Нейронные колебания


Нейронные колебания , или мозговые волны , представляют собой ритмические или повторяющиеся паттерны нейронной активности в центральной нервной системе . Нервная ткань может генерировать колебательную активность разными способами, обусловленную либо механизмами внутри отдельных нейронов , либо взаимодействием между нейронами. В отдельных нейронах колебания могут проявляться либо как колебания мембранного потенциала , либо как ритмические паттерны потенциалов действия , которые затем вызывают колебательную активацию постсинаптических нейронов. На уровне нейронных ансамблей синхронизированная активность большого числа нейронов может вызывать макроскопические колебания, которые можно наблюдать на электроэнцефалограмме . Колебательная активность в группах нейронов обычно возникает в результате обратных связей между нейронами, что приводит к синхронизации их импульсов. Взаимодействие между нейронами может вызывать колебания с частотой, отличной от частоты срабатывания отдельных нейронов. Хорошо известным примером макроскопических нейронных колебаний является альфа-активность .
Нейронные колебания у человека наблюдались исследователями еще в 1924 году ( Ганс Бергер ). Более 50 лет спустя собственное колебательное поведение было обнаружено в нейронах позвоночных, но его функциональная роль до сих пор не до конца понята. [3] Возможная роль нейронных колебаний включает связывание функций , механизмы передачи информации и генерацию ритмической двигательной активности . За последние десятилетия было получено больше информации, особенно благодаря достижениям в области визуализации мозга . Основная область исследований в области нейробиологии связана с определением того, как генерируются колебания и какова их роль. Колебательная активность мозга широко наблюдается на разных уровнях организации и считается, что она играет ключевую роль в обработке нейронной информации. Многочисленные экспериментальные исследования подтверждают функциональную роль нейронных колебаний; однако единая интерпретация все еще отсутствует.
История
[ редактировать ]Ричард Кейтон обнаружил электрическую активность в полушариях головного мозга кроликов и обезьян и представил свои открытия в 1875 году. [4] Адольф Бек опубликовал в 1890 году свои наблюдения за спонтанной электрической активностью мозга кроликов и собак, включавшей измененные светом ритмические колебания, регистрируемые с помощью электродов, помещенных непосредственно на поверхность мозга. [5] До Ганса Бергера Владимир Владимирович Правдич-Неминский опубликовал первые ЭЭГ животных и вызванный потенциал собаки. [6]
Обзор
[ редактировать ]Нейронные колебания наблюдаются во всей центральной нервной системе на всех уровнях и включают в себя серии спайков , потенциалы локального поля и крупномасштабные колебания , которые можно измерить с помощью электроэнцефалографии (ЭЭГ). В общем виде колебания можно охарактеризовать их частотой , амплитудой и фазой . Эти свойства сигнала можно извлечь из нейронных записей с помощью частотно-временного анализа . Считается, что в крупномасштабных колебаниях изменения амплитуды являются результатом изменений синхронизации внутри нейронного ансамбля , также называемых локальной синхронизацией. Помимо локальной синхронизации, может синхронизироваться и колебательная активность отдаленных нейронных структур (одиночных нейронов или нейронных ансамблей). Нейронные колебания и синхронизация связаны со многими когнитивными функциями, такими как передача информации, восприятие, двигательный контроль и память. [7] [8] [9] [10]
Противоположностью синхронизации нейронов является нейронная изоляция, то есть электрическая активность нейронов не синхронизирована во времени. Это когда вероятность того, что нейрон достигнет своего порогового потенциала для распространения сигнала на следующий нейрон, снижается. Это явление обычно наблюдается, когда спектральная интенсивность уменьшается в результате суммирования импульсов этих нейронов, что можно использовать для дифференциации когнитивных функций или изоляции нейронов. Однако были использованы новые нелинейные методы, которые одновременно объединяют временные и спектральные энтропийные отношения, чтобы охарактеризовать то, как изолированы нейроны (неспособность сигнала распространяться на соседние нейроны), что является индикатором нарушения (например, гипоксии). [1]
Нейронные колебания наиболее широко изучались при нейронной активности, генерируемой большими группами нейронов. Масштабную активность можно измерить с помощью таких методов, как ЭЭГ. В целом сигналы ЭЭГ имеют широкий спектральный состав, аналогичный розовому шуму , но также обнаруживают колебательную активность в определенных диапазонах частот. Первый обнаруженный и наиболее известный диапазон частот — альфа-активность (8–12 Гц ). [11] [12] [13] который можно обнаружить в затылочной доле во время спокойного бодрствования и который увеличивается при закрытых глазах. [14] Другие диапазоны частот: дельта (1–4 Гц), тета (4–8 Гц), бета (13–30 Гц), низкая гамма (30–70 Гц), [15] и высокие гамма-диапазоны (70–150 Гц). Более быстрые ритмы, такие как гамма-активность, связаны с когнитивной обработкой. Действительно, сигналы ЭЭГ резко изменяются во время сна и демонстрируют переход от более высоких частот к более медленным, таким как альфа-волны. Фактически, различные стадии сна обычно характеризуются своим спектральным составом. [16] Следовательно, нейронные колебания связаны с когнитивными состояниями, такими как осведомленность и сознание . [17] [18] [15] [13]
Хотя нейронные колебания в активности мозга человека в основном исследуются с помощью записей ЭЭГ, их также можно наблюдать с помощью более инвазивных методов записи, таких как единичные записи . Нейроны могут генерировать ритмические паттерны потенциалов действия или спайков. Некоторые типы нейронов имеют тенденцию срабатывать на определенных частотах, выступая в роли резонаторов. [19] или как собственные генераторы . [2] Взрыв — это еще одна форма ритмических всплесков. Пиковые паттерны считаются фундаментальными для кодирования информации в мозге. Колебательная активность может наблюдаться и в виде подпороговых колебаний мембранного потенциала (т.е. при отсутствии потенциалов действия). [20] Если многочисленные нейроны активируются синхронно , они могут вызвать колебания локальных потенциалов поля . Количественные модели могут оценить силу нейронных колебаний в записанных данных. [21]
Нейронные колебания обычно изучаются в математических рамках и относятся к области «нейродинамики», области исследований в области когнитивных наук , в которой особое внимание уделяется динамическому характеру нейронной активности при описании функций мозга . [22] Он рассматривает мозг как динамическую систему и использует дифференциальные уравнения для описания того, как нейронная активность развивается с течением времени. В частности, он направлен на то, чтобы связать динамические закономерности активности мозга с когнитивными функциями, такими как восприятие и память. В очень абстрактной форме нейронные колебания можно анализировать аналитически . [23] [24] При изучении в более физиологически реалистичных условиях колебательная активность обычно изучается с использованием компьютерного моделирования модели вычислительной .
Функции нейронных колебаний весьма разнообразны и различаются для разных типов колебательной деятельности. Примерами являются генерация ритмической активности, такой как сердцебиение , и нейронная связь сенсорных характеристик восприятия, таких как форма и цвет объекта. Нейронные колебания также играют важную роль при многих неврологических расстройствах , таких как чрезмерная синхронизация во время судорожной активности при эпилепсии или тремор у пациентов с болезнью Паркинсона . Колебательную активность также можно использовать для управления внешними устройствами, такими как интерфейс мозг-компьютер . [25]
Физиология
[ редактировать ]Колебательная активность наблюдается во всей центральной нервной системе на всех уровнях организации. Широко признаны три различных уровня: микромасштаб (активность одного нейрона), мезомасштаб (активность локальной группы нейронов) и макромасштаб (активность различных областей мозга). [26]
микроскопический
[ редактировать ]Нейроны генерируют потенциалы действия в результате изменений электрического мембранного потенциала. Нейроны могут последовательно генерировать несколько потенциалов действия, образуя так называемые последовательности спайков. Эти цепочки спайков являются основой нейронного кодирования и передачи информации в мозгу. Последовательности шипов могут образовывать всевозможные паттерны, такие как ритмические всплески и взрывы , и часто демонстрируют колебательную активность. [27] Колебательную активность в одиночных нейронах можно наблюдать и при подпороговых колебаниях мембранного потенциала. Эти ритмические изменения мембранного потенциала не достигают критического порога и поэтому не приводят к возникновению потенциала действия. Они могут быть результатом постсинаптических потенциалов синхронных входов или внутренних свойств нейронов.
Нейрональные спайки можно классифицировать по характеру активности. Возбудимость нейронов можно разделить на I и II классы. Нейроны класса I могут генерировать потенциалы действия с произвольно низкой частотой в зависимости от силы входного сигнала, тогда как нейроны класса II генерируют потенциалы действия в определенном диапазоне частот, который относительно нечувствителен к изменениям силы входного сигнала. [19] Нейроны класса II также более склонны к подпороговым колебаниям мембранного потенциала.
мезоскопический
[ редактировать ]Группа нейронов также может генерировать колебательную активность. Благодаря синаптическим взаимодействиям паттерны возбуждения различных нейронов могут синхронизироваться, а ритмические изменения электрического потенциала, вызванные их потенциалами действия, могут накапливаться ( конструктивная интерференция ). То есть синхронизированные паттерны возбуждения приводят к синхронизированному входному сигналу в другие области коры, что приводит к колебаниям потенциала локального поля большой амплитуды . Эти крупномасштабные колебания также можно измерить за пределами кожи головы с помощью электроэнцефалографии (ЭЭГ) и магнитоэнцефалографии (МЭГ). Электрические потенциалы, генерируемые отдельными нейронами, слишком малы, чтобы их можно было уловить за пределами кожи головы, а активность ЭЭГ или МЭГ всегда отражает сумму синхронной активности тысяч или миллионов нейронов, имеющих одинаковую пространственную ориентацию. [28]
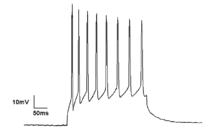
Нейроны в нейронном ансамбле редко срабатывают в один и тот же момент, то есть полностью синхронизируются. Вместо этого вероятность срабатывания ритмически модулируется, так что нейроны с большей вероятностью сработают одновременно, что приводит к колебаниям их средней активности. (См. рисунок вверху страницы.) Таким образом, частота крупномасштабных колебаний не обязательно должна соответствовать схеме срабатывания отдельных нейронов. Изолированные кортикальные нейроны регулярно активируются при определенных условиях, но в неповрежденном мозге корковые клетки бомбардируются сильно меняющимися синаптическими входами и обычно активируются, казалось бы, случайным образом. Однако если вероятность срабатывания большой группы нейронов ритмически модулировать с общей частотой, они будут генерировать колебания в среднем поле. (См. также рисунок вверху страницы.) [27]
Нейронные ансамбли могут генерировать колебательную активность эндогенно посредством локальных взаимодействий между возбуждающими и тормозными нейронами. В частности, тормозные интернейроны играют важную роль в обеспечении синхронности нейронных ансамблей, создавая узкое окно для эффективного возбуждения и ритмически модулируя частоту возбуждения возбуждающих нейронов. [29]
Макроскопический
[ редактировать ]Нейронные колебания также могут возникать в результате взаимодействия между различными областями мозга, связанными через структурный коннектом . временные задержки Здесь важную роль играют . Поскольку все области мозга двунаправленно связаны, эти связи между областями мозга образуют обратной связи петли . Петли положительной обратной связи имеют тенденцию вызывать колебательную активность, частота которой обратно пропорциональна времени задержки. Примером такой петли обратной связи являются связи между таламусом и корой головного мозга – таламокортикальные излучения . Эта таламокортикальная сеть способна генерировать колебательную активность, известную как периодический таламо-кортикальный резонанс . [30] Таламокортикальная сеть играет важную роль в генерации альфа-активности . [31] [32] В сетевой модели всего мозга с реалистичной анатомической связностью и задержками распространения между областями мозга колебания в бета-диапазоне частот возникают в результате частичной синхронизации подмножеств областей мозга, колеблющихся в гамма-диапазоне (генерируемых на мезоскопическом уровне). [33]
Механизмы
[ редактировать ]Нейрональные свойства
[ редактировать ]Ученые определили некоторые внутренние свойства нейронов , которые играют важную роль в генерации колебаний мембранного потенциала. В частности, потенциалзависимые ионные каналы имеют решающее значение для генерации потенциалов действия. Динамика этих ионных каналов отражена в хорошо зарекомендовавшей себя модели Ходжкина-Хаксли , которая описывает, как потенциалы действия инициируются и распространяются с помощью набора дифференциальных уравнений. Используя бифуркационный анализ , можно определить различные колебательные разновидности этих нейронных моделей, что позволяет классифицировать типы нейрональных ответов. Колебательная динамика нейрональных импульсов, определенная в модели Ходжкина-Хаксли, близко согласуется с эмпирическими данными.
Помимо периодических всплесков, подпороговые колебания мембранного потенциала , то есть резонансное поведение, которое не приводит к возникновению потенциалов действия, также могут способствовать колебательной активности, способствуя синхронной активности соседних нейронов. [34] [35]
Подобно нейронам-водителям ритма в генераторах центральных паттернов , подтипы корковых клеток ритмично выдают всплески импульсов (краткие группы спайков) на предпочтительных частотах. [2] Взрывные нейроны могут служить кардиостимуляторами для синхронных колебаний сети, а всплески спайков могут лежать в основе или усиливать нейрональный резонанс. [27] Многие из этих нейронов можно считать собственными осцилляторами, а именно нейронами, которые сами генерируют свои колебания, поскольку их частоты колебаний могут быть изменены локальным применением глутамата in vivo. [36]
Свойства сети
[ редактировать ]Помимо внутренних свойств нейронов, биологических нейронных сетей важным источником колебательной активности также являются свойства . Нейроны общаются друг с другом через синапсы и влияют на время прохождения импульсов в постсинаптических нейронах. В зависимости от свойств связи, таких как сила связи, временная задержка и является ли связь возбуждающей или тормозящей , серии спайков взаимодействующих нейронов могут синхронизироваться . [37] Нейроны локально связаны, образуя небольшие кластеры, называемые нейронными ансамблями . Определенные сетевые структуры способствуют колебательной активности на определенных частотах. Например, активность нейронов, генерируемая двумя популяциями взаимосвязанных тормозных и возбуждающих клеток, может демонстрировать спонтанные колебания, которые описываются моделью Уилсона-Коуэна .
Если группа нейронов участвует в синхронизированной колебательной активности, нейронный ансамбль можно математически представить как один осциллятор. [26] Различные нейронные ансамбли связаны между собой дальнодействующими связями и образуют сеть слабо связанных осцилляторов следующего пространственного масштаба. Слабо связанные осцилляторы могут генерировать ряд динамики, включая колебательную активность. [38] Дальние связи между различными структурами мозга, такими как таламус и кора головного мозга (см. Таламокортикальные колебания ), включают временные задержки из-за конечной скорости проводимости аксонов. Поскольку большинство связей взаимны, они образуют петли обратной связи , поддерживающие колебательную активность. Колебания, записанные из нескольких областей коры, могут синхронизироваться, образуя крупномасштабные мозговые сети , динамику и функциональные связи которых можно изучать с помощью спектрального анализа и причинно-следственных связей Грейнджера . [39] Когерентная деятельность крупномасштабной деятельности мозга может образовывать динамические связи между областями мозга, необходимые для интеграции распределенной информации. [18]
Было показано, что микроглия — основные иммунные клетки мозга — играет важную роль в формировании сетевых связей и, таким образом, влияет на колебания нейронной сети как ex vivo , так и in vivo . [40]
Нейромодуляция
[ редактировать ]Помимо быстрых прямых синаптических взаимодействий между нейронами, образующими сеть, колебательная активность регулируется нейромодуляторами в гораздо более медленном временном масштабе. То есть известно, что уровни концентрации определенных нейротрансмиттеров регулируют величину колебательной активности. Например, ГАМК положительно коррелирует с частотой колебаний индуцированных стимулов. было показано, что концентрация [41] Ряд ядер в стволе мозга имеют диффузные проекции по всему мозгу, влияющие на уровень концентрации нейротрансмиттеров, таких как норадреналин , ацетилхолин и серотонин . Эти нейромедиаторные системы влияют на физиологическое состояние, например, на бодрствование или возбуждение , и оказывают выраженное влияние на амплитуду различных мозговых волн, таких как альфа-активность. [42]
Математическое описание
[ редактировать ]Колебания часто можно описать и проанализировать с помощью математики. Математики определили несколько динамических механизмов, генерирующих ритмичность. Среди наиболее важных — гармонические (линейные) генераторы, генераторы предельного цикла и с запаздывающей обратной связью . генераторы [43] Гармонические колебания очень часто возникают в природе — примерами являются звуковые волны, движение маятника и всякого рода вибрации. Обычно они возникают, когда физическая система в небольшой степени выходит из состояния с минимальной энергией , и хорошо понятны математически.
Гармонические генераторы, управляемые шумом, реалистично имитируют альфа-ритм на ЭЭГ бодрствования, а также медленные волны и веретена на ЭЭГ сна. Успешные алгоритмы анализа ЭЭГ были основаны на таких моделях. Некоторые другие компоненты ЭЭГ лучше описываются колебаниями предельного цикла или с задержкой обратной связи.
Колебания предельного цикла возникают в физических системах, которые демонстрируют большие отклонения от равновесия , тогда как колебания с запаздывающей обратной связью возникают, когда компоненты системы влияют друг на друга после значительных временных задержек. Колебания предельного цикла могут быть сложными, но для их анализа существуют мощные математические инструменты; математика колебаний с запаздывающей обратной связью по сравнению с этим примитивна. Линейные генераторы и генераторы предельного цикла качественно различаются по тому, как они реагируют на колебания входного сигнала. В линейном генераторе частота более или менее постоянна, но амплитуда может сильно меняться. В генераторе с предельным циклом амплитуда имеет тенденцию быть более или менее постоянной, но частота может сильно меняться. Сердцебиение . является примером колебаний предельного цикла, при котором частота ударов варьируется в широких пределах, в то время как каждый отдельный удар продолжает перекачивать примерно одинаковое количество крови
Вычислительные модели используют различные абстракции для описания сложной колебательной динамики, наблюдаемой в активности мозга. В этой области используется множество моделей, каждая из которых определена на разном уровне абстракции и пытается смоделировать различные аспекты нейронных систем. Они варьируются от моделей кратковременного поведения отдельных нейронов до моделей того, как динамика нейронных цепей возникает в результате взаимодействия между отдельными нейронами, до моделей того, как поведение может возникать из абстрактных нейронных модулей, которые представляют собой целые подсистемы.
Модель одного нейрона
[ редактировать ]
Модель биологического нейрона — это математическое описание свойств нервных клеток или нейронов, предназначенное для точного описания и прогнозирования его биологических процессов. Одной из наиболее успешных моделей нейронов является модель Ходжкина-Хаксли, за которую Ходжкин и Хаксли получили в 1963 году Нобелевскую премию по физиологии и медицине. Модель основана на данных гигантского аксона кальмара и состоит из нелинейных дифференциальных уравнений, которые аппроксимируют электрические характеристики нейрона, включая генерацию и распространение потенциалов действия . Модель настолько успешно описывает эти характеристики, что вариации ее формулировки, основанной на проводимости, продолжают использоваться в моделях нейронов более полувека спустя. [44]
Модель Ходжкина-Хаксли слишком сложна для понимания с помощью классических математических методов, поэтому исследователи часто обращаются к упрощениям, таким как модель ФитцХью-Нагумо и модель Хиндмарша-Роуза , или к сильно идеализированным моделям нейронов, таким как дырявый нейрон, интегрирующий и запускающий. , первоначально разработанный Лапиком в 1907 году. [45] [46] Такие модели фиксируют только существенные динамики мембран, такие как всплески или разрывы, за счет биофизических деталей, но они более эффективны в вычислительном отношении, позволяя моделировать более крупные биологические нейронные сети .
Пиковая модель
[ редактировать ]Модель нейронной сети описывает популяцию физически связанных между собой нейронов или группу разрозненных нейронов, чьи входы или цели передачи сигналов определяют узнаваемую схему. Целью этих моделей является описание того, как динамика нейронных цепей возникает в результате взаимодействия между отдельными нейронами. Локальные взаимодействия между нейронами могут привести к синхронизации пиковой активности и составить основу колебательной активности. В частности, было показано, что модели взаимодействующих пирамидных клеток и тормозных интернейронов генерируют мозговые ритмы, такие как гамма-активность . [47] Аналогичным образом было показано, что моделирование нейронных сетей с феноменологической моделью сбоев реакции нейронов может предсказывать спонтанные широкополосные нейронные колебания. [48]
Модель нейронной массы
[ редактировать ]
Модели нейронного поля являются еще одним важным инструментом в изучении нейронных колебаний и представляют собой математическую основу, описывающую эволюцию таких переменных, как средняя скорострельность в пространстве и времени. При моделировании активности большого количества нейронов основная идея состоит в том, чтобы довести плотность нейронов до предела континуума , что приведет к созданию пространственно непрерывных нейронных сетей . Вместо моделирования отдельных нейронов этот подход аппроксимирует группу нейронов по ее средним свойствам и взаимодействиям. Он основан на подходе среднего поля — области статистической физики , которая занимается крупномасштабными системами. Модели, основанные на этих принципах, использовались для математического описания нейронных колебаний и ритмов ЭЭГ. Например, их использовали для исследования зрительных галлюцинаций. [50]
Модель Курамото
[ редактировать ]Модель Курамото связанных фазовых генераторов. [51] — одна из наиболее абстрактных и фундаментальных моделей, используемых для исследования нейронных колебаний и синхронизации. Он фиксирует активность локальной системы (например, отдельного нейрона или нейронного ансамбля) только по ее круговой фазе и, следовательно, игнорирует амплитуду колебаний (амплитуда постоянна). [52] Взаимодействия между этими осцилляторами представлены простой алгебраической формой (например, синусоидальной функцией) и вместе создают динамическую картину в глобальном масштабе.
Модель Курамото широко используется для изучения колебательной активности мозга, и было предложено несколько расширений, повышающих ее нейробиологическую достоверность, например, за счет включения топологических свойств локальных корковых связей. [53] В частности, оно описывает, как активность группы взаимодействующих нейронов может синхронизироваться и генерировать крупномасштабные колебания.
Моделирование с использованием модели Курамото с реалистичными кортикальными связями на большие расстояния и взаимодействиями с задержкой во времени выявило появление медленных шаблонных флуктуаций, которые воспроизводят BOLD функциональные карты состояния покоя, которые можно измерить с помощью фМРТ . [54]
Модели активности
[ редактировать ]Как отдельные нейроны, так и группы нейронов могут спонтанно генерировать колебательную активность. Кроме того, они могут демонстрировать колебательные реакции на перцептивные входные или двигательные сигналы. Некоторые типы нейронов будут ритмично активироваться при отсутствии какого-либо синаптического сигнала. Аналогичным образом, активность всего мозга обнаруживает колебательную активность, в то время как испытуемые не участвуют в какой-либо деятельности, так называемую активность в состоянии покоя . Эти постоянные ритмы могут меняться по-разному в ответ на перцептивные или двигательные воздействия. Колебательная активность может реагировать увеличением или уменьшением частоты и амплитуды или временным прерыванием, которое называется сбросом фазы. Кроме того, внешняя активность может вообще не взаимодействовать с текущей деятельностью, что приводит к аддитивной реакции.
 Частота . продолжающейся колебательной активности увеличивается между t1 и t2
Частота . продолжающейся колебательной активности увеличивается между t1 и t2 Амплитуда продолжающейся колебательной активности увеличивается между t1 и t2.
Амплитуда продолжающейся колебательной активности увеличивается между t1 и t2. Фаза продолжающейся колебательной активности сбрасывается в момент t1.
Фаза продолжающейся колебательной активности сбрасывается в момент t1. Активность линейно добавляется к продолжающейся колебательной активности между t1 и t2.
Активность линейно добавляется к продолжающейся колебательной активности между t1 и t2.




Текущая деятельность
[ редактировать ]Спонтанная активность — это активность мозга в отсутствие явной задачи, такой как сенсорная или двигательная активность, и поэтому ее также называют активностью в состоянии покоя. Она противоположна индуцированной активности, то есть активности мозга, вызываемой сенсорными стимулами или двигательными реакциями.
Термин продолжающаяся мозговая активность используется в электроэнцефалографии и магнитоэнцефалографии для тех компонентов сигнала, которые не связаны с обработкой раздражителя или возникновением других конкретных событий, например, с перемещением части тела, т.е. событий, не образующих вызванные потенциалы / вызванные поля , или индуцированная активность.
Спонтанная активность обычно считается шумом , если человек интересуется обработкой стимулов; однако считается, что спонтанная активность играет решающую роль в развитии мозга, например, в формировании сети и синаптогенезе. Спонтанная активность может быть информативной относительно текущего психического состояния человека (например, бодрствование, бдительность) и часто используется при исследованиях сна. Определенные виды колебательной активности, например альфа-волны , являются частью спонтанной активности. Статистический анализ колебаний мощности альфа-активности выявляет бимодальное распределение, т.е. высоко- и низкоамплитудный режим, и, следовательно, показывает, что активность в состоянии покоя не просто отражает шумовой процесс. [55]
В случае фМРТ спонтанные колебания сигнала , зависящего от уровня кислорода в крови (ЖИРНЫЙ), выявляют закономерности корреляции, которые связаны с сетями состояний покоя, такими как сеть по умолчанию . [56] Временная эволюция сетей состояний покоя коррелирует с колебаниями колебательной активности ЭЭГ в разных диапазонах частот. [57]
Текущая активность мозга также может играть важную роль в восприятии, поскольку она может взаимодействовать с деятельностью, связанной с поступающими стимулами. Действительно, исследования ЭЭГ показывают, что зрительное восприятие зависит как от фазы, так и от амплитуды корковых колебаний. Например, амплитуда и фаза альфа-активности в момент зрительной стимуляции предсказывают, будет ли субъект воспринимать слабый стимул. [58] [59] [60]
Частотная характеристика
[ редактировать ]В ответ на входной сигнал нейрон или ансамбль нейронов могут изменить частоту своих колебаний, тем самым изменяя скорость их всплесков. Часто частота срабатывания нейрона зависит от суммарной активности, которую он получает. Изменения частоты также часто наблюдаются в генераторах центральных паттернов и напрямую связаны со скоростью двигательной активности , например с частотой шагов при ходьбе. Однако изменения относительной частоты колебаний между различными областями мозга встречаются не так часто, поскольку частота колебательной активности часто связана с временными задержками между областями мозга.
Амплитудный отклик
[ редактировать ]Помимо вызванной активности, нейронная активность, связанная с обработкой стимулов, может привести к индуцированной активности. Индуцированная активность означает модуляцию текущей активности мозга, вызванную обработкой стимулов или подготовкой к движению. Следовательно, они отражают косвенный ответ в отличие от вызванных ответов. Хорошо изученным видом индуцированной активности является изменение амплитуды колебательной активности. Например, гамма-активность часто увеличивается во время повышенной умственной активности, например, во время представления объектов. [61] Поскольку индуцированные отклики могут иметь разные фазы в разных измерениях и, следовательно, будут компенсироваться во время усреднения, их можно получить только с помощью частотно-временного анализа . Индуцированная активность обычно отражает активность многочисленных нейронов: считается, что изменения амплитуды колебательной активности возникают в результате синхронизации нейронной активности, например, за счет синхронизации времени спайков или колебаний мембранного потенциала отдельных нейронов. Поэтому увеличение колебательной активности часто называют событийной синхронизацией, а снижение — событийной десинхронизацией. [62]
Сброс фазы
[ редактировать ]Сброс фазы происходит, когда входные данные нейрона или ансамбля нейронов сбрасывают фазу продолжающихся колебаний. [63] Это очень распространено в одиночных нейронах, где время импульсов регулируется в зависимости от входного сигнала нейрона (нейрон может генерировать импульсы с фиксированной задержкой в ответ на периодический входной сигнал, что называется фазовой синхронизацией). [19] ), а также может возникать в нейронных ансамблях, когда фазы их нейронов регулируются одновременно. Сброс фазы имеет основополагающее значение для синхронизации разных нейронов или разных областей мозга. [18] [38] потому что время спайков может быть привязано к активности других нейронов.
Сброс фазы также позволяет изучать вызванную активность - термин, используемый в электроэнцефалографии и магнитоэнцефалографии для обозначения реакций активности мозга, которые напрямую связаны с активностью, связанной со стимулом . Вызванные потенциалы и потенциалы, связанные с событием, получают из электроэнцефалограммы путем усреднения с привязкой к стимулу, т. е. усреднения различных проб с фиксированными задержками вокруг предъявления стимула. Как следствие, те компоненты сигнала, которые одинаковы в каждом отдельном измерении, сохраняются, а все остальные, то есть текущая или спонтанная активность, усредняются. То есть потенциалы, связанные с событием, отражают только колебания активности мозга, синхронизированные по фазе со стимулом или событием. Вызванная активность часто считается независимой от текущей мозговой активности, хотя этот вопрос является предметом постоянных дискуссий. [64] [65]
Асимметричная амплитудная модуляция
[ редактировать ]Недавно было высказано предположение, что даже если фазы в разных исследованиях не совпадают, индуцированная активность все равно может вызывать потенциалы, связанные с событиями , поскольку текущие колебания мозга могут быть несимметричными, и, таким образом, амплитудные модуляции могут привести к сдвигу базовой линии, который не усредняется. [66] [67] Эта модель подразумевает, что медленные реакции, связанные с событиями, такие как асимметричная альфа-активность, могут быть результатом асимметричных модуляций амплитуды колебаний мозга, таких как асимметрия внутриклеточных токов, которые распространяются вперед и назад по дендритам. [68] Согласно этому предположению, асимметрия дендритного тока может вызвать асимметрию колебательной активности, измеряемой с помощью ЭЭГ и МЭГ, поскольку обычно считается, что дендритные токи в пирамидных клетках генерируют сигналы ЭЭГ и МЭГ, которые можно измерить на коже головы. [69]
Межчастотная связь
[ редактировать ]Межчастотная связь (CFC) описывает связь (статистическую корреляцию) между медленной и быстрой волной. Существует много видов, обычно обозначаемых как связь AB, что означает, что A медленной волны соединяется с B быстрой волны. Например, фазово-амплитудная связь - это когда фаза медленной волны связана с амплитудой быстрой волны. [70]
Тета -гамма-код представляет собой связь между тета-волной и гамма-волной в сети гиппокампа. Во время тета-волны последовательно активируются от 4 до 8 непересекающихся ансамблей нейронов. Было высказано предположение, что это формирует нейронный код, представляющий несколько элементов во временном кадре. [71] [72]
Функция
[ редактировать ]Нейронная синхронизация может модулироваться ограничениями задачи, такими как внимание , и считается, что она играет роль в привязке функций . [73] нейронная связь , [7] и координация движений . [9] Нейрональные колебания стали горячей темой в нейробиологии в 1990-х годах, когда исследования зрительной системы мозга Грея, Сингера и других подтвердили гипотезу нейронных связей . [74] Согласно этой идее, синхронные колебания в ансамблях нейронов связывают нейроны, представляющие разные признаки объекта. Например, когда человек смотрит на дерево, нейроны зрительной коры, представляющие ствол дерева, и нейроны, представляющие ветви того же дерева, будут колебаться синхронно, образуя единое представление дерева. Это явление лучше всего видно на потенциалах локального поля , которые отражают синхронную активность локальных групп нейронов, но оно также было показано на записях ЭЭГ и МЭГ, что дает все больше доказательств тесной связи между синхронной колебательной активностью и различными когнитивными функциями, такими как перцептивная активность. группировка [73] и контроль внимания сверху вниз. [15] [13] [12]
Кардиостимулятор
[ редактировать ]Клетки синоатриального узла , расположенного в правом предсердии сердца, спонтанно деполяризуются примерно 100 раз в минуту. Хотя все клетки сердца обладают способностью генерировать потенциалы действия, вызывающие сокращение сердца, обычно его инициирует синоатриальный узел просто потому, что он генерирует импульсы немного быстрее, чем другие области. Следовательно, эти клетки генерируют нормальный синусовый ритм и называются клетками-водителями ритма, поскольку они напрямую контролируют частоту сердечных сокращений . При отсутствии внешнего нервного и гормонального контроля клетки в СА-узле будут ритмично разряжаться. Синоатриальный узел обильно иннервируется вегетативной нервной системой , которая регулирует частоту спонтанных импульсов пейсмекерных клеток.
Центральный генератор шаблонов
[ редактировать ]Синхронная активация нейронов также составляет основу периодических двигательных команд на ритмические движения. Эти ритмические сигналы производятся группой взаимодействующих нейронов, образующих сеть, называемую центральным генератором паттернов . Генераторы центральных паттернов — это нейронные цепи, которые при активации могут создавать ритмические двигательные паттерны при отсутствии сенсорных или нисходящих входных сигналов, несущих конкретную временную информацию. Примерами являются ходьба , дыхание и плавание . [75] Большинство доказательств существования центральных генераторов паттернов получено от низших животных, таких как минога , но есть также доказательства существования спинальных генераторов центральных паттернов у людей. [76] [77]
Обработка информации
[ редактировать ]Нейрональные спайки обычно считаются основой передачи информации в мозгу. Для такой передачи информация должна быть закодирована в виде импульсов. Были предложены различные типы схем кодирования, такие как скоростное кодирование и временное кодирование . Нейронные колебания могут создавать периодические временные окна, в которых входные импульсы оказывают большее влияние на нейроны, тем самым обеспечивая механизм декодирования временных кодов. [78]
Временное декодирование
[ редактировать ]Одноклеточные собственные генераторы служат ценным инструментом для декодирования временно закодированной сенсорной информации. Эта информация кодируется через интервалы между спайками, и внутренние генераторы могут действовать как «временные линейки» для точного измерения этих интервалов. Одним из примечательных механизмов достижения этого является нейронная фазовая автоподстройка частоты (NPLL). В этом механизме корковые осцилляторы подвергаются модуляции под влиянием частоты импульсов таламокортикальных «детекторов фазы», которые, в свою очередь, измеряют несоответствие между корковой и сенсорной периодичностью. [79]
Восприятие
[ редактировать ]Синхронизация возбуждения нейронов может служить средством группировки пространственно разделенных нейронов, которые реагируют на один и тот же стимул, чтобы связать эти ответы для дальнейшей совместной обработки, т.е. использовать временную синхронность для кодирования отношений. Сначала были предложены чисто теоретические формулировки гипотезы связывания синхронностью. [80] но впоследствии были получены обширные экспериментальные данные, подтверждающие потенциальную роль синхронности как реляционного кода. [81]
Функциональная роль синхронизированной колебательной активности головного мозга была установлена главным образом в экспериментах, проведенных на бодрствующих котятах с несколькими имплантированными в зрительную кору электродами. Эти эксперименты показали, что группы пространственно разделенных нейронов участвуют в синхронной колебательной активности при активации зрительными стимулами. Частота этих колебаний находилась в диапазоне 40 Гц и отличалась от периодической активации, индуцированной решеткой, что позволяет предположить, что колебания и их синхронизация обусловлены внутренними нейрональными взаимодействиями. [81] Аналогичные результаты были параллельно продемонстрированы группой Экхорна, что предоставило дополнительные доказательства функциональной роли нейронной синхронизации в связывании признаков. [82] С тех пор многочисленные исследования повторили эти результаты и распространили их на различные методы, такие как ЭЭГ, предоставив обширные доказательства функциональной роли гамма -колебаний в зрительном восприятии.
Жиль Лоран и его коллеги показали, что колебательная синхронизация играет важную функциональную роль в восприятии запахов. Восприятие разных запахов приводит к тому, что разные подмножества нейронов активируются в разных наборах колебательных циклов. [83] Эти колебания могут быть нарушены ГАМК блокатором пикротоксином . [84] а нарушение колебательной синхронизации приводит к ухудшению поведенческой дискриминации химически близких запахов у пчел, [85] и к более схожим реакциям на запахи в нижестоящих нейронах β-доли. [86] Недавнее продолжение этой работы показало, что колебания создают периодические окна интеграции для клеток Кеньона в теле гриба насекомого , так что входящие шипы из усиков более эффективны в активации клеток Кеньона только на определенных фазах колебательного цикла. [78]
Считается также, что нейронные колебания связаны с чувством времени. [87] и в соматосенсорном восприятии. [88] Однако недавние результаты свидетельствуют против часовой функции кортикальных гамма-колебаний. [89]
Координация движений
[ редактировать ]В двигательной системе часто наблюдаются колебания. Пфурчеллер и его коллеги обнаружили снижение альфа (8–12 Гц) и бета (13–30 Гц) колебаний в активности ЭЭГ , когда испытуемые совершали движение. [62] [90] Используя внутрикорковые записи, аналогичные изменения колебательной активности были обнаружены в моторной коре, когда обезьяны выполняли двигательные действия, требующие значительного внимания. [91] [92] Кроме того, колебания на уровне позвоночника синхронизируются с бета-колебаниями в моторной коре во время постоянной мышечной активации, что определяется кортико-мышечной когерентностью . [93] [94] [95] Аналогичным образом, мышечная активность различных мышц демонстрирует межмышечную когерентность на нескольких различных частотах, отражающих основные нейронные схемы, участвующие в координации движений . [96] [97]
Недавно было обнаружено, что корковые колебания распространяются в виде бегущих волн по поверхности моторной коры вдоль доминирующих пространственных осей, характерных для локальной схемы моторной коры. [98] Было высказано предположение, что двигательные команды в форме бегущих волн могут пространственно фильтроваться нисходящими волокнами для избирательного управления мышечной силой. [99] Моделирование показало, что продолжающаяся волновая активность в коре головного мозга может вызвать устойчивую мышечную силу с физиологическим уровнем когерентности ЭЭГ-ЭМГ. [100]
Колебательные ритмы частотой 10 Гц были зарегистрированы в области мозга, называемой нижней оливой , которая связана с мозжечком. [20] Эти колебания наблюдаются также в двигательной активности физиологического тремора. [101] и при выполнении медленных движений пальцами. [102] Эти результаты могут указывать на то, что человеческий мозг периодически контролирует непрерывные движения. В подтверждение было показано, что эти прерывания движений напрямую коррелируют с колебательной активностью в мозжечково-таламо-кортикальной петле, которая может представлять собой нейронный механизм прерывистого моторного контроля. [103]
Память
[ редактировать ]Нейронные колебания, в частности тета- активность, во многом связаны с функцией памяти. Тета-ритмы очень сильны в гиппокампе и энторинальной коре грызунов во время обучения и восстановления памяти, и считается, что они жизненно важны для индукции долговременной потенциации — потенциального клеточного механизма обучения и памяти. связь между тета- и гамма- Считается, что активностью жизненно важна для функций памяти, включая эпизодическую память . [104] [105] Тесная координация всплесков одиночных нейронов с локальными тета-колебаниями связана с успешным формированием памяти у людей, поскольку более стереотипные всплески предсказывают лучшую память. [106]
Сон и сознание
[ редактировать ]Сон — это естественно повторяющееся состояние, характеризующееся снижением или отсутствием сознания и происходящее в циклах сна с быстрым движением глаз (REM) и с медленным движением глаз сна (NREM). Стадии сна характеризуются спектральным составом ЭЭГ : например, стадия N1 относится к переходу мозга от альфа-волн (обычных в состоянии бодрствования) к тета-волнам, тогда как стадия N3 (глубокий или медленноволновой сон) характеризуется наличие дельта-волн. [107] Нормальный порядок стадий сна: N1 → N2 → N3 → N2 → REM. [ нужна ссылка ]
Разработка
[ редактировать ]Нейронные колебания могут играть роль в развитии нервной системы. Например, считается, что волны сетчатки обладают свойствами, которые определяют раннюю связь цепей и синапсов между клетками сетчатки. [108]
Патология
[ редактировать ]

Определенные типы нервных колебаний могут возникать и при патологических ситуациях, например, при болезни Паркинсона или эпилепсии . Эти патологические колебания часто представляют собой аберрантную версию нормальных колебаний. Например, одним из наиболее известных типов являются спайк-волновые колебания, типичные для генерализованных или абсансных эпилептических припадков и напоминающие нормальные колебания веретена сна.
Тремор
[ редактировать ]Тремор – это непроизвольное, несколько ритмичное сокращение и расслабление мышц, включающее возвратно-поступательные движения одной или нескольких частей тела. Это наиболее распространенное из всех непроизвольных движений, которое может поражать кисти, руки, глаза, лицо, голову, голосовые связки, туловище и ноги. Чаще всего тремор возникает в руках. У некоторых людей тремор является симптомом другого неврологического расстройства. Было идентифицировано множество различных форм тремора, таких как эссенциальный тремор или паркинсонический тремор. Утверждается, что тремор, вероятно, имеет многофакторное происхождение, на него влияют нейронные колебания в центральной нервной системе, а также периферические механизмы, такие как резонансы рефлекторных петель. [109]
Эпилепсия
[ редактировать ]Эпилепсия – распространенное хроническое неврологическое заболевание, характеризующееся судорогами . Эти припадки являются временными признаками и/или симптомами аномальной, чрезмерной или гиперсинхронной активности нейронов головного мозга. [110]
Таламокортикальная аритмия
[ редактировать ]При таламокортикальной аритмии (ТКД) нормальный таламокортикальный резонанс нарушается . Таламическая потеря входного сигнала позволяет частоте таламо-кортикального столба снижаться до тета- или дельта-диапазона, что определяется МЭГ и ЭЭГ с помощью машинного обучения. [111] TCD можно лечить нейрохирургическими методами, такими как таламотомия .
Приложения
[ редактировать ]Клинические конечные точки
[ редактировать ]Нейронные колебания чувствительны к нескольким препаратам, влияющим на активность мозга; соответственно, биомаркеры, основанные на нейронных колебаниях, становятся вторичными конечными точками в клинических испытаниях и при количественной оценке эффектов в доклинических исследованиях. Эти биомаркеры часто называют «биомаркерами ЭЭГ» или «нейрофизиологическими биомаркерами», и их количественную оценку проводят с помощью количественной электроэнцефалографии (кЭЭГ) . Биомаркеры ЭЭГ можно извлечь из ЭЭГ с помощью набора инструментов нейрофизиологических биомаркеров с открытым исходным кодом .
Интерфейс мозг-компьютер
[ редактировать ]Нейронные колебания применялись в качестве управляющего сигнала в различных интерфейсах мозг-компьютер (BCI). [112] Например, можно создать неинвазивный BCI, поместив электроды на кожу головы и затем измерив слабые электрические сигналы. Хотя активность отдельных нейронов не может быть записана с помощью неинвазивного ИМК, поскольку череп демпфирует и размывает электромагнитные сигналы, колебательную активность все же можно надежно обнаружить. BCI был представлен Видалем в 1973 году. [113] как проблема использования сигналов ЭЭГ для управления объектами вне человеческого тела.
После испытания BCI в 1988 году альфа-ритм использовался в BCI на основе ритма мозга для управления физическим объектом, роботом. [114] [115] BCI на основе альфа-ритма был первым BCI для управления роботом. [116] [117] В частности, некоторые формы BCI позволяют пользователям управлять устройством путем измерения амплитуды колебательной активности в определенных диапазонах частот, включая мю- и бета -ритмы.
Примеры
[ редактировать ]Неполный список типов колебательной активности, встречающихся в центральной нервной системе:
- Альфа-волна
- Бета-волна
- взрыв
- Сердечный цикл
- Дельта-волна
- Эпилептический припадок
- Гамма-волна
- Математическое моделирование электрофизиологической активности при эпилепсии
- Му волна
- волны ПГО
- Таламокортикальные колебания
- Комплексы острая волна–рябь
- Спящий шпиндель
- Подпороговые колебания мембранного потенциала
- Тета-волна
См. также
[ редактировать ]- Кибернетика
- Теория динамических систем
- ЭЭГ-анализ
- нейрокибернетика
- Колебательная нейронная сеть
- Системная нейробиология
- Тета-хилинг
- Сброс фазы в нейронах
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Наполи, Николас Дж.; Демас, Мэтью; Стивенс, Чад Л.; Кеннеди, Келли Д.; Харривел, Анджела Р.; Барнс, Лаура Э.; Поуп, Алан Т. (03 марта 2020 г.). «Сложность активации: инструмент когнитивных нарушений для характеристики нейроизоляции» . Научные отчеты . 10 (1): 3909. Бибкод : 2020NatSR..10.3909N . дои : 10.1038/s41598-020-60354-2 . ISSN 2045-2322 . ПМК 7054256 . ПМИД 32127579 .
 В эту статью включен текст из этого источника, доступного по лицензии CC BY 4.0 .
В эту статью включен текст из этого источника, доступного по лицензии CC BY 4.0 . - ^ Перейти обратно: а б с Ахиссар Э. и Ваадиа Э. Колебательная активность отдельных единиц соматосенсорной коры бодрствующей обезьяны и их возможная роль в анализе текстуры. Proc Natl Acad Sci USA 87, 8935-8939 (1990).
- ^ Ллинас Р.Р. (2014). «Внутренние электрические свойства нейронов млекопитающих и функции ЦНС: историческая перспектива» . Границы клеточной нейронауки . 8 : 320. дои : 10.3389/fncel.2014.00320 . ПМК 4219458 . ПМИД 25408634 .
- ^ «Катон, Ричард - Электрические токи мозга» . echo.mpiwg-berlin.mpg.de . Проверено 21 декабря 2018 г.
- ^ Коэнен А., Файн Е., Заячковская О. (2014). «Адольф Бек: забытый пионер электроэнцефалографии». Журнал истории нейронаук . 23 (3): 276–86. дои : 10.1080/0964704x.2013.867600 . ПМИД 24735457 . S2CID 205664545 .
- ^ Правдич-Неминский В.В. (1913). «Попытка зарегистрировать электрические явления мозга». Центральный журнал физиологии . 27 :951–60.
- ^ Перейти обратно: а б Фрис П. (октябрь 2005 г.). «Механизм когнитивной динамики: нейронная связь через когерентность нейронов». Тенденции в когнитивных науках . 9 (10): 474–80. дои : 10.1016/j.tics.2005.08.011 . ПМИД 16150631 . S2CID 6275292 .
- ^ Фелл Дж., Аксмахер Н. (февраль 2011 г.). «Роль фазовой синхронизации в процессах памяти». Обзоры природы. Нейронаука . 12 (2): 105–18. дои : 10.1038/nrn2979 . ПМИД 21248789 . S2CID 7422401 .
- ^ Перейти обратно: а б Шницлер А., Гросс Дж. (апрель 2005 г.). «Нормальная и патологическая колебательная связь в мозге». Обзоры природы. Нейронаука . 6 (4): 285–96. дои : 10.1038/nrn1650 . ПМИД 15803160 . S2CID 2749709 .
- ^ Балдауф Д., Дезимона Р. (апрель 2014 г.). «Нейронные механизмы объектного внимания» . Наука . 344 (6182): 424–7. Бибкод : 2014Sci...344..424B . дои : 10.1126/science.1247003 . ПМИД 24763592 . S2CID 34728448 .
- ^ Фостер Дж. Дж., Саттерер Д. В., Серенсес Дж. Т., Фогель Е. К., Ау Э (июль 2017 г.). «Колебания альфа-диапазона позволяют отслеживать скрытое пространственное внимание с пространственным и временным разрешением» . Психологическая наука . 28 (7): 929–941. дои : 10.1177/0956797617699167 . ПМЦ 5675530 . ПМИД 28537480 .
- ^ Перейти обратно: а б де Врис И.Е., Маринато Г., Балдауф Д. (август 2021 г.). «Декодирование объектно-ориентированного слухового внимания по альфа-колебаниям МЭГ, восстановленным по источнику» . Журнал неврологии . 41 (41): 8603–8617. doi : 10.1523/JNEUROSCI.0583-21.2021 . ПМЦ 8513695 . ПМИД 34429378 .
- ^ Перейти обратно: а б с Багерзаде И., Балдауф Д., Пантазис Д., Дезимона Р. (февраль 2020 г.). «Альфа-синхрония и нейробиоуправление пространственным вниманием» . Нейрон . 105 (3): 577–587.e5. дои : 10.1016/j.neuron.2019.11.001 . hdl : 11572/252726 . ПМИД 31812515 . S2CID 208614924 .
- ^ Бергер Х, Грей СМ (1929). «Об электроэнцефалограмме человека». Главный психиатр Невропатия 87 :527-570. дои : 10.1007/BF01797193 . hdl : 11858/00-001M-0000-002A-5DE0-7 . S2CID 10835361 .
- ^ Перейти обратно: а б с Балдауф Д., Дезимона Р. (апрель 2014 г.). «Нейронные механизмы объектного внимания» . Наука . 344 (6182): 424–7. Бибкод : 2014Sci...344..424B . дои : 10.1126/science.1247003 . ПМИД 24763592 . S2CID 34728448 .
- ^ Демент В., Клейтман Н. (ноябрь 1957 г.). «Циклические изменения ЭЭГ во время сна и их связь с движениями глаз, подвижностью тела и сновидениями». Электроэнцефалография и клиническая нейрофизиология . 9 (4): 673–90. дои : 10.1016/0013-4694(57)90088-3 . ПМИД 13480240 .
- ^ Энгель А.К., Сингер В. (январь 2001 г.). «Временная привязка и нейронные корреляты сенсорного осознания». Тенденции в когнитивных науках . 5 (1): 16–25. дои : 10.1016/S1364-6613(00)01568-0 . ПМИД 11164732 . S2CID 11922975 .
- ^ Перейти обратно: а б с Варела Ф., Лашо Дж.П., Родригес Э., Мартинери Дж. (апрель 2001 г.). «Мозговая сеть: фазовая синхронизация и крупномасштабная интеграция». Обзоры природы. Нейронаука . 2 (4): 229–39. дои : 10.1038/35067550 . ПМИД 11283746 . S2CID 18651043 .
- ^ Перейти обратно: а б с Ижикевич Э.М. (2007). Динамические системы в нейробиологии . Кембридж, Массачусетс: MIT Press.
- ^ Перейти обратно: а б Ллинас Р., Яром Ю. (июль 1986 г.). «Осцилляционные свойства нейронов нижней оливы морской свинки и их фармакологическая модуляция: исследование in vitro» . Журнал физиологии . 376 : 163–82. дои : 10.1113/jphysicalol.1986.sp016147 . ПМЦ 1182792 . ПМИД 3795074 .
- ^ Мурешан Р.К., Юрьюц О.Ф., Мока В.В., Сингер В., Николич Д. (март 2008 г.). «Оценка колебаний: эффективный метод оценки силы колебаний нейрональной активности». Журнал нейрофизиологии . 99 (3): 1333–53. дои : 10.1152/jn.00772.2007 . ПМИД 18160427 .
- ^ Берроу Т. (1943). «Нейродинамика поведения. Филобиологическое предисловие». Философия науки . 10 (4): 271–288. дои : 10.1086/286819 . S2CID 121438105 .
- ^ Амари, Сюн-ичи (1 июня 1977 г.). «Динамика формирования паттернов в нейронных полях латерального тормозного типа» . Биологическая кибернетика . 27 (2): 77–87. дои : 10.1007/BF00337259 . ISSN 1432-0770 . ПМИД 911931 . S2CID 2811608 .
- ^ Шёнер, Г.; Келсо, JAS (25 марта 1988 г.). «Генерация динамических паттернов в поведенческих и нейронных системах» . Наука . 239 (4847): 1513–1520. Бибкод : 1988Sci...239.1513S . дои : 10.1126/science.3281253 . ISSN 0036-8075 . ПМИД 3281253 .
- ^ Ванстинзель М.Дж., Пелс Э.Г., Бляйхнер М.Г., Бранко М.П., Денисон Т., Фрейденбург З.В. и др. (ноябрь 2016 г.). «Полностью имплантированный интерфейс мозг-компьютер у изолированного пациента с БАС» . Медицинский журнал Новой Англии . 375 (21): 2060–2066. дои : 10.1056/NEJMoa1608085 . hdl : 1874/344360 . ПМК 5326682 . ПМИД 27959736 .
- ^ Перейти обратно: а б Хакен Х (1996). Принципы работы мозга . Спрингер. ISBN 978-3-540-58967-9 .
- ^ Перейти обратно: а б с Ван XJ (июль 2010 г.). «Нейрофизиологические и вычислительные принципы корковых ритмов в познании» . Физиологические обзоры . 90 (3): 1195–268. doi : 10.1152/physrev.00035.2008 . ПМЦ 2923921 . ПМИД 20664082 .
- ^ Нуньес П.Л., Сринивасан Р. (1981). Электрические поля мозга: Нейрофизика ЭЭГ . Издательство Оксфордского университета. ISBN 9780195027969 .
- ^ Карден Дж.А., Карлен М., Мелетис К., Кноблих У., Чжан Ф., Дейссерот К. и др. (июнь 2009 г.). «Управление быстродействующими клетками индуцирует гамма-ритм и контролирует сенсорные реакции» . Природа . 459 (7247): 663–7. Бибкод : 2009Natur.459..663C . дои : 10.1038/nature08002 . ПМЦ 3655711 . ПМИД 19396156 .
- ^ Ллинас Р., Рибари У., Контрерас Д., Педроарена К. (ноябрь 1998 г.). «Нейронная основа сознания» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 353 (1377): 1841–9. дои : 10.1098/rstb.1998.0336 . ПМЦ 1692417 . ПМИД 9854256 .
- ^ Боллимунта А., Мо Дж., Шредер С.Э., Дин М. (март 2011 г.). «Нейрональные механизмы и модуляция внимания кортикоталамических α-колебаний» . Журнал неврологии . 31 (13): 4935–43. doi : 10.1523/JNEUROSCI.5580-10.2011 . ПМК 3505610 . ПМИД 21451032 .
- ^ Суффчинский П., Калицин С., Пфурчеллер Г., Лопес да Силва Ф.Х. (декабрь 2001 г.). «Вычислительная модель таламо-кортикальных сетей: динамический контроль альфа-ритмов в зависимости от фокусного внимания». Международный журнал психофизиологии . 43 (1): 25–40. дои : 10.1016/S0167-8760(01)00177-5 . ПМИД 11742683 .
- ^ Кабрал Дж., Лакху Х., Вулрич М., Йоенссон М., Мохсени Х., Бейкер А. и др. (апрель 2014 г.). «Изучение механизмов спонтанной функциональной связности в МЭГ: как задержанные сетевые взаимодействия приводят к структурированным амплитудным огибающим колебаний с полосовой фильтрацией» . НейроИмидж . 90 : 423–35. doi : 10.1016/j.neuroimage.2013.11.047 . hdl : 10230/23081 . ПМИД 24321555 .
- ^ Ллинас Р.Р. (декабрь 1988 г.). «Внутренние электрофизиологические свойства нейронов млекопитающих: понимание функции центральной нервной системы». Наука . 242 (4886): 1654–64. Бибкод : 1988Sci...242.1654L . дои : 10.1126/science.3059497 . ПМИД 3059497 .
- ^ Ллинас Р.Р., Грейс А.А., Яром Ю. (февраль 1991 г.). «In vitro нейроны 4-го слоя коры млекопитающих проявляют внутреннюю колебательную активность в диапазоне частот от 10 до 50 Гц» . Труды Национальной академии наук Соединенных Штатов Америки . 88 (3): 897–901. Бибкод : 1991PNAS...88..897L . дои : 10.1073/pnas.88.3.897 . ПМК 50921 . ПМИД 1992481 .
- ^ Ахиссар Э., Хайдарлиу С. и Заксенхаус М. (1997). Декодирование временно закодированной сенсорной информации с помощью корковых колебаний и компараторов таламических фаз. Proc Natl Acad Sci USA 94, 11633-11638.
- ^ Цейтлер М., Даффертсхофер А., Гилен CC (июнь 2009 г.). «Асимметрия в импульсных генераторах с задержкой» (PDF) . Физический обзор E . 79 (6 Pt 2): 065203. Бибкод : 2009PhRvE..79f5203Z . дои : 10.1103/PhysRevE.79.065203 . hdl : 1871/29169 . ПМИД 19658549 .
- ^ Перейти обратно: а б Пиковский А., Розенблюм М., Куртс Дж. (2001). Синхронизация: универсальное понятие в нелинейных науках . Издательство Кембриджского университета. ISBN 978-0-521-53352-2 .
- ^ Андреа Бровелли, Стивен Л. Бресслер и их коллеги, 2004 г.
- ^ Питер, Берки; Чаба, Череп; Жужанна, Кёрней (2024). «Микроглия способствует нейрональной синхронизации, несмотря на фенотипическую трансформацию, связанную с эндогенной АТФ, в острых срезах мозга мышей» . Природные коммуникации . дои : 10.1038/s41467-024-49773-1 . ПМЦ 11208608 . ПМИД 38926390 .
- ^ Мутукумарасвами С.Д., Эдден Р.А., Джонс Д.К., Светтенхэм Дж.Б., Сингх К.Д. (май 2009 г.). «Концентрация ГАМК в состоянии покоя предсказывает пиковую частоту гамма-излучения и амплитуду фМРТ в ответ на зрительную стимуляцию у людей» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (20): 8356–61. Бибкод : 2009PNAS..106.8356M . дои : 10.1073/pnas.0900728106 . ПМЦ 2688873 . ПМИД 19416820 .
- ^ Моруцци Дж., Магун Х.В. (ноябрь 1949 г.). «Ретикулярная формация ствола мозга и активация ЭЭГ» . Электроэнцефалография и клиническая нейрофизиология . 1 (4): 455–73. дои : 10.1016/0013-4694(49)90219-9 . ПМИД 18421835 .
- ^ Бужаки Г., Драгун А. (июнь 2004 г.). «Нейрональные колебания в корковых сетях». Наука . 304 (5679): 1926–9. Бибкод : 2004Sci...304.1926B . дои : 10.1126/science.1099745 . ПМИД 15218136 . S2CID 8002293 .
- ^ Каттералл, Вашингтон, Раман, И.М., Робинсон, HPC, Сейновски, Т.Дж., Полсен, О. (2 октября 2012 г.). «Наследие Ходжкина-Хаксли: от каналов к схемам» . Журнал неврологии . 32 (41): 14064–14073. doi : 10.1523/JNEUROSCI.3403-12.2012 . ПМК 3500626 . ПМИД 23055474 .
- ^ Лапик, LM (1907). «Количественные исследования электрического возбуждения нервов». J Физиол Париж . 9 : 620–635.
- ^ Эбботт, Ларри (1999). «Представление Лапиком модели нейрона, объединяющей и запускающей (1907)». Бюллетень исследований мозга . 50 (5): 303–304. дои : 10.1016/S0361-9230(99)00161-6 . ПМИД 10643408 . S2CID 46170924 .
- ^ Уиттингтон М.А., Трауб Р.Д., Копелл Н., Эрментраут Б., Буль Э.Х. (декабрь 2000 г.). «Ритмы, основанные на торможении: экспериментальные и математические наблюдения за сетевой динамикой». Международный журнал психофизиологии . 38 (3): 315–36. CiteSeerX 10.1.1.16.6410 . дои : 10.1016/S0167-8760(00)00173-2 . ПМИД 11102670 .
- ^ Голденталь А., Варди Р., Сарди С., Сабо П., Кантер И. (2015). «Широкополосные макроскопические кортикальные колебания возникают в результате внутренних нарушений реакции нейронов» . Границы в нейронных цепях . 9 : 65.arXiv : 1511.00235 . дои : 10.3389/fncir.2015.00065 . ПМЦ 4626558 . ПМИД 26578893 .
- ^ Вендлинг Ф., Беллангер Дж. Дж., Бартоломей Ф., Човель П. (октябрь 2000 г.). «Актуальность нелинейных моделей с сосредоточенными параметрами для анализа эпилептических сигналов глубинной ЭЭГ». Биологическая кибернетика . 83 (4): 367–78. дои : 10.1007/s004220000160 . ПМИД 11039701 . S2CID 8751526 .
- ^ Бресслофф ПК, Коуэн Дж.Д. (2003)Спонтанное формирование паттернов в первичной зрительной коре. В: Дж. Хоган, А.Р. Краускопф, М. ди Бернадо, Р.Э. Уилсон (ред.), Нелинейная динамика и хаос: куда нам идти дальше?
- ^ Курамото Ю. (1984). Химические колебания, волны и турбулентность . Дуврские публикации.
- ^ Эрментроут Б (1994). «Введение в нейронные осцилляторы». В Ф. Вентриглии (ред.). Нейронное моделирование и нейронные сети . стр. 79–110.
- ^ Брейкспир М., Хайтманн С., Даффертсхофер А. (2010). «Генеративные модели корковых колебаний: нейробиологические последствия модели Курамото» . Границы человеческой неврологии . 4 : 190. дои : 10.3389/fnhum.2010.00190 . ПМЦ 2995481 . ПМИД 21151358 .
- ^ Кабрал Дж., Хьюг Э., Спорнс О., Деку Дж. (июль 2011 г.). «Роль колебаний локальной сети в функциональной связности в состоянии покоя» . НейроИмидж . 57 (1): 130–139. doi : 10.1016/j.neuroimage.2011.04.010 . ПМИД 21511044 . S2CID 13959959 .
- ^ Фрейер Ф., Акино К., Робинсон П.А., Риттер П., Брейкспир М. (июль 2009 г.). «Бистабильность и негауссовы колебания спонтанной корковой активности» . Журнал неврологии . 29 (26): 8512–24. doi : 10.1523/JNEUROSCI.0754-09.2009 . ПМК 6665653 . ПМИД 19571142 .
- ^ Фокс, доктор медицинских наук, Рэйкл М.Е. (сентябрь 2007 г.). «Спонтанные колебания активности мозга, наблюдаемые с помощью функциональной магнитно-резонансной томографии». Обзоры природы. Нейронаука . 8 (9): 700–11. дои : 10.1038/nrn2201 . ПМИД 17704812 . S2CID 15979590 .
- ^ Лауфс Х., Краков К., Стерцер П., Эгер Э., Байерле А., Салек-Хаддади А., Кляйншмидт А. (сентябрь 2003 г.). «Электроэнцефалографические признаки режимов внимания и когнитивных дефолтов при спонтанных колебаниях активности мозга в состоянии покоя» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (19): 11053–8. Бибкод : 2003PNAS..10011053L . дои : 10.1073/pnas.1831638100 . ЧВК 196925 . ПМИД 12958209 .
- ^ Мэтьюсон К.Э., Граттон Дж., Фабиани М., Бек Д.М., Ро Т. (март 2009 г.). «Видеть или не видеть: альфа-фаза предстимула предсказывает зрительную осведомленность» . Журнал неврологии . 29 (9): 2725–32. doi : 10.1523/JNEUROSCI.3963-08.2009 . ПМЦ 2724892 . ПМИД 19261866 .
- ^ Буш Н.А., Дюбуа Дж., ВанРуллен Р. (июнь 2009 г.). «Фаза продолжающихся колебаний ЭЭГ предсказывает зрительное восприятие» . Журнал неврологии . 29 (24): 7869–76. doi : 10.1523/jneurosci.0113-09.2009 . ПМЦ 6665641 . ПМИД 19535598 .
- ^ ван Дейк Х., Шоффелен Дж. М., Остенвельд Р., Йенсен О. (февраль 2008 г.). «Престимульная колебательная активность в альфа-диапазоне предсказывает способность к визуальному различению» . Журнал неврологии . 28 (8): 1816–23. doi : 10.1523/jneurosci.1853-07.2008 . ПМК 6671447 . ПМИД 18287498 .
- ^ Таллон-Бодри С., Бертран О. (апрель 1999 г.). «Колебательная гамма-активность у человека и ее роль в представлении объектов». Тенденции в когнитивных науках . 3 (4): 151–162. дои : 10.1016/S1364-6613(99)01299-1 . ПМИД 10322469 . S2CID 1308261 .
- ^ Перейти обратно: а б Пфурчеллер Г., Лопес да Силва Ф.Х. (ноябрь 1999 г.). «Синхронизация и десинхронизация ЭЭГ/МЭГ, связанная с событиями: основные принципы». Клиническая нейрофизиология . 110 (11): 1842–57. дои : 10.1016/S1388-2457(99)00141-8 . ПМИД 10576479 . S2CID 24756702 .
- ^ ТАСС ПА (2007). Фазовый сброс в медицине и биологии: стохастическое моделирование и анализ данных . Берлин Гейдельберг: Springer-Verlag. ISBN 978-3-540-65697-5 .
- ^ Макейг С., Вестерфилд М., Юнг Т.П., Энгхофф С., Таунсенд Дж., Куршен Е., Сейновски Т.Дж. (январь 2002 г.). «Динамические мозговые источники зрительных вызванных реакций». Наука . 295 (5555): 690–4. Бибкод : 2002Sci...295..690M . дои : 10.1126/science.1066168 . ПМИД 11809976 . S2CID 15200185 .
- ^ Мякинен В., Тийтинен Х., Мэй П. (февраль 2005 г.). «Слуховые реакции, связанные с событиями, генерируются независимо от текущей активности мозга». НейроИмидж . 24 (4): 961–8. doi : 10.1016/j.neuroimage.2004.10.020 . ПМИД 15670673 . S2CID 16210275 .
- ^ Никулин В.В., Линкенкаер-Хансен К., Нолте Г., Лемм С., Мюллер К.Р. , Ильмониеми Р.Дж., Курио Г. (май 2007 г.). «Новый механизм вызванных реакций в человеческом мозге». Европейский журнал неврологии . 25 (10): 3146–54. дои : 10.1111/j.1460-9568.2007.05553.x . ПМИД 17561828 . S2CID 12113334 .
- ^ Мазахери А., Дженсен О (июль 2008 г.). «Асимметричные амплитудные модуляции колебаний мозга порождают медленные вызванные реакции» . Журнал неврологии . 28 (31): 7781–7. doi : 10.1523/JNEUROSCI.1631-08.2008 . ПМК 6670375 . ПМИД 18667610 .
- ^ Мазахери А., Дженсен О (2008). «Ритмическая пульсация: связь текущей активности мозга с вызванными реакциями» . Границы человеческой неврологии . 4 : 177. дои : 10.3389/fnhum.2010.00177 . ПМЦ 2972683 . ПМИД 21060804 .
- ^ Хамалайнен М., Хари Р., Ильмониеми Р.Ю., Кнуутила Дж., Лоунасмаа О.В. (1993). «Магнитоэнцефалография. Теория, приборы и приложения к неинвазивным исследованиям работающего мозга человека» . Ред. Мод Физ . 65 (2): 413–497. Бибкод : 1993RvMP...65..413H . дои : 10.1103/RevModPhys.65.413 .
- ^ Хиафил, Александр; Жиро, Анн-Лиза; Фонтолан, Лоренцо; Гуткин, Борис (01 ноября 2015 г.). «Нейронная межчастотная связь: соединяющие архитектуры, механизмы и функции» . Тенденции в нейронауках . 38 (11): 725–740. дои : 10.1016/j.tins.2015.09.001 . ISSN 0166-2236 . ПМИД 26549886 . S2CID 3545001 .
- ^ Лисман, Джон Э.; Йенсен, Оле (20 марта 2013 г.). «Нейронный код Тета-гамма» . Нейрон . 77 (6): 1002–1016. дои : 10.1016/j.neuron.2013.03.007 . ISSN 0896-6273 . ПМЦ 3648857 . ПМИД 23522038 .
- ^ Дриё, Селин; Зугаро, Микаэль (2019). «Последовательности гиппокампа во время исследования: механизмы и функции» . Границы клеточной нейронауки . 13 : 232. дои : 10.3389/fncel.2019.00232 . ISSN 1662-5102 . ПМК 6584963 . ПМИД 31263399 .
- ^ Перейти обратно: а б Певица В. (1993). «Синхронизация корковой активности и ее предполагаемая роль в обработке информации и обучении». Ежегодный обзор физиологии . 55 : 349–74. дои : 10.1146/annurev.ph.55.030193.002025 . ПМИД 8466179 .
- ^ Певец В., Грей СМ (1995). «Интеграция визуальных функций и гипотеза временной корреляции». Ежегодный обзор неврологии . 18 : 555–86. CiteSeerX 10.1.1.308.6735 . дои : 10.1146/annurev.ne.18.030195.003011 . ПМИД 7605074 .
- ^ Мардер Э., Бучер Д. (ноябрь 2001 г.). «Центральные генераторы узоров и контроль ритмических движений» . Современная биология . 11 (23): Р986-96. Бибкод : 2001CBio...11.R986M . дои : 10.1016/S0960-9822(01)00581-4 . ПМИД 11728329 . S2CID 1294374 .
- ^ Димитриевич М.Р., Герасименко Ю., Пинтер М.М. (ноябрь 1998 г.). «Доказательства существования генератора центральных паттернов спинного мозга у людей». Анналы Нью-Йоркской академии наук . 860 (1): 360–76. Бибкод : 1998NYASA.860..360D . дои : 10.1111/j.1749-6632.1998.tb09062.x . ПМИД 9928325 . S2CID 102514 .
- ^ Даннер С.М., Хофстоеттер США, Фрейндл Б., Биндер Х., Майр В., Раттай Ф., Минасян К. (март 2015 г.). «Управление спинальной локомоторикой человека основано на гибко организованных импульсных генераторах» . Мозг . 138 (Часть 3): 577–88. дои : 10.1093/brain/awu372 . ПМЦ 4408427 . ПМИД 25582580 .
- ^ Перейти обратно: а б Гупта Н., Сингх С.С., Стопфер М. (декабрь 2016 г.). «Осцилляционные окна интеграции в нейронах» . Природные коммуникации . 7 : 13808. Бибкод : 2016NatCo...713808G . дои : 10.1038/ncomms13808 . ПМК 5171764 . ПМИД 27976720 .
- ^ Ахиссар Э., Нелингер Г., Асса Э., Карп О. и Сараф-Синик И. Таламокортикальные петли как временные демодуляторы чувств. Коммуникационная биология 6, 562 (2023).
- ^ Милнер ПМ (ноябрь 1974 г.). «Модель визуального распознавания форм». Психологический обзор . 81 (6): 521–35. дои : 10.1037/h0037149 . ПМИД 4445414 .
- ^ Перейти обратно: а б Грей СМ, Кениг П., Энгель А.К., Сингер В. (март 1989 г.). «Колебательные реакции зрительной коры головного мозга кошки демонстрируют межколоночную синхронизацию, которая отражает глобальные свойства стимула». Природа . 338 (6213): 334–7. Бибкод : 1989Natur.338..334G . дои : 10.1038/338334a0 . ПМИД 2922061 . S2CID 4281744 .
- ^ Экхорн Р., Бауэр Р., Джордан В., Брош М., Крузе В., Мунк М., Рейтбёк Х.Дж. (1988). «Когерентные колебания: механизм связывания признаков в зрительной коре? Многоэлектродный и корреляционный анализ у кошки». Биологическая кибернетика . 60 (2): 121–30. дои : 10.1007/BF00202899 . ПМИД 3228555 . S2CID 206771651 .
- ^ Вер М., Лоран Г. (ноябрь 1996 г.). «Кодирование запаха временными последовательностями срабатываний в колеблющихся нейронных агрегатах». Природа . 384 (6605): 162–6. Бибкод : 1996Natur.384..162W . дои : 10.1038/384162a0 . ПМИД 8906790 . S2CID 4286308 .
- ^ Маклеод К., Лоран Дж. (ноябрь 1996 г.). «Отличные механизмы синхронизации и временного формирования нейронных агрегатов, кодирующих запах». Наука . 274 (5289): 976–9. Бибкод : 1996Sci...274..976M . дои : 10.1126/science.274.5289.976 . ПМИД 8875938 . S2CID 10744144 .
- ^ Стопфер М., Багаван С., Смит Б.Х., Лоран Дж. (ноябрь 1997 г.). «Нарушение распознавания запахов из-за десинхронизации нейронных агрегатов, кодирующих запах». Природа . 390 (6655): 70–4. Бибкод : 1997Natur.390...70S . дои : 10.1038/36335 . ПМИД 9363891 . S2CID 205024830 .
- ^ Маклауд К., Беккер А., Лоран Г. (октябрь 1998 г.). «Кто считывает временную информацию, содержащуюся в синхронизированных и осциллирующих последовательностях импульсов?». Природа . 395 (6703): 693–8. Бибкод : 1998Natur.395..693M . дои : 10.1038/27201 . ПМИД 9790189 . S2CID 4424801 .
- ^ Бухуси CV, Мек WH (октябрь 2005 г.). «Что нами движет? Функциональные и нейронные механизмы определения интервалов времени». Обзоры природы. Нейронаука . 6 (10): 755–65. дои : 10.1038/nrn1764 . ПМИД 16163383 . S2CID 29616055 .
- ^ Ахиссар Э., Заксенхаус М (2001). «Глава 6 Временное и пространственное кодирование вибриссной системы крысы». Достижения в кодировании нейронной популяции . Прогресс в исследованиях мозга. Том. 130. С. 75–87. дои : 10.1016/S0079-6123(01)30007-9 . ISBN 9780444501103 . ПМИД 11480290 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Бернс С.П., Син Д., Шепли Р.М. (июнь 2011 г.). «Является ли активность гамма-диапазона в потенциале локального поля коры V1 «часами» или отфильтрованным шумом?» . Журнал неврологии . 31 (26): 9658–64. doi : 10.1523/jneurosci.0660-11.2011 . ПМЦ 3518456 . ПМИД 21715631 .
- ^ Пфурчеллер Г., Аранибар А. (июнь 1977 г.). «Связанная с событием корковая десинхронизация, обнаруженная с помощью измерения мощности ЭЭГ кожи головы». Электроэнцефалография и клиническая нейрофизиология . 42 (6): 817–26. дои : 10.1016/0013-4694(77)90235-8 . ПМИД 67933 .
- ^ Мурти В.Н., Фетц Э.Э. (декабрь 1996 г.). «Колебательная активность сенсомоторной коры бодрствующих обезьян: синхронизация потенциалов локальных полей и связь с поведением». Журнал нейрофизиологии . 76 (6): 3949–67. дои : 10.1152/jn.1996.76.6.3949 . ПМИД 8985892 .
- ^ Санес Дж. Н., Донохью Дж. П. (май 1993 г.). «Колебания локальных потенциалов поля моторной коры приматов при произвольных движениях» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (10): 4470–4. Бибкод : 1993PNAS...90.4470S . дои : 10.1073/pnas.90.10.4470 . ПМЦ 46533 . ПМИД 8506287 .
- ^ Конвей Б.А., Холлидей Д.М., Фармер С.Ф., Шахани У., Маас П., Вейр А.И., Розенберг-младший (декабрь 1995 г.). «Синхронизация между моторной корой и мотонейронным пулом спинного мозга при выполнении постоянной двигательной задачи у человека» . Журнал физиологии . 489 (Часть 3) (3): 917–24. doi : 10.1113/jphysicalol.1995.sp021104 . ПМЦ 1156860 . ПМИД 8788955 .
- ^ Салениус С., Портин К., Кайола М., Салмелин Р., Хари Р. (июнь 1997 г.). «Корковый контроль активности мотонейронов человека во время изометрического сокращения». Журнал нейрофизиологии . 77 (6): 3401–5. дои : 10.1152/jn.1997.77.6.3401 . ПМИД 9212286 . S2CID 2178927 .
- ^ Бейкер С.Н., Оливье Э., Лемон Р.Н. (май 1997 г.). «Когерентные колебания в моторной коре обезьян и ЭМГ мышц рук демонстрируют модуляцию, зависящую от задачи» . Журнал физиологии . 501 (Часть 1) (1): 225–41. дои : 10.1111/j.1469-7793.1997.225bo.x . ПМЦ 1159515 . ПМИД 9175005 .
- ^ Бунстра Т.В., Данна-Дос-Сантос А., Се Х.Б., Рёрдинк М., Стинс Дж.Ф., Брейкспир М. (декабрь 2015 г.). «Мышечные сети: анализ связности активности ЭМГ во время постурального контроля» . Научные отчеты . 5 : 17830. Бибкод : 2015NatSR...517830B . дои : 10.1038/srep17830 . ПМЦ 4669476 . ПМИД 26634293 .
- ^ Керкман Дж.Н., Даффертсхофер А., Голло Л.Л., Брейкспир М., Бунстра Т.В. (июнь 2018 г.). «Сетевая структура опорно-двигательного аппарата человека формирует нейронные взаимодействия в нескольких временных масштабах» . Достижения науки . 4 (6): eaat0497. Бибкод : 2018SciA....4..497K . дои : 10.1126/sciadv.aat0497 . ПМК 6021138 . ПМИД 29963631 .
- ^ Рубино Д., Роббинс К.А., Хацопулос Н.Г. (декабрь 2006 г.). «Распространяющиеся волны опосредуют передачу информации в моторной коре». Природная неврология . 9 (12): 1549–57. дои : 10.1038/nn1802 . ПМИД 17115042 . S2CID 16430438 .
- ^ Хейтманн С., Бунстра Т., Гонг П., Брейкспир М., Эрментраут Б. (2015). «Ритмы устойчивой позы: двигательные команды как пространственно организованные модели колебаний». Нейрокомпьютинг . 170 : 3–14. дои : 10.1016/j.neucom.2015.01.088 .
- ^ Хейтманн С., Бунстра Т., Брейкспир М. (октябрь 2013 г.). «Дендритный механизм декодирования бегущих волн: принципы и применение в моторной коре» . PLOS Вычислительная биология . 9 (10): е1003260. Бибкод : 2013PLSCB...9E3260H . дои : 10.1371/journal.pcbi.1003260 . ПМЦ 3814333 . ПМИД 24204220 .
- ^ Аллум Дж. Х., Дитц В., Фройнд Х. Дж. (май 1978 г.). «Нейрональные механизмы, лежащие в основе физиологического тремора». Журнал нейрофизиологии . 41 (3): 557–71. дои : 10.1152/jn.1978.41.3.557 . ПМИД 660226 .
- ^ Вальбо А.Б., Вессберг Дж. (сентябрь 1993 г.). «Организация двигательной деятельности при медленных движениях пальцев у человека» . Журнал физиологии . 469 : 673–91. doi : 10.1113/jphysicalol.1993.sp019837 . ПМЦ 1143894 . ПМИД 8271223 .
- ^ Гросс Дж., Тиммерманн Л., Куджала Дж., Диркс М., Шмитц Ф., Салмелин Р., Шницлер А. (февраль 2002 г.). «Нейронная основа прерывистого двигательного контроля у человека» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (4): 2299–302. Бибкод : 2002PNAS...99.2299G . дои : 10.1073/pnas.032682099 . ПМЦ 122359 . ПМИД 11854526 .
- ^ Бузаки Г (2006). Ритмы мозга . Издательство Оксфордского университета.
- ^ Найхус Э., Карран Т. (июнь 2010 г.). «Функциональная роль гамма- и тета-колебаний в эпизодической памяти» . Неврологические и биоповеденческие обзоры . 34 (7): 1023–35. doi : 10.1016/j.neubiorev.2009.12.014 . ПМЦ 2856712 . ПМИД 20060015 .
- ^ Рутисхаузер Ю, Росс И.Б., Мамелак А.Н., Шуман Э.М. (апрель 2010 г.). «Сила человеческой памяти прогнозируется с помощью фазовой блокировки тета-частоты отдельных нейронов» (PDF) . Природа . 464 (7290): 903–7. Бибкод : 2010Natur.464..903R . дои : 10.1038/nature08860 . ПМИД 20336071 . S2CID 4417989 .
- ^ Бранкаччо А., Табарелли Д., Бигика М., Балдауф Д. (апрель 2020 г.). «Локализация коркового источника специфической колебательной активности стадии сна» . Научные отчеты . 10 (1): 6976. Бибкод : 2020NatSR..10.6976B . дои : 10.1038/s41598-020-63933-5 . ПМЦ 7181624 . ПМИД 32332806 .
- ^ Феллер М.Б. (июль 2009 г.). «Волны на сетчатке, вероятно, управляют формированием специфичных для глаз проекций ретиногенекулята» . Нейронное развитие . 4:24 . дои : 10.1186/1749-8104-4-24 . ПМК 2706239 . ПМИД 19580682 .
- ^ Маколи Дж. Х., компакт-диск Марсдена (август 2000 г.). «Физиологический и патологический тремор и ритмическая центральная двигательная регуляция» . Мозг . 123 (Часть 8) (8): 1545–67. дои : 10.1093/мозг/123.8.1545 . ПМИД 10908186 .
- ^ Шустерман В., Трой В.К. (июнь 2008 г.). «От исходного уровня к эпилептиформной активности: путь к синхронизированной ритмичности в крупномасштабных нейронных сетях». Физический обзор E . 77 (6 Pt 1): 061911. Бибкод : 2008PhRvE..77f1911S . дои : 10.1103/PhysRevE.77.061911 . ПМИД 18643304 . S2CID 13928602 .
- ^ Ваннесте С., Сонг Джей Джей, Де Риддер Д. (март 2018 г.). «Таламокортикальная аритмия, обнаруженная с помощью машинного обучения» . Природные коммуникации . 9 (1): 1103. Бибкод : 2018NatCo...9.1103V . дои : 10.1038/s41467-018-02820-0 . ПМЦ 5856824 . ПМИД 29549239 .
- ^ Бирбаумер Н. (ноябрь 2006 г.). «Нарушая тишину: интерфейсы мозг-компьютер (BCI) для связи и управления движением» . Психофизиология . 43 (6): 517–32. дои : 10.1111/j.1469-8986.2006.00456.x . ПМИД 17076808 .
- ^ Видал Джей-Джей (1973). «На пути к прямой связи мозг-компьютер» . Ежегодный обзор биофизики и биоинженерии . 2 : 157–80. дои : 10.1146/annurev.bb.02.060173.001105 . ПМИД 4583653 .
- ^ Божиновский С., Сестаков М., Божиновская Л. (ноябрь 1988 г.). «Использование альфа-ритма ЭЭГ для управления мобильным роботом». Материалы ежегодной международной конференции Общества инженеров IEEE в медицине и биологии . Новый Орлеан: IEEE. стр. 1515–1516. дои : 10.1109/IEMBS.1988.95357 . ISBN 0-7803-0785-2 . S2CID 62179588 .
- ^ Божиновский С (август 1990 г.). «Управление траекторией мобильного робота: от фиксированных рельсов к прямому биоэлектрическому управлению». В Кайнаке О (ред.). Материалы международного семинара IEEE по интеллектуальному управлению движением . Том. 2. Стамбул: IEEE. стр. 463–467. дои : 10.1109/IMC.1990.687362 . S2CID 60642344 .
- ^ Лебедев М (2016). «Усиление сенсомоторных функций с помощью нейронных протезов». Опера медика и физиология . 2 (3–4): 211–227. doi : 10.20388/OMP.003.0035 (неактивен 31 января 2024 г.).
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Лебедев М.А., Николелис М.А. (апрель 2017 г.). «Интерфейсы мозг-машина: от фундаментальной науки к нейропротезам и нейрореабилитации». Физиологические обзоры . 97 (2): 767–837. doi : 10.1152/physrev.00027.2016 . ПМИД 28275048 .
Дальнейшее чтение
[ редактировать ]- Бужаки Г (2006). Ритмы мозга . Издательство Оксфордского университета. ISBN 978-0-19-530106-9 .
- Фриман В. (1975). Массовое действие в нервной системе . Академическая пресса. ISBN 978-0124120471 . Архивировано из оригинала 5 июля 2015 г.