Тромбоцит
| Тромбоциты | |
|---|---|
 Изображение, полученное световым микроскопом (500 ×) окрашенного по Гимзе, мазка периферической крови, показывает тромбоциты (маленькие фиолетовые точки), окруженные эритроцитами (большие серые круглые структуры) | |
| Подробности | |
| Предшественник | Мегакариоциты |
| Функция | Образование тромбов; профилактика кровотечений |
| Идентификаторы | |
| латинский | тромбоцит |
| МеШ | D001792 |
| ФМА | 62851 |
| Анатомические термины микроанатомии | |
Тромбоциты или тромбоциты (от древнегреческого θρόμβος ( thrómbos ) «сгусток» и κύτος ( kútos ) «клетка») представляют собой компонент крови , функция которого (наряду с факторами свертывания крови ) заключается в реакции на кровотечение из- за повреждения кровеносного сосуда путем слипания, тем самым инициирование образования тромба . [1] Тромбоциты не имеют клеточного ядра ; представляют собой фрагменты цитоплазмы мегакариоцитов . [2] костного мозга или легких, [3] которые затем поступают в обращение. Тромбоциты встречаются только у млекопитающих, тогда как у других позвоночных (например, птиц , амфибий ) тромбоциты циркулируют в виде интактных мононуклеаров . [4] : 3
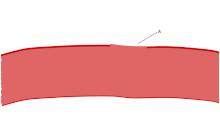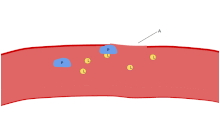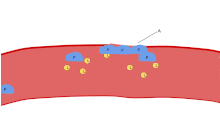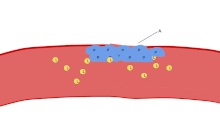
Одной из основных функций тромбоцитов является участие в гемостазе : процессе остановки кровотечения в месте прерванного эндотелия . Они собираются на объекте и, если нарушение физически не слишком велико, затыкают дыру. Во-первых, тромбоциты прикрепляются к веществам за пределами прерванного эндотелия: адгезия . Во-вторых, они меняют форму, включают рецепторы и выделяют химические посланники : активацию . В-третьих, они соединяются друг с другом посредством рецепторных мостиков: агрегация . [5] Формирование этой пробки тромбоцитов (первичный гемостаз) связано с активацией каскада свертывания крови с последующим отложением и связыванием фибрина (вторичный гемостаз). Эти процессы могут перекрываться: спектр варьируется от преимущественно тромбоцитарной пробки или «белого сгустка» до преимущественно фибрина или «красного сгустка» или более типичной смеси. Берридж добавляет ретракцию и ингибирование тромбоцитов в качестве четвертого и пятого шагов. [6] в то время как другие добавили бы шестой шаг — заживление ран . [ нужна ссылка ] Тромбоциты участвуют как во врожденных [7] и адаптивный [8] внутрисосудистые иммунные реакции.
Помимо облегчения процесса свертывания крови, тромбоциты содержат цитокины и факторы роста , которые могут способствовать заживлению ран и регенерации поврежденных тканей. [9] [10]
Срок
[ редактировать ]Термин тромбоцит (тромбоцит) вошел в употребление в начале 1900-х годов и иногда используется как синоним тромбоцита; но обычно не встречается в научной литературе, за исключением корневого слова для других терминов, связанных с тромбоцитами (например, тромбоцитопения, означающая низкий уровень тромбоцитов). [4] : v3 Термин тромбоциты подходит для мононуклеарных клеток, обнаруженных в крови немлекопитающих позвоночных: они являются функциональным эквивалентом тромбоцитов, но циркулируют как интактные клетки, а не как цитоплазматические фрагменты мегакариоцитов костного мозга. [4] : 3
В некоторых контекстах слово тромб используется взаимозаменяемо со словом сгусток , независимо от его состава (белый, красный или смешанный). В других контекстах его используют для противопоставления нормального сгустка от аномального: тромб возникает в результате физиологического гемостаза, тромбоз возникает из-за патологического и чрезмерного количества сгустка. [11] В третьем контексте оно используется для противопоставления результата и процесса: тромб — это результат, тромбоз — это процесс.
Морфология
[ редактировать ]Структура
[ редактировать ]Структурно тромбоцит можно разделить на четыре зоны, от периферической до самой внутренней: [ нужна ссылка ]
- Периферическая зона – богата гликопротеинами, необходимыми для адгезии, активации и агрегации тромбоцитов. Например, GPIb/IX/V ; ГПВИ ; ГПIIб/IIIа
- Золь-гель зона – богата микротрубочками и микрофиламентами , что позволяет тромбоцитам сохранять дискоидную форму.
- Зона органелл – богата гранулами тромбоцитов. Альфа-гранулы содержат медиаторы свертывания крови, такие как фактор V , фактор VIII , фибриноген , фибронектин , фактор роста тромбоцитов и хемотаксические агенты . Дельта-гранулы, или плотные тельца , содержат АДФ , кальций и серотонин , которые являются медиаторами, активирующими тромбоциты.
- Мембранозная зона – мембраны, происходящие из мегакариоцитов, гладкой эндоплазматической сети организованные в плотную трубчатую систему, отвечающую за синтез тромбоксана А2 . Эта плотная трубчатая система соединена с поверхностной мембраной тромбоцитов, способствуя высвобождению тромбоксана А2.
Форма
[ редактировать ]Циркулирующие инактивированные тромбоциты представляют собой двояковыпуклые дискоидные (линзообразные) структуры. [12] [4] : 117–118 Наибольший диаметр 2–3 мкм. [13] Активированные тромбоциты имеют выступы клеточной мембраны, покрывающие их поверхность.
В первом приближении форму можно считать похожей на сплюснутые сфероиды , с соотношением полуосей 2 к 8. [14] Это приближение можно использовать для моделирования гидродинамических и оптических свойств популяции, а также для восстановления геометрических параметров отдельных измеренных тромбоцитов методом проточной цитометрии . [15] Более точные биофизические модели морфологии поверхности тромбоцитов, моделирующие их форму на основе первых принципов, позволяют получить более реалистичную геометрию тромбоцитов в спокойном и активированном состоянии. [16]
Разработка
[ редактировать ]
- Производство мегакариоцитов и тромбоцитов регулируется тромбопоэтином , гормоном, вырабатываемым в почках и печени.
- Каждый мегакариоцит за свою жизнь производит от 1000 до 3000 тромбоцитов.
- В среднем 10 11 Тромбоциты производятся ежедневно у здорового взрослого человека.
- Резервные тромбоциты хранятся в селезенке и высвобождаются при необходимости в результате сокращения селезенки, вызываемого симпатической нервной системой.

- Средняя продолжительность жизни циркулирующих тромбоцитов составляет от 8 до 9 дней. [17] Продолжительность жизни отдельных тромбоцитов контролируется внутренним путем регуляции апоптоза, который имеет Bcl-x L. таймер [18]
- Старые тромбоциты разрушаются путем фагоцитоза в селезенке и печени.
Гемостаз
[ редактировать ]
Основная функция тромбоцитов — слипаться, чтобы остановить острое кровотечение. Этот процесс сложен, поскольку в динамике тромбоцитов участвуют более 193 белков и 301 взаимодействие. [5] Несмотря на значительное совпадение, функцию тромбоцитов можно смоделировать в три этапа:
Адгезия
[ редактировать ]Образование тромба на интактном эндотелии предотвращается оксидом азота . [19] простациклин , [20] и CD39 . [21]
Эндотелиальные клетки прикрепляются к субэндотелиальному коллагену с помощью фактора фон Виллебранда (ФВ), который производят эти клетки. Фактор Виллебранда также хранится в тельцах Вейбеля-Палада эндотелиальных клеток и конститутивно секретируется в кровь. Тромбоциты хранят фактор Виллебранда в своих альфа-гранулах.
Когда эндотелиальный слой разрушается, коллаген и ФВ прикрепляют тромбоциты к субэндотелию. Рецептор тромбоцитов GP1b-IX-V связывается с ФВ; а рецептор GPVI и интегрин α2β1 связываются с коллагеном. [22]
Активация
[ редактировать ]
Торможение
[ редактировать ]Неповрежденная эндотелиальная выстилка ингибирует активацию тромбоцитов путем выработки оксида азота , эндотелиальной АДФазы и PGI 2 (простациклина). Эндотелиальная АДФаза разрушает активатор тромбоцитов АДФ . [ нужна ссылка ]
Покоящиеся тромбоциты поддерживают активный отток кальция посредством циклического АМФ -активируемого кальциевого насоса. Внутриклеточная концентрация кальция определяет статус активации тромбоцитов, поскольку он является вторичным мессенджером , который вызывает конформационные изменения и дегрануляцию тромбоцитов. Эндотелиальный простациклин связывается с простаноидными рецепторами на поверхности покоящихся тромбоцитов. Это событие стимулирует связанный Gs- белок увеличивать активность аденилатциклазы и увеличивает выработку цАМФ, что еще больше способствует оттоку кальция и снижает доступность внутриклеточного кальция для активации тромбоцитов. [ нужна ссылка ]
С другой стороны, АДФ связывается с пуринергическими рецепторами на поверхности тромбоцитов. Поскольку тромбоцитарный пуринергический рецептор P2Y12 связан с Gi -белками, АДФ снижает активность аденилатциклазы тромбоцитов и выработку цАМФ, что приводит к накоплению кальция внутри тромбоцитов за счет инактивации кальциевого насоса цАМФ. Другой АДФ-рецептор P2Y1 соединяется с Gq, который активирует фосфолипазу C-бета 2 ( PLCB2 ), что приводит к образованию инозитол-1,4,5-трифосфата (IP3) и внутриклеточному высвобождению большего количества кальция. Все это вместе вызывает активацию тромбоцитов. Эндотелиальная АДФаза разрушает АДФ и предотвращает это. Клопидогрел и родственные ему антиагреганты также действуют как пуринергических рецепторов P2Y12 антагонисты . [ нужна ссылка ] Данные свидетельствуют о том, что ADP активирует путь PI3K/Akt во время первой волны агрегации, что приводит к образованию тромбина и активации PAR-1 , что вызывает вторую волну агрегации. [23]
Триггер (индукция)
[ редактировать ]Активация тромбоцитов начинается через несколько секунд после возникновения адгезии. Он запускается, когда коллаген из субэндотелия связывается с его рецепторами ( рецептор GPVI и интегрин α2β1) на тромбоцитах. GPVI связан с гамма-цепью рецептора Fc и приводит через активацию тирозинкиназного каскада, в конечном итоге, к активации PLC-гамма2 ( PLCG2 ) и большему высвобождению кальция. [ нужна ссылка ]
Тканевой фактор также связывается с фактором VII в крови, который инициирует внешний каскад свертывания крови , увеличивая выработку тромбина . Тромбин является мощным активатором тромбоцитов, действующим через Gq и G12. Это рецепторы, связанные с G-белком , и они включают опосредованные кальцием сигнальные пути внутри тромбоцитов, преодолевая исходный отток кальция. Семейства трех G-белков (Gq, Gi, G12) действуют вместе для полной активации. Тромбин также способствует вторичному укреплению тромбоцитарной пробки фибрином. Активация тромбоцитов, в свою очередь, приводит к дегрануляции и высвобождению фактора V и фибриногена , усиливая каскад свертывания крови. Закупоривание тромбоцитов и коагуляция происходят одновременно, причем каждый из них индуцирует образование окончательного тромба, сшитого фибрином. [ нужна ссылка ]
Компоненты (последствия)
[ редактировать ]Активация GPIIb/IIIa
[ редактировать ]Коллаген-опосредованная передача сигналов GPVI увеличивает выработку тромбоцитами тромбоксана А2 (TXA2) и снижает выработку простациклина . тромбоцитов Это происходит за счет изменения метаболического потока пути синтеза эйкозаноидов , который включает ферменты фосфолипазу А2 , циклооксигеназу 1 и тромбоксан-А-синтазу . Тромбоциты секретируют тромбоксан А2, который действует на собственные рецепторы тромбоксана на поверхности тромбоцитов (отсюда так называемый механизм «наружу внутрь») и на рецепторы других тромбоцитов. Эти рецепторы запускают внутритромбоцитарную передачу сигналов, которая преобразует рецепторы GPIIb/IIIa в их активную форму, чтобы инициировать агрегацию . [5]
Секреция гранул
[ редактировать ]
Тромбоциты содержат плотные гранулы , лямбда-гранулы и альфа-гранулы . Активированные тромбоциты секретируют содержимое этих гранул через свои канальцевые системы наружу. Связанные и активированные тромбоциты дегранулируются, высвобождая хемотаксические агенты тромбоцитов и привлекая больше тромбоцитов к месту повреждения эндотелия. Характеристики гранул:
- α-гранулы (альфа-гранулы) – содержат P-селектин , фактор тромбоцитов 4 , трансформирующий фактор роста-β1 , фактор роста тромбоцитов , фибронектин , B-тромбоглобулин , фактор Виллебранда , фибриноген и факторы свертывания крови V и XIII.
- δ-гранулы (дельта- или плотные гранулы) – содержат АДФ или АТФ , кальций и серотонин.
- γ-гранулы (гамма-гранулы) – похожи на лизосомы и содержат несколько гидролитических ферментов.
- Гранулы λ (лямбда-гранулы) – содержимое, участвующее в резорбции на более поздних стадиях восстановления сосудов.
Изменение морфологии
[ редактировать ]Как показали методы проточной цитометрии и электронной микроскопии , наиболее чувствительным признаком активации при воздействии на тромбоциты с помощью АДФ являются морфологические изменения. [24] Гиперполяризация митохондрий является ключевым событием в инициировании морфологических изменений. [25] Концентрация кальция внутри тромбоцитов увеличивается, стимулируя взаимодействие между комплексом микротрубочки/актиновые филаменты. Непрерывные изменения формы тромбоцитов от неактивированных до полностью активированных лучше всего наблюдать с помощью сканирующей электронной микроскопии . Три этапа на этом пути называются ранним дендритным , ранним распространением и распространением . Поверхность неактивированных тромбоцитов похожа на поверхность мозга – морщинистая из-за многочисленных неглубоких складок, которые увеличивают площадь поверхности; ранний дендритный осьминог с множеством рук и ног; ранний разворот , сырое яйцо, жаренное на сковороде, «желток» — центральное тело; и спред - приготовленная яичница с более плотной центральной частью.
Все эти изменения вызваны взаимодействием комплекса микротрубочки/актин с клеточной мембраной тромбоцитов и открытой канальцевой системой (OCS), которая представляет собой расширение и инвагинацию этой мембраны. Этот комплекс расположен прямо под этими мембранами и является химическим мотором, который вытягивает инвагинированный OCS из внутренней части тромбоцита, как выворачивает карманы брюк наизнанку, создавая дендриты. Этот процесс аналогичен механизму сокращения мышечной клетки . [26] Таким образом, вся OCS становится неотличимой от исходной мембраны тромбоцитов, поскольку она образует «жареное яйцо». Такое резкое увеличение площади поверхности происходит без растяжения и добавления фосфолипидов к мембране тромбоцитов. [27]
Взаимодействие тромбоцитов и факторов свертывания: облегчение свертывания крови
[ редактировать ]Активация тромбоцитов приводит к тому, что поверхность их мембраны становится отрицательно заряженной. Один из сигнальных путей включает скрамблазу , которая перемещает отрицательно заряженные фосфолипиды с внутренней на внешнюю поверхность мембраны тромбоцитов. Эти фосфолипиды затем связывают комплексы теназы и протромбиназы , два места взаимодействия между тромбоцитами и каскадом свертывания крови. Ионы кальция необходимы для связывания этих факторов свертывания крови.
Помимо взаимодействия с фактором Виллебранда и фибрином, тромбоциты взаимодействуют с тромбином, факторами X, Va, VIIa, XI, IX и протромбином, завершая формирование через каскад свертывания крови. [28] [29] Человеческие тромбоциты не экспрессируют тканевой фактор . [28] Крысиные тромбоциты экспрессируют белок тканевого фактора и несут как пре-мРНК тканевого фактора, так и зрелую мРНК. [30]
Агрегация
[ редактировать ]
Агрегация тромбоцитов начинается через несколько минут после активации и происходит в результате включения рецептора GPIIb/IIIa , что позволяет этим рецепторам связываться с vWF или фибриногеном . [5] Каждый тромбоцит имеет около 60 000 таких рецепторов. [31] Когда во время активации включается любой один или несколько из по меньшей мере девяти различных поверхностных рецепторов тромбоцитов, внутритромбоцитарные сигнальные пути заставляют существующие рецепторы GpIIb/IIIa менять форму – из свернутой в прямую – и, таким образом, становиться способными к связыванию. [5]
Поскольку фибриноген представляет собой палочковидный белок с узелками на обоих концах, способный связывать GPIIb/IIIa, активированные тромбоциты с обнаженным GPIIb/IIIa могут связывать фибриноген с образованием агрегатов. GPIIb/IIIa может также дополнительно закреплять тромбоциты на субэндотелиальном факторе Виллебранда для дополнительной структурной стабилизации.
Классически считалось, что это единственный механизм, участвующий в агрегации, но были идентифицированы три других механизма, которые могут инициировать агрегацию, в зависимости от скорости кровотока (т.е. диапазона сдвига). [32]
Иммунная функция
[ редактировать ]Тромбоциты играют центральную роль во врожденном иммунитете , инициируя и участвуя в многочисленных воспалительных процессах, непосредственно связывая и даже уничтожая патогены. Клинические данные показывают, что у многих пациентов с серьезными бактериальными или вирусными инфекциями наблюдается тромбоцитопения , что снижает их вклад в воспаление. Агрегаты тромбоцитов и лейкоцитов (PLA), обнаруживаемые в кровообращении, типичны при сепсисе или воспалительном заболевании кишечника , что указывает на связь между тромбоцитами и иммунными клетками. [33]
Мембрана тромбоцитов имеет рецепторы для коллагена. После разрыва стенки кровеносного сосуда тромбоциты обнажаются и прикрепляются к коллагену в окружающей ткани.
Иммунотромбоз
[ редактировать ]Поскольку гемостаз является основной функцией тромбоцитов у млекопитающих, он также находит применение при возможном сдерживании инфекции. [7] В случае травмы тромбоциты вместе с каскадом свертывания крови обеспечивают первую линию защиты, образуя тромб. Таким образом, гемостаз и защита хозяина в эволюции были переплетены. Например, у атлантического мечехвоста (возраст которого оценивается более 400 миллионов лет) единственный тип клеток крови, амебоциты , способствует как гемостатической функции, так и инкапсуляции и фагоцитозу патогенов посредством экзоцитоза внутриклеточных гранул, содержащих бактерицидную защиту. молекулы. Свертывание крови поддерживает иммунную функцию, улавливая бактерии. [34]
Хотя тромбоз, свертывание крови в интактных сосудах, обычно рассматривают как патологический иммунный ответ, приводящий к обтурации просвета сосуда и последующему гипоксическому повреждению тканей, в ряде случаев направленный тромбоз, называемый иммунотромбозом, может локально контролировать распространение тромбоза. инфекция. Тромбоз направлен в соответствии тромбоцитов, нейтрофилов и моноцитов . Процесс инициируется либо иммунными клетками путем активации их рецепторов распознавания образов (PRR), либо путем связывания тромбоцитов с бактериями. Тромбоциты могут связываться с бактериями напрямую через тромбоцитарные PRR. [33] и бактериальные поверхностные белки или через белки плазмы, которые связываются как с тромбоцитами, так и с бактериями. [35] Моноциты реагируют на молекулярные паттерны, связанные с бактериальными патогенами (PAMP) или молекулярные паттерны, связанные с повреждением (DAMP), активируя внешний путь коагуляции. Нейтрофилы облегчают свертывание крови посредством НЕТоза , а тромбоциты облегчают НЕТоз нейтрофилов. Сети связывают тканевой фактор, связывая центры свертывания крови с местом инфекции. Они также активируют внутренний путь свертывания крови, обеспечивая отрицательно заряженную поверхность фактора XII. Другие выделения нейтрофилов, такие как протеолитические ферменты, расщепляющие ингибиторы свертывания крови, также способствуют этому процессу. [7]
При дисбалансе всей регуляции иммунотромбоза этот процесс может стать аберрантным. Предполагается, что регуляторные дефекты иммунотромбоза являются основным фактором патологического тромбоза в таких формах, как диссеминированное внутрисосудистое свертывание крови (ДВС-синдром) или тромбоз глубоких вен . ДВС-синдром при сепсисе является ярким примером как нарушения регуляции процесса свертывания крови, так и чрезмерной системной воспалительной реакции, приводящей к образованию множества микротромбов, сходных по составу с таковым при физиологическом иммунотромбозе – фибрина, тромбоцитов, нейтрофилов и НЭО. [7]
Воспаление
[ редактировать ]Тромбоциты быстро перемещаются в места повреждения или инфекции и потенциально модулируют воспалительные процессы, взаимодействуя с лейкоцитами и секретируя цитокины , хемокины и другие медиаторы воспаления. [36] [37] [38] [39] [40] Тромбоциты также секретируют фактор роста тромбоцитов (PDGF).
Тромбоциты модулируют нейтрофилы, образуя тромбоцитарно-лейкоцитарные агрегаты (PLA). Эти образования индуцируют повышенную продукцию интегрина αmβ2 ( Mac-1 ) в нейтрофилах. Взаимодействие с PLA также вызывает дегрануляцию и усиление фагоцитоза нейтрофилов.
Тромбоциты являются крупнейшим источником растворимого CD40L , который индуцирует выработку активных форм кислорода (АФК) и усиливает экспрессию молекул адгезии, таких как E-селектин , ICAM-1 и VCAM-1 , в нейтрофилах, активирует макрофаги и активирует цитотоксический ответ в клетках нейтрофилов. Т- и В-лимфоциты . [33]
Тромбоциты млекопитающих, лишенные ядра, способны осуществлять автономное передвижение. [41] Тромбоциты являются активными поглотителями, очищающими стенки кровеносных сосудов и реорганизующими тромб. Они способны распознавать многие поверхности, включая бактерии, и прикрепляться к ним, а также могут окутывать их в свою открытую канальцевую систему (OCP), что приводит к предложению назвать этот процесс коверцитозом ( OCS), а не фагоцитозом, поскольку OCS - это просто инвагинация. внешней плазматической мембраны. Эти пучки тромбоцитов и бактерий обеспечивают платформу взаимодействия нейтрофилов, которые уничтожают бактерии с помощью НЕТоза и фагоцитоза.
Тромбоциты также участвуют в хронических воспалительных заболеваниях, таких как синовит или ревматоидный артрит . [42] Тромбоциты активируются гликопротеином рецептора коллагена IV (GPVI). Провоспалительные микровезикулы тромбоцитов вызывают постоянную секрецию цитокинов из соседних фибробластоподобных синовиоцитов , наиболее заметно IL-6 и IL-8 . Воспалительное повреждение окружающего внеклеточного матрикса постоянно приводит к увеличению количества коллагена, поддерживая выработку микровезикул.
Адаптивный иммунитет
[ редактировать ]Активированные тромбоциты способны участвовать в адаптивном иммунитете , взаимодействуя с антителами . Они способны специфически связывать IgG через FcγRIIA , рецептор константного фрагмента IgG (Fc). При активации и связывании с опсонизированными IgG бактериями тромбоциты высвобождают активные формы кислорода (АФК), антимикробные пептиды, дефензины , киноцидины и протеазы , непосредственно убивая бактерии. [43] Тромбоциты также секретируют провоспалительные и прокоагулянтные медиаторы, такие как неорганические полифосфаты или фактор тромбоцитов 4 (PF4), связывая врожденный и адаптивный иммунные реакции. [43] [44]
Признаки и симптомы расстройств
[ редактировать ]Спонтанные и обильные кровотечения могут возникать из-за нарушений тромбоцитов. Это кровотечение может быть вызвано недостаточным количеством тромбоцитов, дисфункциональными тромбоцитами или плотностью тромбоцитов более 1 миллиона / микролитр. (Чрезмерное количество создает относительный дефицит фактора фон Виллебранда из-за секвестрации.) [45] [46]
Кровотечение, вызванное нарушением тромбоцитов или нарушением фактора свертывания крови, можно отличить по характеристикам и локализации кровотечения. [4] : 815, Таблица 39-4 Тромбоцитарное кровотечение включает в себя кровотечение из пореза, быстрое и обильное, но его можно остановить давлением; спонтанное кровотечение на кожу, вызывающее пурпурное пятно, названное по размеру: петехии , пурпура , экхимозы ; кровотечение на слизистые оболочки, вызывающее кровоточивость десен, носовое кровотечение и желудочно-кишечное кровотечение; меноррагия; и интраретинальное и внутричерепное кровотечение.
Чрезмерное количество тромбоцитов и/или нормальные тромбоциты, реагирующие на аномальные стенки сосудов, могут привести к венозному тромбозу и артериальному тромбозу . Симптомы зависят от места тромбоза.
Измерение и тестирование
[ редактировать ]Измерение
[ редактировать ]Концентрацию тромбоцитов в крови (т.е. количество тромбоцитов) можно измерить вручную с помощью гемоцитометра или путем помещения крови в автоматический анализатор тромбоцитов с использованием подсчета частиц, например счетчика Коултера , или оптических методов. [47] Наиболее распространенные методы анализа крови включают в себя подсчет тромбоцитов, обычно обозначаемый как PLT . [48]
Концентрация тромбоцитов варьируется у разных людей и с течением времени: в среднем в популяции она составляет от 250 000 до 260 000 клеток на мм. 3 (эквивалент микролитра), но типичный лабораторно принятый нормальный диапазон составляет от 150 000 до 400 000 клеток на мм. 3 или 150–400 × 10 9 за литр. [48] [47]

На окрашенном мазке крови тромбоциты выглядят как темно-фиолетовые пятна размером около 20% от диаметра эритроцитов. Мазок выявляет размер, форму, качественное количество и комкование . Здоровый взрослый человек обычно имеет в 10–20 раз больше эритроцитов, чем тромбоцитов.
Время кровотечения
[ редактировать ]Время кровотечения было разработано Дьюком в 1910 году как тест функции тромбоцитов. [49] В тесте Дьюка измерялось время, необходимое для остановки кровотечения из стандартизированной раны на мочке уха, которую промокали каждые 30 секунд, считая нормой менее 3 минут. [50] Нормальное время кровотечения отражает достаточное количество и функцию тромбоцитов, а также нормальную микроциркуляторную сеть .
Многоэлектродная агрегометрия
[ редактировать ]При многоэлектродной агрегометрии цельную кровь с антикоагулянтом смешивают с физиологическим раствором и агонистом тромбоцитов в одноразовой кювете с двумя парами электродов. Увеличение импеданса между электродами по мере агрегации на них тромбоцитов измеряется и визуализируется в виде кривой. [51] [52]
| АДП | Адреналин | Коллаген | Ристоцетин | |
|---|---|---|---|---|
| рецептора P2Y Дефект [53] (включая клопидогрель ) | Снижение | Нормальный | Нормальный | Нормальный |
| адренергических рецепторов Дефект [53] | Нормальный | Снижение | Нормальный | Нормальный |
| рецептора коллагена Дефект [53] | Нормальный | Нормальный | Снижено или отсутствует | Нормальный |
| Нормальный | Нормальный | Нормальный | Снижено или отсутствует | |
| Снижение | Снижение | Снижение | Нормальный или сниженный | |
| Недостаток пула хранения [54] | Отсутствие второй волны | Частичный | ||
| Аспирин или аспириноподобное расстройство | Отсутствие второй волны | Отсутствующий | Нормальный | |
Агрегометрия светопропускания
[ редактировать ]При светотрансмиссионной агрегометрии (LTA) богатая тромбоцитами плазма помещается между источником света и фотоэлементом . Неагрегированная плазма пропускает относительно мало света. После добавления агониста тромбоциты агрегируют, увеличивая светопропускание, что фиксируется фотоэлементом. [55]
Импедансная агрегометрия цельной крови
[ редактировать ]Импедансная агрегометрия цельной крови (WBA) измеряет изменение электрического импеданса между двумя электродами, когда агрегация тромбоцитов индуцируется агонистом. Люмиагрегометрия цельной крови может повысить чувствительность теста к нарушению секреции тромбоцитарных гранул. [56]
ПФА-100
[ редактировать ]PFA -100 (Анализ функции тромбоцитов – 100) представляет собой систему для анализа функции тромбоцитов, в которой цитратная цельная кровь аспирируется через одноразовый картридж, содержащий отверстие внутри мембраны, покрытой либо коллагеном и адреналином, либо коллагеном и АДФ. Эти агонисты вызывают адгезию, активацию и агрегацию тромбоцитов, что приводит к быстрой окклюзии отверстия и прекращению кровотока, называемому временем закрытия (CT). Повышенный КТ с ЭПИ и коллагеном может указывать на внутренние дефекты, такие как болезнь фон Виллебранда , уремия или ингибиторы циркулирующих тромбоцитов. Последующий тест с участием коллагена и АДФ используется, чтобы определить, было ли аномальное КТ с коллагеном и ЭПИ вызвано воздействием ацетилсульфосалициловой кислоты (аспирина) или лекарств, содержащих ингибиторы. [57]
расстройства
[ редактировать ]Низкая концентрация тромбоцитов называется тромбоцитопенией и возникает либо из-за снижения выработки, либо из-за повышенного разрушения. Повышенная концентрация тромбоцитов называется тромбоцитозом и бывает либо врожденной , реактивной (на цитокины ), либо обусловлена нерегулируемой выработкой: одним из миелопролиферативных новообразований или некоторыми другими миелоидными новообразованиями . Нарушение функции тромбоцитов называется тромбоцитопатией или нарушением функции тромбоцитов. [4] : vii
Нормальные тромбоциты могут реагировать на аномалии на стенке сосуда, а не на кровоизлияние, что приводит к неадекватной адгезии/активации тромбоцитов и тромбозу : образованию сгустка внутри неповрежденного сосуда. Этот тип тромбоза возникает по механизмам, отличным от механизмов нормального тромба: расширение фибрина венозного тромбоза ; расширение нестабильной или разорвавшейся артериальной бляшки, вызывающее артериальный тромбоз ; и микроциркуляторный тромбоз. Артериальный тромб может частично блокировать кровоток, вызывая ишемию ниже по течению , или может полностью блокировать его, вызывая гибель тканей ниже по течению .: [4] : vii
Три широкие категории нарушений тромбоцитов: «недостаточно», «дисфункциональны» и «слишком много». [4] : vii
Тромбоцитопения
[ редактировать ]- Иммунная тромбоцитопения (ИТП) – ранее известная как иммунная тромбоцитопеническая пурпура и идиопатическая тромбоцитопеническая пурпура.
- Спленомегалия
- Семейная тромбоцитопения [58] [59]
- Химиотерапия
- Бабезиоз
- Лихорадка Денге
- Ониялай
- Тромботическая тромбоцитопеническая пурпура
- HELLP-синдром
- Гемолитико-уремический синдром
- Лекарственная тромбоцитопеническая пурпура (пять известных препаратов, наиболее проблематичной является гепарин-индуцированная тромбоцитопения (ГИТ)
- Связанный с беременностью
- Неонатальный аллоиммунный ассоциированный
- Апластическая анемия
- Связанный с переливанием крови
- Псевдотромбоцитопения
- Вакцино-индуцированная иммунная тромботическая тромбоцитопения (ВИТТ)
Изменение функции тромбоцитов (тромбоцитопатия)
[ редактировать ]- Врожденный
- Нарушения адгезии
- Нарушения активации
- Нарушения количества или высвобождения гранул
- Синдром Германского-Пудлака
- Синдром серых тромбоцитов
- Дефект рецептора АДФ
- Снижение активности циклооксигеназы
- Дефицит пула хранения тромбоцитов
- Нарушения агрегации
- Нарушения свертывающей активности
- Приобретенный
- Нарушения адгезии
- Пароксизмальная ночная гемоглобинурия
- Астма [60]
- Респираторное заболевание, обостряющееся приемом аспирина (AERD/триада Самтера) [61]
- Рак [62]
- Малярия [63]
- Снижение активности циклооксигеназы
- Нарушения адгезии
Тромбоцитоз и тромбоцитемия
[ редактировать ]- Реактивный
- Хроническая инфекция
- Хроническое воспаление
- Злокачественное новообразование
- Гипоспленизм (после спленэктомии)
- Дефицит железа
- Острая кровопотеря
- Миелопролиферативные новообразования – тромбоциты одновременно повышены и активированы.
- Связан с другими миелоидными новообразованиями.
- Врожденный
Фармакология
[ редактировать ]Противовоспалительные препараты
[ редактировать ]Некоторые препараты, используемые для лечения воспаления, имеют нежелательный побочный эффект, заключающийся в подавлении нормальной функции тромбоцитов. Это нестероидные противовоспалительные препараты (НПВП). Аспирин необратимо нарушает функцию тромбоцитов, ингибируя циклооксигеназу -1 (ЦОГ1) и, следовательно, нормальный гемостаз. Образующиеся тромбоциты не способны производить новую циклооксигеназу, поскольку у них нет ДНК. Нормальная функция тромбоцитов не возвращается до тех пор, пока прием аспирина не прекратится и достаточное количество пораженных тромбоцитов не будет заменено новыми, что может занять более недели. Ибупрофен , другой НПВП , не имеет такого длительного эффекта: функция тромбоцитов обычно возвращается в течение 24 часов. [64] а прием ибупрофена перед приемом аспирина предотвращает необратимые эффекты аспирина. [65]
Препараты, подавляющие функцию тромбоцитов
[ редактировать ]Эти препараты используются для предотвращения образования тромбов.
Пероральные агенты
[ редактировать ]Препараты, стимулирующие выработку тромбоцитов
[ редактировать ]Внутривенные средства
[ редактировать ]- Абциксимаб
- Эптифибатид
- Тирофибан
- Прочие: опрелвекин , ромиплостим , элтромбопаг , аргатробан.
Терапия
[ редактировать ]Переливание
[ редактировать ]Показания
[ редактировать ]Переливание тромбоцитов чаще всего используется для коррекции необычно низкого количества тромбоцитов или для предотвращения спонтанного кровотечения (обычно при количестве менее 10×10). 9 /L) или в ожидании медицинских процедур, которые обязательно связаны с кровотечением. Например, у пациентов, перенесших операцию , уровень ниже 50×10 9 /L связан с аномальным хирургическим кровотечением, и процедур регионарной анестезии, таких как эпидуральная анестезия. при уровнях ниже 80×10 следует избегать 9 /Л. [66] Тромбоциты также можно переливать, если количество тромбоцитов в норме, но тромбоциты дисфункциональны, например, когда человек принимает аспирин или клопидогрель . [67] Наконец, тромбоциты могут быть перелиты в рамках протокола массивной переливания , при котором три основных компонента крови (эритроциты, плазма и тромбоциты) переливаются для устранения тяжелого кровотечения . Переливание тромбоцитов противопоказано при тромботической тромбоцитопенической пурпуре (ТТП), поскольку оно усиливает коагулопатию . Переливание тромбоцитов обычно неэффективно и, следовательно, противопоказано для профилактики иммунной тромбоцитопении (ИТП), поскольку перелитые тромбоциты немедленно выводятся из организма; однако он показан для лечения кровотечений. [68]
Коллекция
[ редактировать ]
Тромбоциты либо выделяют из собранных единиц цельной крови и объединяют для получения терапевтической дозы, либо собирают путем афереза тромбоцитов : кровь берется у донора, пропускается через устройство, которое удаляет тромбоциты, а остаток возвращается донору в замкнутый контур. Промышленный стандарт предусматривает проверку тромбоцитов на наличие бактерий перед переливанием крови, чтобы избежать септических реакций, которые могут быть фатальными. Недавно отраслевые стандарты AABB для банков крови и служб переливания крови (5.1.5.1) разрешили использование технологии снижения количества патогенов в качестве альтернативы бактериальному скринингу тромбоцитов. [69]
Объединенные тромбоциты цельной крови, иногда называемые «случайными» тромбоцитами, разделяются одним из двух методов. [70] В США порцию цельной крови помещают в большую центрифугу , что называется «мягким вращением». При таких настройках тромбоциты остаются во взвешенном состоянии в плазме. ( Богатая тромбоцитами плазма PRP) отделяется от эритроцитов, затем центрифугируется на более высокой скорости для сбора тромбоцитов из плазмы. В других регионах мира порция цельной крови центрифугируется с использованием настроек, которые заставляют тромбоциты суспендироваться в слое « лейкоцитной пленки », который включает тромбоциты и лейкоциты. «Лейкочную пленку» изолируют в стерильном пакете, суспендируют в небольшом количестве эритроцитов и плазмы, затем снова центрифугируют, чтобы отделить тромбоциты и плазму от эритроцитов и лейкоцитов. Независимо от исходного способа приготовления, несколько донаций могут быть объединены в один контейнер с помощью стерильного соединительного устройства для изготовления единого препарата с необходимой терапевтической дозой.
Тромбоциты для афереза собираются с помощью механического устройства, которое забирает кровь у донора и центрифугирует собранную кровь для отделения тромбоцитов и других компонентов, подлежащих сбору. Оставшаяся кровь возвращается донору. Преимущество этого метода состоит в том, что одна донация обеспечивает по крайней мере одну терапевтическую дозу, в отличие от многократной донации тромбоцитов цельной крови. Это означает, что реципиент контактирует с меньшим количеством доноров и имеет меньший риск заболеваний, передающихся при переливании крови, и других осложнений. Иногда человек, например больной раком , которому требуется регулярное переливание тромбоцитов, получает повторные донации от конкретного донора, чтобы минимизировать риск. Уменьшение количества патогенов в тромбоцитах с помощью, например, обработки рибофлавином и ультрафиолетовым излучением может снизить инфекционную нагрузку патогенов, содержащихся в продуктах донорской крови. [71] [72] Другой процесс фотохимической обработки с использованием амотосалена и УФА-излучения был разработан для инактивации вирусов, бактерий, паразитов и лейкоцитов. [73] Кроме того, аферезные тромбоциты, как правило, содержат меньше загрязняющих эритроцитов, поскольку метод сбора более эффективен, чем центрифугирование с мягким вращением.
Хранилище
[ редактировать ]Тромбоциты, собранные любым методом, имеют типичный срок хранения пять дней. Это приводит к нехватке поставок, поскольку тестирование пожертвований часто занимает целый день. Эффективных консервантов для тромбоцитов не разработано.
Тромбоциты хранятся при постоянном перемешивании при температуре 20–24 °C (68–75 °F). Препараты нельзя хранить в холодильнике, поскольку это приводит к изменению формы тромбоцитов и потере их функции. Хранение при комнатной температуре создает среду, в которой любые занесенные бактерии могут размножаться и впоследствии вызывать бактериемию . Соединенные Штаты требуют, чтобы продукты перед переливанием проверялись на наличие бактериального заражения. [74]

Доставка
[ редактировать ]Тромбоциты не обязательно должны принадлежать к той же группе крови АВО, что и реципиент, или быть перекрестно совместимыми для обеспечения иммунной совместимости между донором и реципиентом, если только они не содержат значительное количество эритроцитов (эритроцитов). Присутствие эритроцитов придает продукту красновато-оранжевый цвет и обычно связано с тромбоцитами цельной крови. На некоторых участках могут набираться тромбоциты, но это не критично.
Перед выдачей тромбоцитов реципиенту их можно облучить, чтобы предотвратить развитие реакции «трансплантат против хозяина», связанной с переливанием крови , или их можно промыть для удаления плазмы.
Изменение количества тромбоцитов у реципиента после переливания называется «приращением» и рассчитывается путем вычитания количества тромбоцитов до переливания из количества тромбоцитов после переливания. На прирост влияют многие факторы, включая размер тела, количество перелитых тромбоцитов и клинические особенности, которые могут вызвать преждевременное разрушение перелитых тромбоцитов. Когда реципиенты не демонстрируют адекватного прироста после трансфузии, это называется рефрактерностью к переливанию тромбоцитов .
Тромбоциты, полученные как путем афереза, так и от случайных доноров, могут быть обработаны посредством процесса уменьшения объема. В этом процессе тромбоциты вращаются в центрифуге и удаляется плазма, оставляя от 10 до 100 мл концентрата тромбоцитов. Такие тромбоциты с уменьшенным объемом обычно переливают только новорожденным и детям, когда большой объем плазмы может перегрузить малую систему кровообращения ребенка. Меньший объем плазмы также снижает вероятность возникновения неблагоприятной трансфузионной реакции на белки плазмы. [75] Срок хранения тромбоцитов с уменьшенным объемом составляет четыре часа. [76]
Ремонт ран
[ редактировать ]Сгусток крови — лишь временное решение для остановки кровотечения; требуется восстановление тканей. Небольшие нарушения в эндотелии регулируются физиологическими механизмами; большие перерывы со стороны хирурга-травматолога. [77] Фибрин медленно растворяется фибринолитическим ферментом плазмином , а тромбоциты очищаются путем фагоцитоза . [78]
Тромбоциты высвобождают фактор роста тромбоцитов (PDGF), мощный хемотаксический агент; и TGF бета , который стимулирует отложение внеклеточного матрикса ; фактор роста фибробластов , инсулиноподобный фактор роста 1 тромбоцитов , эпидермальный фактор роста и фактор роста эндотелия сосудов . Местное применение этих факторов в повышенных концентрациях через плазму, богатую тромбоцитами (PRP), используется в качестве вспомогательного средства при заживлении ран. [79]
немлекопитающие
[ редактировать ]Вместо тромбоцитов у немлекопитающих позвоночных имеются ядросодержащие тромбоциты, которые по морфологии напоминают В-лимфоциты . Они агрегируют в ответ на тромбин, но не на АДФ, серотонин или адреналин, как это делают тромбоциты. [80] [81]
История
[ редактировать ]- Джордж Гулливер в 1841 году нарисовал тромбоциты. [82] с помощью двухлинзового (составного) микроскопа, изобретенного в 1830 году Джозефом Джексоном Листером . [83] Этот микроскоп настолько улучшил разрешение, что впервые можно было увидеть тромбоциты.
- Уильям Аддисон в 1842 году нарисовал изображение тромбоцитарно-фибринового сгустка. [84]
- Лайонел Бил в 1864 году первым опубликовал рисунок тромбоцитов. [85]
- Макс Шульце в 1865 году описал то, что он назвал «сферулами», которые, как он отметил, были намного меньше эритроцитов, иногда слипались и иногда обнаруживались в скоплениях фибринового материала. [86]
- Джулио Биццозеро в 1882 году изучал кровь амфибий микроскопически in vivo . Он назвал сферулы Шульца (ит.) пиастрином : маленькие пластинки. [87] [88] Биццозеро, возможно, предложил название Blutplattchen. [89]
- Уильям Ослер наблюдал тромбоциты и в опубликованных лекциях в 1886 году назвал их третьим тельцем и кровяной бляшкой ; и описал их как «бесцветный протоплазматический диск». [90]
- Джеймс Райт исследовал мазки крови, используя краситель, названный в его честь, и использовал терминальные пластинки в своей публикации 1906 года: [91] переход на тромбоциты в его публикации 1910 года. [92]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Лаки К. (декабрь 1972 г.). «Наше древнее наследие в области свертывания крови и некоторые его последствия». Анналы Нью-Йоркской академии наук . 202 (1): 297–307. Бибкод : 1972NYASA.202..297L . дои : 10.1111/j.1749-6632.1972.tb16342.x . ПМИД 4508929 . S2CID 45051688 .
- ^ Махлус К.Р., Тон Дж.Н., Итальяно Дж.Э. (апрель 2014 г.). «Интерпретация танца развития мегакариоцитов: обзор клеточных и молекулярных процессов, опосредующих образование тромбоцитов». Британский журнал гематологии . 165 (2): 227–236. дои : 10.1111/bjh.12758 . ПМИД 24499183 . S2CID 42595581 .
- ^ Лефрансэ, Эмма; Ортис-Муньос, Гваделупа; Кодрилье, Аксель; Маллавия, Беньят; Лю, Фэнчунь; Сайя, Дэвид М.; Торнтон, Эмили Э.; Хедли, Марк Б.; Дэвид, Тово; Кофлин, Шон Р.; Краммель, Мэтью Ф. (апрель 2017 г.). «Легкие являются местом биогенеза тромбоцитов и резервуаром гемопоэтических предшественников» . Природа . 544 (7648): 105–109. Бибкод : 2017Natur.544..105L . дои : 10.1038/nature21706 . ISSN 1476-4687 . ПМК 5663284 . ПМИД 28329764 .
- ^ Jump up to: а б с д и ж г час Майкельсон, Алан Д. (2013). Тромбоциты (3-е изд.). Академический. ISBN 978-0123878373 .
- ^ Jump up to: а б с д и Йип Дж., Шен Ю., Берндт М.К., Эндрюс Р.К. (февраль 2005 г.). «Первичные рецепторы адгезии тромбоцитов» . ИУБМБ Жизнь . 57 (2): 103–108. дои : 10.1080/15216540500078962 . ПМИД 16036569 . S2CID 12054259 .
- ^ Берридж, Майкл Дж. (1 октября 2014 г.). «Модуль 11: Клеточный стресс, воспалительные реакции и гибель клеток» (PDF) . Биология клеточной сигнализации . Том. 6. Портленд Пресс. стр. 11-1–11-30. doi : 10.1042/csb0001011 (неактивен 11 февраля 2024 г.).
{{cite book}}: CS1 maint: DOI неактивен по состоянию на февраль 2024 г. ( ссылка )
- ^ Jump up to: а б с д Гертнер Ф., Массберг С. (декабрь 2016 г.). «Свертывание крови при иммунотромбозах – на переднем крае внутрисосудистого иммунитета». Семинары по иммунологии . 28 (6): 561–569. дои : 10.1016/j.smim.2016.10.010 . ПМИД 27866916 .
- ^ Хэмптон Т. (апрель 2018 г.). «Роль тромбоцитов в адаптивном иммунитете может способствовать сепсису и шоку». ДЖАМА . 319 (13): 1311–1312. дои : 10.1001/jama.2017.12859 . ПМИД 29614158 .
- ^ Цечерска-Херич Е, Гошка М, Долеговска Б (2022). «Применение регенеративной способности тромбоцитов в современной медицине». Обзоры цитокинов и факторов роста . 64 : 84–94. doi : 10.1016/j.cytogfr.2021.11.003 . ПМИД 34924312 .
- ^ Сюй Дж, Гоу Л, Цю С (2020). «Богатая тромбоцитами плазма и регенеративная стоматология» . Австралийский стоматологический журнал . 65 (2): 131–142. дои : 10.1111/прилаг.12754 . ПМК 7384010 . ПМИД 32145082 .
- ^ Furie B, Furie BC (август 2008 г.). «Механизмы тромбообразования». Медицинский журнал Новой Англии . 359 (9): 938–949. дои : 10.1056/NEJMra0801082 . ПМИД 18753650 .
- ^ Джайн, Северная Каролина (июнь 1975 г.). «Сканирующее электронное микроскопическое исследование тромбоцитов некоторых видов животных». Тромбоз и геморрагический диатез . 33 (3): 501–507. ПМИД 1154309 .
- ^ Паулюс Дж. М. (сентябрь 1975 г.). «Размер тромбоцитов у человека» . Кровь . 46 (3): 321–336. дои : 10.1182/blood.V46.3.321.321 . ПМИД 1097000 .
- ^ Фроймович М.М. (1976). «Геометрия нормальных тромбоцитов млекопитающих по данным количественных микроскопических исследований» . Биофизический журнал . 16 (9): 1071–1089. Бибкод : 1976BpJ....16.1071F . дои : 10.1016/s0006-3495(76)85756-6 . ПМЦ 1334946 . ПМИД 786400 .
- ^ Москаленский А.Е., Юркин М.А., Конохова А.И., Строкотов Д.И., Некрасов В.М., Чернышев А.В., Цветовская Г.А., Чикова Е.Д., Мальцев В.П. (2013). «Точное измерение объема и формы покоящихся и активированных тромбоцитов крови по светорассеянию» . Журнал биомедицинской оптики . 18 (1): 017001. Бибкод : 2013JBO....18a7001M . дои : 10.1117/1.JBO.18.1.017001 . ПМИД 23288415 . S2CID 44626047 .
- ^ Москаленский А.Е., Юркин М.А., Мулюков А.Р., Литвиненко А.Л., Некрасов В.М., Чернышев А.В., Мальцев В.П. (2018). «Метод моделирования формы тромбоцитов и ее эволюции при активации» . PLOS Вычислительная биология . 14 (3): e1005899. Бибкод : 2018PLSCB..14E5899M . дои : 10.1371/journal.pcbi.1005899 . ПМК 5860797 . ПМИД 29518073 .
- ^ Харкер Л.А., Роскос Л.К., Маржец У.М., Картер Р.А., Черри Дж.К., Санделл Б., Чунг Э.Н., Терри Д., Шеридан В. (апрель 2000 г.). «Влияние фактора роста и развития мегакариоцитов на производство тромбоцитов, продолжительность жизни тромбоцитов и функцию тромбоцитов у здоровых добровольцев». Кровь . 95 (8): 2514–2522. дои : 10.1182/blood.V95.8.2514 . ПМИД 10753829 .
- ^ Мейсон К.Д., Карпинелли М.Р., Флетчер Дж.И., Коллиндж Дж.Э., Хилтон А.А., Эллис С., Келли П.Н., Экерт П.Г., Меткалф Д., Робертс А.В., Хуанг Д.С., Кайл Б.Т. (март 2007 г.). «Запрограммированная гибель ануклеарных клеток ограничивает продолжительность жизни тромбоцитов» . Клетка . 128 (6): 1173–1186. дои : 10.1016/j.cell.2007.01.037 . ПМИД 17382885 . S2CID 7492885 .
- ^ Палмер Р.М., Ферриге АГ, Монкада С (1987). «Высвобождение оксида азота отвечает за биологическую активность релаксирующего фактора эндотелиального происхождения». Природа . 327 (6122): 524–526. Бибкод : 1987Natur.327..524P . дои : 10.1038/327524a0 . ПМИД 3495737 . S2CID 4305207 .
- ^ Джонс С.И., Барретт Н.Е., Мораес Л.А., Гиббинс Дж.М., Джексон Д.Э. (2012). «Эндогенные тормозные механизмы и регуляция функции тромбоцитов». Тромбоциты и мегакариоциты . Методы молекулярной биологии. Том. 788. стр. 341–366. дои : 10.1007/978-1-61779-307-3_23 . ISBN 978-1-61779-306-6 . ПМИД 22130718 .
- ^ Маркус А.Дж., Брукман М.Дж., Дросопулос Дж.Х., Олсон К.Е., Ислам Н., Пинский DJ, Леви Р. (апрель 2005 г.). «Роль CD39 (NTPDase-1) в тромборегуляции, церебропротекции и кардиопротекции». Семинары по тромбозам и гемостазу . 31 (2): 234–246. дои : 10.1055/s-2005-869528 . ПМИД 15852226 . S2CID 41764516 .
- ^ Дюбуа С., Панико-Дюбуа Л., Меррилл-Сколофф Г., Фьюри Б., Фьюри BC (май 2006 г.). «Гликопротеин VI-зависимые и -независимые пути тромбообразования in vivo» . Кровь . 107 (10): 3902–3906. дои : 10.1182/кровь-2005-09-3687 . ПМК 1895285 . ПМИД 16455953 .
- ^ Цзян, Л.; Сюй, К.; Ю, С.; Лю, П.; Луо, Д.; Чжоу, К.; Гао, К.; Ху, Х. (2013). «Критическая роль тромбина/PAR-1 в АДФ-индуцированной секреции тромбоцитов и второй волне агрегации» . Журнал тромбозов и гемостаза . 11 (5): 930–940. дои : 10.1111/jth.12168 . ISSN 1538-7933 . ПМИД 23406164 .
- ^ Литвинов Р.И., Вайзель Й.В., Андрианова И.А., Пешкова А.Д., Минь Г.Л. (2018). «Дифференциальная чувствительность различных маркеров активации тромбоцитов аденозиндифосфатом» . Бионанонаука . 9 (1): 53–58. дои : 10.1007/s12668-018-0586-4 . ПМК 6750022 . ПМИД 31534882 .
- ^ Матарресе П., Страфасе Э., Палумбо Г., Ансельми М., Гамбарделла Л., Асционе Б., Дель Принсипе Д., Малорни В. (февраль 2009 г.). «Митохондрии регулируют метаморфоз тромбоцитов, индуцированный опсонизированным зимозаном А – активация и долгосрочная приверженность клеточной гибели» . Журнал ФЭБС . 276 (3): 845–856. дои : 10.1111/j.1742-4658.2008.06829.x . ПМИД 19143843 .
- ^ Уайт JG (декабрь 1987 г.). «Обзор структурной физиологии тромбоцитов». Сканирующая микросхема . 1 (4): 1677–1700. ПМИД 3324323 .
- ^ Бенке О (1970). «Морфология мембранных систем тромбоцитов». Серия Гематологическая . 3 (4): 3–16. ПМИД 4107203 .
- ^ Jump up to: а б Бушар Б.А., Манн К.Г., Бутенас С. (август 2010 г.). «Нет доказательств влияния тканевого фактора на тромбоциты» . Кровь . 116 (5): 854–855. doi : 10.1182/blood-2010-05-285627 . ПМЦ 2918337 . ПМИД 20688968 .
- ^ Ахмад С.С., Равала-Шейх Р., Уолш П.Н. (1992). «Компоненты и сборка комплекса, активирующего фактор Х». Семинары по тромбозам и гемостазу . 18 (3): 311–323. дои : 10.1055/s-2007-1002570 . ПМИД 1455249 . S2CID 28765989 .
- ^ Тьяги Т, Ахмад С, Гупта Н, Саху А, Ахмад Й, Наир В, Чаттерджи Т, Баджадж Н, Сенгупта С, Ганджу Л, Сингх СБ, Ашраф МЗ (февраль 2014 г.). «Измененная экспрессия белков тромбоцитов и активность кальпаина опосредуют протромботический фенотип, индуцированный гипоксией» . Кровь . 123 (8): 1250–1260. doi : 10.1182/blood-2013-05-501924 . ПМИД 24297866 .
- ^ О'Халлоран А.М., Кертин Р., О'Коннор Ф., Дули М., Фицджеральд А., О'Брайен Дж.К., Фицджеральд DJ, Шилдс, округ Колумбия (февраль 2006 г.). «Влияние генетической изменчивости в области гена GPIIIa на систематическую ошибку экспрессии Pl и плотность рецепторов GPIIb/IIIa в тромбоцитах». Британский журнал гематологии . 132 (4): 494–502. дои : 10.1111/j.1365-2141.2005.05897.x . ПМИД 16412022 . S2CID 41983626 .
- ^ Коллер Б.С., Череш Д.А., Аш Э., Селигсон У (январь 1991 г.). «Экспрессия рецептора витронектина тромбоцитов отличает иракских евреев от арабских пациентов с тромбастенией Гланцмана в Израиле» . Кровь . 77 (1): 75–83. дои : 10.1182/blood.V77.1.75.75 . ПМИД 1702031 .
- ^ Jump up to: а б с Дженне К.Н., Уррутиа Р., Кубес П. (июнь 2013 г.). «Тромбоциты: мостик гемостаза, воспаления и иммунитета» . Международный журнал лабораторной гематологии . 35 (3): 254–261. дои : 10.1111/ijlh.12084 . ПМИД 23590652 .
- ^ Левин Дж. (2007), «Эволюция тромбоцитов млекопитающих», Тромбоциты , Elsevier, стр. 3–22, doi : 10.1016/B978-012369367-9/50763-1 , ISBN 978-0123693679
- ^ Кокс Д., Керриган С.В., Уотсон С.П. (июнь 2011 г.). «Тромбоциты и врожденная иммунная система: механизмы бактериальной активации тромбоцитов» . Журнал тромбозов и гемостаза . 9 (6): 1097–1107. дои : 10.1111/j.1538-7836.2011.04264.x . ПМИД 21435167 .
- ^ Вейрих А.С., Циммерман Г.А. (сентябрь 2004 г.). «Тромбоциты: сигнальные клетки в иммунном континууме». Тенденции в иммунологии . 25 (9): 489–495. дои : 10.1016/j.it.2004.07.003 . ПМИД 15324742 .
- ^ Вагнер Д.Д., Burger PC (декабрь 2003 г.). «Тромбоциты при воспалении и тромбозе» . Атеросклероз, тромбоз и сосудистая биология . 23 (12): 2131–2137. doi : 10.1161/01.ATV.0000095974.95122.EC . ПМИД 14500287 .
- ^ Дьяково Т.Г., Пури К.Д., Варнок Р.А., Спрингер Т.А., фон Андриан У.Х. (июль 1996 г.). «Тромбоцит-опосредованная доставка лимфоцитов в венулы с высоким эндотелием». Наука . 273 (5272): 252–255. Бибкод : 1996Sci...273..252D . дои : 10.1126/science.273.5272.252 . ПМИД 8662511 . S2CID 21334521 .
- ^ Ианнаконе М., Ситиа Г., Исогава М., Маркезе П., Кастро М.Г., Ловенштейн П.Р., Чисари Ф.В., Руджери З.М., Гуидотти Л.Г. (ноябрь 2005 г.). «Тромбоциты опосредуют цитотоксическое повреждение печени, вызванное Т-лимфоцитами» . Природная медицина . 11 (11): 1167–1169. дои : 10.1038/nm1317 . ПМК 2908083 . ПМИД 16258538 .
- ^ Олерс, Стефан Х.; Тобин, Дэвид М.; Бриттон, Уорвик Дж.; Шавит, Джордан А.; Нгуен, Туонг; Йохансен, Мэтт Д.; Джонсон, Хелси Э.; Хортл, Элинор (2019). «Ингибирование тромбоцитов восстанавливает защитный иммунитет к микобактериальной инфекции у рыбок данио» . Журнал инфекционных болезней . 220 (3): 524–534. дои : 10.1093/infdis/jiz110 . ПМК 6603966 . ПМИД 30877311 .
- ^ Гертнер Ф, Ахмад З, Розенбергер Г, Фан С, Николай Л, Буш Б, Явуз Г, Люкнер М, Исикава-Анкерхольд Х, Хеннел Р, Бенечет А, Лоренц М, Чандраратне С, Шуберт И, Хелмер С, Стридниг Б, Старк К., Янко М., Бётчер Р.Т., Вершор А., Леон К., Гаше К., Гудерманн Т., Медерос Ю., Шницлер М., Пинкус З., Яннаконе М., Хаас Р., Ваннер Г., Лаубер К., Сикст М., Массберг С. (ноябрь 2017 г.) ). «Мигрирующие тромбоциты — это механопоглотители, которые собирают и объединяют бактерии» . Клетка . 171 (6): 1368–1382.e23. дои : 10.1016/j.cell.2017.11.001 . ПМИД 29195076 .
- ^ Бойлард Э., Нигрович П.А., Лараби К., Уоттс Г.Ф., Коблин Дж.С., Вайнблатт М.Е., Массаротти Э.М., Ремолд-О'Доннелл Э., Фарндейл Р.В., Уэр Дж., Ли Д.М. (январь 2010 г.). «Тромбоциты усиливают воспаление при артрите за счет выработки коллаген-зависимых микрочастиц» . Наука . 327 (5965): 580–583. Бибкод : 2010Sci...327..580B . дои : 10.1126/science.1181928 . ПМЦ 2927861 . ПМИД 20110505 .
- ^ Jump up to: а б Паланкар Р., Колер Т.П., Крауэль К., Веше Дж., Хаммершмидт С., Грейнахер А. (июнь 2018 г.). «Тромбоциты убивают бактерии, соединяя врожденный и адаптивный иммунитет посредством фактора тромбоцитов 4 и FcγRIIA» . Журнал тромбозов и гемостаза . 16 (6): 1187–1197. дои : 10.1111/jth.13955 . ПМИД 29350833 .
- ^ МакМорран Б.Дж., Вечорски Л., Дрисдейл К.Е., Чан Дж.А., Хуанг Х.М., Смит С., Митику С., Бисон Дж.Г., Бурджио Дж., Фут С.Дж. (декабрь 2012 г.). «Тромбоцитный фактор 4 и антиген Даффи, необходимые для уничтожения тромбоцитов Plasmodium falciparum» . Наука . 338 (6112): 1348–1351. Бибкод : 2012Sci...338.1348M . дои : 10.1126/science.1228892 . ПМИД 23224555 . S2CID 206544569 .
- ^ Муракава М., Окамура Т., Цуцуми К., Таногути С., Камура Т., Сибуя Т., Харада М., Нихо Ю. (1992). «Приобретенная болезнь фон Виллебранда в сочетании с эссенциальной тромбоцитемией: регресс после лечения». Акта гематологическая . 87 (1–2): 83–87. дои : 10.1159/000204725 . ПМИД 1585777 .
- ^ ван Гендерен П.Дж., Линкнегт Х., Михилс Дж.Дж., Бадде У. (сентябрь 1996 г.). «Приобретенная болезнь Виллебранда при миелопролиферативных заболеваниях». Лейкемия и лимфома . 22 (Приложение 1): 79–82. дои : 10.3109/10428199609074364 . ПМИД 8951776 .
- ^ Jump up to: а б Жесткий, Патрик Дж. (1990). Уокер, Х. Кеннет; Холл, В. Даллас; Херст, Дж. Уиллис (ред.). Клинические методы: история, физические и лабораторные исследования (3-е изд.). Бостон: Баттервортс. ISBN 978-0-409-90077-4 . ПМИД 21250105 .
- ^ Jump up to: а б «Нарушения тромбоцитов: тромбоцитопения» . Национальный институт сердца, легких и крови (NHLBI). 24 марта 2022 г. Проверено 18 ноября 2022 г.
- ^ Линд, Стюарт Э.; Куркджян, Карла Д. (2011). «Время кровотечения» . В Майкельсоне, Алан Д. (ред.). Тромбоциты (2-е изд.). Эльзевир. п. 485. ИСБН 978-0080465869 .
- ^ Герцог В.В. (1910). «Отношение тромбоцитов к геморрагической болезни» . ДЖАМА . 55 (14): 1185–1192. дои : 10.1001/jama.1910.04330140029009 .
- ^ Рануччи, Марко; Симиони, Паоло (2016). Тесты на месте тяжелого кровотечения: Руководство по диагностике и лечению . Спрингер. стр. 40–42. ISBN 978-3-319-24795-3 .
- ^ Маркуччи, Карло; Шетткер, Патрик (2014). Периоперационный гемостаз: коагуляция для анестезиологов . Спрингер. стр. 54–56. ISBN 978-3-642-55004-1 .
- ^ Jump up to: а б с д и Борхани, Мунира; Пахоре, Заен; ул Кадр, Зишан; Рехан, Мухаммед; Наз, Арши; Хан, Асиф; Ансари, Сакиб; Фарзана, Тасним; Надим, Мухаммед; Раза, Сайед Амир; Шамси, Тахир (2010). «Кровокровные расстройства в племени: результат кровнородственного размножения» . Сиротский журнал редких заболеваний . 5 (1). дои : 10.1186/1750-1172-5-23 . ISSN 1750-1172 . ПМИД 20822539 .
- ^ Jump up to: а б «Зачем выполнять агрегацию тромбоцитов?» . Хелена Биологические науки . 2015 год
- ^ Кукер, Адам (2014). «Агрегометрия светопропускания». Гематолог . 11 (2). дои : 10.1182/hem.V11.2.2555 . ISSN 1551-8779 .
- ^ МакГлассон Д.Л., Фрицма Г.А. Агрегометрия тромбоцитов цельной крови и исследование функции тромбоцитов. Семин Тромб Гемост. Март 2009 г.;35(2):168-80. дои: 10.1055/s-0029-1220325. Epub, 30 апреля 2009 г. PMID 19408190.
- ^ «Часто задаваемые вопросы по анализу функции тромбоцитов» (PDF) . Отделение патологии . Университет Содружества Вирджинии . Проверено 27 марта 2017 г.
- ^ Уоррен, Джей Ти; Ди Паола, Дж (2 июня 2022 г.). «Генетика наследственных тромбоцитопений» . Кровь . 139 (22): 3264–3277. дои : 10.1182/blood.2020009300 . ПМЦ 9164741 . ПМИД 35167650 .
- ^ Печчи, А; Балдуини, CL (июль 2021 г.). «Наследственные тромбоцитопении: обновленное руководство для врачей». Обзоры крови . 48 : 100784. doi : 10.1016/j.blre.2020.100784 . ПМИД 33317862 . S2CID 229178137 .
- ^ Корнеруп К.Н., Пейдж CP (август 2007 г.). «Роль тромбоцитов в патофизиологии астмы». Тромбоциты . 18 (5): 319–328. дои : 10.1080/09537100701230436 . ПМИД 17654302 . S2CID 7923694 .
- ^ Лэйдлоу Т.М., Киддер М.С., Бхаттачарья Н., Син В., Шен С., Милн Г.Л., Кастельс М.С., Чхай Х., Бойс Дж.А. (апрель 2012 г.). «Перепроизводство цистеиниллейкотриена при обострении респираторного заболевания, вызванного приемом аспирина, обусловлено лейкоцитами, прилипшими к тромбоцитам» . Кровь . 119 (16): 3790–3798. дои : 10.1182/blood-2011-10-384826 . ПМЦ 3335383 . ПМИД 22262771 .
- ^ Эрпенбек Л., член парламента Шёна (апрель 2010 г.). «Смертельные союзники: фатальное взаимодействие между тромбоцитами и метастазирующими раковыми клетками» . Кровь . 115 (17): 3427–3436. дои : 10.1182/кровь-2009-10-247296 . ПМЦ 2867258 . ПМИД 20194899 .
- ^ Пожалуйста, RJ (июль 2009 г.). «Сила тромбоцитов: липкие проблемы для липких паразитов?» . Тенденции в паразитологии . 25 (7): 296–299. дои : 10.1016/j.pt.2009.04.002 . ПМК 3116138 . ПМИД 19539528 .
- ^ «Краткая информация для пациентов. Функция тромбоцитов после приема ибупрофена в течение 1 недели» . Анналы внутренней медицины . 142 (7): I–54. Апрель 2005 г. doi : 10.7326/0003-4819-142-7-200504050-00004 . ПМИД 15809457 .
- ^ Рао Г.Х., Джонсон Г.Г., Редди К.Р., Уайт Дж.Г. (1983). «Ибупрофен защищает циклооксигеназу тромбоцитов от необратимого ингибирования аспирином» . Атеросклероз . 3 (4): 383–388. дои : 10.1161/01.ATV.3.4.383 . ПМИД 6411052 . S2CID 3229482 .
- ^ Ван Вин Дж., Нокс Т. Дж., Макрис М. (январь 2010 г.). «Риск спинальной гематомы после нейроаксиальной анестезии или люмбальной пункции у лиц с тромбоцитопенией» . Британский журнал гематологии . 148 (1): 15–25. дои : 10.1111/j.1365-2141.2009.07899.x . ПМИД 19775301 .
- ^ Робак Дж., Гроссман Б., Харрис Т., Хиллер С., ред. (2011). Техническое руководство (17-е изд.). Bethesda MD: AABB. п. 580. ИСБН 978-1-56395-315-6 .
- ^ Прован Д., Арнольд Д.М., Бассел Дж.Б., Чонг Б.Х., Купер Н., Гернсхаймер Т., Ганима В., Годо Б., Гонсалес-Лопес Т.Дж., Грейнджер Дж., Хоу М., Круз С., Макдональд В., Мишель М., Ньюленд AC, Паворд С. , Родегьеро Ф., Скалли М., Томияма Ю., Вонг Р.С., Заджа Ф., Кутер DJ. Обновленный международный консенсусный отчет по исследованию и лечению первичной иммунной тромбоцитопении. Кровь Адв. 26 ноября 2019 г.; 3 (22): 3780-3817. doi: 10.1182/bloodadvances.2019000812. PMID: 31770441; PMCID: PMC6880896.
- ^ Американская ассоциация банков крови (2003). «5.1.5.1». Стандарты для банков крови и служб переливания крови (22-е изд.). Bethesda MD: AABB.
- ^ Хёгман CF (январь 1992 г.). «Новые тенденции в получении и хранении тромбоцитов» . Переливание . 32 (1): 3–6. дои : 10.1046/j.1537-2995.1992.32192116428.x . ПМИД 1731433 .
- ^ Руан П.Х., Эдрич Р., Гамп Д., Кейл С.Д., Леонард Р.Л., Гудрич Р.П. (июнь 2004 г.). «Фотохимическая инактивация отдельных вирусов и бактерий в концентратах тромбоцитов с использованием рибофлавина и света». Переливание . 44 (6): 877–885. дои : 10.1111/j.1537-2995.2004.03355.x . ПМИД 15157255 . S2CID 24109912 .
- ^ Перес-Пухоль С., Тонда Р., Лозано М., Фусте Б., Лопес-Вильчес И., Галан А.М., Ли Дж., Гудрич Р., Эсколар Г. (июнь 2005 г.). «Влияние новой технологии снижения патогенов (Mirasol PRT) на функциональные аспекты концентратов тромбоцитов». Переливание . 45 (6): 911–919. дои : 10.1111/j.1537-2995.2005.04350.x . ПМИД 15934989 . S2CID 23169569 .
- ^ Резюме Проуза (апрель 2013 г.). «Инактивация патогенов компонентов: критический обзор». Вокс Сангвинис . 104 (3): 183–199. дои : 10.1111/j.1423-0410.2012.01662.x . ПМИД 23134556 . S2CID 38392712 .
- ^ ААББ (2009). Стандарты для банков крови и служб переливания крови (26-е изд.). Bethesda MD: AABB.
- ^ Шенфельд Х., Шпис С., Якоб С. (март 2006 г.). «Концентраты тромбоцитов с уменьшенным объемом». Текущие гематологические отчеты . 5 (1): 82–88. ПМИД 16537051 .
- ^ CBBS: Промытые и уменьшенные по объему блоки тромбоцитарфереза. Архивировано 14 апреля 2014 г. в Wayback Machine . Cbbsweb.org (25 октября 2001 г.). Проверено 14 ноября 2011 г.
- ^ Нгуен, Д.Т., Оргилл Д.П., Мерфи Г.Ф. (2009). Глава 4: Патофизиологические основы заживления ран и регенерации кожи. Биоматериалы для лечения потери кожи. Woodhead Publishing (Великобритания/Европа) и CRC Press (США), Кембридж/Бока-Ратон, стр. 25–57. ( ISBN 978-1-4200-9989-8 ISBN 978-1-84569-363-3 )
- ^ Моват Х.З., Вайзер В.Дж., Глинн М.Ф., Мастард Дж.Ф. (декабрь 1965 г.). «Фагоцитоз и агрегация тромбоцитов» . Журнал клеточной биологии . 27 (3): 531–543. дои : 10.1083/jcb.27.3.531 . ПМК 2106759 . ПМИД 4957257 .
- ^ Гаваз М., Фогель С. (октябрь 2013 г.). «Тромбоциты в восстановлении тканей: контроль апоптоза и взаимодействие с регенеративными клетками» . Кровь . 122 (15): 2550–2554. дои : 10.1182/blood-2013-05-468694 . ПМИД 23963043 .
- ^ Шмайер А.А., Сталкер Т.Дж., Рунге Дж.Дж., Ли Д., Нагасвами С., Мерико П., Чен М., Клише С., Гариепи К., Брасс Л.Ф., Хаммер Д.А., Вайзель Дж.В., Розенталь К., Кан М.Л. (сентябрь 2011 г.). «Оклюзивные тромбы возникают у млекопитающих, но не у птиц, в ответ на повреждение артерий: эволюционный взгляд на сердечно-сосудистые заболевания человека» . Кровь . 118 (13): 3661–3669. дои : 10.1182/blood-2011-02-338244 . ПМЦ 3186337 . ПМИД 21816834 .
- ^ Беламарич Ф.А., Шепро Д., Кин М. (ноябрь 1968 г.). «АДФ не участвует в индуцированной тромбином агрегации тромбоцитов позвоночных, не являющихся млекопитающими». Природа . 220 (5166): 509–510. Бибкод : 1968Natur.220..509B . дои : 10.1038/220509a0 . ПМИД 5686175 . S2CID 4269208 .
- ^ Ланцет, 1882, ii. 916; Записки исследований Гулливера по анатомии, физиологии, патологии и ботанике, 1880 г.; Физиология Карпентера, изд. Power, 9-е изд., см. Указатель в разделе «Гулливер».
- ^ Годли, сэр Рикман (1917). Лорд Листер . Лондон: Macmillan & Co.
- ^ Робб-Смит АХ (июль 1967 г.). «Почему были обнаружены тромбоциты». Британский журнал гематологии . 13 (4): 618–637. дои : 10.1111/j.1365-2141.1967.tb00769.x . ПМИД 6029960 . S2CID 5742616 .
- ^ Бил Л.С. (1864 г.). «О зародышевом веществе крови с замечаниями по образованию фибрина». Труды Микроскопического общества и журнала . 12 : 47–63. дои : 10.1111/j.1365-2818.1864.tb01625.x .
- ^ Шульце М (1865). «Нагретый столик и его применение при исследованиях крови» . Арх Микроск Анат . 1 (1): 1–42. дои : 10.1007/BF02961404 . S2CID 84919090 .
- ^ Биззозеро, Дж. (1882). «О новом компоненте крови и его роли в тромбообразовании и свертывании крови» . Арх Патол Анат Физ Клин Мед . 90 (2): 261–332. дои : 10.1007/BF01931360 . S2CID 37267098 .
- ^ Брюэр Д.Б. (май 2006 г.). «Макс Шульце (1865), Г. Биццозеро (1882) и открытие тромбоцитов» . Британский журнал гематологии . 133 (3): 251–258. дои : 10.1111/j.1365-2141.2006.06036.x . ПМИД 16643426 .
- ^ Научный американец . Манн и компания. 1882. с. 105.
- ^ Ослер В. (1886 г.). «О некоторых проблемах физиологии форменных элементов крови». Медицинские новости . 48 : 421–425.
- ^ Райт Дж. Х. (1906). «Происхождение и природа кровяных пластинок» . Бостонский медицинский и хирургический журнал . 154 (23): 643–645. дои : 10.1056/NEJM190606071542301 .
- ^ Райт Дж. Х. (1910). «Гистогенез тромбоцитов» . Журнал морфологии . 21 (2): 263–278. дои : 10.1002/jmor.1050210204 . hdl : 2027/hvd.32044107223588 . S2CID 84877594 .