Нейронное колебание


Нейронные колебания , или мозговые волны , являются ритмичными или повторяющимися паттернами нервной активности в центральной нервной системе . Нейронная ткань может вызвать колебательную активность во многих отношениях, управляемой либо механизмами в отдельных нейронах , либо путем взаимодействия между нейронами. В отдельных нейронах колебания могут появляться либо в виде колебаний в мембранном потенциале , либо в качестве ритмических паттернов потенциалов действия , которые затем вызывают колебательную активацию постсинаптических нейронов. На уровне нейронных ансамблей синхронизированная активность большого количества нейронов может привести к макроскопическим колебаниям, которые можно наблюдать на электроэнцефалограмме . Колебательная активность в группах нейронов обычно возникает из -за связей обратной связи между нейронами, которые приводят к синхронизации их схемы стрельбы. Взаимодействие между нейронами может привести к колебаниям на различной частоте, чем частота стрельбы отдельных нейронов. Хорошо известным примером макроскопических нейронных колебаний является альфа -активность .
Нейронные колебания у людей наблюдали исследователи еще в 1924 году ( Ханс Бергер ). Более 50 лет спустя в нейронах позвоночных встречались внутреннее колебательное поведение, но его функциональная роль до сих пор не до конца понятна. [ 3 ] Возможные роли нервных колебаний включают привязку к признакам , механизмы передачи информации и генерацию ритмического моторного выхода . За последние десятилетия было получено более понимания, особенно с достижениями в области визуализации мозга . Основная область исследований в области нейробиологии включает в себя определение того, как генерируются колебания и каковы их роли. Колебательная активность в мозге широко наблюдается на разных уровнях организации и, как полагают, играет ключевую роль в обработке нейронной информации. Многочисленные экспериментальные исследования подтверждают функциональную роль нервных колебаний; Однако унифицированное толкование все еще не хватает.
История
[ редактировать ]Ричард Катон обнаружил электрическую активность в полушариях головного мозга кроликов и обезьян и представил свои результаты в 1875 году. [ 4 ] Адольф Бек опубликовал в 1890 году его наблюдения спонтанной электрической активности мозга кроликов и собак, которые включали ритмические колебания, измененные светом, обнаруженные с помощью электродов, расположенных непосредственно на поверхности мозга. [ 5 ] Перед Гансом Бергером Владимир Владимирович Правдич-Неминский опубликовал первое ЭЭГ животного и вызванный потенциал собаки. [ 6 ]
Обзор
[ редактировать ]Нейронные колебания наблюдаются по всей центральной нервной системе на всех уровнях и включают в себя шипы , локальные полевые потенциалы и крупномасштабные колебания , которые могут быть измерены с помощью электроэнцефалографии (ЭЭГ). В целом, колебания могут характеризоваться их частотой , амплитудой и фазой . Эти свойства сигнала могут быть извлечены из нейронных записей, используя частотный анализ . Считается, что при крупномасштабных колебаниях изменения амплитуды являются результатом изменений синхронизации в нейронном ансамбле , также называемом локальной синхронизацией. В дополнение к локальной синхронизации, колебательная активность отдаленных нервных структур (отдельные нейроны или нейронные ансамбли) может синхронизировать. Нейронные колебания и синхронизация были связаны со многими когнитивными функциями, такими как передача информации, восприятие, управление двигателем и память. [ 7 ] [ 8 ] [ 9 ] [ 10 ]
Противоположностью синхронизации нейронов является нейронная изоляция, а это когда электрическая активность нейронов не синхронизируется во времени. Это когда вероятность того, что нейрона достигнет своего порогового потенциала для распространения сигнала в следующем нейроне уменьшается. Это явление обычно наблюдается, когда спектральная интенсивность уменьшается от суммирования этих нейронов, которые можно использовать для дифференциации когнитивной функции или нервной изоляции. Тем не менее, были использованы новые нелинейные методы, что пара временных и спектральных энтропийных отношений одновременно, чтобы охарактеризовать, как выделяются нейроны (неспособность сигнала распространять на соседние нейроны), индикатор нарушений (например, гипоксия). [ 1 ]
Нейронные колебания наиболее широко изучались в нейронной активности, генерируемой большими группами нейронов. Крупномасштабная активность может быть измерена с помощью таких методов, как ЭЭГ. В целом, сигналы ЭЭГ имеют широкое спектральное содержание, аналогичное розовому шуму , но также выявляют колебательную активность в определенных полосах частот. Первая обнаруженная и наиболее известная частотная полоса- альфа-активность (8–12 Гц ) [ 11 ] [ 12 ] [ 13 ] Это может быть обнаружено из затылочной доли во время расслабленного бодрствования и увеличивается, когда глаза закрыты. [ 14 ] Другие полосы частот: Delta (1–4 Гц), тета (4–8 Гц), бета (13–30 Гц), низкая гамма (30–70 Гц), [ 15 ] и высокая частота гамма (70–150 Гц). Более быстрые ритмы, такие как гамма -активность, были связаны с когнитивной обработкой. Действительно, сигналы ЭЭГ резко меняются во время сна. На самом деле, разные стадии сна обычно характеризуются их спектральным содержанием. [ 16 ] Следовательно, нейронные колебания были связаны с когнитивными состояниями, такими как осознание и сознание . [ 17 ] [ 18 ] [ 15 ] [ 13 ]
Хотя нервные колебания в активности мозга человека в основном исследуются с использованием записей ЭЭГ, они также наблюдаются с использованием более инвазивных методов записи, таких как одноразовые записи . Нейроны могут генерировать ритмические паттерны потенциалов действия или шипов. Некоторые типы нейронов имеют тенденцию стрелять на определенных частотах, либо в качестве резонаторов [ 19 ] или как внутренние генераторы . [ 2 ] Разрыв - еще одна форма ритмического всплеска. Паттерны пика считаются фундаментальными для кодирования информации в мозге. Колебательная активность также может наблюдаться в форме колебаний подпороговой мембраны (т.е. в отсутствие потенциалов действия). [ 20 ] Если многочисленные нейроны в синхронности вспыхивают , они могут привести к колебаниям в местных полевых потенциалах . Количественные модели могут оценить силу нейронных колебаний в зарегистрированных данных. [ 21 ]
Нейронные колебания обычно изучаются в математической рамках и принадлежат к области «невродинамики», области исследований в когнитивных науках , которая уделяет сильное внимание динамическому характеру нейронной активности при описании функции мозга . [ 22 ] Он считает мозг динамической системой и использует дифференциальные уравнения, чтобы описать, как развивается нейронная активность с течением времени. В частности, он стремится связывать динамические паттерны мозговой активности с когнитивными функциями, такими как восприятие и память. В очень абстрактной форме можно анализировать нейронные колебания . [ 23 ] [ 24 ] При изучении в более физиологически реалистичных условиях колебательная активность обычно изучается с использованием компьютерного моделирования вычислительной модели .
Функции нервных колебаний широко распространены и различаются для различных типов колебательной активности. Примерами являются генерация ритмической активности, такой как сердцебиение и нервное связывание сенсорных особенностей в восприятии, таких как форма и цвет объекта. Нейронные колебания также играют важную роль во многих неврологических расстройствах , таких как чрезмерная синхронизация во время судорожной активности при эпилепсии или тремор у пациентов с болезнью Паркинсона . Колебательная активность также может использоваться для контроля внешних устройств, таких как интерфейс мозга и компута . [ 25 ]
Физиология
[ редактировать ]Колебательная активность наблюдается по всей центральной нервной системе на всех уровнях организации. Три разных уровня были широко признаны: микромасштабная (активность одного нейрона), мезомасштаб (активность локальной группы нейронов) и макромасштаб (активность различных областей мозга). [ 26 ]
Микроскопический
[ редактировать ]Нейроны генерируют потенциалы действия , возникающие в результате изменений в потенциале электрической мембраны. Нейроны могут генерировать множественные потенциалы действия в последовательности, образующих так называемые шиповые поезда. Эти шипы являются основой для нейронного кодирования и переноса информации в мозге. Поезда Spike могут образовывать все виды узоров, такие как ритмичный всплеск и разрыв , и часто демонстрируют колебательную активность. [ 27 ] Колебательная активность в отдельных нейронах также может наблюдаться в субпороговых колебаниях в мембранном потенциале. Эти ритмические изменения в мембранном потенциале не достигают критического порога и, следовательно, не приводят к потенциалу действия. Они могут быть результатом постсинаптических потенциалов от синхронных входов или из внутренних свойств нейронов.
Нейрональный всплеск может быть классифицирован по схеме активности. Возбудимость нейронов может быть подразделена в классе I и II. Нейроны класса I могут генерировать потенциалы действия с произвольно низкой частотой в зависимости от прочности входа, тогда как нейроны класса II генерируют потенциалы действия в определенной полосе частот, что относительно нечувствительно к изменениям в прочности входа. [ 19 ] Нейроны класса II также более склонны к демонстрации субпороговых колебаний в мембранном потенциале.
Мезоскопический
[ редактировать ]Группа нейронов также может вызывать колебательную активность. Благодаря синаптическим взаимодействиям модели стрельбы различных нейронов могут стать синхронизированными, и ритмические изменения в электрическом потенциале, вызванные их потенциалом действия, могут накапливаться ( конструктивные помехи ). То есть, синхронизированные схемы стрельбы приводят к синхронизированному входу в другие области кортикальной коры, что приводит к возникновению колебаний большой амплитуды локального потенциала поля . Эти крупномасштабные колебания также могут быть измерены за пределами кожи головы с использованием электроэнцефалографии (ЭЭГ) и магнитоэнцефалографии (МЭГ). Электрические потенциалы, генерируемые отдельными нейронами, слишком малы, чтобы их можно было поднять вне кожи головы, а активность ЭЭГ или МЭГ всегда отражает суммирование синхронной активности тысяч или миллионов нейронов, которые имеют сходную пространственную ориентацию. [ 28 ]
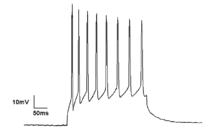
Нейроны в нейронном ансамбле редко бывают в одном и том же моменте, т.е. полностью синхронизировано. Вместо этого вероятность стрельбы ритмично модулируется таким образом, что нейроны с большей вероятностью будут стрелять одновременно, что порождает колебания в их средней активности. (См. Рисунок в верхней части страницы.) Таким образом, частота крупномасштабных колебаний не должна соответствовать схеме стрельбы отдельных нейронов. Изолированные корковые нейроны регулярно стреляют в определенных условиях, но в интактном мозге корковые клетки бомбардируют очень колеблющимися синаптическими входами и обычно стреляют в случайном. Однако, если вероятность большой группы запуска нейронов ритмично модулирована на общей частоте, они будут генерировать колебания в среднем поле. (См. Также рисунок в верхней части страницы.) [ 27 ]
Нейронные ансамбли могут генерировать колебательную активность эндогенно посредством локальных взаимодействий между возбуждающими и ингибирующими нейронами. В частности, ингибирующие интернейроны играют важную роль в производстве синхронности нейронных ансамблей путем генерации узкого окна для эффективного возбуждения и ритмично модуляции скорости стрельбы возбуждающих нейронов. [ 29 ]
Макроскопический
[ редактировать ]Нейронные колебания также могут возникнуть в результате взаимодействия между различными областями мозга, связанными с структурным разъемом . Задержки времени играют важную роль здесь. Поскольку все области мозга связаны двунаправленными, эти связи между областями мозга образуют петли обратной связи . Положительные петли обратной связи имеют тенденцию вызывать колебательную активность, когда частота обратно связана со временем задержки. Примером такой петли обратной связи является связь между таламусом и корой - таламокортикальным излучением . Эта таламокортикальная сеть способна генерировать колебательную активность, известную как рецидивирующий таламо-кортикальный резонанс . [ 30 ] Таламокортикальная сеть играет важную роль в генерации альфа -активности . [ 31 ] [ 32 ] В сетевой модели всего мозга с реалистичной анатомической связностью и задержками распространения между областями мозга колебания в бета-частотном диапазоне вытекают из частичной синхронизации подмножествам мозга, колеблющихся в гамма-диапазоне (генерируется на мезоскопическом уровне). [ 33 ]
Механизмы
[ редактировать ]Нейрональные свойства
[ редактировать ]Ученые идентифицировали некоторые внутренние нейрональные свойства , которые играют важную роль в создании колебаний мембранных потенциалов. В частности, ионные каналы, управляемые напряжением, имеют решающее значение для генерации потенциалов действия. Динамика этих ионных каналов была захвачена в устоявшейся модели Ходжкина-Хексли, которая описывает, как потенциалы действий инициируются и распространяются с помощью набора дифференциальных уравнений. Используя анализ бифуркации , могут быть определены различные колебательные сорта этих нейрональных моделей, что позволяет классифицировать типы нейрональных реакций. Колебательная динамика всплеска нейронов, как идентифицировано в модели Ходжкина - Хексли, тесно согласна с эмпирическими результатами.
В дополнение к периодическому всплеску, потенциальные колебания подпороговой мембраны , то есть резонансное поведение, которое не приводит к потенциалам действия, также может способствовать колебательной активности, способствуя синхронной активности соседних нейронов. [ 34 ] [ 35 ]
Подобно нейронам кардиостимулятора в центральных генераторах рисунков , подтипы корковых клеток огры стрелков из шипов (короткие кластеры шипов) ритмично на предпочтительных частотах. [ 2 ] Разрывающие нейроны могут служить кардиостимуляторами для синхронных колебаний сети, а всплески шипов могут лежать в основе или усилить нейрональный резонанс. [ 27 ] Многие из этих нейронов можно считать внутренними осцилляторами, а именно, нейроны, которые генерируют свои колебания по сути, поскольку их частоты колебаний могут быть изменены местными применениями глутамата in vivo. [ 36 ]
Свойства сети
[ редактировать ]Помимо внутренних свойств нейронов, свойства биологической нейронной сети также являются важным источником колебательной активности. Нейроны общаются друг с другом через синапсы и влияют на время шиповских поездов в постсинаптических нейронах. В зависимости от свойств соединения, таких как прочность связи, задержка времени и то, является ли связь возбуждающей или ингибирующей , всплывающие поезда взаимодействующих нейронов могут стать синхронизированными . [ 37 ] Нейроны локально связаны, образуя небольшие кластеры, которые называются нейронными ансамблями . Некоторые сетевые структуры способствуют колебательной активности на определенных частотах. Например, нейрональная активность, генерируемая двумя популяциями взаимосвязанных ингибирующих и возбуждающих клеток, может показать спонтанные колебания, которые описаны моделью Уилсон-Коуэна .
Если группа нейронов участвует в синхронизированной колебательной активности, нейронный ансамбль может быть математически представлен как один генератор. [ 26 ] Различные нейронные ансамбли связаны с помощью дальних соединений и образуют сеть слабо связанных генераторов в следующем пространственном масштабе. Слабо связанные генераторы могут генерировать ряд динамики, включая колебательную активность. [ 38 ] Большой диапазон между различными структурами мозга, такими как таламус и кора (см. Таламокортикальные колебания ), включают в себя сроки по времени из-за конечной скорости проводимости аксонов. Поскольку большинство соединений являются взаимными, они формируют петли питательной спины , которые поддерживают колебательную активность. Колебания, зарегистрированные из множества областей коры, могут стать синхронизированными с образованием крупномасштабных сетей мозга , чья динамика и функциональная связь могут быть изучены с помощью спектрального анализа и причинности Грейнджера . измерений [ 39 ] Когерентная активность крупномасштабной активности мозга может образовывать динамические связи между областями мозга, необходимыми для интеграции распределенной информации. [ 18 ]
Было показано, что микроглия - основные иммунные клетки мозга - играют важную роль в формировании сетевой связности и, таким образом, влияя на колебания нейрональной сети как ex vivo , так и in vivo . [ 40 ]
Нейромодуляция
[ редактировать ]В дополнение к быстрому прямым синаптическим взаимодействиям между нейронами, образующими сеть, колебательная активность регулируется нейромодуляторами в гораздо более медленной масштабе времени. То есть, как известно, уровни концентрации определенных нейротрансмиттеров регулируют количество колебательной активности. Например, ГАМК положительно коррелирует с частотой колебаний в индуцированных стимулах. было показано, что концентрация [ 41 ] Ряд ядер в стволе мозга имеет диффузные проекции по всему мозгу, влияющие на уровни концентрации нейротрансмиттеров, таких как норадреналин , ацетилхолин и серотонин . Эти нейротрансмиттерные системы влияют на физиологическое состояние, например, бодрствование или возбуждение и оказывают выраженное влияние на амплитуду различных мозговых волн, таких как альфа -активность. [ 42 ]
Математическое описание
[ редактировать ]Колебания часто можно описать и анализировать с использованием математики. Математики определили несколько динамических механизмов, которые генерируют ритмичность. Среди наиболее важных- гармонические (линейные) осцилляторы, осцилляторы ограниченного цикла и генераторы с отсроченной обратной связью . [ 43 ] Гармонические колебания появляются очень часто в природе - эксплуатации - это звуковые волны, движение маятника и вибрации любого рода. Как правило, они возникают, когда физическая система возмущается небольшой степенью из состояния с минимальной энергией и хорошо изучена математически.
Хармонические генераторы, управляемые шумом, реалистично имитируют альфа-ритм в бодрствующем ЭЭГ, а также медленные волны и шпинции в ЭЭГ сна. Успешные алгоритмы анализа ЭЭГ были основаны на таких моделях. Несколько других компонентов ЭЭГ лучше описаны с помощью предельного цикла или колебаний с отсроченным обратном обратной связью.
Долаживания предельного цикла возникают из-за физических систем, которые показывают большие отклонения от равновесия , тогда как колебания с задержкой обратной связи возникают, когда компоненты системы влияют друг на друга после значительных временных задержек. Долаживание предельного цикла может быть сложными, но для их анализа существуют мощные математические инструменты; Математика колебаний с отсроченной обратной связью примитивна по сравнению. Линейные осцилляторы и осцилляторы ограниченного цикла качественно различаются с точки зрения того, как они реагируют на колебания при вводе. В линейном генераторе частота более или менее постоянна, но амплитуда может сильно различаться. В генераторе ограниченного цикла амплитуда имеет тенденцию быть более или менее постоянной, но частота может сильно различаться. Сердцебиение . является примером колебаний с ограниченным циклом в том смысле, что частота ритмов варьируется широко, в то время как каждый отдельный бит продолжает качать примерно одинаковое количество крови
Вычислительные модели принимают различные абстракции, чтобы описать сложную колебательную динамику, наблюдаемую в мозговой активности. Многие модели используются в этой области, каждая из которых определяется на разных уровнях абстракции и пытается смоделировать различные аспекты нейронных систем. Они варьируются от моделей краткосрочного поведения отдельных нейронов за счет моделей того, как динамика нейронной схемы возникает от взаимодействия между отдельными нейронами, до моделей того, как поведение может возникнуть из абстрактных нейронных модулей, которые представляют полные подсистемы.
Одиночный нейрон модель
[ редактировать ]
Модель биологического нейрона представляет собой математическое описание свойств нервных клеток или нейронов, которое предназначено для точного описания и прогнозирования его биологических процессов. Одной из наиболее успешных моделей нейронов является модель Ходжкина - Хексли, для которой Ходжкин и Хаксли получили Нобелевскую премию 1963 года в области физиологии или медицины. Модель основана на данных из гигантского аксона кальмара и состоит из нелинейных дифференциальных уравнений, которые приближают электрические характеристики нейрона, включая генерацию и распространение потенциалов действия . Модель настолько успешна в описании этих характеристик, что вариации ее «основанной на проводимости» по-прежнему используются в моделях нейронов более полувека спустя. [ 44 ]
Модель Hodgkin-Huxley слишком сложна, чтобы понять, используя классические математические методы, поэтому исследователи часто обращаются к упрощениям, таким как модель Fitzhugh-Nagumo и модель Hindmarsh-Rose , или модели с высококвалифицированными нейронами, такими как протекающий интегрированный нейроны первоначально разработан Лапиком в 1907 году. [ 45 ] [ 46 ] Такие модели отражают только динамику мембраны, такую как всплеск или разрыв за счет биофизических деталей, но более эффективно вычислительно, что обеспечивает моделирование более крупных биологических нейронных сетей .
Модель всплеска
[ редактировать ]Модель нейронной сети описывает популяцию физически взаимосвязанных нейронов или группу разнородных нейронов, входные данные или сигнальные цели, определяют узнаваемую схему. Эти модели направлены на то, чтобы описать, как динамика нейронной схемы возникает в результате взаимодействия между отдельными нейронами. Локальные взаимодействия между нейронами могут привести к синхронизации активности всплеска и формировать основу колебательной активности. В частности, было показано, что модели взаимодействующих пирамидальных клеток и ингибирующих интернейронов генерируют ритмы мозга, такие как гамма -активность . [ 47 ] Точно так же было показано, что моделирование нейронных сетей с феноменологической моделью для сбоев нейронального ответа может предсказать спонтанные широкополосные нейронные колебания. [ 48 ]
Нейронная модель массы
[ редактировать ]
Модели нейронных поля являются еще одним важным инструментом в изучении нейронных колебаний и представляют собой математическую структуру, описывающую эволюцию переменных, таких как средняя скорость стрельбы в пространстве и времени. При моделировании активности большого количества нейронов центральная идея состоит в том, чтобы перенести плотность нейронов до предела континуума , что приводит к пространственно непрерывным нейронным сетям . Вместо моделирования отдельных нейронов этот подход приближается к группе нейронов по его средним свойствам и взаимодействиям. Он основан на подходе среднего полевого поля , области статистической физики , которая занимается крупномасштабными системами. Модели, основанные на этих принципах, использовались для предоставления математических описаний нервных колебаний и ритмов ЭЭГ. Например, они использовались для исследования визуальных галлюцинаций. [ 50 ]
Модель Курамото
[ редактировать ]Модель курамото связанных фазовых осцилляторов [ 51 ] является одной из самых абстрактных и фундаментальных моделей, используемых для изучения нейронных колебаний и синхронизации. Он отражает активность локальной системы (например, единого нейрона или нейронного ансамбля) только своей круглой фазой и, следовательно, игнорирует амплитуду колебаний (амплитуда постоянна). [ 52 ] Взаимодействия между этими генераторами вводятся простой алгебраической формой (такой как функция синации ) и коллективно генерируют динамический шаблон в глобальном масштабе.
Модель Курамото широко используется для изучения колебательной активности мозга, и было предложено несколько расширений, которые увеличивают ее нейробиологическую достоверность, например, путем включения топологических свойств локальной связности коры. [ 53 ] В частности, в нем описывается, как активность группы взаимодействующих нейронов может стать синхронизированной и генерировать крупномасштабные колебания.
Моделирование с использованием модели Курамото с реалистичной дальнейшей связностью кортикальной связи и задержкой времени, показывают появление медленных колебаний с рисунком, которые воспроизводят функциональные карты, которые воспроизводят жирные карты , которые можно измерить с использованием МРТ . [ 54 ]
Модели деятельности
[ редактировать ]Как отдельные нейроны, так и группы нейронов могут создавать колебательную активность спонтанно. Кроме того, они могут показать колебательные ответы на воспринимаемый вход или моторный выход. Некоторые типы нейронов будут ритмично стрелять в отсутствие какого -либо синаптического ввода. Аналогичным образом, активность всего мозга выявляет колебательную активность, в то время как субъекты не участвуют в какой-либо активности, так называемой активности отдыха . Эти текущие ритмы могут меняться по -разному в ответ на вход восприятия или моторный выход. Колебательная активность может реагировать на увеличение или уменьшение частоты и амплитуды или показывать временное прерывание, которое называется сбросом фазы. Кроме того, внешняя активность может вообще не взаимодействовать с постоянной деятельностью, что приводит к аддитивному ответу.
-
 Частота продолжающейся колебательной активности увеличивается между T1 и T2.
Частота продолжающейся колебательной активности увеличивается между T1 и T2. -
 Амплитуда . непрерывной колебательной активности увеличивается между T1 и T2
Амплитуда . непрерывной колебательной активности увеличивается между T1 и T2 -
 Фаза . постоянной колебательной активности сбросится при T1
Фаза . постоянной колебательной активности сбросится при T1 -
 Активность линейно добавляется к продолжающейся колебательной активности между T1 и T2.
Активность линейно добавляется к продолжающейся колебательной активности между T1 и T2.




Постоянная деятельность
[ редактировать ]Спонтанная активность-это мозговая активность в отсутствие явной задачи, такой как сенсорный вход или моторный выход, а также, следовательно, также называется активностью отдыха. Это противоречит индуцированной активности, т.е. активности мозга, которая вызвана сенсорными стимулами или моторными реакциями.
Термин непрерывная активность мозга используется при электроэнцефалографии и магнитоэнцефалографии для тех сигнальных компонентов, которые не связаны с обработкой стимула или возникновением конкретных других событий, таких как перемещение части тела, т.е., которые не образуют вызванных потенциалов / вызывает поля или индуцированная активность.
Спонтанная активность обычно считается шумом , если кто -то заинтересован в обработке стимулов; Однако, как считается, спонтанная активность играет решающую роль во время развития мозга, например, в формировании сети и синаптогенезе. Спонтанная деятельность может быть информативной в отношении нынешнего психического состояния человека (например, бодрствования, бдительности) и часто используется в исследованиях сна. Некоторые типы колебательной активности, такие как альфа -волны , являются частью спонтанной активности. Статистический анализ флуктуаций мощности альфа-активности показывает бимодальное распределение, то есть режим высокой и низкой амплитуды, и, следовательно, показывает, что активность в состоянии покоя не только отражает процесс шума . [ 55 ]
В случае MRI спонтанные колебания в зависимом (жирном)) сигнале уровня крови выявляют паттерны корреляции, которые связаны с сетями состояния покоя, такими как сеть по умолчанию . [ 56 ] Временная эволюция сетей покоя в состоянии покоя коррелирует с колебаниями колебательной активности ЭЭГ в различных полосах частот. [ 57 ]
Непрерывная активность мозга также может играть важную роль в восприятии, поскольку она может взаимодействовать с активностью, связанной с входящими стимулами. Действительно, исследования ЭЭГ предполагают, что визуальное восприятие зависит как от фазы, так и от амплитуды коры колебаний. Например, амплитуда и фаза альфа -активности в момент визуальной стимуляции предсказывают, будет ли субъект слабый стимул. [ 58 ] [ 59 ] [ 60 ]
Частотная характеристика
[ редактировать ]В ответ на входные данные, нейронный или нейрональный ансамбль может изменить частоту, с которой он колеблется, тем самым изменяя скорость , с которой он пирует. Часто скорость стрельбы нейрона зависит от суммированной деятельности, которую он получает. Изменения частоты также обычно наблюдаются в генераторах центральных рисунков и непосредственно связаны со скоростью моторной активности , такой как шаг частота ходьбы. Тем не менее, изменения в частоте относительных колебаний между различными областями мозга не столь распространены, поскольку частота колебательной активности часто связана с задержками времени между областями мозга.
Амплитудный ответ
[ редактировать ]Помимо вызванной активности, нейронная активность, связанная с обработкой стимула, может привести к индуцированной активности. Индуцированная активность относится к модуляции в продолжающейся активности мозга, вызванной обработкой стимулов или подготовки движения. Следовательно, они отражают косвенный ответ в отличие от вызванных ответов. Хорошо изученным типом индуцированной активности является изменение амплитуды в колебательной активности. Например, гамма -активность часто увеличивается во время повышенной умственной активности, например, во время представления объекта. [ 61 ] Поскольку индуцированные ответы могут иметь разные фазы по измерениям и, следовательно, отменяются во время усреднения, их можно получить только с использованием частотного анализа . Индуцированная активность обычно отражает активность многочисленных нейронов: изменения амплитуды в колебательной активности, как полагают, возникают в результате синхронизации нейронной активности, например, путем синхронизации времени всплеска или колебаний мембранного потенциала отдельных нейронов. Поэтому увеличение колебательной активности часто называют синхронизацией, связанной с событиями, в то время как снижение называется десинхронизацией, связанной с событиями. [ 62 ]
Сброс фазы
[ редактировать ]Сброс фазы происходит, когда вход в нейронный или нейрональный ансамбль сбрасывает фазу текущих колебаний. [ 63 ] Это очень часто встречается в отдельных нейронах, где сроки всплеска корректируются с учетом нейронального ввода (нейрон может вспыхивать при фиксированной задержке в ответ на периодический вход, который называется фазовой блокировкой [ 19 ] ) и может также происходить в нейрональных ансамблях, когда фазы их нейронов регулируются одновременно. Сброс фазы является фундаментальным для синхронизации различных нейронов или различных областей мозга [ 18 ] [ 38 ] Потому что сроки шипов могут стать фазой, заблокированными для активности других нейронов.
Сброс фазы также позволяет изучать вызванную активность, термин, используемый при электроэнцефалографии и магнитоэнцефалографии для ответов в активности мозга, которые непосредственно связаны с активностью, связанной с стимулом . Вызванные потенциалы и потенциалы, связанные с событиями, получают из электроэнцефалограммы путем усреднения в среднем, т.е. в среднем различных испытаний при фиксированных задержек вокруг представления стимула. Как следствие, эти компоненты сигнала, которые одинаковы в каждом отдельном измерении, сохраняются, и все остальные, т.е. непрерывная или спонтанная активность, усредняются. То есть, связанные с событиями потенциалы отражают только колебания в мозговой активности, которые связаны с фазой для стимула или события. Призывая активность часто считается независимой от продолжающейся мозговой активности, хотя это продолжающаяся дебаты. [ 64 ] [ 65 ]
Асимметричная амплитуда модуляция
[ редактировать ]Недавно было предложено, что даже если фазы не выровнены в разных исследованиях, индуцированная активность может по-прежнему вызывать связанные с событиями потенциалы, потому что текущие колебания мозга могут не быть симметричными, и, следовательно, амплитудные модуляции могут привести к базовому сдвигу, который не в среднем. [ 66 ] [ 67 ] Эта модель подразумевает, что медленные ответы, связанные с событиями, такие как асимметричная альфа-активность, могут быть результатом асимметричных модуляций амплитуды колебаний мозга, таких как асимметрия внутриклеточных токов, которые распространяют вперед и назад вниз по дендритам. [ 68 ] При этом предположении асимметрия в дендритном токе будет вызывать асимметрию в колебательной активности, измеренной ЭЭГ и МЭГ, поскольку, как правило, дендритные токи в пирамидальных клетках генерируют сигналы ЭЭГ и МЕГ, которые могут быть измерены на коже. [ 69 ]
Межчастотная связь
[ редактировать ]Поперечная связь (CFC) описывает связь (статистическая корреляция) между медленной волной и быстрой волной. Есть много видов, обычно написанных как соединение AB, а это означает, что A медленной волны связана с B быстрой волны. Например, фазовая амплитудная связь - это то, где фаза медленной волны сочетается с амплитудой быстрой волны. [ 70 ]
Тета -гамма-код -это связь между тета-волной и гамма-волной в сети гиппокампа. Во время тета-волны от 4 до 8 непересекающихся ансамблей нейронов активируются последовательно. Это было предположительно, чтобы сформировать нейронный код, представляющий несколько элементов во временной рамке [ 71 ] [ 72 ]
Функция
[ редактировать ]Нейронная синхронизация может быть модулирована с помощью ограничений задач, таких как внимание , и, как полагают, играет роль в связывании признаков , [ 73 ] нейрональная связь , [ 7 ] и моторная координация . [ 9 ] Нейрональные колебания стали горячей темой в нейробиологии в 1990 -х годах, когда исследования визуальной системы мозга от Grey, Singer и других, по -видимому, поддерживали гипотезу связывания нервного нерва . [ 74 ] Согласно этой идее, синхронные колебания в нейрональных ансамблях связывают нейроны, представляющие различные признаки объекта. Например, когда человек смотрит на дерево, нейроны визуальной коры, представляющие ствол дерева, и те, которые представляют ветви того же дерева, будут колебаться в синхронности, образуя единое представление дерева. Это явление лучше всего проявляется в локальных полевых потенциалах , которые отражают синхронную активность локальных групп нейронов, но также были показаны в записях ЭЭГ и MEG , обеспечивающих все больше доказательств для тесной связи между синхронной колебательной активностью и различными когнитивными функциями, такими как перцепция группировка [ 73 ] и внимание к сверху вниз. [ 15 ] [ 13 ] [ 12 ]
Кардиостимулятор
[ редактировать ]Клетки в синоатриальном узле , расположенные в правом предсердии сердца, спонтанно деполяризует примерно 100 раз в минуту. Хотя все клетки сердца обладают способностью генерировать потенциалы действия, которые вызывают сокращение сердца, синоатриальный узел обычно инициирует его, просто потому, что он генерирует импульсы немного быстрее, чем другие области. Следовательно, эти клетки генерируют нормальный ритм пазухи и называются клетками кардиостимулятора, поскольку они непосредственно контролируют частоту сердечных сокращений . В отсутствие внешнего нейронного и гормонального контроля клетки в узле SA будут ритмично разряжаться. Синоатриальный узел богато иннервируется вегетативной нервной системой , которая вверх или вниз регулирует частоту спонтанного стрельбы клеток кардиостимулятора.
Центральный генератор шаблонов
[ редактировать ]Синхронизированная стрельба нейронов также образует основу периодических моторных команд для ритмических движений. Эти ритмические выходы производятся группой взаимодействующих нейронов, которые образуют сеть, называемую генератором центрального рисунка . Центральные генераторы рисунков - это нейрональные цепи, которые - когда активируются - могут производить ритмические моторные паттерны при отсутствии сенсорных или нисходящих входов, которые содержат определенную информацию о времени. Примерами являются ходьба , дыхание и плавание , [ 75 ] Большинство доказательств генераторов центральных рисунков поступают от низших животных, таких как миног , но есть также свидетельство генераторов центральных моделей позвоночника у людей. [ 76 ] [ 77 ]
Обработка информации
[ редактировать ]Нейрональный всплеск обычно считается основой для переноса информации в мозге. Для такой передачи информация должна быть закодирована в схеме пика. Были предложены различные типы схем кодирования, такие как кодирование скорости и временное кодирование . Нейронные колебания могут создавать периодические временные окна, в которых входные шипы оказывают большее влияние на нейроны, тем самым обеспечивая механизм для декодирования временных кодов. [ 78 ]
Временное декодирование
[ редактировать ]Внутренние одноклеточные генераторы служат ценными инструментами для декодирования, кодируемой временно, сенсорной информации. Эта информация кодируется через интервалы международных интервалов, и внутренние генераторы могут выступать в качестве «временных правителей» для точного измерения этих интервалов. Одним из заметных механизма для достижения этого является петля нейрональной фазы, закрепленную (NPLL). В этом механизме корковые генераторы подвергаются модуляции, на которую влияют скорости стрельбы таламокортикальных «фазовых детекторов», которые, в свою очередь, оценивают несоответствие между корковой и сенсорной периодичностью. [ 79 ]
Восприятие
[ редактировать ]Синхронизация увольнения нейронов может служить средством для группировки пространственно сегрегированных нейронов, которые реагируют на тот же стимул, чтобы связывать эти ответы для дальнейшей совместной обработки, то есть для использования временной синхронности для кодирования отношений. Сначала были предложены чисто теоретические формулировки гипотезы о связывании по синхронности, были предложены, [ 80 ] Но впоследствии сообщалось об обширных экспериментальных данных, подтверждающих потенциальную роль синхронности как реляционного кодекса. [ 81 ]
Функциональная роль синхронизированной колебательной активности в мозге была в основном установлена в экспериментах, проведенных на бодрствующих котятах с несколькими электродами, имплантированными в зрительную кору. Эти эксперименты показали, что группы пространственно сегрегированных нейронов участвуют в синхронной колебательной активности при активации визуальными стимулами. Частота этих колебаний находилась в диапазоне 40 Гц и отличалась от периодической активации, вызванной решеткой, что позволяет предположить, что колебания и их синхронизация были вызваны внутренними нейрональными взаимодействиями. [ 81 ] Подобные результаты были показаны параллельно группой Экхорна, предоставляя дополнительные доказательства функциональной роли нервной синхронизации в связывании признаков. [ 82 ] С тех пор многочисленные исследования воспроизводили эти результаты и расширили их до различных методов, таких как ЭЭГ, предоставив широкие доказательства функциональной роли гамма -колебаний в визуальном восприятии.
Жиль Лоран и его коллеги показали, что колебательная синхронизация играет важную функциональную роль в восприятии запаха. Восприятие разных запахов приводит к тому, что различные подмножества нейронов стреляют в разные наборы колебательных циклов. [ 83 ] Эти колебания могут быть нарушены GABA блокатором Picrotoxin , [ 84 ] и нарушение колебательной синхронизации приводит к нарушению поведенческой дискриминации химически сходных одорантов в пчелах, [ 85 ] и на более похожие ответы на запахи в нижестоящих нейронах β-добычи. [ 86 ] Недавнее наблюдение за этой работой показало, что колебания создают периодические окна интеграции для клеток кеньон в корпусе грибов насекомых , так что входящие шипы из антенной доли являются более эффективными в активации клеток кеньонов только на определенных фазах колебательного цикла. [ 78 ]
Нейронные колебания также считаются вовлеченными в смысле времени [ 87 ] и в соматосенсорном восприятии. [ 88 ] Тем не менее, недавние результаты спорят против часовой функции кортикальных гамма-колебаний. [ 89 ]
Моторная координация
[ редактировать ]Уверяния обычно сообщаются в моторной системе. Pfurtscheller и коллеги обнаружили снижение колебаний альфа (8–12 Гц) и бета (13–30 Гц) в активности ЭЭГ , когда субъекты делали движение. [ 62 ] [ 90 ] Используя внутрикорятические записи, аналогичные изменения в колебательной активности были обнаружены в моторной коре, когда обезьяны выполняли моторные действия, которые требовали значительного внимания. [ 91 ] [ 92 ] Кроме того, колебания на уровне позвоночника становятся синхронизированными с бета-колебаниями в моторной коре во время постоянной мышечной активации, как определено с помощью когерентности кортико-мышечной . [ 93 ] [ 94 ] [ 95 ] Аналогично, мышечная активность различных мышц выявляет межмышечную когерентность на нескольких различных частотах, отражающих основную нервную схему , участвующую в координации двигательной системы . [ 96 ] [ 97 ]
Недавно было обнаружено, что корковые колебания распространяются в качестве движущихся волн по поверхности моторной коры вдоль доминирующих пространственных оси, характерных для локальной схемы моторной коры. [ 98 ] Было предложено, чтобы моторные команды в виде движущихся волн могут пространственно фильтровать нисходящими волокнами для избирательного контроля мышечной силы. [ 99 ] Моделирование показало, что продолжающаяся волновая активность в коре может вызвать устойчивую мышечную силу с физиологическими уровнями когерентности ЭЭГ-ЭМГ. [ 100 ]
Колебательные ритмы при 10 Гц были зарегистрированы в области мозга, называемой нижней оливкой , которая связана с мозжечкой. [ 20 ] Эти колебания также наблюдаются при моторном выходе физиологического тремора [ 101 ] и при выполнении медленных движений пальцев. [ 102 ] Эти результаты могут указывать на то, что человеческий мозг периодически контролирует непрерывные движения. В поддержку было показано, что эти прерывистости движения напрямую коррелируют с колебательной активностью в циклере-цитатно-кортикальной петле, которая может представлять собой нейронный механизм для прерывистого моторного контроля. [ 103 ]
Память
[ редактировать ]Нейронные колебания, в частности тета -активность, тесно связаны с функцией памяти. Тета-ритмы очень сильны в гиппокампе грызунов и энторинальной коре во время обучения и поиска памяти, и, как полагают, они жизненно важны для индукции долгосрочного потенцирования , потенциального клеточного механизма для обучения и памяти. Считается, что связь между тета и гамма -активностью имеет жизненно важное значение для функций памяти, включая эпизодическую память . [ 104 ] [ 105 ] Пряная координация односпальных пиков с локальными колебаниями тета связана с успешной формированием памяти у людей, поскольку более стереотипные вспышки предсказывают лучшую память. [ 106 ]
Сон и сознание
[ редактировать ]Сон представляет собой естественное повторяющееся состояние, характеризующееся уменьшенным или отсутствующим сознанием , и происходит в циклах быстрого движения глаз (REM) и нерапидного движения глаз (NREM). Стадии сна характеризуются спектральным содержанием ЭЭГ : например, стадия N1 относится к переходу мозга от альфа-волн (общих в состоянии бодрствования) к тета Присутствие дельта -волн. [ 107 ] Нормальный порядок стадий сна - N1 → N2 → N3 → N2 → REM. [ Цитация необходима ]
Разработка
[ редактировать ]Нейронные колебания могут играть роль в развитии нервного нерва. Например, , что волны сетчатки обладают свойствами, которые определяют раннюю связность схем и синапсов между клетками в сетчатке. считается [ 108 ]
Патология
[ редактировать ]

Конкретные виды нервных колебаний также могут появляться в патологических ситуациях, таких как болезнь Паркинсона или эпилепсия . Эти патологические колебания часто состоят из аберрантной версии нормального колебания. Например, одним из самых известных типов является всплеск и волновой колебание, которое типично для обобщенных или отсутствия эпилептических приступов, и которые напоминают нормальные колебания веретена во сне.
Тремор
[ редактировать ]Тремор является непроизвольным, несколько ритмичным, мышечным сокращением и расслаблением, связанным с движениями одной или нескольких частей тела. Это наиболее распространенное из всех непроизвольных движений и может влиять на руки, руки, глаза, лицо, голова, голосовые связки, туловище и ноги. Большинство толчков встречаются в руках. У некоторых людей тремор является симптомом другого неврологического расстройства. Было выявлено много разных форм тремора, таких как необходимый тремор или паркинсонское тремор. Утверждается, что тремор, вероятно, будут многофакторными по происхождению, с вкладами от нервных колебаний в центральных нервных системах, а также из периферических механизмов, таких как рефлекторные петлевые резонансы. [ 109 ]
Эпилепсия
[ редактировать ]Эпилепсия является распространенным хроническим неврологическим расстройством, характеризующимся судорогами . Эти приступы являются временными признаками и/или симптомами аномальной, чрезмерной или гиперсинхронной активности нейронов в мозге. [ 110 ]
Таламокортикальная дисритмия
[ редактировать ]В таламокортикальной дисритмии (TCD) нормальный таламокортикальный резонанс нарушается. Потеря ввода таламуса позволяет замедлить частоту таламо-кортикальной колонки в тета или дельта [ 111 ] TCD можно лечить нейрохирургическими методами, такими как таламотомия .
Приложения
[ редактировать ]Клинические конечные точки
[ редактировать ]Нейронные колебания чувствительны к нескольким препаратам, влияющим на активность мозга; Соответственно, биомаркеры , основанные на нейронных колебаниях, становятся вторичными конечными точками в клинических испытаниях и в количественных эффектах в преклинических исследованиях. Эти биомаркеры часто называют «биомаркерами ЭЭГ» или «нейрофизиологическими биомаркерами» и определяются количественно с использованием количественной электроэнцефалографии (QEEG) . Биомаркеры ЭЭГ могут быть извлечены из ЭЭГ с использованием инструментов инструментов с открытым исходным кодом .
Интерфейс мозга - Компьютер
[ редактировать ]Нейронные колебания были применены в качестве контрольного сигнала в различных интерфейсах мозга и коммутеров (BCIS). [ 112 ] Например, неинвазивный BCI может быть создан путем размещения электродов на коже головы, а затем измерения слабых электрических сигналов. Хотя индивидуальные нейроны не могут быть зарегистрированы через неинвазивную BCI, потому что череп сыт и размывает электромагнитные сигналы, колебательная активность все еще может быть надежно обнаружена. BCI был представлен Vidal в 1973 году [ 113 ] в качестве проблемы использования сигналов ЭЭГ для контроля объектов вне тела человека.
После вызова BCI в 1988 году альфа -ритм использовался в BCI на основе ритма мозга для контроля над физическим объектом, роботом. [ 114 ] [ 115 ] BCI на основе альфа -ритма был первым BCI для контроля робота. [ 116 ] [ 117 ] В частности, некоторые формы BCI позволяют пользователям управлять устройством, измеряя амплитуду колебательной активности в определенных полосах частот, включая MU и бета -ритмы.
Примеры
[ редактировать ]Не инклюзивный список видов колебательной активности, обнаруженный в центральной нервной системе:
- Альфа -волна
- Бета -волна
- Разрыв
- Сердечный цикл
- Дельта волна
- Эпилептический захват
- Гамма -волна
- Математическое моделирование электрофизиологической активности при эпилепсии
- MU WAVE
- PGO волны
- Таламокортикальные колебания
- Острые волновые комплексы
- Сон шпиндель
- Потенциальные колебания подпороговой мембраны
- Тета -волна
Смотрите также
[ редактировать ]- Кибернетика
- Теория динамических систем
- ЭЭГ -анализ
- Neurocybernetics
- Колебательная нейронная сеть
- Системная нейробиология
- Теталирование
- Сброс фазы в нейронах
Ссылки
[ редактировать ]- ^ Jump up to: а беременный Наполи, Николас Дж.; Демас, Мэтью; Стивенс, Чад Л.; Кеннеди, Келли Д.; Харривел, Анжела Р.; Барнс, Лора Э.; Папа, Алан Т. (2020-03-03). «Сложность активации: инструмент когнитивных нарушений для характеристики нейроизоляции» . Научные отчеты . 10 (1): 3909. Bibcode : 202020natsr..10.3909n . doi : 10.1038/s41598-020-60354-2 . ISSN 2045-2322 . PMC 7054256 . PMID 32127579 .
 Эта статья включает текст из этого источника, который доступен по лицензии CC по 4,0 .
Эта статья включает текст из этого источника, который доступен по лицензии CC по 4,0 .
- ^ Jump up to: а беременный в Ahissar, E. & Vaadia, E. колебательная активность отдельных единиц в соматосенсорной коре бодрствующей обезьяны и их возможная роль в анализе текстур. Proc Natl Acad Sci USA 87, 8935-8939 (1990).
- ^ Llinás RR (2014). «Внутренние электрические свойства нейронов млекопитающих и функции ЦНС: историческая перспектива» . Границы в клеточной нейробиологии . 8 : 320. DOI : 10.3389/fncel.2014.00320 . PMC 4219458 . PMID 25408634 .
- ^ «Катон, Ричард - электрические токи мозга» . echo.mpiwg-berlin.mpg.de . Получено 2018-12-21 .
- ^ Coenen A, Fine E, Zayachkivska O (2014). «Адольф Бек: забытый пионер в области электроэнцефалографии». Журнал истории нейронаук . 23 (3): 276–86. doi : 10.1080/0964704x.2013.867600 . PMID 24735457 . S2CID 205664545 .
- ^ Правдич-Неминский В.В. (1913). «Попытка зарегистрировать электрические симптомы мозга». Центральный лист для физиологии . 27 : 951–60.
- ^ Jump up to: а беременный Фри P (октябрь 2005 г.). «Механизм когнитивной динамики: нейрональная связь через нейрональную когерентность». Тенденции в когнитивных науках . 9 (10): 474–80. doi : 10.1016/j.tics.2005.08.011 . PMID 16150631 . S2CID 6275292 .
- ^ Fell J, Axmacher N (февраль 2011 г.). «Роль фазовой синхронизации в процессах памяти». Природные обзоры. Нейробиология . 12 (2): 105–18. doi : 10.1038/nrn2979 . PMID 21248789 . S2CID 7422401 .
- ^ Jump up to: а беременный Schnitzler A, Gross J (апрель 2005 г.). «Нормальная и патологическая колебательная связь в мозге». Природные обзоры. Нейробиология . 6 (4): 285–96. doi : 10.1038/nrn1650 . PMID 15803160 . S2CID 2749709 .
- ^ Baldauf D, Desimone R (апрель 2014 г.). «Нейронные механизмы объектного внимания» . Наука . 344 (6182): 424–7. Bibcode : 2014sci ... 344..424b . doi : 10.1126/science.1247003 . PMID 24763592 . S2CID 34728448 .
- ^ Foster JJ, Sutterer DW, Serens JT, Vogel EK, Awh E (июль 2017 г.). «Альфа-диапазоны позволяют пространственно и временно разрешить отслеживание скрытого пространственного внимания» . Психологическая наука . 28 (7): 929–941. doi : 10.1177/0956797617699167 . PMC 5675530 . PMID 28537480 .
- ^ Jump up to: а беременный De Vries IE, Marinato G, Baldauf D (август 2021 г.). «Декодирование объектного слухового внимания от источника, реконструированных Alpha-колебаний MEG» . Журнал нейробиологии . 41 (41): 8603–8617. doi : 10.1523/jneurosci.0583-21.2021 . PMC 8513695 . PMID 34429378 .
- ^ Jump up to: а беременный в Bagherzadeh Y, Baldauf D, Pantazis D, Desimone R (февраль 2020 г.). «Альфа -синхронность и контроль нейробиоуправления пространственного внимания» . Нейрон . 105 (3): 577–587.e5. doi : 10.1016/j.neuron.2019.11.001 . HDL : 11572/252726 . PMID 31812515 . S2CID 208614924 .
- ^ Berger H, Grey CM (1929). «О человеческой электрической фалограмме». Arch Psychiat Nervenkr . 87 : 527–570. Doi : 10.1007/bf01797193 . HDL : 11858/00-001M-0000-002A-5DE0-7 . S2CID 10835361 .
- ^ Jump up to: а беременный в Baldauf D, Desimone R (апрель 2014 г.). «Нейронные механизмы объектного внимания» . Наука . 344 (6182): 424–7. Bibcode : 2014sci ... 344..424b . doi : 10.1126/science.1247003 . PMID 24763592 . S2CID 34728448 .
- ^ Dement W, Kleitman N (ноябрь 1957 г.). «Циклические вариации в ЭЭГ во время сна и их связь с движениями глаз, подвижностью тела и сновидениями». Электроэнцефалография и клиническая нейрофизиология . 9 (4): 673–90. doi : 10.1016/0013-4694 (57) 90088-3 . PMID 13480240 .
- ^ Энгель А.К., Сингер В. (январь 2001 г.). «Временное связывание и нейронные корреляты сенсорного осознания». Тенденции в когнитивных науках . 5 (1): 16–25. doi : 10.1016/s1364-6613 (00) 01568-0 . PMID 11164732 . S2CID 11922975 .
- ^ Jump up to: а беременный в Varela F, Lachaux JP, Rodriguez E, Martinerie J (апрель 2001 г.). «BrainWeb: фазовая синхронизация и крупномасштабная интеграция». Природные обзоры. Нейробиология . 2 (4): 229–39. doi : 10.1038/35067550 . PMID 11283746 . S2CID 18651043 .
- ^ Jump up to: а беременный в Izhikevich EM (2007). Динамические системы в нейробиологии . Кембридж, Массачусетс: MIT Press.
- ^ Jump up to: а беременный Llinás R, Yarom Y (июль 1986 г.). «Колебные свойства нижних олиговых нейронов Гвинея и их фармакологическая модуляция: исследование in vitro» . Журнал физиологии . 376 : 163–82. doi : 10.1113/jphysiol.1986.sp016147 . PMC 1182792 . PMID 3795074 .
- ^ Mureşan RC, Jurjuţ OF, Moca VV, Singer W, Nikolić D (март 2008 г.). «Оценка колебаний: эффективный метод оценки силы колебаний в нейрональной активности». Журнал нейрофизиологии . 99 (3): 1333–53. doi : 10.1152/jn.00772.2007 . PMID 18160427 .
- ^ Burrow T (1943). «Невродинамика поведения. Филобиологическое предисловие». Философия науки . 10 (4): 271–288. doi : 10.1086/286819 . S2CID 121438105 .
- ^ Amari, Shun-Ichi (1977-06-01). «Динамика формирования паттернов в нервных полях типа бокового ингибирования» . Биологическая кибернетика . 27 (2): 77–87. doi : 10.1007/bf00337259 . ISSN 1432-0770 . PMID 911931 . S2CID 2811608 .
- ^ Schöner, G.; Келсо, Джас (1988-03-25). «Динамическое генерация шаблона в поведенческих и нейронных системах» . Наука . 239 (4847): 1513–1520. Bibcode : 1988sci ... 239.1513S . doi : 10.1126/science.3281253 . ISSN 0036-8075 . PMID 3281253 .
- ^ Vansteensel MJ, Pels EG, Bleichner MG, Branco MP, Denison T, Freudenburg ZV, et al. (Ноябрь 2016). «Полностью имплантированный интерфейс мозга-компьютер у запертого пациента с БАС» . Новая Англия Журнал медицины . 375 (21): 2060–2066. doi : 10.1056/nejmoa1608085 . HDL : 1874/344360 . PMC 5326682 . PMID 27959736 .
- ^ Jump up to: а беременный Haken H (1996). Принципы функционирования мозга . Спрингер. ISBN 978-3-540-58967-9 .
- ^ Jump up to: а беременный в Ван XJ (июль 2010 г.). «Нейрофизиологические и вычислительные принципы кортикальных ритмов в познании» . Физиологические обзоры . 90 (3): 1195–268. doi : 10.1152/physrev.00035.2008 . PMC 2923921 . PMID 20664082 .
- ^ Nunez PL, Srinivasan R (1981). Электрические поля мозга: нейрофизика ЭЭГ . Издательство Оксфордского университета. ISBN 9780195027969 .
- ^ Кардин Дж.А., Карлен М., Мелетис К., Кноблич У, Чжан Ф., Дейссерот К. и др. (Июнь 2009 г.). «Приводящие быстрорастущие клетки индуцируют гамма-ритм и контролируют сенсорные ответы» . Природа . 459 (7247): 663–7. Bibcode : 2009natur.459..663c . doi : 10.1038/nature08002 . PMC 3655711 . PMID 19396156 .
- ^ Llinás R, Ribary U, Crongras D, Pedroarena C (ноябрь 1998 г.). «Нейрональная основа для сознания» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 353 (1377): 1841–9. doi : 10.1098/rstb.1998.0336 . PMC 1692417 . PMID 9854256 .
- ^ Bollimunta A, Mo J, Schroeder CE, Ding M (март 2011 г.). «Нейрональные механизмы и модуляция внимания кортикоталамических α -колебаний» . Журнал нейробиологии . 31 (13): 4935–43. doi : 10.1523/jneurosci.5580-10.2011 . PMC 3505610 . PMID 21451032 .
- ^ Suffczynski P, Kalitzin S, Pfurtscheller G, Lopes Da Silva FH (декабрь 2001 г.). «Вычислительная модель таламо-кортикальных сетей: динамический контроль альфа-ритмов в отношении очагового внимания». Международный журнал психофизиологии . 43 (1): 25–40. doi : 10.1016/s0167-8760 (01) 00177-5 . PMID 11742683 .
- ^ Cabral J, Luckhoo H, Woolrich M, Joensson M, Mohseni H, Baker A, et al. (Апрель 2014). «Изучение механизмов спонтанной функциональной связности в MEG: как отсроченные сетевые взаимодействия приводят к структурированным амплитудным конвертам отфильтрованных колебаний с полосой» . Нейроамиж . 90 : 423–35. doi : 10.1016/j.neuroimage.2013.11.047 . HDL : 10230/23081 . PMID 24321555 .
- ^ Llinás RR (декабрь 1988 г.). «Внутренние электрофизиологические свойства нейронов млекопитающих: понимание функции центральной нервной системы». Наука . 242 (4886): 1654–64. Bibcode : 1988sci ... 242.1654L . doi : 10.1126/science.3059497 . PMID 3059497 .
- ^ Llinás RR, Grace AA, Yarom Y (февраль 1991 г.). «Нейроны in vitro в кортикальном слое млекопитающих 4 проявляют внутреннюю колебательную активность в диапазоне частот от 10 до 50 Гц» . Труды Национальной академии наук Соединенных Штатов Америки . 88 (3): 897–901. Bibcode : 1991pnas ... 88..897L . doi : 10.1073/pnas.88.3.897 . PMC 50921 . PMID 1992481 .
- ^ Ahissar, E., Haidarliu, S. и Zacksenhouse, M. (1997). Декодирование временно кодируемого сенсорного ввода с помощью корковых колебаний и таламических фазовых компараторов. Proc Natl Acad Sci USA 94, 11633-11638.
- ^ Zeitler M, Daffertshofer A, Gielen CC (июнь 2009 г.). «Асимметрия в осцилляторах с импульсом с задержкой» (PDF) . Физический обзор e . 79 (6 Pt 2): 065203. Bibcode : 2009frve..79f5203Z . doi : 10.1103/physreve.79.065203 . HDL : 1871/29169 . PMID 19658549 .
- ^ Jump up to: а беременный Пиковский А., Розенблум М., Куртс Дж. (2001). Синхронизация: универсальная концепция в нелинейных науках . Издательство Кембриджского университета. ISBN 978-0-521-53352-2 .
- ^ Андреа Бровелли, Стивен Л. Бресслер и их коллеги, 2004
- ^ Петр, Берки; Csaba, cserep; Zsuzsanna, Környei (2024). «Microglia способствует синхронности нейронов, несмотря на эндогенную фенотипическую трансформацию, связанную с АТФ, в острых срезах мозга мыши» . Природная связь . 15 doi : 10.1038/s41467-024-49773-1 . PMC 11208608 . PMID 38926390 .
- ^ Muthukumaraswamy SD, Edden RA, Jones DK, Swettenham JB, Singh KD (май 2009 г.). «Концентрация покоя GABA предсказывает пиковую гамма -частоту и амплитуду МРТ в ответ на визуальную стимуляцию у людей» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (20): 8356–61. Bibcode : 2009pnas..106.8356m . doi : 10.1073/pnas.0900728106 . PMC 2688873 . PMID 19416820 .
- ^ Moruzzi G, Magoun HW (ноябрь 1949 г.). «Стволовая образование ствола мозга и активация ЭЭГ» . Электроэнцефалография и клиническая нейрофизиология . 1 (4): 455–73. doi : 10.1016/0013-4694 (49) 90219-9 . PMID 18421835 .
- ^ Buzsáki G, Draguhn A (июнь 2004 г.). «Нейрональные колебания в корковых сетях». Наука . 304 (5679): 1926–9. Bibcode : 2004sci ... 304.1926b . doi : 10.1126/science.1099745 . PMID 15218136 . S2CID 8002293 .
- ^ Catterall, WA, Raman, IM, Robinson, HPC, Sejnowski, TJ, Paulsen, O. (2 октября 2012 г.). «Hodgkin-Huxley Heritage: от каналов до цепей» . Журнал нейробиологии . 32 (41): 14064–14073. doi : 10.1523/jneurosci.3403-12.2012 . PMC 3500626 . PMID 23055474 .
- ^ Lapicque, LM (1907). «Количественные исследования электрического возбуждения нервов». J Physiol Paris . 9 : 620–635.
- ^ Эбботт, Ларри (1999). «Внедрение Lapicque модели интегрирования и огня (1907)». Бюллетень исследования мозга . 50 (5): 303–304. doi : 10.1016/s0361-9230 (99) 00161-6 . PMID 10643408 . S2CID 46170924 .
- ^ Уиттингтон М.А., Трауб Р.Д., Копелл Н., Эрментрут Б., Буль Э.Х. (декабрь 2000 г.). «Ритмы на основе ингибирования: экспериментальные и математические наблюдения по динамике сети». Международный журнал психофизиологии . 38 (3): 315–36. Citeseerx 10.1.1.16.6410 . doi : 10.1016/s0167-8760 (00) 00173-2 . PMID 11102670 .
- ^ Goldental A, Vardi R, Sardi S, Sabo P, Kanter I (2015). «Широкополосные макроскопические колебания коры возникают из -за внутренних сбоев нейрональных ответов» . Границы в нейронных цепях . 9 : 65. Arxiv : 1511.00235 . doi : 10.3389/fncir.2015.00065 . PMC 4626558 . PMID 26578893 .
- ^ Wendling F, Bellanger JJ, Bartolomei F, Chauvel P (октябрь 2000 г.). «Актуальность нелинейных моделей с смешным параметром при анализе эпилептических сигналов глубины-EEG». Биологическая кибернетика . 83 (4): 367–78. doi : 10.1007/s004220000160 . PMID 11039701 . S2CID 8751526 .
- ^ Bressloff PC, Cowan JD (2003) Спонтанное образование рисунка в первичной зрительной коре. В: J Hogan, Ar Krauskopf, M Di Bernado, Re Wilson (Eds.), Нелинейная динамика и хаос: куда мы идем отсюда?
- ^ Kuramoto Y (1984). Химические колебания, волны и турбулентность . Dover Publications.
- ^ Ermentrout B (1994). «Введение в нервные генераторы». В F Ventriglia (ред.). Нейронные моделирование и нейронные сети . С. 79–110.
- ^ Breakspear M, Heitmann S, Daffertshofer A (2010). «Генеративные модели коры колебаний: нейробиологические последствия модели Курамото» . Границы в нейробиологии человека . 4 : 190. doi : 10.3389/fnhum.2010.00190 . PMC 2995481 . PMID 21151358 .
- ^ Cabral J, Hugues E, Sporns O, Deco G (июль 2011 г.). «Роль локальных колебаний сети в функциональном подключении в состоянии покоя» . Нейроамиж . 57 (1): 130–139. doi : 10.1016/j.neuroimage.2011.04.010 . PMID 21511044 . S2CID 13959959 .
- ^ Фрейер Ф., Акино К., Робинсон П.А., Риттер П., Брейспир М (июль 2009 г.). «Бистатичность и негауссовые колебания в спонтанной корковой активности» . Журнал нейробиологии . 29 (26): 8512–24. doi : 10.1523/jneurosci.0754-09.2009 . PMC 6665653 . PMID 19571142 .
- ^ Fox MD, Raichle Me (сентябрь 2007 г.). «Спонтанные колебания активности мозга, наблюдаемые при функциональной магнитно -резонансной томографии». Природные обзоры. Нейробиология . 8 (9): 700–11. doi : 10.1038/nrn2201 . PMID 17704812 . S2CID 15979590 .
- ^ Laufs H, Krakow K, Sterzer P, Eger E, Beyerle A, Salek-Haddadi A, Kleinschmidt A (сентябрь 2003 г.). «Электроэнцефалографические сигнатуры мод внимания и когнитивных по умолчанию в спонтанных колебаниях активности мозга в покое» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (19): 11053–8. Bibcode : 2003pnas..10011053L . doi : 10.1073/pnas.1831638100 . PMC 196925 . PMID 12958209 .
- ^ Мэтьюсон К.Е., Граттон Г., Фабиани М., Бек Д.М., Ро Т (март 2009 г.). «Чтобы увидеть или не видеть: Prestimulus Alpha Phase предсказывает визуальное осознание» . Журнал нейробиологии . 29 (9): 2725–32. doi : 10.1523/jneurosci.3963-08.2009 . PMC 2724892 . PMID 19261866 .
- ^ Busch NA, Dubois J, Vanrullen R (июнь 2009 г.). «Фаза текущих колебаний ЭЭГ предсказывает визуальное восприятие» . Журнал нейробиологии . 29 (24): 7869–76. doi : 10.1523/jneurosci.0113-09.2009 . PMC 6665641 . PMID 19535598 .
- ^ Van Dijk H, Schoffelen JM, Oostenveld R, Jensen O (февраль 2008 г.). «Претимул колебательная активность в альфа -полосе предсказывает способность визуальной дискриминации» . Журнал нейробиологии . 28 (8): 1816–23. doi : 10.1523/jneurosci.1853-07.2008 . PMC 6671447 . PMID 18287498 .
- ^ Таллон-Бодри С., Бертран О (апрель 1999 г.). «Колебательная гамма -активность у людей и ее роль в представлении объектов». Тенденции в когнитивных науках . 3 (4): 151–162. doi : 10.1016/s1364-6613 (99) 01299-1 . PMID 10322469 . S2CID 1308261 .
- ^ Jump up to: а беременный Pfurtscheller G, Lopes Da Silva FH (ноябрь 1999 г.). «Связанная с событиями синхронизация и десинхронизация ЭЭГ/МЕГ: основные принципы». Клиническая нейрофизиология . 110 (11): 1842–57. doi : 10.1016/s1388-2457 (99) 00141-8 . PMID 10576479 . S2CID 24756702 .
- ^ Tass PA (2007). Сброс фазы в медицине и биологии: стохастическое моделирование и анализ данных . Берлин Хейдельберг: Springer-Verlag. ISBN 978-3-540-65697-5 .
- ^ Makeig S, Westerfield M, Jung TP, Enghoff S, Townsend J, Couchesne E, Sejnowski TJ (январь 2002 г.). «Динамические источники мозга визуальных реакций». Наука . 295 (5555): 690–4. Bibcode : 2002sci ... 295..690M . doi : 10.1126/science.1066168 . PMID 11809976 . S2CID 15200185 .
- ^ Mäkinen V, Tiitinen H, May P (февраль 2005 г.). «Ответы, связанные с слуховыми событиями, генерируются независимо от продолжающейся мозговой деятельности». Нейроамиж . 24 (4): 961–8. doi : 10.1016/j.neuroimage.2004.10.020 . PMID 15670673 . S2CID 16210275 .
- ^ Никулин В.В., Линкенкаер-Хансен К., Нолте Г., Лемм С., Мюллер К.Р. , Илмомими Р.Дж., Курйо Г. (май 2007). «Новый механизм вызванных реакций в человеческом мозге». Европейский журнал нейробиологии . 25 (10): 3146–54. doi : 10.1111/j.1460-9568.2007.05553.x . PMID 17561828 . S2CID 12113334 .
- ^ Mazaheri A, Jensen O (июль 2008 г.). «Асимметричные амплитудные модуляции колебаний мозга генерируют медленные вызванные реакции» . Журнал нейробиологии . 28 (31): 7781–7. doi : 10.1523/jneurosci.1631-08.2008 . PMC 6670375 . PMID 18667610 .
- ^ Mazaheri A, Jensen O (2008). «Ритмичный пульсирующий: связывание продолжающейся мозговой активности с вызванными реакциями» . Границы в нейробиологии человека . 4 : 177. DOI : 10.3389/fnhum.2010.00177 . PMC 2972683 . PMID 21060804 .
- ^ Hamalainen M, Hari R, Ilmoniemi RJ, Knuutila J, Lounasmaa OV (1993). «Магнитоэнцефалография - теория, инструментация и применение к неинвазивным исследованиям работающего человеческого мозга» . Rev Mod Phys . 65 (2): 413–497. Bibcode : 1993rvmp ... 65..413H . doi : 10.1103/revmodphys.65.413 .
- ^ Хайафил, Александр; Жиро, Энн-Лиз; Фонтолан, Лоренцо; Gutkin, Boris (2015-11-01). «Нейронная перекрестная связь: соединение архитектур, механизмов и функций» . Тенденции в нейронауках . 38 (11): 725–740. doi : 10.1016/j.tins.2015.09.001 . ISSN 0166-2236 . PMID 26549886 . S2CID 3545001 .
- ^ Лисман, Джон Э.; Дженсен, Оле (2013-03-20). «Тета-гаммайский нейронный код» . Нейрон . 77 (6): 1002–1016. doi : 10.1016/j.neuron.2013.03.007 . ISSN 0896-6273 . PMC 3648857 . PMID 23522038 .
- ^ Drieu, Céline; Zugaro, Michaël (2019). «Последовательности гиппокампа во время исследования: механизмы и функции» . Границы в клеточной нейробиологии . 13 : 232. DOI : 10.3389/fncel.2019.00232 . ISSN 1662-5102 . PMC 6584963 . PMID 31263399 .
- ^ Jump up to: а беременный Singer W (1993). «Синхронизация кортикальной активности и ее предполагаемая роль в обработке и обучении информации». Ежегодный обзор физиологии . 55 : 349–74. doi : 10.1146/annurev.ph.55.030193.002025 . PMID 8466179 .
- ^ Singer W, Grey CM (1995). «Интеграция визуальной особенности и гипотеза временной корреляции». Ежегодный обзор нейробиологии . 18 : 555–86. Citeseerx 10.1.1.308.6735 . doi : 10.1146/annurev.ne.18.030195.003011 . PMID 7605074 .
- ^ Marder E, Bucher D (ноябрь 2001 г.). «Центральные генераторы рисунков и управление ритмическими движениями» . Текущая биология . 11 (23): R986-96. Bibcode : 2001cbio ... 11.r986m . doi : 10.1016/s0960-9822 (01) 00581-4 . PMID 11728329 . S2CID 1294374 .
- ^ Dimitrijevic MR, Gerasimenko Y, Pinter MM (ноябрь 1998). «Свидетельство о генераторе центрального паттерна позвоночника у людей». Анналы нью -йоркской академии наук . 860 (1): 360–76. Bibcode : 1998nyasa.860..360d . doi : 10.1111/j.1749-6632.1998.tb09062.x . PMID 9928325 . S2CID 102514 .
- ^ Danner SM, Hofstoetter US, Freundl B, Binder H, Mayr W, Rattay F, Minassian K (март 2015 г.). «Городской позвоночный двигатель контроль основан на гибко организованных генераторах взрыва» . Мозг . 138 (Pt 3): 577–88. doi : 10.1093/brain/awu372 . PMC 4408427 . PMID 25582580 .
- ^ Jump up to: а беременный Гупта Н., Сингх С.С., Стопфер М (декабрь 2016 г.). «Окна колебательных интеграций в нейронах» . Природная связь . 7 : 13808. Bibcode : 2016natco ... 713808g . doi : 10.1038/ncomms13808 . PMC 5171764 . PMID 27976720 .
- ^ Ahissar, E., Nelinger, G., Assa, E., Karp, O. & Saraf-Sinik, I. Thalamocortical Cyples как височные демодуляторы в чувствах. Биология связи 6, 562 (2023).
- ^ Милнер премьер -министр (ноябрь 1974 г.). «Модель для распознавания визуальной формы». Психологический обзор . 81 (6): 521–35. doi : 10.1037/h0037149 . PMID 4445414 .
- ^ Jump up to: а беременный Грей К.М., Кениг П., Энгель А.К., Сингер В (март 1989 г.). «Колебные ответы в визуальной коре кошки демонстрируют межколонную синхронизацию, которая отражает глобальные свойства стимула». Природа . 338 (6213): 334–7. Bibcode : 1989natur.338..334G . doi : 10.1038/338334a0 . PMID 2922061 . S2CID 4281744 .
- ^ Экхорн Р., Бауэр Р., Джордан В., Брош М., Крузе В., Мунк М., Рейтбук Х.Дж. (1988). «Когерентные колебания: механизм функции, связывающего в зрительной коре? Многократный электрод и анализ корреляции в кошке». Биологическая кибернетика . 60 (2): 121–30. doi : 10.1007/bf00202899 . PMID 3228555 . S2CID 206771651 .
- ^ Wehr M, Laurent G (ноябрь 1996 г.). «Кодирование запаха временными последовательностями стрельбы в колеблющихся нейронных сбоях». Природа . 384 (6605): 162–6. Bibcode : 1996natur.384..162W . doi : 10.1038/384162A0 . PMID 8906790 . S2CID 4286308 .
- ^ Macleod K, Laurent G (ноябрь 1996 г.). «Отдельные механизмы синхронизации и временного паттерна нейронных сборов, кодирующих запах». Наука . 274 (5289): 976–9. Bibcode : 1996sci ... 274..976m . doi : 10.1126/science.274.5289.976 . PMID 8875938 . S2CID 10744144 .
- ^ Stopfer M, Bhagavan S, Smith BH, Laurent G (ноябрь 1997 г.). «Обнаруженная дискриминация запаха в отношении десинхронизации нейронных собраний, кодирующих одур». Природа . 390 (6655): 70–4. Bibcode : 1997natur.390 ... 70S . doi : 10.1038/36335 . PMID 9363891 . S2CID 205024830 .
- ^ Macleod K, Bäcker A, Laurent G (октябрь 1998 г.). «Кто читает временную информацию, содержащуюся в синхронизированных и колебательных поездах шипа?». Природа . 395 (6703): 693–8. Bibcode : 1998natur.395..693m . doi : 10.1038/27201 . PMID 9790189 . S2CID 4424801 .
- ^ Buhusi CV, Meck WH (октябрь 2005 г.). «Что заставляет нас тикать? Функциональные и нервные механизмы интервального времени». Природные обзоры. Нейробиология . 6 (10): 755–65. doi : 10.1038/nrn1764 . PMID 16163383 . S2CID 29616055 .
- ^ Ахиссар Е., Заксенхаус М. (2001). «Глава 6 Временное и пространственное кодирование в системе колебаний крысы». Достижения в области кодирования нейронного населения . Прогресс в исследовании мозга. Тол. 130. С. 75–87. doi : 10.1016/s0079-6123 (01) 30007-9 . ISBN 9780444501103 Полем PMID 11480290 .
{{cite book}}:|journal=игнорируется ( помощь ) - ^ Burns SP, Xing D, Shapley RM (июнь 2011 г.). «Является ли активность гамма-диапазона в локальном потенциале поля коры V1« часы »или отфильтрованный шум?» Полем Журнал нейробиологии . 31 (26): 9658–64. doi : 10.1523/jneurosci.0660-11.2011 . PMC 3518456 . PMID 21715631 .
- ^ Pfurtscheller G, Aranibar A (июнь 1977 г.). «Связанная с событием кортикальная десинхронизация, обнаруженная путем измерения мощности ЭЭГ скальпа». Электроэнцефалография и клиническая нейрофизиология . 42 (6): 817–26. doi : 10.1016/0013-4694 (77) 90235-8 . PMID 67933 .
- ^ Мурти Вн, Фетц Е.Е. (декабрь 1996 г.). «Колебательная активность в сенсомоторной коре бодрствующих обезьян: синхронизация локальных потенциалов поля и связь с поведением». Журнал нейрофизиологии . 76 (6): 3949–67. doi : 10.1152/jn.1996.76.6.3949 . PMID 8985892 .
- ^ SANES JN, Donoghue JP (май 1993). «Колебания в локальном полевом потенциале моторной коры приматов во время добровольного движения» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (10): 4470–4. Bibcode : 1993pnas ... 90.4470s . doi : 10.1073/pnas.90.10.4470 . PMC 46533 . PMID 8506287 .
- ^ Конвей Б.А., Хэллидей Д.М., Фармер С.Ф., Шахани У, Маас П., Вейр А.И., Розенберг младший (декабрь 1995 г.). «Синхронизация между моторной корой и пул позвоночника мотонейронального бассейна во время выполнения поддерживаемой моторной задачи у человека» . Журнал физиологии . 489 (Pt 3) (3): 917–24. doi : 10.1113/jphysiol.1995.sp021104 . PMC 1156860 . PMID 8788955 .
- ^ Салениус С., Портин К, Каджола М., Салмелин Р., Хари Р (июнь 1997 г.). «Корковый контроль над мотонейроном человеческого мотонейрона во время изометрического сокращения». Журнал нейрофизиологии . 77 (6): 3401–5. doi : 10.1152/jn.1997.77.6.3401 . PMID 9212286 . S2CID 2178927 .
- ^ Бейкер С.Н., Оливье Э., Лимон Р.Н. (май 1997). «Когерентные колебания в коре моторной коры обезьяны и мышцы ручной мышцы показывают зависимую от задачи модуляцию» . Журнал физиологии . 501 (Pt 1) (1): 225–41. doi : 10.1111/j.1469-7793.1997.225bo.x . PMC 1159515 . PMID 9175005 .
- ^ Boostra TW, Danna-Dos-Santos A, Xie HB, Roerdink M, Stins JF, Breakspear M (декабрь 2015 г.). «Мышечные сети: анализ связи активности ЭМГ во время постурального контроля» . Научные отчеты . 5 : 17830. Bibcode : 2015natsr ... 517830b . doi : 10.1038/srep17830 . PMC 4669476 . PMID 26634293 .
- ^ Керкман Дж.Н., Даффертсхофер А., Голло Л.Л., Брейспир М., Бустерна Т.В. (июнь 2018 г.). «Сетчатая структура человеческой опорно -двигательной системы формирует нервные взаимодействия в нескольких временных масштабах» . Наука достижения . 4 (6): EAAT0497. Bibcode : 2018scia .... 4..497K . doi : 10.1126/sciadv.aat0497 . PMC 6021138 . PMID 29963631 .
- ^ Рубино Д., Роббинс К.А., Хасопулос Нг (декабрь 2006 г.). «Распространенные волны опосредуют передачу информации в моторной коре». Nature Neuroscience . 9 (12): 1549–57. doi : 10.1038/nn1802 . PMID 17115042 . S2CID 16430438 .
- ^ Heitmann S, Boostra T, Gong P, Breakspear M, Ermentrout B (2015). «Ритмы устойчивой осанки: моторные команды как пространственно организованные шаблоны колебаний». Нейрокомпьютинг . 170 : 3–14. doi : 10.1016/j.neucom.2015.01.088 .
- ^ Heitmann S, Boostra T, Breakspear M (октябрь 2013 г.). «Дендритный механизм для декодирования путешествующих волн: принципы и применение к моторной коре» . PLOS Computational Biology . 9 (10): E1003260. BIBCODE : 2013PLSCB ... 9E3260H . doi : 10.1371/journal.pcbi.1003260 . PMC 3814333 . PMID 24204220 .
- ^ Allum JH, Dietz V, Freund HJ (май 1978). «Нейрональные механизмы, лежащие в основе физиологического тремора». Журнал нейрофизиологии . 41 (3): 557–71. doi : 10.1152/jn.1978.41.3.557 . PMID 660226 .
- ^ Vallbo AB, Wessberg J (сентябрь 1993 г.). «Организация моторного производства в медленных движениях пальцев в человеке» . Журнал физиологии . 469 : 673–91. doi : 10.1113/jphysiol.1993.sp019837 . PMC 1143894 . PMID 8271223 .
- ^ Гросс Дж., Тиммерманн Л., Куджала Дж., Диркс М., Шмитц Ф., Салмелин Р., Шницлер А (февраль 2002 г.). «Нейронная основа прерывистого моторного контроля у людей» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (4): 2299–302. Bibcode : 2002pnas ... 99.2299G . doi : 10.1073/pnas.032682099 . PMC 122359 . PMID 11854526 .
- ^ Buszaki G (2006). Ритмы мозга . Издательство Оксфордского университета.
- ^ Nyhus E, Curran T (июнь 2010 г.). «Функциональная роль гамма и колебаний тета в эпизодической памяти» . Нейробиологии и биобиоэвиральные обзоры . 34 (7): 1023–35. doi : 10.1016/j.neubiorev.2009.12.014 . PMC 2856712 . PMID 20060015 .
- ^ Rutishauser U, Ross IB, Mamelak AN, Schuman EM (апрель 2010 г.). «Сила памяти человека прогнозируется по тета-частотной фазовой блокировке отдельных нейронов» (PDF) . Природа . 464 (7290): 903–7. Bibcode : 2010natur.464..903r . doi : 10.1038/nature08860 . PMID 20336071 . S2CID 4417989 .
- ^ Бранкаччо А., Табарелли Д., Бигика М., Балдауф Д. (апрель 2020 г.). «Локализация источника коры с удельной колебательной активностью коры» . Научные отчеты . 10 (1): 6976. Bibcode : 202020natsr..10.6976b . doi : 10.1038/s41598-020-63933-5 . PMC 7181624 . PMID 32332806 .
- ^ Feller MB (июль 2009 г.). «Волны сетчатки, вероятно, будут обучать образованию специфичных для глаз ретиногеногентных проекций» . Нейронное развитие . 4 : 24. DOI : 10.1186/1749-8104-4-24 . PMC 2706239 . PMID 19580682 .
- ^ Макаули Дж.Х., Марсден CD (август 2000 г.). «Физиологические и патологические тремор и ритмичный центральный моторный контроль» . Мозг . 123 (Pt 8) (8): 1545–67. doi : 10.1093/мозг/123,8.1545 . PMID 10908186 .
- ^ Shusterman V, Troy WC (июнь 2008 г.). «От исходной до эпилептиформной активности: путь к синхронизированной ритмичности в крупномасштабных нейронных сетях». Физический обзор e . 77 (6 pt 1): 061911. Bibcode : 2008frve..77f1911s . doi : 10.1103/physreve.77.061911 . PMID 18643304 . S2CID 13928602 .
- ^ Vanneste S, Song JJ, De Ridder D (март 2018 г.). «Таламокортикальная дисритмия, обнаруженная путем машинного обучения» . Природная связь . 9 (1): 1103. Bibcode : 2018natco ... 9.1103V . doi : 10.1038/s41467-018-02820-0 . PMC 5856824 . PMID 29549239 .
- ^ Birbaumer N (ноябрь 2006 г.). «Разрушение тишины: интерфейсы мозга-компьютера (BCI) для связи и моторного управления» . Психофизиология . 43 (6): 517–32. doi : 10.1111/j.1469-8986.2006.00456.x . PMID 17076808 .
- ^ Видал Дж.Дж. (1973). «На пути к прямой коммуникации мозга» . Ежегодный обзор биофизики и биоинженерии . 2 : 157–80. doi : 10.1146/annurev.bb.02.060173.001105 . PMID 4583653 .
- ^ Bozinovski S, Sestakov M, Bozinovska L (ноябрь 1988). «Использование альфа -ритма ЭЭГ для управления мобильным роботом». Материалы ежегодной Международной конференции Общества инженерии IEEE в области медицины и биологии . Новый Орлеан: IEEE. С. 1515–1516. doi : 10.1109/iembs.1988.95357 . ISBN 0-7803-0785-2 Полем S2CID 62179588 .
- ^ Bozinovski S (август 1990). «Управление траекторией мобильных роботов: от фиксированных рельсов до прямого биоэлектрического контроля». В Кайнак О (ред.). Материалы Международного семинара IEEE по интеллектуальному управлению движением . Тол. 2. Стамбул: IEEE. С. 463–467. doi : 10.1109/imc.1990.687362 . S2CID 60642344 .
- ^ Лебедев М. (2016). «Увеличение сенсомоторных функций с нейронными протезами». Opera Medica et physiologica . 2 (3–4): 211–227. doi : 10.20388/omp.003.0035 (неактивный 2024-09-12).
{{cite journal}}: CS1 Maint: doi неактивен по состоянию на сентябрь 2024 года ( ссылка ) - ^ Лебедев Ма, Николелис Ма (апрель 2017 г.). «Интерфейсы мозговых машин: от основной науки до нейропротезы и нейрореабилитации». Физиологические обзоры . 97 (2): 767–837. doi : 10.1152/physrev.00027.2016 . PMID 28275048 .
Дальнейшее чтение
[ редактировать ]- Бузсаки Г. (2006). Ритмы мозга . Издательство Оксфордского университета. ISBN 978-0-19-530106-9 .
- Freeman W (1975). Массовое действие в нервной системе . Академическая пресса. ISBN 978-0124120471 Полем Архивировано с оригинала 2015-07-05.