Зеркальный нейрон
Эту статью необходимо обновить . ( апрель 2020 г. ) |
| Зеркальная система | |
|---|---|
| Идентификаторы | |
| МеШ | D059167 |
| Анатомические термины нейроанатомии | |
Зеркальный нейрон — это нейрон , который срабатывает как тогда, когда животное действует, так и когда животное наблюдает то же действие, выполняемое другим. [1] [2] [3] Таким образом, нейрон «отражает» поведение другого, как если бы действовал сам наблюдатель. Зеркальные нейроны не всегда физиологически отличаются от других типов нейронов мозга; их основным отличительным фактором является характер реагирования. [4] По этому определению такие нейроны наблюдались непосредственно у людей. [5] и виды приматов , [6] и у птиц. [7]
У людей активность мозга, соответствующая активности зеркальных нейронов, обнаружена в премоторной коре , дополнительной двигательной области , первичной соматосенсорной коре и нижней теменной коре . [8] Функция зеркальной системы у человека является предметом многочисленных спекуляций. Было показано, что птицы обладают имитирующим резонансным поведением, а неврологические данные свидетельствуют о наличии некоторой формы зеркальной системы. [6] [9] На сегодняшний день не предложено ни одной широко распространенной нейронной или вычислительной модели, описывающей, как активность зеркальных нейронов поддерживает когнитивные функции. [10] [11] [12]
Тема зеркальных нейронов продолжает вызывать интенсивные споры. В 2014 году журнал Philosophical Transactions of the Royal Society B опубликовал специальный выпуск, полностью посвященный исследованиям зеркальных нейронов. [13] Некоторые исследователи предполагают, что зеркальные системы могут имитировать наблюдаемые действия и, таким образом, способствовать теории умственных способностей. [14] [15] в то время как другие связывают зеркальные нейроны с языковыми способностями. [16] Нейробиологи, такие как Марко Якобони, утверждают, что системы зеркальных нейронов в человеческом мозге помогают людям понимать действия и намерения других людей. Кроме того, Якобони утверждал, что зеркальные нейроны являются нейронной основой способности человека к эмоциям, таким как сочувствие . [17]
Открытие
[ редактировать ]В 1980-х и 1990-х годах нейрофизиологи Джакомо Риццолатти , Джузеппе Ди Пеллегрино, Лучано Фадига , Леонардо Фогасси и Витторио Галлезе из Пармского университета поместили электроды в вентральную премоторную кору макак, чтобы изучить нейроны, специализирующиеся на контроле рук и рта. действия; например, взять объект и манипулировать им. В ходе каждого эксперимента исследователи позволяли обезьяне дотягиваться до кусочков еды и записывали данные по отдельным нейронам в мозгу обезьяны, измеряя таким образом реакцию нейрона на определенные движения. [18] [19] Они обнаружили, что некоторые нейроны реагировали, когда обезьяна наблюдала, как человек берет кусок еды, а также когда обезьяна сама брала еду.Открытие было первоначально представлено журналу Nature , но было отклонено из-за «отсутствия общего интереса» перед публикацией в менее конкурентоспособном журнале. [20]
Несколько лет спустя та же группа опубликовала еще одну эмпирическую статью, в которой обсуждалась роль системы зеркальных нейронов в распознавании действий и предполагалось, что зона Брока человека является гомологической областью вентральной премоторной коры обезьяны. [21] Хотя в этих статьях сообщалось о наличии зеркальных нейронов, реагирующих на действия рук, последующее исследование Пьера Франческо Феррари и его коллег [22] описали наличие зеркальных нейронов, реагирующих на движения рта и мимические жесты.
Дальнейшие эксперименты подтвердили, что около 10% нейронов нижней лобной и нижнетеменной коры обезьян обладают «зеркальными» свойствами и дают сходные реакции на выполняемые движения рук и наблюдаемые действия. В 2002 году Кристиан Кейзерс и его коллеги сообщили, что как у людей, так и у обезьян зеркальная система также реагирует на звуки действий. [3] [23] [24]
Отчеты о зеркальных нейронах были широко опубликованы. [21] и подтвердил [25] зеркальные нейроны обнаружены как в нижнелобных, так и нижнетеменных областях мозга. Недавно данные функциональной нейровизуализации убедительно свидетельствуют о том, что у людей есть схожие системы зеркальных нейронов: исследователи определили области мозга, которые реагируют как во время действия, так и во время наблюдения за действием. Неудивительно, что эти области мозга включают те, что есть у макак. [1] Однако функциональная магнитно-резонансная томография (фМРТ) может исследовать весь мозг сразу и предполагает, что у людей гораздо более широкая сеть областей мозга проявляет зеркальные свойства, чем считалось ранее. Эти дополнительные области включают соматосенсорную кору и, как полагают, заставляют наблюдателя почувствовать, как он чувствует себя при движении наблюдаемым образом. [26] [27]
Источник
[ редактировать ]Многие неявно предполагают, что зеркальная функция зеркальных нейронов обусловлена прежде всего наследственными генетическими факторами и что генетическая предрасположенность к развитию зеркальных нейронов развилась потому, что они облегчают понимание действий. [28] Напротив, ряд теоретических теорий утверждают, что зеркальные нейроны могут просто возникнуть из-за выученных ассоциаций, включая теорию Хеббиана . [29] теория ассоциативного обучения , [28] и канализация . [30]
У обезьян
[ редактировать ]
Первым животным, у которого исследователи изучали зеркальные нейроны индивидуально, стала макака . У этих обезьян зеркальные нейроны обнаруживаются в нижней лобной извилине (область F5) и нижней теменной доле . [1]
Считается, что зеркальные нейроны способствуют пониманию поведения других животных . Например, зеркальный нейрон, который срабатывает, когда обезьяна рвет лист бумаги, также срабатывает, когда обезьяна видит, как человек рвет бумагу, или слышит, как рвется бумага (без визуальных сигналов). Эти свойства заставили исследователей поверить в то, что зеркальные нейроны кодируют абстрактные концепции действий, таких как «разрыв бумаги», независимо от того, выполняется ли это действие обезьяной или другим животным. [1]
Функция зеркальных нейронов у макак остается неизвестной. Взрослые макаки, похоже, не учатся путем подражания. Недавние эксперименты Феррари и его коллег показывают, что детеныши макак могут имитировать движения лица человека, но только в новорожденном возрасте и в течение ограниченного временного окна. [31] Даже если это еще не было эмпирически продемонстрировано, было высказано предположение, что зеркальные нейроны вызывают такое поведение и другие имитационные явления. [32] Действительно, существует ограниченное понимание того, в какой степени обезьяны демонстрируют подражательное поведение. [10]
У взрослых обезьян зеркальные нейроны могут позволить обезьяне понимать, что делает другая обезьяна, или распознавать действия другой обезьяны. [33]
У грызунов
[ редактировать ]Ряд исследований показал, что крысы и мыши проявляют признаки дистресса, когда видят, как другого грызуна трясут ногами. [34] Группа Кристиана Кейзерса, записанная с нейронов, когда крысы испытывали боль или были свидетелями боли других, выявила наличие зеркальных нейронов боли в передней поясной извилине крысы, то есть нейронов, которые реагируют как в то время, когда животное испытывает боль, так и в то время, когда животное становится свидетелем боли. других. [35] Деактивация этой области поясной извилины привела к уменьшению эмоционального заражения у крыс, так что крысы-наблюдатели демонстрировали меньший стресс, когда были свидетелями того, как другая крыса испытывает боль. [35] Гомологичная часть передней поясной извилины связана с сочувствием к боли у людей. [36] что позволяет предположить гомологию между системами, участвующими в эмоциональном заражении у грызунов и эмпатией/эмоциональным заражением боли у людей.
У людей
[ редактировать ]
Обычно невозможно изучить отдельные нейроны в человеческом мозге, поэтому большинство доказательств существования зеркальных нейронов у людей являются косвенными. Эксперименты по визуализации мозга с использованием функциональной магнитно-резонансной томографии (фМРТ) показали, что нижняя лобная кора и верхняя теменная доля человека активны, когда человек выполняет действие, а также когда человек видит, как другой человек выполняет действие. Было высказано предположение, что эти области мозга содержат зеркальные нейроны, и они были определены как система зеркальных нейронов человека. [37] Более поздние эксперименты показали, что даже на уровне отдельных участников, сканированных с помощью фМРТ, большие области, содержащие несколько вокселей фМРТ, повышают их активность как во время наблюдения, так и при выполнении действий. [26]
Нейропсихологические исследования, изучающие области поражения, вызывающие нарушение навыков действий, интерпретации пантомимы и дефицит восприятия биологических движений , указали на причинную связь между целостностью нижней лобной извилины и этим поведением. [38] [39] [40] Исследования транскраниальной магнитной стимуляции также подтвердили это. [41] [42] Эти результаты показывают, что активация в областях, связанных с зеркальными нейронами, вряд ли будет просто эпифеноменальной.
В исследовании, опубликованном в апреле 2010 года, сообщается о записях отдельных нейронов с зеркальными свойствами в человеческом мозге. [43] Мукамель и др. (Current Biology, 2010) зарегистрировано в мозге 21 пациента, находившегося на лечении в Медицинском центре Рональда Рейгана Калифорнийского университета в Лос-Анджелесе от трудноизлечимой эпилепсии . Пациентам были имплантированы внутричерепные глубинные электроды для выявления очагов судорог для возможного хирургического лечения. Расположение электродов определялось исключительно клиническими критериями; исследователи, с согласия пациентов, использовали те же электроды для «совмещения» своих исследований. Исследователи обнаружили небольшое количество нейронов, которые активировались или проявляли наибольшую активность как тогда, когда человек выполнял задачу, так и когда он наблюдал за ней. Другие нейроны обладали антизеркальными свойствами: они реагировали, когда участник выполнял действие, но тормозились, когда участник видел это действие.
Обнаруженные зеркальные нейроны располагались в дополнительной двигательной области и медиальной височной коре (остальные области мозга не исследовались). По чисто практическим причинам эти области не совпадают с теми, в которых зеркальные нейроны были зарегистрированы у обезьян: исследователи в Парме изучали вентральную премоторную кору и связанную с ней нижнюю теменную долю, две области, в которых эпилепсия возникает редко, и следовательно, записи отдельных клеток в этих регионах у людей обычно не производятся. С другой стороны, до сих пор никому не приходилось искать зеркальные нейроны в дополнительной двигательной области или медиальной височной доле у обезьяны. Таким образом, в совокупности это не означает, что у людей и обезьян зеркальные нейроны находятся в разных местах, а, скорее, они могут иметь зеркальные нейроны как в вентральной премоторной коре, так и в нижней теменной доле, где они были зарегистрированы у обезьян, и в дополнительной доле. двигательные области и медиальную височную долю, где они были зарегистрированы у человека, особенно потому, что подробный анализ фМРТ человека предполагает активность, совместимую с наличием зеркальных нейронов во всех этих областях. [26]
Другое исследование показало, что у людей не обязательно больше зеркальных нейронов, чем у обезьян, но вместо этого существует основной набор зеркальных нейронов, используемый для наблюдения и выполнения действий. Однако для других предполагаемых функций зеркальных нейронов зеркальная система может обладать способностью задействовать другие области мозга при выполнении своих слуховых, соматосенсорных и аффективных компонентов. [44]
Разработка
[ редактировать ]Данные о младенцах, полученные с использованием методов отслеживания глаз, позволяют предположить, что система зеркальных нейронов развивается до 12-месячного возраста и что эта система может помогать младенцам понимать действия других людей. [45] Критический вопрос касается того, как зеркальные нейроны приобретают зеркальные свойства. Две тесно связанные модели предполагают, что зеркальные нейроны обучаются посредством Хеббиана. [46] или Ассоциативное обучение [47] [48] [12] (см. Ассоциативное последовательное обучение ). Однако если премоторные нейроны необходимо тренировать действием, чтобы приобрести зеркальные свойства, то неясно, как новорожденные дети способны имитировать мимику другого человека (имитация невидимых действий), как это предполагает работа Мельцова и Мура. . Одна из возможностей заключается в том, что вид высовывания языка задействует врожденный механизм высвобождения у новорожденных. Тщательный анализ показывает, что «имитация» этого единственного жеста может объяснить почти все сообщения о мимике лица у новорожденных. [49]
Возможные функции
[ редактировать ]Понимание намерений
[ редактировать ]Многие исследования связывают зеркальные нейроны с пониманием целей и намерений. Фогасси и др. (2005) [25] зарегистрировали активность 41 зеркального нейрона в нижней теменной доле (IPL) двух макак-резус. IPL уже давно признана ассоциативной корой головного мозга, которая объединяет сенсорную информацию. Обезьяны наблюдали, как экспериментатор либо хватал яблоко и подносил его ко рту, либо хватал какой-то предмет и помещал его в чашку.
- В общей сложности 15 зеркальных нейронов энергично активизировались, когда обезьяна наблюдала движение «схватить-есть», но не зарегистрировали никакой активности в условиях «схватывания-места».
- Для четырех других зеркальных нейронов справедливо обратное: они активировались в ответ на то, что экспериментатор в конце концов положил яблоко в чашку, но не съел его.
Только тип действия, а не кинематическая сила, с которой модели манипулировали объектами, определял активность нейронов. Также было важно, что нейроны сработали до того, как обезьяна увидела, как человеческая модель начинает второй двигательный акт (поднесение предмета ко рту или помещение его в чашку). Следовательно, нейроны IPL «кодируют одно и то же действие (схватывание) по-разному в зависимости от конечной цели действия, в которое заложено это действие». [25] Они могут обеспечить нейронную основу для прогнозирования последующих действий другого человека и определения намерений. [25]
Содействие обучению
[ редактировать ]Другой возможной функцией зеркальных нейронов может быть облегчение обучения. Зеркальные нейроны кодируют конкретное представление действия, то есть представление, которое активировалось бы, если бы наблюдатель действовал. Это позволило бы нам неявно (в мозгу) смоделировать (повторить внутренне) наблюдаемое действие, собрать собственные двигательные программы наблюдаемых действий и подготовиться к воспроизведению действий позже. Это неявное обучение. Благодаря этому наблюдатель будет производить действие явно (в своем поведении) с ловкостью и ловкостью. Это происходит за счет ассоциативных процессов обучения. Чем чаще активируется синаптическая связь, тем сильнее она становится. [50]
Сочувствие
[ редактировать ]Стефани Престон и Франс де Ваал [51] Жан Десети , [52] [53] и Витторио Галлезе [54] [55] и Кристиан Кейзерс [3] независимо утверждали, что система зеркальных нейронов участвует в эмпатии . Большое количество экспериментов с использованием фМРТ, электроэнцефалографии (ЭЭГ) и магнитоэнцефалографии (МЭГ) показали, что определенные области мозга (в частности, передняя островковая доля , передняя поясная извилина и нижняя лобная кора) активны, когда люди испытывают эмоции (отвращение, счастье). , боль и т. д.), а также когда они видят другого человека, испытывающего эмоцию. [56] [57] [58] [59] [60] [61] [62] Дэвид Фридберг и Витторио Галлезе также выдвинули идею о том, что эта функция системы зеркальных нейронов имеет решающее значение для эстетического опыта. [63] Тем не менее эксперимент, направленный на исследование активности зеркальных нейронов при эмпатии, проведенный Сукайной Беккали и Питером Энтикоттом в Университете Дикина, дал иной результат. Проанализировав данные отчета, они пришли к двум выводам о моторной эмпатии и эмоциональной эмпатии. Во-первых, нет никакой связи между моторной эмпатией и активностью зеркальных нейронов. Во-вторых, имеются лишь слабые доказательства активности этих нейронов в нижней лобной извилине (НФГ) и нет доказательств эмоциональной эмпатии, связанной с зеркальными нейронами в ключевых областях мозга (нижняя теменная долька: IPL). Другими словами, не было сделано точного вывода о роли зеркальных нейронов в эмпатии и о том, необходимы ли они для человеческой эмпатии. [64] Однако эти области мозга не совсем такие же, как те, которые отражают действия рук, а зеркальные нейроны, отвечающие за эмоциональные состояния или сочувствие, у обезьян еще не описаны.
В недавнем исследовании, проведенном в 2022 году, для каждого задания требовалось шестнадцать действий руками. Задание отображало как фазу слова активности, так и фазу предполагаемого слова. Действия рук были выбраны в «следах», каждый из которых вводился дважды. Один раз был с фазой соответствия, а другой раз с фазой вводящего в заблуждение слова. Слова действия изображались двумя-тремя словами, каждое из которых начиналось со слова «к». Например, «указать» (действие) или «вращать» (намерение).
Участники должны были ответить, соответствует ли правильная фаза слова соответствующему слову действия или намерения. На фазу слова нужно было ответить в течение 3000 мс, с черным экраном между каждым изображением 1000 мс. Цель создания черных экранов заключалась в том, чтобы обеспечить достаточное количество времени между ответами. Участники нажимали на клавиатуре «x» или «m», чтобы обозначить свои ответы в формате «да/нет». [65]
Кристиан Кейзерс из Social Brain Lab и его коллеги показали, что люди, которые, согласно анкетам самоотчета, более эмпатичны, имеют более сильную активацию как в зеркальной системе, так и в отношении действий рук. [66] и зеркальная система для эмоций, [61] обеспечивая более прямую поддержку идеи о том, что зеркальная система связана с эмпатией.Некоторые исследователи заметили, что система зеркал человека не пассивно реагирует на наблюдение за действиями, а находится под влиянием мышления наблюдателя. [67] Исследователи наблюдали связь зеркальных нейронов во время эмпатического участия в уходе за пациентами. [68]
Исследования на крысах показали, что передняя поясная извилина содержит зеркальные нейроны боли, то есть нейроны, реагирующие как во время непосредственного переживания боли, так и во время наблюдения за болью других. [35] ингибирование этой области приводит к снижению эмоционального заражения у крыс. [35] и мыши, [34] и снижение отвращения к причинению вреда другим. [69] Это дает причинно-следственную связь между зеркальными нейронами боли, эмоциональным заражением и просоциальным поведением — двумя явлениями, связанными с эмпатией, у грызунов. Эта мозговая активность в гомологичной области мозга связана с индивидуальной вариабельностью эмпатии у людей. [36] предполагает, что аналогичный механизм может действовать и у млекопитающих.
Самосознание человека
[ редактировать ]В.С. Рамачандран предположил, что зеркальные нейроны могут обеспечить неврологическую основу самосознания человека. [70] В эссе, написанном для Edge Foundation в 2009 году, Рамачандран дал следующее объяснение своей теории: «...Я также предположил, что эти нейроны могут не только помогать моделировать поведение других людей, но и могут быть направлены «вовнутрь» — так сказать, — чтобы создавать представления второго порядка или мета-представления ваших собственных более ранних мозговых процессов. Это может быть нейронной основой самоанализа и взаимности самосознания и другого осознания. Очевидно, здесь возникает вопрос о курице или яйце. эволюционировали первыми, но... Главное, что эти двое эволюционировали совместно, взаимно обогащая друг друга, создавая зрелое представление о себе, которое характеризует современных людей». [71]
Язык
[ редактировать ]Функциональные МРТ-исследования у людей сообщили об обнаружении областей, гомологичных системе зеркальных нейронов обезьяны, в нижней лобной коре, близкой к зоне Брока, одной из предполагаемых речевых областей мозга. Это привело к предположениям, что человеческий язык развился из системы исполнения/понимания жестов, реализованной в зеркальных нейронах. Говорят, что зеркальные нейроны могут обеспечить механизм понимания действий, имитационного обучения и моделирования поведения других людей. [72] Эта гипотеза подтверждается некоторой цитоархитектонической гомологией между премоторной областью F5 обезьяны и областью Брока человека. [73] Скорость расширения словарного запаса связана со способностью детей вокально отражать не-слова и, таким образом, приобретать новое произношение слов. Такое повторение речи происходит автоматически, быстро [74] и отдельно в мозгу для восприятия речи . [75] [76] Более того, такая голосовая имитация может происходить без понимания, например, при затенении речи. [77] и эхолалия . [78]
Дополнительные доказательства этой связи получены в недавнем исследовании, в котором мозговая активность двух участников измерялась с помощью фМРТ, когда они жестикулировали друг другу, используя жесты рук во время игры в шарады – метод, который, по мнению некоторых, может представлять собой эволюционного предшественника. человеческого языка. Анализ данных с использованием причинности Грейнджера показал, что система зеркальных нейронов наблюдателя действительно отражает образец активности двигательной системы отправителя, что подтверждает идею о том, что двигательная концепция, связанная со словами, действительно передается от одного мозга к другому. с помощью зеркальной системы [79]
Система зеркальных нейронов, по-видимому, по своей сути неадекватна для того, чтобы играть какую-либо роль в синтаксисе , учитывая, что это определяющее свойство человеческих языков, реализованное в иерархической рекурсивной структуре, сводится к линейным последовательностям фонем, что делает рекурсивную структуру недоступной для сенсорного обнаружения. [80]
Автоматическая имитация
[ редактировать ]Этот термин обычно используется для обозначения случаев, когда человек, наблюдая за движением тела, непреднамеренно выполняет аналогичное движение тела или изменяет способ выполнения движения тела. Автоматическая имитация редко предполагает открытое выполнение совпадающих реакций. Вместо этого эффекты обычно состоят во времени реакции, а не в точности, различиях между совместимыми и несовместимыми исследованиями.Исследования показывают, что существование автоматической имитации, которая является скрытой формой имитации, отличается от пространственной совместимости. Это также указывает на то, что, хотя автоматическое подражание подлежит входной модуляции с помощью процессов внимания и выходной модуляции с помощью тормозных процессов, оно опосредовано приобретенными, долговременными сенсомоторными ассоциациями, которые не могут быть изменены непосредственно интенциональными процессами. Многие исследователи полагают, что автоматическое подражание осуществляется системой зеркальных нейронов. [81] Кроме того, есть данные, которые показывают, что наш постуральный контроль нарушается, когда люди слушают предложения о других действиях. Например, если задача состоит в том, чтобы сохранить осанку, люди делают это хуже, когда слушают такие предложения: «Я встаю, надеваю тапочки, иду в ванную». Это явление может быть связано с тем, что при восприятии действия происходит аналогичная активация моторной коры, как если бы то же действие выполнял человек (система зеркальных нейронов). [82]
Двигательная мимикрия
[ редактировать ]В отличие от автоматической имитации, моторная мимикрия наблюдается в (1) натуралистических социальных ситуациях и (2) посредством измерения частоты действий в течение сеанса, а не измерения скорости и/или точности в ходе испытаний. [83]
Объединение исследований моторной мимикрии и автоматического подражания могло бы выявить убедительные доказательства того, что эти явления зависят от одних и тех же психологических и нервных процессов. Однако предварительные данные получены в исследованиях, показывающих, что социальный прайминг оказывает аналогичное влияние на моторную мимику. [84] [85]
Тем не менее, сходство между автоматической имитацией, зеркальными эффектами и моторной мимикрией побудило некоторых исследователей предположить, что автоматическая имитация опосредована системой зеркальных нейронов и что это строго контролируемый лабораторный эквивалент моторной мимикрии, наблюдаемой в натуралистических социальных контекстах. Если это правда, то автоматическую имитацию можно использовать как инструмент для исследования того, как система зеркальных нейронов способствует когнитивному функционированию и как моторная мимика способствует просоциальным установкам и поведению. [86] [87]
Метаанализ исследований имитации на людях показывает, что существует достаточно доказательств активации зеркальной системы во время имитации, что вероятно вовлечение зеркальных нейронов, хотя ни в одном опубликованном исследовании не зафиксирована активность отдельных нейронов. Однако для двигательной имитации этого, вероятно, недостаточно. Исследования показывают, что при подражании в равной степени активируются области лобных и теменных долей, выходящие за пределы классической зеркальной системы. Это говорит о том, что другие области, наряду с зеркальной системой, имеют решающее значение для имитации поведения. [8]
Аутизм
[ редактировать ]Также было высказано предположение, что проблемы с системой зеркальных нейронов могут лежать в основе когнитивных расстройств, особенно аутизма . [88] [89] Однако связь между дисфункцией зеркальных нейронов и аутизмом является предварительной, и еще предстоит продемонстрировать, как зеркальные нейроны связаны со многими важными характеристиками аутизма. [10]
Некоторые исследователи утверждают, что существует связь между дефицитом зеркальных нейронов и аутизмом . Записи ЭЭГ двигательных областей подавляются, когда кто-то наблюдает за движением другого человека, и этот сигнал может относиться к системе зеркальных нейронов. Это подавление было меньше у детей с аутизмом. [88] Хотя эти результаты были повторены несколькими группами, [90] [91] другие исследования не обнаружили доказательств дисфункции системы зеркальных нейронов при аутизме. [10] В 2008 году Оберман и др. опубликовал исследовательскую работу, в которой были представлены противоречивые данные ЭЭГ. Оберман и Рамачандран обнаружили типичное мю-подавление для знакомых стимулов, но не для незнакомых стимулов, что привело их к выводу, что система зеркальных нейронов детей с РАС (расстройством аутистического спектра) функциональна, но менее чувствительна, чем у типичных детей. [92] Основываясь на противоречивых данных, представленных экспериментами по подавлению мю-волн, Патриция Черчленд предупредила, что результаты подавления мю-волн не могут использоваться в качестве достоверного показателя для измерения производительности систем зеркальных нейронов. [93] Недавние исследования показывают, что зеркальные нейроны не играют роли в аутизме:
...не существует четких доказательств фундаментального дефицита зеркальной системы при аутизме. Поведенческие исследования показали, что люди с аутизмом хорошо понимают цели действий. Более того, два независимых исследования нейровизуализации показали, что теменной компонент зеркальной системы обычно функционирует у людей с аутизмом. [94]
Некоторые анатомические различия были обнаружены в областях мозга, связанных с зеркальными нейронами, у взрослых с расстройствами аутистического спектра по сравнению со взрослыми, не страдающими аутизмом. Все эти области коры были тоньше, и степень истончения коррелировала с тяжестью симптомов аутизма, причем эта корреляция почти ограничивалась этими областями мозга. [95] Основываясь на этих результатах, некоторые исследователи утверждают, что аутизм вызван нарушениями в системе зеркальных нейронов, что приводит к нарушениям социальных навыков, подражания, эмпатии и теории разума. [ ВОЗ? ]
Многие исследователи отмечают, что теория аутизма «разбитых зеркал» слишком упрощена, и одни лишь зеркальные нейроны не могут объяснить различия, обнаруженные у людей с аутизмом. Прежде всего, как отмечалось выше, ни одно из этих исследований не было прямым измерением активности зеркальных нейронов - другими словами, активность фМРТ или подавление ритма ЭЭГ не однозначно индексируют зеркальные нейроны. Динштейн и его коллеги обнаружили нормальную активность зеркальных нейронов у людей с аутизмом с помощью фМРТ. [96] У людей с аутизмом не всегда обнаруживаются нарушения понимания намерения, понимания действий и восприятия биологического движения (ключевые функции зеркальных нейронов). [97] [98] или зависят от задачи. [99] [100] Сегодня очень немногие люди верят, что проблема зеркальной системы по принципу «все или ничего» может лежать в основе аутизма. Вместо этого «необходимо провести дополнительные исследования и проявлять больше осторожности при обращении к средствам массовой информации». [101]
Исследования 2010 года [96] пришли к выводу, что у аутистов нет дисфункции зеркальных нейронов, хотя небольшой размер выборки ограничивает степень обобщения этих результатов. В более позднем обзоре утверждается, что неврологических данных недостаточно, чтобы поддержать эту «теорию разбитого зеркала» об аутизме. [102]
Теория разума
[ редактировать ]В философии разума зеркальные нейроны стали основным объединяющим призывом теоретиков моделирования относительно нашей « теории разума ». «Теория разума» относится к нашей способности делать выводы о психическом состоянии другого человека (т. е. убеждениях и желаниях) на основе опыта или его поведения.
Существует несколько конкурирующих моделей, пытающихся объяснить нашу теорию разума; наиболее примечательной в отношении зеркальных нейронов является теория моделирования. Согласно теории симуляции, теория разума доступна, потому что мы подсознательно сопереживаем человеку, за которым наблюдаем, и, учитывая соответствующие различия, представляем, чего бы мы хотели и во что верили в этом сценарии. [103] [104] Зеркальные нейроны интерпретировались как механизм, с помощью которого мы моделируем других, чтобы лучше их понять, и поэтому их открытие было воспринято некоторыми как подтверждение теории моделирования (которая появилась за десять лет до открытия зеркальных нейронов). [54] Совсем недавно Теория разума и моделирование рассматривались как взаимодополняющие системы с разными временными путями развития. [105] [106] [107]
На уровне нейронов в исследовании 2015 года, проведенном Керен Харуш и Зивом Уильямсом с использованием совместно взаимодействующих приматов, выполняющих повторяющуюся игру «Дилемма заключенного», авторы выявили нейроны в передней поясной извилине , которые выборочно предсказывали еще неизвестные решения противника или скрытое состояние ума. Эти «нейроны, предсказывающие другие», различали собственные решения и решения других и были уникально чувствительны к социальному контексту, но они не кодировали наблюдаемые действия противника или получение вознаграждения. Таким образом, эти поясные клетки могут существенно дополнять функцию зеркальных нейронов, предоставляя дополнительную информацию о других социальных агентах, которую невозможно сразу наблюдать или знать. [108]
Половые различия
[ редактировать ]Серия недавних исследований, проведенных Явеем Ченгом с использованием различных нейрофизиологических показателей, включая МЭГ , [109] спинномозговая рефлекторная возбудимость, [110] электроэнцефалография, [111] [112] задокументировали наличие гендерных различий в системе зеркальных нейронов человека: участники-женщины демонстрировали более сильный двигательный резонанс, чем участники-мужчины.
В другом исследовании половые различия между механизмами зеркальных нейронов были подтверждены тем, что данные показали повышенную эмпатическую способность у женщин по сравнению с мужчинами. [ нужна ссылка ] . Во время эмоционального социального взаимодействия женщины показали большую способность к эмоциональной перспективе, принимая во внимание [ нужны разъяснения ] чем мужчины при общении с другим человеком лицом к лицу. Однако данные исследования показали, что когда дело дошло до распознавания эмоций других, способности всех участников были очень похожими, и между испытуемыми мужского и женского пола не было ключевой разницы. [113]
Сонный паралич
[ редактировать ]Баланд Джалал и В.С. Рамачандран выдвинули гипотезу, что система зеркальных нейронов играет важную роль в возникновении галлюцинаций незваного гостя и внетелесных переживаний во время сонного паралича . [114] Согласно этой теории, сонный паралич приводит к растормаживанию системы зеркальных нейронов, открывая путь галлюцинациям человекоподобных призрачных существ. В качестве механизма такого расторможения зеркальных нейронов предлагается деафферентация сенсорной информации при сонном параличе. [114] Авторы предполагают, что их гипотезу о роли системы зеркальных нейронов можно проверить:
«Эти идеи можно изучить с помощью нейровизуализации, чтобы изучить избирательную активацию областей мозга, связанных с активностью зеркальных нейронов, когда человек галлюцинирует незваного гостя или испытывает опыт выхода из тела во время сонного паралича». [114]
Функция зеркальных нейронов, психоз и эмпатия при шизофрении
[ редактировать ]Недавние исследования, в которых измерялось подавление мю-волн, показывают, что активность зеркальных нейронов положительно коррелирует с психотическими симптомами (т.е. большее подавление мю/активность зеркальных нейронов было самым высоким среди субъектов с большей тяжестью психотических симптомов). Исследователи пришли к выводу, что «более высокая активность зеркальных нейронов может лежать в основе дефицита сенсорных ворот при шизофрении и может способствовать сенсорным ошибочным атрибуциям, особенно в ответ на социально значимые стимулы, а также быть предполагаемым механизмом бреда и галлюцинаций». [115]
Сомнения относительно зеркальных нейронов
[ редактировать ]Хотя некоторые представители научного сообщества выразили восторг по поводу открытия зеркальных нейронов, есть учёные, которые выразили сомнения как по поводу существования, так и по поводу роли зеркальных нейронов в организме человека. Сегодня, похоже, все согласны с тем, что важность так называемых зеркальных нейронов сильно преувеличена. По мнению таких ученых, как Хикок, Пасколо и Динштейн, неясно, действительно ли зеркальные нейроны образуют отдельный класс клеток (в отличие от случайного явления, наблюдаемого в клетках, выполняющих другие функции). [116] и является ли зеркальная активность отдельным типом реакции или просто артефактом общего облегчения двигательной системы. [11]
В 2008 году Илан Динштейн и др. утверждал, что первоначальный анализ был неубедительным, поскольку он был основан на качественном описании свойств отдельных клеток и не принимал во внимание небольшое количество нейронов с сильной зеркальной избирательностью в двигательных областях. [10] Другие учёные утверждают, что измерения задержки срабатывания нейронов, похоже, несовместимы со стандартным временем реакции. [116] и отметил, что никто не сообщил о том, что прерывание двигательных областей в F5 приведет к снижению распознавания действий. [11] (Критики этого аргумента ответили, что эти авторы пропустили исследования нейропсихологии человека и ТМС, в которых сообщалось, что нарушения в этих областях действительно вызывают дефицит активности. [39] [41] не затрагивая другие виды восприятия.) [40]
В 2009 году Линнау и др. провели эксперимент, в котором они сравнили двигательные действия, которые сначала наблюдались, а затем выполнялись, с двигательными действиями, которые сначала выполнялись, а затем наблюдались. Они пришли к выводу, что между этими двумя процессами существует значительная асимметрия, которая указывает на то, что зеркальные нейроны не существуют у людей. Они заявили: «Важно отметить, что мы не обнаружили никаких признаков адаптации для двигательных действий, которые сначала выполнялись, а затем наблюдались. Неспособность найти кросс-модальную адаптацию для выполняемых и наблюдаемых двигательных действий несовместима с основным предположением.теории зеркальных нейронов, которая утверждает, что распознавание и понимание действий основаны на двигательном моделировании». [117] Однако в том же году Килнер и др. показали, что если в качестве стимулов используются целенаправленные действия, как IPL, так и премоторные области демонстрируют подавление повторения между наблюдением и выполнением, которое предсказывают зеркальные нейроны. [118]
В 2009 году Грег Хикок опубликовал обширный аргумент против утверждения о том, что зеркальные нейроны участвуют в понимании действий: «Восемь проблем теории понимания действий зеркальных нейронов у обезьян и людей». Он пришел к выводу, что «ранняя гипотеза о том, что эти клетки лежат в основе понимания действий, также является интересной и разумной на первый взгляд идеей. Однако, несмотря на ее широкое признание, это предложение никогда не было должным образом проверено на обезьянах, а на людях имеются убедительные эмпирические доказательства, форме физиологической и нейропсихологической (двойной) диссоциации, против иска». [11]
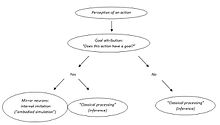
Владимир Косоногов видит еще одно противоречие. Сторонники теории понимания действий зеркальных нейронов постулируют, что зеркальные нейроны кодируют цели действий других людей, поскольку они активируются, если наблюдаемое действие целенаправленно. Однако зеркальные нейроны активируются только тогда, когда наблюдаемое действие является целенаправленным (предметное действие или коммуникативный жест, который, безусловно, тоже имеет цель). Откуда они «узнают», что определенное действие целенаправленно? На каком этапе своей активации они обнаруживают цель движения или ее отсутствие? По его мнению, система зеркальных нейронов может активироваться только после того, как цель наблюдаемого действия будет присвоена какими-то другими структурами мозга. [50]
Нейрофилософы, такие как Патрисия Черчленд, выразили как научные, так и философские возражения против теории, согласно которой зеркальные нейроны отвечают за понимание намерений других. В главе 5 своей книги Braintrust 2011 года Черчленд указывает, что утверждение о том, что зеркальные нейроны участвуют в понимании намерений (посредством моделирования наблюдаемых действий), основано на предположениях, затуманенных нерешенными философскими проблемами. Она утверждает, что намерения понимаются (кодируются) на более сложном уровне нейронной активности, чем уровень активности отдельных нейронов. Черчленд утверждает, что «нейрон, хотя и сложен в вычислительном отношении, это всего лишь нейрон. Это не разумный гомункул. Если нейронная сеть представляет что-то сложное, например намерение [оскорбить], она должна иметь правильные входные данные и находиться в правильное место в нейронной схеме, чтобы сделать это». [119]
Сесилия Хейес выдвинула теорию, согласно которой зеркальные нейроны являются побочным продуктом ассоциативного обучения, а не эволюционной адаптации. Она утверждает, что зеркальные нейроны у людей являются продуктом социального взаимодействия, а не эволюционной адаптацией к пониманию действий. В частности, Хейес отвергает выдвинутую В.С. Рамачандраном теорию о том, что зеркальные нейроны стали «движущей силой большого скачка вперед в эволюции человека». [12] [120]
См. также
[ редактировать ]- Обучение ассоциативной последовательности
- Общая теория кодирования
- Эмоциональное заражение
- Сочувствие
- Синестезия зеркального прикосновения
- Зеркальное отображение (психология)
- Двигательное познание
- Двигательная теория восприятия речи
- Об интеллекте
- Позитронно-эмиссионная томография
- Теория моделирования эмпатии
- Повторение речи
- Веретенообразный нейрон
Ссылки
[ редактировать ]- ^ Jump up to: а б с д Риццолатти Дж., Крейгеро Л. (2004). «Система зеркальных нейронов». Ежегодный обзор неврологии . 27 (1): 169–192. дои : 10.1146/annurev.neuro.27.070203.144230 . ПМИД 15217330 . S2CID 1729870 .
- ^ Кейзерс С. (ноябрь 2009 г.). «Зеркальные нейроны» . Современная биология . 19 (21): Р971–Р973. дои : 10.1016/j.cub.2009.08.026 . hdl : 20.500.11755/c62977db-8a4e-4cc9-b82a-5013b52a0bc6 . ПМИД 19922849 . S2CID 12668046 .
- ^ Jump up to: а б с Кейзерс С (23 июня 2011 г.). Эмпатический мозг . Разжечь.
- ^ Ачарья, Сурья; Шукла, Самарт (2012). «Зеркальные нейроны: загадка метафизического модульного мозга» . Журнал естественных наук, биологии и медицины . 3 (2): 118–124. дои : 10.4103/0976-9668.101878 . ПМЦ 3510904 . ПМИД 23225972 .
- ^ Мукамель Р., Экстром А.Д., Каплан Дж., Якобони М., Фрид И. (апрель 2010 г.). «Однонейронные реакции у человека при выполнении и наблюдении действий» . Современная биология . 20 (8): 750–756. дои : 10.1016/j.cub.2010.02.045 . ПМК 2904852 . ПМИД 20381353 .
- ^ Jump up to: а б Риццолатти Дж., Фадига Л., Фогасси Л., Галлезе В. (май 1999 г.). «Резонансное поведение и зеркальные нейроны» . Итальянский биологический архив . 137 (2–3): 85–100. ПМИД 10349488 .
- ^ Пратер Дж. Ф., Питерс С., Новицки С., Муни Р. (январь 2008 г.). «Точное слухо-вокальное зеркалирование в нейронах для обучения речевому общению». Природа . 451 (7176): 305–310. Бибкод : 2008Natur.451..305P . дои : 10.1038/nature06492 . ПМИД 18202651 . S2CID 4344150 .
- ^ Jump up to: а б Моленбергс П., Каннингтон Р., Мэттингли Дж.Б. (июль 2009 г.). «Участвует ли система зеркальных нейронов в подражании? Краткий обзор и метаанализ». Неврологические и биоповеденческие обзоры . 33 (7): 975–980. doi : 10.1016/j.neubiorev.2009.03.010 . ПМИД 19580913 . S2CID 25620637 .
- ^ Акинс К.К., Кляйн Э.Д., Зенталл Т.Р. (август 2002 г.). «Имитативное обучение японского перепела (Coturnix japonica) с использованием процедуры двунаправленного управления» . Обучение и поведение животных . 30 (3): 275–281. дои : 10.3758/bf03192836 . ПМИД 12391793 .
- ^ Jump up to: а б с д и Динштейн И., Томас С., Берманн М., Хигер DJ (январь 2008 г.). «Зеркало природы» . Современная биология . 18 (1): С13–С18. дои : 10.1016/j.cub.2007.11.004 . ПМК 2517574 . ПМИД 18177704 .
- ^ Jump up to: а б с д Хикок Дж. (июль 2009 г.). «Восемь задач для понимания теории действия зеркальных нейронов у обезьян и людей» . Журнал когнитивной нейронауки . 21 (7): 1229–1243. дои : 10.1162/jocn.2009.21189 . ПМЦ 2773693 . ПМИД 19199415 .
- ^ Jump up to: а б с Привет, Сесилия (март 2010 г.). «Откуда берутся зеркальные нейроны?». Неврологические и биоповеденческие обзоры . 34 (4): 575–583. doi : 10.1016/j.neubiorev.2009.11.007 . ПМИД 19914284 . S2CID 2578537 .
- ^ Феррари ПФ, Риццолатти Дж (2014). «Исследование зеркальных нейронов: прошлое и будущее» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 369 (1644): 20130169. doi : 10.1098/rstb.2013.0169 . ПМК 4006175 . ПМИД 24778369 .
- ^ Кейзерс С., Газзола В. (2006). «На пути к объединяющей нейронной теории социального познания». Понимание эмоций . Прогресс в исследованиях мозга. Том. 156. стр. 379–401. дои : 10.1016/S0079-6123(06)56021-2 . ISBN 9780444521828 . ПМИД 17015092 .
- ^ Арбиб М.А. (апрель 2012 г.). Как мозг обрел язык: гипотеза зеркальной системы . Издательство Оксфордского университета.
- ^ Теорет Х., Паскуаль-Леоне А. (октябрь 2002 г.). «Овладение языком: делай, как слышишь» . Современная биология . 12 (21): Р736–Р737. дои : 10.1016/S0960-9822(02)01251-4 . ПМИД 12419204 .
- ^ Блейксли С. (10 января 2006 г.). «Клетки, читающие мысли» . Нью-Йорк Таймс | Наука .
- ^ ди Пеллегрино Дж., Фадига Л., Фогасси Л., Галлезе В., Риццолатти Дж. (1992). «Понимание двигательных событий: нейрофизиологическое исследование». Экспериментальное исследование мозга . 91 (1): 176–180. дои : 10.1007/bf00230027 . ПМИД 1301372 . S2CID 206772150 .
- ^ Риццолатти Дж., Фадига Л., Галлезе В., Фогасси Л. (март 1996 г.). «Премоторная кора и распознавание двигательных действий». Исследования мозга. Когнитивные исследования мозга . 3 (2): 131–141. дои : 10.1016/0926-6410(95)00038-0 . ПМИД 8713554 .
- ^ Риццолатти Дж., Фаббри-Дестро М. (январь 2010 г.). «Зеркальные нейроны: от открытия до аутизма». Экспериментальное исследование мозга . 200 (3–4): 223–237. дои : 10.1007/s00221-009-2002-3 . ПМИД 19760408 . S2CID 3342808 .
- ^ Jump up to: а б Галлезе В., Фадига Л., Фогасси Л., Риццолатти Дж. (апрель 1996 г.). «Распознавание действий в премоторной коре» . Мозг . 119 (Часть 2) (2): 593–609. дои : 10.1093/мозг/119.2.593 . ПМИД 8800951 .
- ^ Феррари П.Ф., Галлезе В., Риццолатти Дж., Фогасси Л. (апрель 2003 г.). «Зеркальные нейроны, реагирующие на наблюдение за пищевыми и коммуникативными действиями рта в вентральной премоторной коре обезьяны». Европейский журнал неврологии . 17 (8): 1703–1714. дои : 10.1046/j.1460-9568.2003.02601.x . ПМИД 12752388 . S2CID 1915143 .
- ^ Колер Э., Кейзерс С., Умильта М.А., Фогасси Л., Галлезе В., Риццолатти Дж. (август 2002 г.). «Слух звуков, понимание действий: представление действий в зеркальных нейронах». Наука . 297 (5582). Нью-Йорк, штат Нью-Йорк: 846–8. Бибкод : 2002Sci...297..846K . дои : 10.1126/science.1070311 . ПМИД 12161656 . S2CID 16923101 .
- ^ Газзола В., Азиз-Заде Л., Кейзерс С. (сентябрь 2006 г.). «Эмпатия и соматотопическая слуховая зеркальная система у человека» . Современная биология . 16 (18): 1824–9. дои : 10.1016/j.cub.2006.07.072 . ПМИД 16979560 . S2CID 5223812 .
- ^ Jump up to: а б с д Фогасси Л., Феррари П.Ф., Гезерих Б., Роззи С., Черси Ф., Риццолатти Дж. (апрель 2005 г.). «Теменная доля: от организации действия к пониманию намерения». Наука . 308 (5722). Нью-Йорк, штат Нью-Йорк: 662–7. Бибкод : 2005Sci...308..662F . дои : 10.1126/science.1106138 . ПМИД 15860620 . S2CID 5720234 .
- ^ Jump up to: а б с Газзола В., Кейзерс С. (июнь 2009 г.). «Наблюдение и выполнение действий разделяют двигательные и соматосенсорные воксели у всех испытуемых: анализ несглаженных данных фМРТ для одного субъекта» . Кора головного мозга . 19 (6): 1239–1255. дои : 10.1093/cercor/bhn181 . ПМЦ 2677653 . ПМИД 19020203 .
- ^ Кейзерс С., Каас Дж. Х., Газзола В. (июнь 2010 г.). «Соматосенсация в социальном восприятии». Обзоры природы. Нейронаука . 11 (6): 417–428. дои : 10.1038/nrn2833 . ПМИД 20445542 . S2CID 12221575 .
- ^ Jump up to: а б Кук Р., Бёрд Дж., Кэтмур С., Пресс С., Хейес С. (апрель 2014 г.). «Зеркальные нейроны: от происхождения к функции» (PDF) . Поведенческие и мозговые науки . 37 (2): 177–192. дои : 10.1017/S0140525X13000903 . ПМИД 24775147 . S2CID 52873194 .
- ^ Кейзерс С., Газзола В. (2014). «Хеббианское обучение и прогнозирующие зеркальные нейроны действий, ощущений и эмоций» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 369 (1644): 20130175. doi : 10.1098/rstb.2013.0175 . ПМК 4006178 . ПМИД 24778372 .
- ^ Дель Джудис М., Манера В., Кейзерс С. (март 2009 г.). «Запрограммированы на обучение? Онтогенез зеркальных нейронов». Наука развития . 12 (2): 350–363. дои : 10.1111/j.1467-7687.2008.00783.x . hdl : 2318/133096 . ПМИД 19143807 . S2CID 15093476 .
- ^ Феррари П.Ф., Визальберги Э., Паукнер А., Фогасси Л., Руджеро А., Суоми С.Дж. (сентябрь 2006 г.). «Неонатальная имитация у макак-резус» . ПЛОС Биология . 4 (9): е302. doi : 10.1371/journal.pbio.0040302 . ПМК 1560174 . ПМИД 16953662 .
- ^ Феррари ПФ, Бонини Л, Фогасси Л (август 2009 г.). «От зеркальных нейронов обезьяны к поведению приматов: возможные «прямые» и «косвенные» пути» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 364 (1528): 2311–23. дои : 10.1098/rstb.2009.0062 . ПМК 2865083 . ПМИД 19620103 .
- ^ Риццолатти Дж., Арбиб М.А. (май 1998 г.). «Язык в пределах нашей досягаемости». Тенденции в нейронауках . 21 (5): 188–194. дои : 10.1016/S0166-2236(98)01260-0 . hdl : 11858/00-001M-0000-002C-4B59-6 . ПМИД 9610880 . S2CID 679023 .
- ^ Jump up to: а б Кым С., Шин Х.С. (октябрь 2019 г.). «Нейронная основа обучения страху посредством наблюдения: потенциальная модель аффективной эмпатии» . Нейрон . 104 (1): 78–86. дои : 10.1016/j.neuron.2019.09.013 . ПМИД 31600517 . S2CID 203932127 .
- ^ Jump up to: а б с д Каррильо М., Хан Ю., Мильорати Ф., Лю М., Газзола В., Кейзерс К. (апрель 2019 г.). «Эмоциональные зеркальные нейроны в передней поясной извилине крысы» . Современная биология . 29 (8): 1301–1312.e6. дои : 10.1016/j.cub.2019.03.024 . ПМК 6488290 . ПМИД 30982647 .
- ^ Jump up to: а б Ламм С., Десети Дж., Сингер Т. (февраль 2011 г.). «Метааналитические данные об общих и различных нейронных сетях, связанных с непосредственно переживаемой болью и сочувствием к боли». НейроИмидж . 54 (3): 2492–2502. doi : 10.1016/j.neuroimage.2010.10.014 . ПМИД 20946964 . S2CID 6021487 .
- ^ Якобони М., Вудс Р.П., Брасс М., Бекеринг Х. , Мацциотта Дж.К., Риццолатти Дж. (декабрь 1999 г.). «Корковые механизмы подражания человеку». Наука 286 (5449): 2526–2528. дои : 10.1126/science.286.5449.2526 . ПМИД 10617472 .
- ^ Сайгин А.П., Уилсон С.М., Дронкерс Н.Ф., Бейтс Э. (2004). «Понимание действий при афазии: лингвистические и нелингвистические дефициты и их корреляты». Нейропсихология . 42 (13): 1788–1804. CiteSeerX 10.1.1.544.9071 . doi : 10.1016/j.neuropsychologia.2004.04.016 . ПМИД 15351628 . S2CID 11622224 .
- ^ Jump up to: а б Транел Д., Кеммерер Д., Адольфс Р., Дамасио Х., Дамасио А.Р. (май 2003 г.). «Нейронные корреляты концептуальных знаний для действий». Когнитивная нейропсихология . 20 (3): 409–432. дои : 10.1080/02643290244000248 . ПМИД 20957578 . S2CID 16131 .
- ^ Jump up to: а б Сайгин А.П. (сентябрь 2007 г.). «Верхние височные и премоторные области мозга, необходимые для восприятия биологического движения» . Мозг . 130 (Часть 9): 2452–2461. дои : 10.1093/brain/awm162 . ПМИД 17660183 .
- ^ Jump up to: а б Побрич Дж., Гамильтон А.Ф. (март 2006 г.). «Понимание действий требует левой нижней лобной коры» . Современная биология . 16 (5): 524–529. дои : 10.1016/j.cub.2006.01.033 . ПМИД 16527749 .
- ^ Кандиди М., Ургези С., Ионта С., Аглиоти С.М. (2008). «Виртуальное поражение вентральной премоторной коры ухудшает зрительное восприятие биомеханически возможных, но не невозможных действий». Социальная нейронаука . 3 (3–4): 388–400. дои : 10.1080/17470910701676269 . ПМИД 18979387 . S2CID 37390465 .
- ^ Кейзерс К., Газзола В. (апрель 2010 г.). «Социальная нейробиология: зеркальные нейроны, зарегистрированные у людей» . Современная биология . 20 (8): Р353–Р354. дои : 10.1016/j.cub.2010.03.013 . hdl : 20.500.11755/351f0172-b06b-41de-a013-852c64e197fa . ПМИД 21749952 . S2CID 3609747 .
- ^ Моленбергс П., Каннингтон Р., Мэттингли Дж.Б. (январь 2012 г.). «Области мозга с зеркальными свойствами: метаанализ 125 исследований фМРТ человека». Неврологические и биоповеденческие обзоры . 36 (1): 341–349. doi : 10.1016/j.neubiorev.2011.07.004 . ПМИД 21782846 . S2CID 37871374 .
- ^ Фальк-Иттер Т., Гредебек Г., фон Хофстен С. (июль 2006 г.). «Младенцы предсказывают цели действий других людей». Природная неврология . 9 (7): 878–9. дои : 10.1038/nn1729 . ПМИД 16783366 . S2CID 2409686 .
- ^ Кейзерс С., Перретт Д.И. (ноябрь 2004 г.). «Демистификация социального познания: взгляд Хебба». Тенденции в когнитивных науках . 8 (11): 501–507. дои : 10.1016/j.tics.2004.09.005 . ПМИД 15491904 . S2CID 8039741 .
- ^ Эйес С. (июнь 2001 г.). «Причины и последствия подражания». Тенденции в когнитивных науках . 5 (6): 253–261. дои : 10.1016/S1364-6613(00)01661-2 . ПМИД 11390296 . S2CID 15602731 .
- ^ Брасс М., Хейес С. (октябрь 2005 г.). «Имитация: решает ли когнитивная нейронаука проблему соответствия?». Тенденции в когнитивных науках . 9 (10): 489–95. дои : 10.1016/j.tics.2005.08.007 . ПМИД 16126449 . S2CID 1594505 .
- ^ Анисфельд М (1996). «Новорожденным подходит только моделирование высовывания языка». Обзор развития . 16 (2): 149–161. дои : 10.1006/drev.1996.0006 .
- ^ Jump up to: а б Косоногов, В. (декабрь 2012 г.). «Почему зеркальные нейроны не могут поддерживать понимание действий». Нейрофизиология . 44 (6): 499–502. дои : 10.1007/s11062-012-9327-4 . S2CID 254867235 .
- ^ Престон С.Д., де Ваал Ф.Б. (февраль 2002 г.). «Эмпатия: ее конечные и ближайшие основы». Поведенческие и мозговые науки . 25 (1): 1–72. CiteSeerX 10.1.1.554.2794 . дои : 10.1017/s0140525x02000018 . ПМИД 12625087 .
- ^ Десети, Дж (2002). «Натурализующая эмпатия» [Эмпатия натурализованная]. L’Encephale (на французском языке). 28 (1): 9–20. OCLC 110778688 . ИНИСТ 13554627 .
- ^ Десети Дж., Джексон П.Л. (июнь 2004 г.). «Функциональная архитектура человеческой эмпатии». Обзоры поведенческой и когнитивной нейронауки . 3 (2): 71–100. дои : 10.1177/1534582304267187 . ПМИД 15537986 . S2CID 145310279 .
- ^ Jump up to: а б Галлезе В., Голдман А. (декабрь 1998 г.). «Зеркальные нейроны и симуляционная теория чтения мыслей». Тенденции в когнитивных науках . 2 (12): 493–501. дои : 10.1016/s1364-6613(98)01262-5 . ПМИД 21227300 . S2CID 10108122 .
- ^ Галлезе В. (2001). «Гипотеза «Общего многообразия»: от зеркальных нейронов к эмпатии» . Журнал исследований сознания . 8 : 33–50.
- ^ Ботвиник М., Джа А.П., Билсма Л.М., Фабиан С.А., Соломон П.Е., Пркачин К.М. (март 2005 г.). «Наблюдение за выражением боли на лице задействует области коры, участвующие в непосредственном ощущении боли». НейроИмидж . 25 (1): 312–319. doi : 10.1016/j.neuroimage.2004.11.043 . ПМИД 15734365 . S2CID 24988672 .
- ^ Ченг Ю, Ян С.И., Линь С.П., Ли П.Л., Десети Дж. (май 2008 г.). «Восприятие боли у других подавляет соматосенсорные колебания: исследование магнитоэнцефалографии». НейроИмидж . 40 (4): 1833–1840. doi : 10.1016/j.neuroimage.2008.01.064 . ПМИД 18353686 . S2CID 1827514 .
- ^ Моррисон И., Ллойд Д., ди Пеллегрино Дж., Робертс Н. (июнь 2004 г.). «Викарные реакции на боль в передней поясной извилине: является ли эмпатия мультисенсорной проблемой?» . Когнитивная, аффективная и поведенческая нейронаука . 4 (2): 270–278. дои : 10.3758/CABN.4.2.270 . ПМИД 15460933 .
- ^ Уикер Б., Кейзерс С., Плайи Дж., Ройе Дж.П., Галлезе В., Риццолатти Дж. (октябрь 2003 г.). «Мы оба испытываем отвращение в Моей островке: общей нейронной основе видения и чувства отвращения» . Нейрон . 40 (3): 655–64. дои : 10.1016/s0896-6273(03)00679-2 . ПМИД 14642287 . S2CID 766157 .
- ^ Сингер Т., Сеймур Б., О'Догерти Дж., Кабе Х., Долан Р.Дж., Фрит CD (февраль 2004 г.). «Сочувствие боли включает в себя аффективные, а не сенсорные компоненты боли». Наука . 303 (5661). Нью-Йорк, штат Нью-Йорк: 1157–1162. Бибкод : 2004Sci...303.1157S . дои : 10.1126/science.1093535 . hdl : 21.11116/0000-0001-A020-5 . ПМИД 14976305 . S2CID 14727944 .
- ^ Jump up to: а б Джабби М., Сварт М., Кейзерс С. (февраль 2007 г.). «Эмпатия к положительным и отрицательным эмоциям во вкусовой коре». НейроИмидж . 34 (4): 1744–1753. doi : 10.1016/j.neuroimage.2006.10.032 . ПМИД 17175173 . S2CID 13988152 .
- ^ Ламм С., Бэтсон С.Д., Десети Дж. (январь 2007 г.). «Нейральный субстрат человеческой эмпатии: эффекты взгляда на перспективу и когнитивной оценки». Журнал когнитивной нейронауки . 19 (1): 42–58. дои : 10.1162/jocn.2007.19.1.42 . ПМИД 17214562 . S2CID 2828843 .
- ^ Фридберг Д., Галлезе В. (май 2007 г.). «Движение, эмоция и сопереживание в эстетическом опыте». Тенденции в когнитивных науках . 11 (5): 197–203. дои : 10.1016/j.tics.2007.02.003 . ПМИД 17347026 . S2CID 1996468 .
- ^ Джарретт С. (25 марта 2019 г.). «Существуют лишь слабые доказательства того, что зеркальные нейроны лежат в основе человеческой эмпатии – новый обзор и метаанализ» . Исследовательский дайджест . [ мертвая ссылка ]
- ^ Томпсон Э.Л., Бёрд Дж., Кэтмур К. (ноябрь 2022 г.). «Области зеркальных нейронов мозга способствуют распознаванию действий, но не намерений» . Картирование человеческого мозга . 43 (16): 4901–4913. дои : 10.1002/hbm.26036 . ПМЦ 9582378 . ПМИД 35906896 .
- ^ Газзола, Азиз-Заде и Кейзерс (2006). «Современная биология» (PDF) . Архивировано из оригинала (PDF) 30 июня 2007 г.
- ^ Моленбергс П., Хейворд Л., Мэттингли Дж.Б., Каннингтон Р. (январь 2012 г.). «Схемы активации во время наблюдения за действиями модулируются контекстом в областях зеркальной системы». НейроИмидж . 59 (1): 608–615. doi : 10.1016/j.neuroimage.2011.07.080 . ПМИД 21840404 . S2CID 13951700 .
- ^ Ходжат М., Луи Д.З., Майо В., Гоннелла Дж.С. (2013). «Эмпатия и качество здравоохранения». Американский журнал медицинского качества . 28 (1): 6–7. дои : 10.1177/1062860612464731 . ПМИД 23288854 . S2CID 12645544 .
- ^ Эрнандес-Лаллемент Дж., Атта А.Т., Сойман Э., Пинхал К.М., Газзола В., Кейзерс К. (март 2020 г.). «Вред другим действует как негативное подкрепление у крыс» . Современная биология . 30 (6): 949–961.e7. дои : 10.1016/j.cub.2020.01.017 . hdl : 20.500.11755/ee7ae8ac-7393-4276-84ce-1bad1b8e5e0d . ПМИД 32142701 . S2CID 212424287 .
- ^ Оберман Л., Рамачандран В.С. (2009). «Размышления о системе зеркальных нейронов: их эволюционные функции за пределами моторного представления». В Пинеде Дж.А. (ред.). Системы зеркальных нейронов: роль зеркальных процессов в социальном познании . Хумана Пресс. стр. 39–62 . ISBN 978-1-934115-34-3 .
- ^ Рамачандран В.С. (1 января 2009 г.). «Самосознание: последний рубеж, веб-эссе Edge Foundation» . Проверено 26 июля 2011 г.
- ^ Скойлз-младший (2000). «Жест, происхождение языка и правша» (PDF) . Психология . 11 (24).
- ^ Петридес М., Кадорет Г., Макки С. (июнь 2005 г.). «Орофациальные соматомоторные реакции у гомолога зоны Брока у обезьян-макак». Природа . 435 (7046): 1235–1238. Бибкод : 2005Natur.435.1235P . дои : 10.1038/nature03628 . ПМИД 15988526 . S2CID 4397762 .
- ^ Портер Р.Дж., Любкер Дж.Ф. (сентябрь 1980 г.). «Быстрое воспроизведение последовательностей гласных-гласных: свидетельство быстрой и прямой акустико-моторной связи в речи». Журнал исследований речи и слуха . 23 (3): 593–602. дои : 10.1044/jshr.2303.593 . ПМИД 7421161 .
- ^ Маккарти Р., Уоррингтон Э.К. (июнь 1984 г.). «Двухмаршрутная модель производства речи. Данные об афазии» . Мозг . 107 (Часть 2) (2): 463–485. дои : 10.1093/мозг/107.2.463 . ПМИД 6722512 .
- ^ Маккарти Р.А., Уоррингтон Э.К. (2001). «Повторение без семантики: поверхностная дисфазия?». Нейрокейз . 7 (1): 77–87. дои : 10.1093/neucas/7.1.77 . ПМИД 11239078 . S2CID 12988855 .
- ^ Марслен-Уилсон В. (август 1973 г.). «Лингвистическая структура и речевое затенение с очень короткими задержками». Природа . 244 (5417): 522–523. Бибкод : 1973Natur.244..522M . дои : 10.1038/244522a0 . ПМИД 4621131 . S2CID 4220775 .
- ^ Фэй WH, Коулман Р.О. (июль 1977 г.). «Человеческий преобразователь / воспроизводитель звука: временные возможности глубоко эхолалического ребенка». Мозг и язык . 4 (3): 396–402. дои : 10.1016/0093-934X(77)90034-7 . ПМИД 907878 . S2CID 29492873 .
- ^ Шипперс М.Б., Робрук А., Ренкен Р., Нанетти Л., Кейзерс С. (май 2010 г.). «Отображение потока информации от одного мозга к другому во время жестового общения» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (20): 9388–9393. Бибкод : 2010PNAS..107.9388S . дои : 10.1073/pnas.1001791107 . ПМК 2889063 . ПМИД 20439736 .
- ^ Моро А (2008). Границы Вавилона. Мозг и загадка невозможных языков . МТИ Пресс . п. 257. ИСБН 978-0-262-13498-9 .
- ^ Лонго М.Р., Кособуд А., Бертенталь Б.И. (апрель 2008 г.). «Автоматическая имитация биомеханически возможных и невозможных действий: эффекты стартовых движений и целей» (PDF) . Журнал экспериментальной психологии. Человеческое восприятие и производительность . 34 (2): 489–501. дои : 10.1037/0096-1523.34.2.489 . ПМИД 18377184 .
- ^ Косоногов В (октябрь 2011 г.). «Прослушивание предложений, связанных с действием, ухудшает постуральный контроль». Журнал электромиографии и кинезиологии . 21 (5): 742–745. doi : 10.1016/j.jelekin.2011.05.007 . ПМИД 21705230 .
- ^ Чартран Т.Л., Барг Дж.А. (июнь 1999 г.). «Эффект хамелеона: связь восприятия и поведения и социальное взаимодействие». Журнал личности и социальной психологии . 76 (6): 893–910. дои : 10.1037/0022-3514.76.6.893 . ПМИД 10402679 . S2CID 11818459 .
- ^ Лакин Дж.Л., Чартран Т.Л. (июль 2003 г.). «Использование бессознательной поведенческой мимикрии для создания принадлежности и взаимопонимания». Психологическая наука . 14 (4): 334–339. дои : 10.1111/1467-9280.14481 . ПМИД 12807406 . S2CID 8458849 .
- ^ ван Баарен Р.Б., Мэддукс В.В., Чартран Т.Л., де Бутер С., ван Книппенберг А. (май 2003 г.). «Чтобы имитировать, нужны двое: поведенческие последствия самооценки». Журнал личности и социальной психологии . 84 (5): 1093–1102. дои : 10.1037/0022-3514.84.5.1093 . hdl : 2066/63130 . ПМИД 12757151 . S2CID 729948 .
- ^ Привет С (май 2011 г.). «Автоматическая имитация». Психологический вестник . 137 (3): 463–483. дои : 10.1037/a0022288 . ПМИД 21280938 . S2CID 6975248 .
- ^ Паукнер А., Суоми С.Дж., Визальберги Э., Ferrari PF (август 2009 г.). «Обезьяны-капуцины проявляют привязанность к людям, которые им подражают» . Наука . 325 (5942): 880–883. Бибкод : 2009Sci...325..880P . дои : 10.1126/science.1176269 . ПМК 2764469 . ПМИД 19679816 .
- ^ Jump up to: а б Оберман Л.М., Хаббард Э.М., Макклири Дж.П., Альтшулер Э.Л., Рамачандран В.С., Пинеда Дж.А. (июль 2005 г.). «ЭЭГ-доказательства дисфункции зеркальных нейронов при расстройствах аутистического спектра». Исследования мозга. Когнитивные исследования мозга . 24 (2): 190–198. doi : 10.1016/j.cogbrainres.2005.01.014 . ПМИД 15993757 .
- ^ Дапретто М., Дэвис М.С., Пфайфер Дж.Х., Скотт А.А., Сигман М., Букхаймер С.Ю., Якобони М. (январь 2006 г.). «Понимание эмоций других: дисфункция зеркальных нейронов у детей с расстройствами аутистического спектра» . Природная неврология . 9 (1): 28–30. дои : 10.1038/nn1611 . ПМЦ 3713227 . ПМИД 16327784 .
- ^ Дапретто М., Дэвис М.С., Пфайфер Дж.Х., Скотт А.А., Сигман М., Букхаймер С.Ю., Якобони М. (январь 2006 г.). «Понимание эмоций других: дисфункция зеркальных нейронов у детей с расстройствами аутистического спектра» . Природная неврология . 9 (1): 28–30. дои : 10.1038/nn1611 . ПМЦ 3713227 . ПМИД 16327784 .
- ^ Перкинс Т., Стоукс М., Макгилливрей Дж., Биттар Р. (октябрь 2010 г.). «Дисфункция зеркальных нейронов при расстройствах аутистического спектра». Журнал клинической неврологии . 17 (10): 1239–1243. дои : 10.1016/j.jocn.2010.01.026 . ПМИД 20598548 . S2CID 15141982 .
- ^ Оберман Л.М., Рамачандран В.С., Пинеда Дж.А. (апрель 2008 г.). «Модуляция мю-супрессии у детей с расстройствами аутистического спектра в ответ на знакомые или незнакомые стимулы: гипотеза зеркальных нейронов». Нейропсихология . 46 (5): 1558–1565. doi : 10.1016/j.neuropsychologia.2008.01.010 . ПМИД 18304590 . S2CID 14280719 .
- ^ Черкленд PS (2011). «6». Мозговое доверие . Издательство Принстонского университета. п. 156.
- ^ Гамильтон А., Марш Л. (август 2013 г.). «Две системы понимания действий при аутизме: зеркалирование и ментализация». (PDF) . В Барон-Коэн С., Ломбардо М., Тагер-Флюсберг Х. (ред.). Понимание других умов: перспективы социальной нейробиологии развития . Оксфорд: ОУП. стр. 380–396. ISBN 978-0-19-166880-7 .
- ^ Хаджихани Н., Джозеф Р.М., Снайдер Дж., Тагер-Флюсберг Х. (сентябрь 2006 г.). «Анатомические различия в системе зеркальных нейронов и сети социального познания при аутизме» . Кора головного мозга . 16 (9): 1276–1282. дои : 10.1093/cercor/bhj069 . ПМИД 16306324 .
- ^ Jump up to: а б Каллауэй Э (12 мая 2010 г.). «Зеркальные нейроны при аутизме ведут себя нормально» . Новый учёный .
- ^ Гамильтон АФ (август 2009 г.). «Цели, намерения и психические состояния: проблемы теорий аутизма». Журнал детской психологии, психиатрии и смежных дисциплин . 50 (8): 881–892. CiteSeerX 10.1.1.621.6275 . дои : 10.1111/j.1469-7610.2009.02098.x . ПМИД 19508497 .
- ^ Мерфи П., Брэди Н., Фицджеральд М., Трое Н.Ф. (декабрь 2009 г.). «Нет доказательств нарушения восприятия биологического движения у взрослых с расстройствами аутистического спектра». Нейропсихология . 47 (14): 3225–3235. doi : 10.1016/j.neuropsychologia.2009.07.026 . ПМИД 19666038 . S2CID 12495492 .
- ^ Сайгин А.П., Кук Дж., Блейкмор С.Дж. (октябрь 2010 г.). Бейкер К.И. (ред.). «Незатронутые пороги восприятия для биологического и небиологического восприятия форм из движения в условиях аутистического спектра» . ПЛОС ОДИН . 5 (10): e13491. Бибкод : 2010PLoSO...513491S . дои : 10.1371/journal.pone.0013491 . ПМЦ 2956672 . ПМИД 20976151 .
- ^ Кук Дж., Сайгин А.П., Суэйн Р., Блейкмор С.Дж. (декабрь 2009 г.). «Снижение чувствительности к биологическим движениям с минимальными толчками в условиях аутистического спектра» . Нейропсихология . 47 (14): 3275–3278. doi : 10.1016/j.neuropsychologia.2009.07.010 . ПМЦ 2779370 . ПМИД 19632248 .
- ^ Фань Ю.Т., Десети Дж., Ян С.И., Лю Дж.Л., Ченг Ю. (сентябрь 2010 г.). «Неразрушенные зеркальные нейроны при расстройствах аутистического спектра». Журнал детской психологии, психиатрии и смежных дисциплин . 51 (9): 981–988. дои : 10.1111/j.1469-7610.2010.02269.x . ПМИД 20524939 .
- ^ Хейес С., Катмур С. (январь 2022 г.). «Что случилось с зеркальными нейронами?» . Перспективы психологической науки . 17 (1): 153–168. дои : 10.1177/1745691621990638 . ПМЦ 8785302 . ПМИД 34241539 .
- ^ Гордон Р. (1986). «Народная психология как симуляция». Разум и язык . 1 (2): 158–171. дои : 10.1111/j.1468-0017.1986.tb00324.x .
- ^ Гольдман А (1989). «Интерпретация психологизированная». Разум и язык . 4 (3): 161–185. дои : 10.1111/j.1468-0017.1989.tb00249.x .
- ^ Мельцов А.Н., Децети Дж. (март 2003 г.). «Что подражание говорит нам о социальном познании: сближение психологии развития и когнитивной нейробиологии» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 358 (1431): 491–500. дои : 10.1098/rstb.2002.1261 . ПМЦ 1351349 . ПМИД 12689375 .
- ^ Соммервилл Дж. А., Десети Дж. (апрель 2006 г.). «Плетение ткани социального взаимодействия: формулирование психологии развития и когнитивной нейробиологии в области моторного познания». Психономический бюллетень и обзор . 13 (2): 179–200. дои : 10.3758/BF03193831 . ПМИД 16892982 . S2CID 14689479 .
- ^ Кейзерс К., Газзола В. (май 2007 г.). «Интеграция моделирования и теории разума: от себя к социальному познанию». Тенденции в когнитивных науках . 11 (5): 194–196. дои : 10.1016/j.tics.2007.02.002 . ПМИД 17344090 . S2CID 18930071 .
- ^ Харуш К., Уильямс З.М. (март 2015 г.). «Нейронное предсказание поведения противника во время совместного социального обмена у приматов» . Клетка . 160 (6): 1233–1245. дои : 10.1016/j.cell.2015.01.045 . ПМК 4364450 . ПМИД 25728667 .
- ^ Ченг Ю.В., Цзенг О.Дж., Десети Дж., Имада Т., Се Дж.К. (июль 2006 г.). «Гендерные различия в зеркальной системе человека: исследование магнитоэнцефалографии». НейроОтчёт . 17 (11): 1115–1119. дои : 10.1097/01.wnr.0000223393.59328.21 . ПМИД 16837838 . S2CID 18811017 .
- ^ Ченг Ю., Десети Дж., Линь С.П., Се Дж.К., Хунг Д., Цзэн О.Дж. (июнь 2007 г.). «Половые различия в возбудимости позвоночника при наблюдении за двуногой локомоцией». НейроОтчёт . 18 (9): 887–890. дои : 10.1097/WNR.0b013e3280ebb486 . ПМИД 17515795 . S2CID 16295878 .
- ^ Ян С.И., Десети Дж., Ли С., Чен С., Ченг Ю. (январь 2009 г.). «Гендерные различия в мю-ритме при сочувствии боли: электроэнцефалографическое исследование». Исследования мозга . 1251 : 176–184. дои : 10.1016/j.brainres.2008.11.062 . ПМИД 19083993 . S2CID 40145972 .
- ^ Ченг Ю, Ли П.Л., Ян С.И., Линь С.П., Хунг Д., Десети Дж. (май 2008 г.). «Гендерные различия в мю-ритме системы зеркальных нейронов человека» . ПЛОС ОДИН . 3 (5): е2113. Бибкод : 2008PLoSO...3.2113C . дои : 10.1371/journal.pone.0002113 . ПМК 2361218 . ПМИД 18461176 .
- ^ Шульте-Рютер М., Маркович Х.Дж., Шах Н.Дж., Финк Г.Р., Пифке М. (август 2008 г.). «Гендерные различия в мозговых сетях, поддерживающих эмпатию». НейроИмидж . 42 (1): 393–403. doi : 10.1016/j.neuroimage.2008.04.180 . ПМИД 18514546 . S2CID 10461927 .
- ^ Jump up to: а б с Джалал Б., Рамачандран В.С. (2017). «Сонный паралич, «Призрачный злоумышленник в спальне» и внетелесные переживания: роль зеркальных нейронов» . Границы человеческой неврологии . 11:92 . дои : 10.3389/fnhum.2017.00092 . ПМК 5329044 . ПМИД 28293186 .
- ^ Маккормик Л.М., Брамм М.С., Бидл Дж.Н., Парадизо С., Ямада Т., Андреасен Н. (март 2012 г.). «Функция зеркальных нейронов, психоз и эмпатия при шизофрении» . Психиатрические исследования . 201 (3): 233–239. doi : 10.1016/j.pscychresns.2012.01.004 . ПМЦ 3545445 . ПМИД 22510432 .
- ^ Jump up to: а б Пасколо П.Б., Рагонья Р., Росси Р. (2009). «Парадигма системы зеркальных нейронов и ее последовательность». Походка и осанка . 30 (Приложение 1): 65. doi : 10.1016/j.gaitpost.2009.07.064 .
- ^ Лингнау А., Гезерих Б., Карамацца А. (июнь 2009 г.). «Асимметричная адаптация фМРТ не выявила доказательств существования зеркальных нейронов у людей» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (24): 9925–9930. Бибкод : 2009PNAS..106.9925L . дои : 10.1073/pnas.0902262106 . ПМК 2701024 . ПМИД 19497880 .
- ^ Килнер Дж. М., Нил А., Вайскопф Н., Фристон К. Дж., Фрит CD (август 2009 г.). «Свидетельства наличия зеркальных нейронов в нижней лобной извилине человека» . Журнал неврологии . 29 (32): 10153–10159. doi : 10.1523/jneurosci.2668-09.2009 . ПМК 2788150 . ПМИД 19675249 .
- ^ Черчленд, Патриция, Braintrust (2011), глава 6, стр. 142
- ^ Рамачандран В.С. (2000). «Зеркальные нейроны и имитационное обучение как движущая сила «большого скачка вперед» в эволюции человека» . Край . Проверено 13 апреля 2013 г.
Дальнейшее чтение
[ редактировать ]- Кэри Б. (12 февраля 2008 г.). «Ты напоминаешь мне меня» . Нью-Йорк Таймс .
- Хикок Дж. (2014). Миф о зеркальных нейронах . Национальные географические книги. ISBN 978-0-393-08961-5 .
- Кейзерс С (2011). Эмпатический мозг . Независимая издательская платформа CreateSpace. ISBN 978-1-4637-6906-2 .
- Риццолатти Дж., Синигалья К. (2008). Зеркала в мозгу. Как мы делимся своими действиями и эмоциями . Оксфорд (Великобритания): Издательство Оксфордского университета . ISBN 978-0-19-921798-4 .
- Морселла Э., Барг Дж.А., Голлвитцер П.М., ред. (2009). Оксфордский справочник по человеческой деятельности . Нью-Йорк: Издательство Оксфордского университета . ISBN 978-0-19-530998-0 .
- Галлезе В., Гернсбахер М.А., Хейес С., Хикок Г., Якобони М. (июль 2011 г.). «Форум зеркальных нейронов» . Перспективы психологической науки . 6 (4): 369–407. дои : 10.1177/1745691611413392 . ПМК 4266473 . ПМИД 25520744 .
- Якобони М., Мацциотта Дж.К. (сентябрь 2007 г.). «Система зеркальных нейронов: основные открытия и клиническое применение» . Анналы неврологии . 62 (3): 213–218. дои : 10.1002/ana.21198 . ПМИД 17721988 . S2CID 3225339 .
- Кейзерс С., Газзола В. (2006). «На пути к объединяющей нейронной теории социального познания». Понимание эмоций . Прогресс в исследованиях мозга. Том. 156. стр. 379–401. дои : 10.1016/S0079-6123(06)56021-2 . ISBN 9780444521828 . ПМИД 17015092 .
- Пасколо Б (2013). «Зеркальные нейроны: вопрос все еще открыт?» (PDF) . Прогресс в нейробиологии . 1 (1–4): 25–82.
- Престон С.Д., де Ваал Ф.Б. (февраль 2002 г.). «Эмпатия: ее конечные и ближайшие основы». Поведенческие и мозговые науки . 25 (1): 1–72. дои : 10.1017/s0140525x02000018 . ПМИД 12625087 .
- Риццолатти Дж., Фаббри-Дестро М., Каттанео Л. (январь 2009 г.). «Зеркальные нейроны и их клиническое значение». Природная клиническая практика. Неврология . 5 (1): 24–34. дои : 10.1038/ncpneuro0990 . ПМИД 19129788 . S2CID 2979216 .
Внешние ссылки
[ редактировать ]- Хикок Дж., Поппель Д. «Говорящие мозги» .
Новости и взгляды на нейронную организацию языка
- Рамачандран VS. «Нейроны, сформировавшие цивилизацию» . TED беседует . Архивировано из оригинала 10 февраля 2012 года.
- Томас Б. (2012). «Что такого особенного в зеркальных нейронах?» . Гостевой блог Scientific American . (обзор известных исследовательских подходов на основе интервью с Якобони, Хикоком, Хейесом и Галлезе)
- «Зеркальные нейроны» . НОВА наука СЕЙЧАС . Январь 2005 г. Архивировано из оригинала 11 января 2006 г.
| Базы данных органов управления : Национальные |
|---|