SHIP-1 рецепторами. [28] Связывание лиганда с FcγRIIB приводит к фосфорилированию тирозина мотива ITAM. Эта модификация создает сайт связывания фосфатазы, домен узнавания SH2. Отмена передачи сигналов активации ITAM вызвана ингибированием протеинтирозинкиназ семейства Src и гидролизом мембраны PIP3, прерывая дальнейшую нижестоящую передачу сигналов активирующими рецепторами, такими как активация FcγR, TCR, BCR и цитокиновых рецепторов (например, c-Kit ). [29]
Отрицательная передача сигналов с помощью FcγRIIB главным образом важна для регуляции активированных В-клеток. Положительная передача сигналов В-клеток инициируется связыванием чужеродного антигена с поверхностным иммуноглобулином. Секретируется одно и то же антигенспецифическое антитело, которое может подавлять обратную связь или способствовать передаче негативных сигналов. Эту негативную передачу сигналов обеспечивает FcγRIIB: [30] Эксперименты с использованием мутантов с делецией В-клеток и доминантно-негативных ферментов твердо установили важную роль инозитол-5-фосфатазы, содержащей SH2-домен (SHIP), в отрицательной передаче сигналов. Отрицательная передача сигналов через SHIP, по-видимому, ингибирует путь Ras посредством конкуренции домена SH2 с Grb2 и Shc и может включать потребление внутриклеточных липидных медиаторов, которые действуют как активаторы аллостерических ферментов или способствуют проникновению внеклеточного Ca2+. [31]
Клеточная активация
[ редактировать ]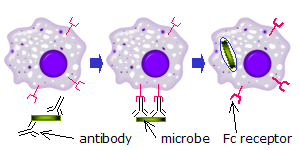
На фагоцитах
[ редактировать ]Когда молекулы IgG , специфичные к определенному антигену или поверхностному компоненту, связываются с патогеном с помощью своей Fab-области (фрагментарной антигенсвязывающей области), их Fc-области направлены наружу, в непосредственный доступ к фагоцитам . Фагоциты связывают эти области Fc со своими рецепторами Fc. [24] Между рецептором и антителом образуется множество взаимодействий с низким сродством, которые работают вместе, прочно связывая микроб, покрытый антителами. Низкое индивидуальное сродство не позволяет рецепторам Fc связывать антитела в отсутствие антигена и, следовательно, снижает вероятность активации иммунных клеток в отсутствие инфекции. Это также предотвращает агглютинацию (свертывание) фагоцитов антителами при отсутствии антигена. После связывания патогена взаимодействие между Fc-участком антитела и Fc-рецепторами фагоцита приводит к инициации фагоцитоза . Патоген поглощается фагоцитом в результате активного процесса, включающего связывание и высвобождение комплекса Fc-область/Fc-рецептор, пока клеточная мембрана фагоцита полностью не окружит патоген. [32]
На NK-клетках
[ редактировать ]Рецептор Fc на NK-клетках распознает IgG, который связан с поверхностью инфицированной патогеном клетки-мишени и называется CD16 или FcγRIII. [33] Активация FcγRIII с помощью IgG вызывает высвобождение цитокинов , таких как IFN-γ , которые передают сигнал другим иммунным клеткам, и цитотоксических медиаторов, таких как перфорин и гранзим , которые проникают в клетку-мишень и способствуют гибели клеток, запуская апоптоз . Этот процесс известен как антителозависимая клеточно-опосредованная цитотоксичность (ADCC). FcγRIII на NK-клетках также может ассоциироваться с мономерным IgG (т.е. с IgG, который не связан с антигеном). Когда это происходит, рецептор Fc ингибирует активность NK-клетки. [34]
О тучных клетках
[ редактировать ]
Антитела IgE связываются антигенами аллергенов с . Эти связанные с аллергеном молекулы IgE взаимодействуют с рецепторами Fcε на поверхности тучных клеток . Активация тучных клеток после взаимодействия с FcεRI приводит к процессу, называемому дегрануляцией , при котором тучные клетки высвобождают предварительно сформированные молекулы из своих цитоплазматических гранул; это смесь соединений, включая гистамин , протеогликаны и сериновые протеазы . [35] Активированные тучные клетки также синтезируют и секретируют медиаторы липидного происхождения (такие как простагландины , лейкотриены и фактор активации тромбоцитов ) и цитокины (такие как интерлейкин 1 , интерлейкин 3 , интерлейкин 4 , интерлейкин 5 , интерлейкин 6 , интерлейкин 13 , фактор некроза опухоли). -альфа , GM-CSF и несколько хемокинов . [36] [37] Эти медиаторы способствуют воспалению , привлекая другие лейкоциты .
Об эозинофилах
[ редактировать ]Крупные паразиты, такие как гельминт (червь) Schistosoma mansoni, слишком велики для поглощения фагоцитами. У них также есть внешняя структура, называемая покровами , которая устойчива к воздействию веществ, выделяемых макрофагами и тучными клетками. Однако эти паразиты могут покрыться IgE и распознаваться FcεRII на поверхности эозинофилов . Активированные эозинофилы высвобождают предварительно сформированные медиаторы, такие как основной основной белок и ферменты , такие как пероксидаза , к которым гельминты не устойчивы. [38] [39] Взаимодействие рецептора FcεRII с Fc-частью IgE, связанного с гельминтами, заставляет эозинофил высвобождать эти молекулы по механизму, аналогичному механизму NK-клетки во время ADCC. [40]
О Т-лимфоцитах
[ редактировать ]CD4+ Т-клетки ( зрелые Т - клетки ) помогают В-клеткам, вырабатывающим антитела. При патологии заболевания наблюдаются несколько подмножеств активированных эффекторных CD4+ Т-клеток. Более ранние исследования, обобщенные Сандерсом и Линчем в 1993 году, предположили критическую роль FcR в иммунных ответах, опосредованных CD4+ Т-клетками, и предположили образование совместного сигнального комплекса между FcR и TCR на поверхности клетки. [41] [42] [43] [44] Чаухан и коллеги сообщили о совместной локализации меченых IC с комплексом CD3 на активированной поверхности CD4+ Т-клеток, что, таким образом, предполагает сосуществование FcR вместе с комплексом TCR. [45] Оба этих рецептора образуют апикальную структуру на мембране активированных CD4+ Т-клеток, что указывает на латеральное движение этих рецепторов. [46] Совместная миграция FcR с комплексом TCR и BCR наблюдается на поверхности клеток, и цитоконъюгаты T:B-клеток демонстрируют такое сосуществование в точке контакта. [47] В более раннем обзоре было предположено, что экспрессия FcR на CD4+ Т-клетках остается открытым вопросом. [48] Это установило действующую парадигму, согласно которой Т-клетки не экспрессируют FcR, и эти результаты никогда не подвергались сомнению и экспериментальной проверке. [49] Чаухан и его коллеги показали связывание иммунных комплексов (ИК), лиганда FcR, с активированными CD4+ Т-клетками. [49] Экспрессия CD16a индуцируется в активированных наивных CD4+ Т-клетках человека, которые экспрессируют CD25, CD69 и CD98, а лигирование с IC приводит к образованию эффекторных клеток памяти. [50] Передача сигналов CD16a опосредуется фосфорилированием Syk (pSyk). [50] [51] [52]
В настоящее время исследование предполагает индуцированную экспрессию CD32a при активации CD4+ Т-клеток человека, аналогичную CD16a. [51] [53] Экспрессия CD32a на CD4+ Т-клетках также была подтверждена тремя независимыми исследованиями исследователей ВИЧ-1. Экспрессия CD16a и CD32a в подмножестве активированных CD4+ Т-клеток теперь подтверждена. [51] [53] FcR на поверхности клетки при связывании с IC, состоящими из нуклеиновых кислот, запускают выработку цитокинов и активируют пути восприятия нуклеиновых кислот. FcR присутствуют как на поверхности клетки, так и в цитозоле. Передача сигналов CD16a усиливает экспрессию нуклеиновых кислот, чувствительных к toll-подобным рецепторам, и перемещает их на поверхность клетки. [50] [54] CD16a является новым костимулирующим сигналом для CD4+ Т-клеток человека, который успешно заменяет потребность в CD28 во время аутоиммунитета. [55] На аутоиммунном фоне Т-клетки CD4+ обходят требование косигнализации CD28, чтобы полностью активироваться. [55] Более того, блокада совместной передачи сигналов CD28 не ингибирует развитие клеток TFH, ключевой субпопуляции для генерации аутоантител, продуцирующих аутореактивные плазматические В-клетки. [56] Для иммунного гомеостаза необходим баланс между костимулирующими и ингибирующими сигналами. Чрезмерная костимуляция и/или недостаточное совместное ингибирование приводит к нарушению толерантности и аутоиммунитету. Костимуляция, опосредованная CD16a, обеспечивает положительный сигнал в активированных CD4+ Т-клетках, а не в покоящихся клетках, в которых отсутствует экспрессия FcγR. [51]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Андерсон Р. (2003). «Манипулирование макромолекулами клеточной поверхности флавивирусами» . Достижения в области исследования вирусов . 59 : 229–74. дои : 10.1016/S0065-3527(03)59007-8 . ISBN 9780120398591 . ПМЦ 7252169 . ПМИД 14696331 .
- ^ Оуэн Дж., Пунт Дж., Стрэнфорд С., Джонс П. (2009). Иммунология (7-е изд.). Нью-Йорк: WH Freeman and Company. п. 423. ИСБН 978-14641-3784-6 .
- ^ Jump up to: а б с д Фридман В.Х. (сентябрь 1991 г.). «Fc-рецепторы и факторы связывания иммуноглобулинов» . Журнал ФАСЭБ . 5 (12): 2684–90. дои : 10.1096/fasebj.5.12.1916092 . ПМИД 1916092 . S2CID 16805557 .
- ^ Индик З.К., Парк Дж.Г., Хантер С., Шрайбер А.Д. (декабрь 1995 г.). «Молекулярное рассечение фагоцитоза, опосредованного гамма-рецептором Fc» . Кровь . 86 (12): 4389–99. doi : 10.1182/blood.V86.12.4389.bloodjournal86124389 . ПМИД 8541526 .
- ^ Харрисон П.Т., Дэвис В., Норман Дж.К., Хокадей А.Р., Аллен Дж.М. (сентябрь 1994 г.). «Связывание мономерного иммуноглобулина G запускает Fc гамма-RI-опосредованный эндоцитоз» . Журнал биологической химии . 269 (39): 24396–402. дои : 10.1016/S0021-9258(19)51097-3 . ПМИД 7929100 .
- ^ Jump up to: а б с д Маверакис Э., Ким К., Шимода М., Гершвин М.Е., Патель Ф., Уилкен Р., Райчаудхури С., Рухаак Л.Р., Лебрилла CB (февраль 2015 г.). «Гликаны в иммунной системе и теория аутоиммунитета с измененными гликанами: критический обзор» . Журнал аутоиммунитета . 57 (6): 1–13. дои : 10.1016/j.jaut.2014.12.002 . ПМЦ 4340844 . ПМИД 25578468 .
- ^ Чжу X, Мэн Дж., Дикинсон Б.Л., Ли X, Мидзогучи Е, Мяо Л., Ван Ю, Роберт С., Ву Б, Смит П.Д., Ленсер В.И., Блумберг Р.С. (март 2001 г.). «Неонатальный Fc-рецептор IgG, родственный MHC класса I, функционально экспрессируется в моноцитах, кишечных макрофагах и дендритных клетках» . Журнал иммунологии . 166 (5): 3266–76. дои : 10.4049/jimmunol.166.5.3266 . ПМЦ 2827247 . ПМИД 11207281 .
- ^ Фиран М., Боудон Р., Раду С., Обер Р.Дж., Икен Д., Антохе Ф., Гети В., Уорд Э.С. (август 2001 г.). «Рецептор FcRn, связанный с MHC класса I, играет важную роль в передаче гамма-глобулина у матери и плода у человека» . Международная иммунология . 13 (8): 993–1002. дои : 10.1093/интимм/13.8.993 . ПМИД 11470769 .
- ^ Симистер Н.Э., Якобовиц Исраэль Э., Ахаус Дж.К., Story CM (май 1997 г.). «Новые функции Fc-рецептора, связанного с MHC класса I, FcRn». Труды Биохимического общества . 25 (2): 481–6. дои : 10.1042/bst0250481 . ПМИД 9191140 .
- ^ Jump up to: а б с Оттен М.А., ван Эгмонд М. (март 2004 г.). «Fc-рецептор IgA (FcalphaRI, CD89)». Письма по иммунологии . 92 (1–2): 23–31. дои : 10.1016/j.imlet.2003.11.018 . ПМИД 15081523 .
- ^ Сибуя А, Хонда С (декабрь 2006 г.). «Молекулярные и функциональные характеристики Fcalpha/muR, нового рецептора Fc для IgM и IgA». Семинары Спрингера по иммунопатологии . 28 (4): 377–82. дои : 10.1007/s00281-006-0050-3 . ПМИД 17061088 . S2CID 23794895 .
- ^ Чо Ю, Усуи К, Хонда С, Тахара-Ханаока С, Сибуя К, Сибуя А (июнь 2006 г.). «Молекулярные характеристики связывания Fc IgA и IgM с Fcalpha/muR» . Связь с биохимическими и биофизическими исследованиями . 345 (1): 474–8. дои : 10.1016/j.bbrc.2006.04.084 . HDL : 2241/102010 . ПМИД 16681999 .
- ^ Очиай К., Ван Б., Ригер А., Килгус О., Маурер Д., Фёдингер Д., Кинет Дж. П., Стингл Г., Томиока Х (1994). «Обзор Fc-эпсилон-RI на эпидермальных клетках Лангерганса человека». Международный архив аллергии и иммунологии . 104 Приложение 1 (1): 63–4. дои : 10.1159/000236756 . ПМИД 8156009 .
- ^ Пруссин С., Меткалф Д.Д. (февраль 2006 г.). «5. IgE, тучные клетки, базофилы и эозинофилы». Журнал аллергии и клинической иммунологии . 117 (2 дополнительных мини-праймера): S450-6. дои : 10.1016/j.jaci.2005.11.016 . ПМИД 16455345 .
- ^ фон Бубнофф Д., Новак Н., Крафт С., Бибер Т. (март 2003 г.). «Центральная роль FcepsilonRI в аллергии». Клиническая и экспериментальная дерматология . 28 (2): 184–7. дои : 10.1046/j.1365-2230.2003.01209.x . ПМИД 12653710 . S2CID 2080598 .
- ^ Кикутани Х., Ёкота А., Учибаяши Н., Юкава К., Танака Т., Сугияма К., Барсумиан Э.Л., Суэмура М., Кисимото Т. (2007). «Структура и функция рецептора Fc II (Fc RII / CD23): точка контакта между эффекторной фазой аллергии и дифференцировкой B-клеток». Симпозиум 147 Фонда Ciba - IgE, тучные клетки и аллергическая реакция . Симпозиум Фонда Новартис. Том. 147. стр. 147-1. 23–35. дои : 10.1002/9780470513866.ch3 . ISBN 9780470513866 . ПМИД 2695308 .
- ^ «Рецептор FCMR Fc mu [Homo sapiens (Человек)] - Ген - NCBI» .
- ^ Кубагава Х., Ока С., Кубагава Ю., Тории И., Такаяма Э., Кан Д.В. и др. (2009). «Идентификация неуловимого рецептора IgMFc (FcmuR) у человека» . Дж. Эксп. Мед . 206 (12): 2779–93. дои : 10.1084/jem.20091107 . ПМК 2806608 . ПМИД 19858324 .
- ^ Лю Дж, Ван Ю, Сюн Э, Хун Р, Лу Ц, Оно Х, Ван Цзюй (2019). «Роль рецептора IgM Fc в иммунитете и толерантности» . Границы в иммунологии . 10 :529. дои : 10.3389/fimmu.2019.00529 . ПМК 6438924 . ПМИД 30967868 .
- ^ Сельварадж П., Фифадара Н., Нагараджан С., Чимино А., Ван Г. (2004). «Функциональная регуляция гамма-рецепторов нейтрофилов человека». Иммунологические исследования . 29 (1–3): 219–30. дои : 10.1385/IR:29:1-3:219 . ПМИД 15181284 . S2CID 85351071 .
- ^ Сулица А., Чемберс В.Х., Манчулеа М., Метес Д., Кори С., Рабинович Х., Уайтсайд Т.Л., Херберман Р.Б. (1995). «Дивергентные пути передачи сигнала и влияние на функции естественных клеток-киллеров, индуцированные взаимодействием рецепторов Fc с физиологическими лигандами или антирецепторными антителами». Естественный иммунитет . 14 (3): 123–33. ПМИД 8832896 .
- ^ Сарфати М., Фурнье С., Ву С.И., Делеспесс Дж. (1992). «Экспрессия, регуляция и функция антигена Fc эпсилон RII (CD23) человека». Иммунологические исследования . 11 (3–4): 260–72. дои : 10.1007/BF02919132 . ПМИД 1287120 . S2CID 83698996 .
- ^ Рагхаван М., Бьоркман П.Дж. (1996). «Fc-рецепторы и их взаимодействие с иммуноглобулинами» (PDF) . Ежегодный обзор клеточной биологии и биологии развития . 12 : 181–220. дои : 10.1146/annurev.cellbio.12.1.181 . ПМИД 8970726 .
- ^ Jump up to: а б Суонсон Дж. А., Хоппе А. Д. (декабрь 2004 г.). «Координация передачи сигналов во время фагоцитоза, опосредованного рецептором Fc». Журнал биологии лейкоцитов . 76 (6): 1093–103. дои : 10.1189/jlb.0804439 . hdl : 2027.42/141562 . ПМИД 15466916 . S2CID 13912335 .
- ^ Сан ПД (2003). «Структура и функция рецепторов естественных клеток-киллеров» . Иммунологические исследования . 27 (2–3): 539–48. дои : 10.1385/IR:27:2-3:539 . ПМИД 12857997 . S2CID 29921323 .
- ^ Душек О., Гойетт Дж., Ван дер Мерве П.А. (ноябрь 2012 г.). «Некаталитические тирозин-фосфорилированные рецепторы». Иммунологические обзоры . 250 (1): 258–276. дои : 10.1111/imr.12008 . ПМИД 23046135 . S2CID 1549902 .
- ^ Камбье Ж.К. (февраль 1995 г.). «Новая номенклатура мотива Reth (или ARH1/TAM/ARAM/YXXL)». Иммунология сегодня . 16 (2): 110. дои : 10.1016/0167-5699(95)80105-7 . ПМИД 7888063 .
- ^ Хуан З.Ю., Хантер С., Ким М.К., Индик З.К., Шрайбер А.Д. (июнь 2003 г.). «Влияние фосфатаз SHP-1 и SHIP-1 на передачу сигналов ITIM- и ITAM-содержащими Fcgamma-рецепторами FcgammaRIIB и FcgammaRIIA» . Журнал биологии лейкоцитов . 73 (6): 823–9. дои : 10.1189/jlb.0902454 . ПМИД 12773515 . S2CID 14502303 .
- ^ Камбье Ж.К. (июнь 1997 г.). «Ингибирующие рецепторы имеются в большом количестве?» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (12): 5993–5. Бибкод : 1997PNAS...94.5993C . дои : 10.1073/pnas.94.12.5993 . ПМК 33673 . ПМИД 9177155 .
- ^ Такай Т., Оно М., Хикида М., Омори Х., Равеч Дж.В. (январь 1996 г.). «Усиленные гуморальные и анафилактические реакции у мышей с дефицитом Fc гамма RII». Природа . 379 (6563): 346–9. Бибкод : 1996Natur.379..346T . дои : 10.1038/379346a0 . ПМИД 8552190 . S2CID 4364705 .
- ^ Коггешолл К.М. (июнь 1998 г.). «Ингибирующая передача сигналов B-клеточным Fc гамма RIIb». Современное мнение в иммунологии . 10 (3): 306–12. дои : 10.1016/s0952-7915(98)80169-6 . ПМИД 9638367 .
- ^ Джоши Т., Бутчар Дж. П., Тридандапани С. (октябрь 2006 г.). «Передача сигналов Fcgamma-рецептора в фагоцитах». Международный журнал гематологии . 84 (3): 210–6. дои : 10.1532/IJH97.06140 . ПМИД 17050193 . S2CID 6501210 .
- ^ Тринкьери Дж., Валианте Н. (1993). «Рецепторы Fc-фрагмента IgG на естественных клетках-киллерах». Естественный иммунитет . 12 (4–5): 218–34. ПМИД 8257828 .
- ^ Сулица А, Галатиук С, Манчуля М, Банку АС, ДеЛео А, Уайтсайд ТЛ, Херберман РБ (апрель 1993 г.). «Регуляция естественной цитотоксичности человека с помощью IgG. IV. Связь между связыванием мономерного IgG с рецепторами Fc на больших гранулярных лимфоцитах и ингибированием активности естественных клеток-киллеров (NK)». Клеточная иммунология . 147 (2): 397–410. дои : 10.1006/cimm.1993.1079 . ПМИД 8453679 .
- ^ Ямасаки С., Сайто Т. (2005). «Регуляция активации тучных клеток с помощью FcepsilonRI». Химическая иммунология и аллергия . 87 : 22–31. дои : 10.1159/000087568 . ISBN 3-8055-7948-9 . ПМИД 16107760 .
- ^ Вакахара С., Фуджи И., Накао Т., Цуритани К., Хара Т., Сайто Х., Ра С. (ноябрь 2001 г.). «Профили экспрессии генов Fc-эпсилон-RI, цитокинов и хемокинов при активации Fc-эпсилон-RI в культивируемых тучных клетках человека, полученных из периферической крови». Цитокин . 16 (4): 143–52. дои : 10.1006/cyto.2001.0958 . ПМИД 11792124 .
- ^ Меткалф Д.Д., Барам Д., Мекори Я.А. (октябрь 1997 г.). «Тучные клетки». Физиологические обзоры . 77 (4): 1033–79. дои : 10.1152/physrev.1997.77.4.1033 . ПМИД 9354811 .
- ^ Дэвид-младший, Баттерворт А.Е., Вадас М.А. (сентябрь 1980 г.). «Механизм взаимодействия, опосредующего уничтожение Schistosoma mansoni эозинофилами человека». Американский журнал тропической медицины и гигиены . 29 (5): 842–8. дои : 10.4269/ajtmh.1980.29.842 . ПМИД 7435788 .
- ^ Капрон М., Сусси Гунни А., Морита М., Труонг М.Дж., Прин Л., Кинет Дж.П., Капрон А. (1995). «Эозинофилы: от низкоаффинных к высокоаффинным рецепторам иммуноглобулина Е» . Аллергия . 50 (25 доп.): 20–3. дои : 10.1111/j.1398-9995.1995.tb04270.x . ПМИД 7677229 . S2CID 36197719 .
- ^ Гунни А.С., Ламхиуед Б., Очиаи К., Танака Ю., Делапорте Е., Капрон А., Кинет Дж.П., Капрон М. (январь 1994 г.). «Высокоаффинный рецептор IgE на эозинофилах участвует в защите от паразитов». Природа . 367 (6459): 183–6. Бибкод : 1994Natur.367..183S . дои : 10.1038/367183a0 . ПМИД 8114916 . S2CID 4331405 .
- ^ Пихлер В.Дж., Лам Л., Бродер С. (1978). «Fc-рецепторы на Т-лимфоцитах человека. I. Переход Tgamma в клетки Tmu». Дж Иммунол . 121 (4): 1540–1548. doi : 10.4049/jimmunol.121.4.1540 . ПМИД 308968 .
- ^ Сандор М., Линч Р.Г. (май 1993 г.). «Fc-рецепторы лимфоцитов: особый случай Т-клеток». Иммунол. Сегодня . 14 (5): 227–31. дои : 10.1016/0167-5699(93)90168-К . ПМИД 8517922 .
- ^ Энгельхардт В., Мацке Дж., Шмидт Р.Э. (1995). «Зависимая от активации экспрессия низкоаффинных рецепторов IgG Fc гамма RII (CD32) и Fc гамма RIII (CD16) в субпопуляциях Т-лимфоцитов человека». Иммунобиология . 192 (5): 297–320. дои : 10.1016/s0171-2985(11)80172-5 . ПМИД 7649565 .
- ^ Моретта Л., Уэбб С.Р., Гросси С.Э., Лидьярд П.М., Купер, доктор медицины (1977). «Функциональный анализ двух субпопуляций Т-клеток человека: помощь и подавление ответов В-клеток Т-клетками, несущими рецепторы для IgM или IgG» . Джей Эксп Мед . 146 (1): 184–200. дои : 10.1084/jem.146.1.184 . ПМК 2180738 . ПМИД 301544 .
- ^ Чаухан А.К., Мур Т.Л. (2011). «Активация Т-клеток терминальным комплексом комплемента и иммунными комплексами» . Журнал биологической химии . 286 (44): 38627–38637. дои : 10.1074/jbc.M111.266809 . ПМК 3207419 . ПМИД 21900254 .
- ^ Чаухан А.К., Мур Т.Л. (2011). «Активация Т-клеток терминальным комплексом комплемента и иммунными комплексами» . Журнал биологической химии . 286 (44): 38627–38637. дои : 10.1074/jbc.M111.266809 . ПМК 3207419 . ПМИД 21900254 .
- ^ Сандор М., Линч Р.Г. (1993). «Fc-рецепторы лимфоцитов: особый случай Т-клеток». Иммунология сегодня . 14 (5): 227–231. дои : 10.1016/0167-5699(93)90168-К . ПМИД 8517922 .
- ^ Ниммерьян Ф, Равеч СП (январь 2008 г.). «Fcгамма-рецепторы как регуляторы иммунных ответов». Нат. Преподобный Иммунол . 8 (1): 34–47. дои : 10.1038/nri2206 . ПМИД 18064051 . S2CID 34597359 .
- ^ Jump up to: а б Брюнс П., Йонссон Ф. (ноябрь 2015 г.). «Эффекторные функции FcR мыши и человека». Иммунол. Преподобный . 268 (1): 25–51. дои : 10.1111/imr.12350 . ПМИД 26497511 . S2CID 19544801 .
- ^ Jump up to: а б с Чаухан А.К., Мур Т.Л., Би Ю, Чен С. (январь 2016 г.). «Ко-сигнал FcγRIIIa-Syk модулирует ответ CD4+ Т-клеток и усиливает экспрессию Toll-подобного рецептора (TLR)» . Ж. Биол. Хим . 291 (3): 1368–86. дои : 10.1074/jbc.M115.684795 . ПМЦ 4714221 . ПМИД 26582197 .
- ^ Jump up to: а б с д Чаухан А.К., Чен С., Мур Т.Л., ДиПаоло Р.Дж. (февраль 2015 г.). «Индуцированная экспрессия FcγRIIIa (CD16a) на CD4+ Т-клетках запускает образование субпопуляции IFN-γhigh» . Ж. Биол. Хим . 290 (8): 5127–40. дои : 10.1074/jbc.M114.599266 . ПМЦ 4335247 . ПМИД 25556651 .
- ^ Чаухан А.К., Мур Т.Л. (2012). «Иммунные комплексы и поздние белки комплемента запускают активацию тирозинкиназы Syk в CD4(+) Т-клетках человека» . Клин Эксп Иммунол . 167 (2): 235–245. дои : 10.1111/j.1365-2249.2011.04505.x . ПМЦ 3278689 . ПМИД 22235999 .
- ^ Jump up to: а б Член парламента Хольгадо, Сананес И., Райден С., Геффнер-младший, Аррувито Л. (2018). «Лигирование CD32 способствует активации CD4+ Т-клеток» . Фронт Иммунол . 9 : 2814. дои : 10.3389/fimmu.2018.02814 . ПМК 6284025 . ПМИД 30555482 .
- ^ Чаухан АК (2017). «Передача сигналов FcgammaRIIIa модулирует эндосомальные ответы TLR в CD4+ Т-клетках человека» . Дж Иммунол . 198 (12): 4596–4606. doi : 10.4049/jimmunol.1601954 . ПМЦ 5505339 . ПМИД 28500073 .
- ^ Jump up to: а б Бур-Жордан Х., Эсенстен Дж.Х., Мартинес-Ллорделла М., Пенаранда С., Штумпф М., Блюстоун Дж.А. (2011). «Внутренний и внешний контроль толерантности периферических Т-клеток с помощью костимулирующих молекул семейства CD28/B7» . Иммунол Рев . 241 (1): 180–205. дои : 10.1111/j.1600-065X.2011.01011.x . ПМК 3077803 . ПМИД 21488898 .
- ^ Вебер Дж.П., Фурманн Ф., Файст Р.К., Ламанн А., Аль Баз М.С., Генц Л.Дж., Ву Ван Д., Магес Х.В., Хафтманн С., Ридель Р., Грюн Дж.Р., Шух В., Крочек Р.А., Радбрух А., Машреги М.Ф., Хутлофф А. 2015). «ICOS поддерживает фенотип Т-фолликулярных хелперных клеток за счет подавления Круппелеподобного фактора 2» . Журнал экспериментальной медицины . 212 (2): 217–233. дои : 10.1084/jem.20141432 . ПМЦ 4322049 . ПМИД 25646266 .
Дальнейшее чтение
[ редактировать ]- Джейнвей Калифорния, Трэверс П., Уолдпорт М., Шломчик М.Дж. (2001). «Глава 9. Гуморальный иммунный ответ» . Иммунобиология: Иммунная система в здоровье и болезнях (5-е изд.). Нью-Йорк: Гарленд. ISBN 978-0-8153-3642-6 .
- Аббас А.К., Лихтман А.Х., Пиллаи С. (2012). «Глава 12: Эффекторные механизмы гуморального иммунитета». Клеточная и молекулярная иммунология (7-е изд.). Филадельфия: Эльзевир/Сондерс. ISBN 978-1-4377-1528-6 .
- Гербер Дж. С., Моссер Д. М. (февраль 2001 г.). «Стимулирующие и тормозящие сигналы, исходящие от рецепторов Fcgamma макрофагов». Микробы и инфекции . 3 (2): 131–9. дои : 10.1016/s1286-4579(00)01360-5 . ПМИД 11251299 .
- Маверакис Э., Ким К., Шимода М., Гершвин М.Е., Патель Ф., Уилкен Р., Райчаудхури С., Рухаак Л.Р., Лебрилла CB (февраль 2015 г.). «Гликаны в иммунной системе и теория аутоиммунитета с измененными гликанами: критический обзор» . Журнал аутоиммунитета . 57 : 1–13. дои : 10.1016/j.jaut.2014.12.002 . ПМЦ 4340844 . ПМИД 25578468 .
Внешние ссылки
[ редактировать ]- Fc + Рецептор в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)