MHC Class I.
| MHC Class I. | |
|---|---|
 Схематическое представление MHC класса I | |
| Идентификаторы | |
| Символ | MHC Class I. |
| Мембраном | 63 |
Молекулы MHC класса I являются одним из двух первичных классов молекул основного комплекса гистосовместимости (MHC) (другой, представляющий собой MHC Class II ) и обнаружены на клеточной поверхности всех зарожденных клеток в телах позвоночных . [ 1 ] [ 2 ] Они также встречаются на тромбоцитах , но не на эритроцитах . Их функция состоит в том, чтобы отображать пептидные фрагменты белков изнутри клетки в цитотоксические Т -клетки ; Это вызовет немедленную реакцию от иммунной системы на конкретный анти-анти-антиген, отображаемый с помощью белка класса I MHC. Поскольку молекулы MHC класса I представляют пептиды, полученные из цитозольных белков, путь представления MHC класса I часто называют цитозольным или эндогенным путем . [ 3 ]
У людей HLA , соответствующие MHC классу I, являются HLA-A , HLA-B и HLA-C .
Функция
[ редактировать ]Молекулы MHC класса I связывают пептиды, генерируемые в основном из -за деградации цитозольных белков протеасомой . MHC I: пептидный комплекс затем вставляется через эндоплазматический ретикулум во внешнюю плазматическую мембрану клетки. Эпитопный пептид связан с внеклеточными частями молекулы MHC класса I. Таким образом, функция MHC класса I состоит в том, чтобы показать внутриклеточные белки в цитотоксические Т -клетки (CTL). Тем не менее, MHC класса I также может представлять пептиды, полученные из экзогенных белков, в процессе, известном как перекрестная презентация .
Нормальная ячейка будет отображать пептиды от нормального текучести клеточного белка на своем MHC класса I, а CTL не будут активированы в ответ на них из -за центральных и периферических механизмов толерантности. Когда клетка экспрессирует инородные белки, такие как после вирусной инфекции, доля MHC класса I будет отображать эти пептиды на клеточной поверхности. Следовательно, CTLS, специфичные для MHC: пептидный комплекс, будут распознавать и убивать представленные клетки.
В качестве альтернативы, сам MHC класса может служить ингибирующим лигандом для естественных клеток -убийц (NKS). Снижение нормальных уровней MHC класса I, механизм, используемый некоторыми вирусами [ 4 ] и определенные опухоли, чтобы уклониться от ответов CTL, активирует убийство NK -клеток.
PIRB и визуальная пластичность
[ редактировать ]Парный иммуноглобулиноподобный рецептор B (PIRB), MHCI-связывающий рецептор, участвует в регуляции визуальной пластичности . [ 5 ] PIRB экспрессируется в центральной нервной системе и уменьшает глазного доминирования пластичность развития в критический период и взрослый возраст. [ 5 ] Когда функция PIRB была отменена у мышей -мутантов, глазного доминирования пластичность стала более выраженной в любом возрасте. [ 5 ] Потеря функциональных мышей PIRB также демонстрировала повышенную пластичность после монокулярной депривации в течение критического периода . [ 5 ] Эти результаты предполагают, что PIRB может участвовать в модуляции синаптической пластичности в зрительной коре .
Структура
[ редактировать ]Молекулы MHC класса I представляют собой гетеродимеры, которые состоят из двух полипептидных цепей, α и β -2 -микроглобулина (B2M). Две цепи связаны нековалентно посредством взаимодействия B2M и α 3 домена. Только α-цепь полиморфна и кодируется геном HLA , в то время как субъединица B2M не является полиморфной и кодируется геном бета-2 микроглобулина . Домен α 3 представляет собой плазматическую мембрану-пропасть и взаимодействует с CD8 со-рецептором Т-клеток . Взаимодействие α 3 -CD8 удерживает молекулу MHC I на месте, в то время как рецептор Т -клеток (TCR) на поверхности цитотоксической Т -клеток связывает его α 1 -α 2 гетеродимерного лиганда и проверяет соединенный пептид на наличие антигеничности. Домены α 1 и α 2 складываются, чтобы составить канавку для связывания пептидов. Молекулы MHC класса I связывают пептиды, которые имеют преимущественно 8-10 аминокислот по длине (Parham 87), но также сообщалось о связывании более длинных пептидов. [ 6 ]
В то время как высокоаффинный пептид и субъединица B2M обычно требуются для поддержания стабильного тройного комплекса MHC с дефицитом пептидов . стабильные, гетеродимеры между пептидом, MHC I и B2M при субфизиологических температурах, наблюдались [ 7 ] [ 8 ] Синтетические стабильные, пептид-рецептивные MHC I-молекулы были получены с использованием дисульфидной связи между MHC I и B2M, названным «Open MHC-I». [ 9 ]
Синтез
[ редактировать ]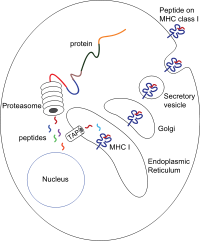
Пептиды генерируются в основном цитозоле протеасомой в . Протеасома - это макромолекула, которая состоит из 28 субъединиц, из которых половина влияет на протеолитическую активность. Протеасома разлагает внутриклеточные белки в мелкие пептиды, которые затем высвобождаются в цитозоль. Протеасомы также могут лигатировать различные пептидные фрагменты (называемые сплайсированными пептидами), продуцируя последовательности, которые являются нерегинных и, следовательно, не линейно шаблонами в геноме. Происхождение сплайсированных пептидных сегментов может происходить из одного и того же белка (цис-рассеяния) или разных белков (транс-рассеяние). [ 10 ] [ 11 ] Пептиды должны быть перемещены из цитозоля в эндоплазматический ретикулум (ER), чтобы встретить молекулу I класса MHC, чей пептид-связывающий сайт находится в просвете ER . У них есть мембрана проксимальная Ig складка
Транслокация и пептидная нагрузка
[ редактировать ]Пептидная транслокация из цитозоля в просвет ER достигается транспортером, связанным с обработкой антигена (TAP). TAP является членом семейства Transporter ABC и является гетеродимерным полипептидом мультимбранного промаза, состоящего из TAP1 и TAP2 . Две субъединицы образуют сайт связывания пептидов и два сайта связывания АТФ, которые сталкиваются с цитозолом. TAP связывает пептиды на цитоплазматической стороне и транслоцирует их при потреблении АТФ в просвет ER. Молекула MHC класса I затем, в свою очередь, загружается пептидами в просвете ER.
Процесс нагрузки пептидов включает в себя несколько других молекул, которые образуют большой мультимерный комплекс, называемый пептидным нагрузкой комплексом [ 12 ] состоящий из крана, тапасина , кальретикулина , кальнексина и ERP57 ( PDIA3 ). Кальнексин действует для стабилизации α -цепи MHC класса I до связывания β2M. После полной сборки молекулы MHC, кальнексин диссоциирует. Молекула MHC, в которой отсутствует связанный пептид, по своей природе нестабильна и требует связывания кальретикулина шаперонов и ERP57. Кроме того, тапасин связывается с молекулой MHC и служит для связи его с белками TAP и облегчает выбор пептида в итерационном процессе, называемом редактированием пептидов, [ 13 ] [ 14 ] [ 15 ] Таким образом, облегчая усиленную нагрузку на пептид и колокализацию.
Как только пептид загружается в молекулу MHC класса I, комплекс диссоциирует и оставляет ER через секреторный путь, чтобы достичь поверхности клетки. Транспорт молекул MHC класса I через секреторный путь включает в себя несколько посттрансляционных модификаций молекулы MHC. Некоторые из посттрансляционных модификаций происходят в ER и включают изменения в области N-гликана белка с последующими обширными изменениями в N-гликанах в аппарате Гольджи . N-гликаны полностью созревают, прежде чем они достигнут клеточной поверхности.
Пептид удаление
[ редактировать ]Пептиды, которые не связывают молекулы MHC класса I в просвете эндоплазматического ретикулума (ER), удаляются из ER через канал SEC61 в цитозоль, [ 16 ] [ 17 ] где они могут подвергаться дальнейшей обрезке в размерах и могут быть перемещены путем привязки к ER для связывания с молекулой класса I MHC.
взаимодействие SEC61 с бычьим альбумином . Например, было обнаружено [ 18 ]
Эффект вирусов
[ редактировать ]Молекулы MHC класса I загружаются пептидами, полученными в результате деградации убиквитинированных цитозольных белков в протеасомах . Поскольку вирусы индуцируют клеточную экспрессию вирусных белков, некоторые из этих продуктов помечены для деградации, причем полученные пептидные фрагменты попадают в эндоплазматический ретикулум и связывают с молекулами MHC I. Таким образом, путь MHC Class I-зависимый путь антигена представления, вирус, инфицированные клетками, сигнализируют о T-клетках, что аномальные белки продуцируются в результате инфекции.
Судьбой вирус-инфицированной клетки почти всегда индукция апоптоза посредством клеточного опосредованного иммунитета , снижая риск заражения соседних клеток. В качестве эволюционного ответа на этот метод иммунного наблюдения, многие вирусы способны подавлять или иным образом предотвращать представление молекул MHC класса I на клеточной поверхности. В отличие от цитотоксических Т -лимфоцитов, природные клетки -убийцы (NK) обычно инактивируются при распознавании молекул MHC I на поверхности клеток. Следовательно, в отсутствие молекул MHC I NK -клетки активируются и распознают клетку как аберрантную, что позволяет предположить, что она может быть инфицирована вирусами, пытающимися избежать иммунного разрушения. Несколько раковых заболеваний также демонстрируют пониженную регуляцию MHC I, что дает трансформированные клетки то же преимущество выживания, позволяющее избежать нормального иммунного наблюдения, предназначенного для уничтожения любых инфицированных или трансформированных клеток. [ 19 ]
Гены и изотипы
[ редактировать ]- Очень полиморфный
- Менее полиморфные
Эволюционная история
[ редактировать ]Гены MHC класса I возникли у самого последнего общего предка всех челюстных позвоночных и были обнаружены у всех живых челюстных позвоночных, которые до сих пор изучались. [ 2 ] С момента их появления в челюстных позвоночных, это семейство генов подвергалось многим дивергентным эволюционным путям, когда видообразования произошли события . Однако существуют задокументированные случаи транс-специфических полиморфизмов конкретный аллель гене MHC, связанном с эволюционным связанным в генах класса I MHC , где эволюционном в разновидность. [ 20 ] Эволюция рождения и смерти является одним из механистических объяснений размера семейства генов класса I MHC.
Рождение генов MHC класса I
[ редактировать ]Эволюция при рождении и смерти утверждает, что события дупликации генов приводят к тому, что геном содержит несколько копий гена, которые затем могут подвергаться отдельным эволюционным процессам. Иногда эти процессы приводят к псевдогенизации (смерти) одной копии гена, хотя иногда этот процесс приводит к двум новым генам с дивергентной функцией. [ 21 ] Вполне вероятно, что локусы IB класса MHC человека (HLA -E, -f и -g), а также Pseudogenes класса I MHC возникли из локусов MHC Class IA (HLA -A, -B и -C) в этом рождении - и процесс смерти. [ 22 ]
Ссылки
[ редактировать ]- ^ Hewitt EW (октябрь 2003 г.). «Путь презентации антигена класса I MHC: стратегии уклонения от вирусного иммунитета» . Иммунология . 110 (2): 163–9. doi : 10.1046/j.1365-2567.2003.01738.x . PMC 1783040 . PMID 14511229 .
- ^ Jump up to: а беременный Kulski JK, Shiina T, Anzai T, Kohara S, Inoko H (декабрь 2002 г.). «Сравнительный геномный анализ MHC: эволюция блоков дублирования класса I, разнообразие и сложности от акулы к человеку». Иммунологические обзоры . 190 : 95–122. doi : 10.1034/j.1600-065x.2002.19008.x . PMID 12493009 . S2CID 41765680 .
- ^ http://users.rcn.com/jkimball.ma.ultranet/biologypages/h/hla.html#class_i_histopamitibility_molecules Архивировано 2016-02-04 на . страницах биологии машины
- ^ Hansen TH, Bouvier M (июль 2009 г.). «Презентация антигена класса I MHC: обучение на стратегиях уклонения от вируса». Природные обзоры. Иммунология . 9 (7): 503–13. doi : 10.1038/nri2575 . PMID 19498380 . S2CID 9278263 .
- ^ Jump up to: а беременный в дюймовый Syken J, Grandpre T, Kanold Po, Shatz CJ (сентябрь 2006 г.). «PIRB ограничивает пластичность глазной доминантности в зрительной коре». Наука . 313 (5794): 1795–800. Bibcode : 2006sci ... 313.1795S . doi : 10.1126/science.1128232 . PMID 16917027 . S2CID 1860730 .
- ^ Барроуз С.Р., Россджон Дж., МакКласки Дж. (Январь 2006 г.). «Мы слишком короткие в картировании эпитопов CTL?». Тенденции в иммунологии . 27 (1): 11–6. doi : 10.1016/j.it.2005.11.001 . PMID 16297661 .
- ^ Lajurgregreen Hg, Stam, St. Ehm B, Witha, Bar, Bicyn J, B, Castin J, Pldinh H, Borth (1990-02). «Пустой MHC класс I Молклулы выходят на холоде» . Природа . 346 (6283): 476-480. Код BIB : 1990nstathe.346..476L . Doi : 10.1038 / 346476A0 . ISSN 0028-0836 . PMID 2198471 .
- ^ Schumacher TN, Heemels MT, Neefjes JJ, Kast W, Melief CJ, Ploegh HL (август 1990 г.). «Прямое связывание пептида с пустыми молекулами MHC класса I на интактных клетках и in vitro» . Клетка . 62 (3): 563–567. doi : 10.1016/0092-8674 (90) 90020-f . PMID 2199065 .
- ^ Sun Y, Young MC, Woodward CH, Danon JN, Truong HV, Gupta S, Winters TJ, Font-Burgada J, Burslem GM, Sgourakis NG (2023-06-20). «Универсальные открытые молекулы MHC-I для быстрой пептидной нагрузки и повышенной сложной стабильности между аллотипами HLA» . Труды Национальной академии наук . 120 (25): E23040555120. Bibcode : 2023pnas..12004055S . doi : 10.1073/pnas.2304055120 . ISSN 0027-8424 . PMC 10288639 . PMID 37310998 .
- ^ Faridi P, Li C, Ramarathinam SH, Vivian JP, Illing PT, Mifsud NA, Ayala R, Song J, Gearing LJ, Hertzog PJ, Ternette N, Rossjohn J, Croft NP, Purcell AW (12 октября 2018 г.). «Подмножество пептидов HLA-I не является геномически шаблонной: доказательствами цис-сплайдных пептидных лигандов» (PDF) . Научная иммунология . 3 (28): EAAR3947. doi : 10.1126/sciimmunol.aar3947 . PMID 30315122 .
- ^ Liepe J, Marino F, Sidney J, Jeko A, Bunting DE, Sette A, Kloetzel PM, Stumpf MP, Heck AJ, Mishto M (21 октября 2016 года). «Большая часть лигандов HLA класса I представляет собой сплайсированные пептиды, созданные протеасомой» (PDF) . Наука . 354 (6310): 354–358. Bibcode : 2016sci ... 354..354L . doi : 10.1126/science.aaf4384 . HDL : 10044/1/42330 . PMID 27846572 . S2CID 41095551 .
- ^ Blees A, Januliene D, Hofmann T, Koller N, Schmidt C, Trowitzsch S, Moeller A, Tampé R (ноябрь 2017). «Структура комплекса пептид-нагрузки человека MHC-I». Природа . 551 (7681): 525–528. Bibcode : 2017natur.551..525b . doi : 10.1038/nature24627 . PMID 29107940 . S2CID 4447406 .
- ^ Ховарт М., Уильямс А., Толстрап А.Б., Эллиотт Т (август 2004 г.). «Тапасин усиливает презентацию пептидов MHC класса I в соответствии с пептидом полураспада» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (32): 11737–42. Bibcode : 2004pnas..10111737H . doi : 10.1073/pnas.0306294101 . PMC 511045 . PMID 15286279 .
- ^ Wearsch PA, Cresswell P (август 2007 г.). «Селективная нагрузка высокоаффинных пептидов на основные молекулы комплекса класса I класса I с помощью гетеродимера Tapasin-ERP57». Природа иммунология . 8 (8): 873–81. doi : 10.1038/ni1485 . PMID 17603487 . S2CID 29762957 .
- ^ Третийборо С.М., Роддик Дж.С., Рэдклифф Дж .н., Ховарт М., Стивенсон Ф.К., Эллиотт Т (февраль 2008 г.). «Тапасин формирует иерархии иммунодоминантности в соответствии с кинетической стабильностью комплексов пептид-MHC класса I» . Европейский журнал иммунологии . 38 (2): 364–9. doi : 10.1002/eji.200737832 . PMID 18196518 . S2CID 28659293 .
- ^ Koopmann Jo, Albring J, Hüter E, Bulbuc N, Spee P, Neefjes J, Hämmerling GJ, Momburg F, et al. (Июль 2000 г.). «Экспорт антигенных пептидов из эндоплазматического ретикулума пересекается с ретроградной транслокацией белка через канал SEC61P» . Иммунитет . 13 (1): 117–27. doi : 10.1016/s1074-7613 (00) 00013-3 . PMID 10933400 .
- ^ Albring J, Koopmann Jo, Hämmerling GJ, Momburg F (январь 2004 г.). «Ретротранслокация тяжелой цепи MHC класса I от эндоплазматической ретикулумы к цитозолю зависит от снабжения АТФ от просвета ER». Молекулярная иммунология . 40 (10): 733–41. doi : 10.1016/j.molimm.2003.08.008 . PMID 14644099 .
- ^ Имай Дж, Хасегава Х., Маруя М., Коясу С., Яхара I (январь 2005 г.). «Экзогенные антигены обрабатываются с помощью эндоплазматической ретикулум, связанной с деградацией (ERAD) при перекрестной презентации дендритными клетками» . Международная иммунология . 17 (1): 45–53. doi : 10.1093/intimm/dxh184 . PMID 15546887 .
- ^ Wang Z, Zhang L, Qiao A, Watson K, Zhang J, Fan GH (февраль 2008 г.). «Активация CXCR4 запускает убиквитинирование и подавление основной гистосовместимости комплекса класса I (MHC-I) на эпителиоидных клетках Hela карциномы» . Журнал биологической химии . 283 (7): 3951–9. doi : 10.1074/jbc.m706848200 . PMID 18083706 .
- ^ Azevedo L, Serrano C, Amorim A, Cooper DN (сентябрь 2015 г.). «Трансвидовый полиморфизм у людей и великих обезьян, как правило, поддерживается путем балансировки отбора, который модулирует иммунный ответ хозяина» . Человеческая геномика . 9 (1): 21. doi : 10.1186/s40246-015-0043-1 . PMC 4559023 . PMID 26337052 .
- ^ Ней М., Руни А.П. (2005-11-14). «Согласованная и эволюция с рождением и смертью многоэблеточных семей» . Ежегодный обзор генетики . 39 (1): 121–52. doi : 10.1146/annurev.genet.39.073003.112240 . PMC 1464479 . PMID 16285855 .
- ^ Хьюз А.Л. (март 1995 г.). «Происхождение и эволюция HLA Pseudogenes класса I» . Молекулярная биология и эволюция . 12 (2): 247–58. doi : 10.1093/oxfordjournals.molbev.a040201 . PMID 7700152 .
Внешние ссылки
[ редактировать ]- Гистосовместимость+антигены+класс+I библиотеки США в Национальной библиотеке Медицинской .
- Гены MHC+Class+I+ в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)