фолликулогенез
- Хотя у многих животных этот процесс аналогичен, в этой статье речь пойдет исключительно о фолликулогенезе человека .

1 - Менструация
2 - Развивающийся фолликул
3 – Зрелый фолликул
4 - Овуляция
5 — желтое тело
6 - Разрушение желтого тела
В биологии фолликулогенез – это созревание фолликула яичника , плотно упакованной оболочки соматических клеток , содержащей незрелый ооцит . Фолликулогенез описывает прогрессирование ряда мелких примордиальных фолликулов в крупные преовуляторные фолликулы , что частично происходит во время менструального цикла .
В отличие от мужского сперматогенеза , который может длиться бесконечно, фолликулогенез заканчивается, когда оставшиеся фолликулы в яичниках неспособны реагировать на гормональные сигналы, которые ранее привлекали некоторые фолликулы к созреванию. Это истощение запасов фолликулов сигнализирует о начале менопаузы .
Обзор
[ редактировать ]Основная роль фолликула – поддержка яйцеклетки . Из всего пула фолликулов, с которыми рождается женщина, только у 0,1% из них произойдет овуляция, тогда как 99,9% разрушатся (в процессе, называемом фолликулярной атрезией ). С самого рождения яичники женщины содержат множество незрелых, примордиальных фолликулов . Каждый из этих фолликулов содержит одинаково незрелый первичный ооцит . В период полового созревания складки фолликулов начинают фолликулогенез, вступая в модель роста, которая заканчивается овуляцией (процессом, при котором яйцеклетка покидает фолликул) или атрезией (гибелью гранулезных клеток фолликула). [ нужна ссылка ]
Во время развития фолликула примордиальные фолликулы претерпевают ряд критических изменений, как гистологических, так и гормональных. Сначала они превращаются в первичные фолликулы, а затем во вторичные фолликулы. Затем фолликулы переходят в третичные или антральные фолликулы . На этом этапе развития они становятся зависимыми от гормонов, особенно от ФСГ, что приводит к существенному увеличению скорости их роста. Поздний третичный или преовуляторный фолликул разрывается и высвобождает яйцеклетку (которая становится вторичным ооцитом ), заканчивая фолликулогенез.
«Отбор» фолликула — это процесс, посредством которого из выбранной когорты или волны выбирается один «доминирующий» фолликул для преимущественного роста. Обычно документально подтверждено, что это происходит один раз в ранней и средней фолликулярной фазе менструального цикла, что приводит к овуляции. [1]
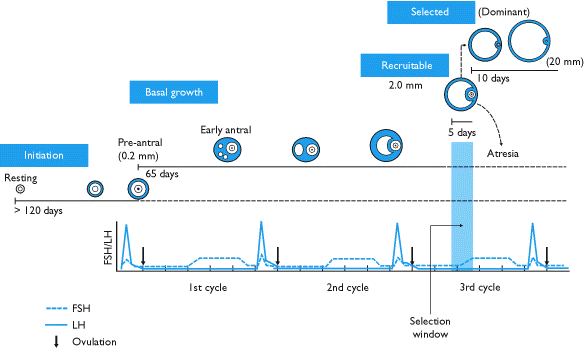
Фазы развития
[ редактировать ]Фолликулогенез непрерывен, а это означает, что в любой момент яичник содержит фолликулы на разных стадиях развития. Большинство фолликулов погибают и никогда не завершают развитие. Некоторые из них полностью развиваются и производят вторичный ооцит, который высвобождается при разрыве фолликула в процессе, называемом овуляцией .
Растущий фолликул проходит следующие отдельные стадии, которые определяются определенными структурными характеристиками:
стадии отидогенеза оогенеза В более широкой перспективе весь фолликулогенез, от примордиального до преовуляторного фолликула, относится к .
| Этап | Описание | Размер |
| Первозданный | Спящие , мелкие, только один слой плоских гранулезных клеток. | Примордиальные фолликулы имеют диаметр около 0,03–0,05 мм. |
| Начальный | Митотические клетки, кубовидные гранулезные клетки | Почти 0,1 мм в диаметре |
| вторичный | Наличие тека-клеток , множественных слоев гранулезных клеток. | Фолликул теперь имеет диаметр 0,2 мм. |
| Раннее высшее образование | Ранний третичный фолликул условно делят на пять классов. Фолликулы класса 1 имеют диаметр 0,2 мм, класса 2 — около 0,4 мм, класса 3 — около 0,9 мм, класса 4 — около 2 мм и класса 5 — около 5 мм. | |
| Позднее высшее образование | Полностью сформированный антральный отдел , отсутствие дальнейшей цитодифференцировки, отсутствие нового прогресса. | Фолликулы 6-го класса имеют диаметр около 10 мм, 7-го класса — около 16 мм и 8-го класса — около 20 мм. Недоминантные фолликулы часто вырастают за пределы 5-го класса, но редко бывает более одного фолликула 8-го класса. |
| Преовуляторный | Наращивание роста концентрации эстрогена, все остальные фолликулы атретические или мертвые. |
Кроме того, фолликулы, образовавшие антральный отдел, называются антральными фолликулами или граафовыми фолликулами. Определения различаются в зависимости от того, где происходит этот сдвиг на приведенной выше стадии: некоторые утверждают, что он происходит при переходе на вторичную стадию. [2] и другие утверждают, что это происходит при переходе на третичную стадию. [3]
До преовуляторной стадии фолликул содержит первичный ооцит, который задерживается в профазе I. мейоза На поздней преовуляторной стадии ооцит продолжает мейоз и становится вторичным ооцитом, остановленным в метафазе II .

Первозданный
[ редактировать ]Через 18–22 недели после зачатия кора женского яичника (яичник плода) содержит максимальное количество фолликулов (в среднем от 4 до 5 миллионов, но отдельные пиковые популяции колеблются от 6 до 7 миллионов). [4] Эти примордиальные фолликулы содержат незрелые ооциты, окруженные плоскими чешуйчатыми гранулезными клетками (опорными клетками), которые отделены от окружающей среды ооцита базальной пластинкой. Они находятся в состоянии покоя и практически не проявляют биологической активности . Поскольку примордиальные фолликулы у человека могут находиться в состоянии покоя до 50 лет, продолжительность овариального цикла не включает это время.
Количество фолликулов немного уменьшается перед рождением и в среднем составляет 500 000 к моменту полового созревания (популяция в период полового созревания колеблется от 25 000 до 1,5 миллионов). [4] Из-за «неэффективного» характера фолликулогенеза (обсуждаемого позже) только 400–500 из этих фолликулов когда-либо достигнут преовуляторной стадии. В период менопаузы остается всего 1000 фолликулов. Кажется вероятным, что ранняя менопауза возникает у женщин с низкой численностью населения при рождении, а поздняя менопауза возникает у женщин с высокой численностью населения при рождении, но клинических доказательств этого пока нет. [4]
Процесс «пробуждения» первичных клеток известен как начальное пополнение. Исследования показали, что первоначальное рекрутирование опосредовано противовесом различных стимулирующих и ингибирующих гормонов и локально продуцируемых факторов роста. [5]
Начальный
[ редактировать ]Во время активации фолликула яичника гранулезные клетки примордиальных фолликулов изменяются с плоской на кубовидную структуру, отмечая начало первичного фолликула . Геном ооцита активируется, и гены начинают транскрибироваться . рудиментарные паракринные сигнальные Формируются пути, жизненно важные для связи между фолликулом и ооцитом. И ооцит, и фолликул резко растут, увеличиваясь почти до 0,1 мм в диаметре. [ нужна ссылка ]
В это время в первичных фолликулах развиваются рецепторы к фолликулостимулирующему гормону (ФСГ), но они не зависят от гонадотропинов до антральной стадии. Однако исследования показали, что присутствие ФСГ ускоряет рост фолликулов in vitro .
Вокруг ооцита образуется гликопротеиновая полимерная капсула, называемая пеллюцидной зоной , отделяющая ее от окружающих гранулезных клеток. Пеллюцидная зона, которая остается с ооцитом после овуляции, содержит ферменты, которые катализируют сперму, обеспечивая проникновение.
вторичный
[ редактировать ]Стромоподобные тека-клетки рекрутируются с помощью сигналов, секретируемых ооцитами. Они окружают самый внешний слой фолликула, базальную пластинку , и подвергаются цитодифференцировке, образуя наружную и внутреннюю теку . Между этими двумя слоями оболочки образуется сложная сеть капиллярных сосудов, которая начинает циркулировать кровь к фолликулу и от него.
Поздний вторичный фолликул гистологически и структурно отмечен полностью выросшим ооцитом, окруженным прозрачной оболочкой, примерно девятью слоями гранулезных клеток, базальной пластинкой, внутренней текой, капиллярной сетью и внешней текой. Развитие антрального отдела также начинается на стадии вторичного фолликула.
Формирование антрума
[ редактировать ]Образование заполненной жидкостью полости, прилегающей к ооциту, называемой антрумом, обозначает фолликул как антральный фолликул , в отличие от так называемого преантрального фолликула , у которого еще нет антрального отверстия. Антральный фолликул также называют граафовым фолликулом .
Определения различаются относительно того, на какой стадии происходит этот сдвиг: некоторые называют фолликулы на вторичной стадии антральными . [2] и другие, обозначающие их как преантральные . [3]
Раннее высшее образование
[ редактировать ]В третичном фолликуле сформирована основная структура зрелого фолликула, и новые клетки не обнаруживаются. Клетки гранулезы и теки продолжают подвергаться митозу одновременно с увеличением объема антрального отдела. Третичные фолликулы могут достигать огромных размеров, чему препятствует только наличие ФСГ, от которого они теперь зависят.
Под действием морфогенного градиента, секретируемого ооцитами, гранулезные клетки третичного фолликула подвергаются дифференцировке на четыре различных подтипа: corona radiata , окружающая пеллюцидную зону; перепонка , внутри базальной пластинки; периантральная , примыкающая к антральному отделу и кучевому отверстию офора , соединяющая перепонку и гранулезные клетки лучистого венца между собой. Каждый тип клеток ведет себя по-разному в ответ на ФСГ.
Клетки Theca interna экспрессируют рецепторы лютеинизирующего гормона ( ЛГ ). ЛГ индуцирует выработку андрогенов тека-клетками, особенно андростендиона , который ароматизируется гранулезными клетками с образованием эстрогенов , в первую очередь эстрадиола . Следовательно, уровень эстрогена начинает повышаться.
Поздняя третичная и преовуляторная (фолликулярная фаза менструального цикла)
[ редактировать ]К этому моменту большая часть группы фолликулов, начавших рост, погибла. Этот процесс гибели фолликула известен как атрезия и характеризуется радикальным апоптозом всех составляющих клеток и ооцита. Хотя неизвестно, что вызывает атрезию, было показано, что присутствие высоких концентраций ФСГ предотвращает ее.
Повышение уровня ФСГ в гипофизе, вызванное распадом желтого тела в конце менструального цикла, ускоряет рекрутирование пяти-семи фолликулов класса 5 для участия в следующем цикле. Эти фолликулы входят в конец предыдущего менструального цикла и переходят в фолликулярную фазу следующего. Выбранные фолликулы, называемые антральными фолликулами, конкурируют друг с другом за стимулирующий рост ФСГ.
Характер появления группы из пяти-семи антральных фолликулов обсуждается. Существуют теории непрерывного рекрутирования антральных фолликулов, теории единичного эпизода рекрутирования в конце лютеиновой фазы, а совсем недавно появились доказательства модели рекрутирования, характеризующейся 2–3 волнами рекрутирования и развития фолликулов в течение менструального цикла. (только одна из которых на самом деле является овуляторной волной). [6]
В ответ на повышение ФСГ антральные фолликулы начинают секретировать эстроген и ингибин , которые оказывают на ФСГ отрицательный эффект обратной связи. [7] Фолликулы, у которых меньше ФСГ-рецепторов , не смогут развиваться дальше; они покажут замедление темпов роста и станут атретическими. В конечном итоге жизнеспособным будет только один фолликул. Этот оставшийся фолликул, называемый доминантным фолликулом , будет быстро и резко расти — до 20 мм в диаметре — и станет преовуляторным фолликулом .
Примечание. Многие источники искажают темпы роста фолликула, а некоторые даже предполагают, что для того, чтобы примордиальный фолликул стал преовуляторным, требуется всего четырнадцать дней. Собственно, фолликулярная фаза менструального цикла означает время между выделением третичного фолликула и последующим его ростом в преовуляторный фолликул. Фактическое время развития фолликула варьируется.
Рост доминантного фолликула во время фолликулярной фазы составляет около 1,5 мм в сутки (±0,1 мм) как в естественных циклах, так и для любого доминантного фолликула, развивающегося на фоне приема комбинированных пероральных контрацептивов . [8] Выполнение контролируемой гиперстимуляции яичников приводит к большему набору фолликулов, их рост составляет около 1,6 мм в день. [8]
Овуляция и желтое тело
[ редактировать ]К концу фолликулярной (или пролиферативной) фазы тринадцатого дня менструального цикла в кумулюсном слое предовуляторного фолликула образуется отверстие, или стигма , и выделяется ооцит с набором кумулюсных клеток в процессе, называемом овуляция . В естественных циклах овуляция может произойти в фолликулах размером не менее 14 мм. [9]
Технически ооцит все еще является вторичным ооцитом, находящимся в метафазе II мейоза. Только после оплодотворения он разовьется в яйцеклетку, а затем быстро в яйцеклетку (посредством завершения мейоза II). Теперь яйцеклетка будет перемещаться по одной из фаллопиевых труб и в конечном итоге выйти из нее во время менструации в случае, если она окажется неоплодотворенной или если она не будет успешно имплантирована в матку (если она была ранее оплодотворена ).
Уровень эстрадиола увеличивается и вызывает овуляторный пик ЛГ (и ФСГ). Этот пик (через AMPc) активирует провоспалительные гены, которые вызывают разрыв стенки фолликула и выход яйцеклетки наружу. Разорвавшийся фолликул претерпевает драматическую трансформацию в желтое тело , стероидиогенный кластер клеток, который поддерживает эндометрий матки путем секреции большого количества прогестерона и незначительного количества эстрогена .
Эти два этапа, хотя и не являются частью фолликулогенеза, включены для полноты картины. Они полностью обсуждаются в соответствующих статьях и рассматриваются в статье о менструальном цикле . Рекомендуется рассмотреть эти три темы.
Гормональная функция
[ редактировать ]Как и большинство вещей, связанных с репродуктивной системой, фолликулогенез контролируется эндокринной системой . Пять гормонов участвуют в сложном процессе положительной и отрицательной обратной связи, регулирующем фолликулогенез. Они есть:
- гонадотропин-рилизинг-гормон (ГнРГ), секретируемый гипоталамусом
- два гонадотропина :
- эстроген
- прогестерон
ГнРГ стимулирует высвобождение ФСГ и ЛГ из передней доли гипофиза, что впоследствии оказывает стимулирующее воздействие на рост фолликулов (однако не сразу, поскольку только антральные фолликулы зависят от ФСГ и ЛГ). Когда тека-клетки образуются в третичном фолликуле, количество эстрогена резко увеличивается (андроген, происходящий из теки, ароматизируется в эстроген гранулезными клетками).
В низкой концентрации эстроген ингибирует гонадотропины, но высокая концентрация эстрогена стимулирует их. Кроме того, по мере секреции большего количества эстрогена тека-клетки производят больше рецепторов ЛГ, побуждая тека-клетки вырабатывать больше андрогена, который впоследствии становится эстрогеном. Эта петля положительной обратной связи вызывает резкий скачок ЛГ, и именно этот скачок вызывает овуляцию.
После овуляции ЛГ стимулирует образование желтого тела. После овуляции уровень эстрогена упал до отрицательного стимулирующего уровня и, следовательно, служит для поддержания концентрации ФСГ и ЛГ. Ингибин, который также секретируется желтым телом, способствует ингибированию ФСГ. Прогестерон, секретируемый желтым телом, подавляет рост фолликулов и поддерживает беременность.
Эндокринная система совпадает с менструальным циклом и проходит тринадцать циклов (и, следовательно, тринадцать всплесков ЛГ) в ходе нормального фолликулогенеза. Однако скоординированная передача сигналов ферментов и специфичная во времени экспрессия гормональных рецепторов гарантируют, что рост фолликулов не будет нарушен во время этих преждевременных всплесков.
Количество фолликулов
[ редактировать ]
Недавно две публикации подвергли сомнению идею о том, что во время рождения закладывается конечное число фолликулов. [11] [12] Сообщалось об обновлении фолликулов яичников из стволовых клеток зародышевой линии (происходящих из костного мозга и периферической крови) в постнатальных яичниках мышей. Исследования, пытающиеся воспроизвести эти результаты, продолжаются, но исследование популяций в 325 яичниках человека не выявило подтверждающих доказательств пополнения фолликулов. [4]
В 2010 году исследователи из Эдинбургского университета определили, что к 30 годам у женщин остается только 10% нерастущих фолликулов (NGF). [10] При рождении у женщины все фолликулы готовы к фолликулогенезу, и их количество неуклонно уменьшается до наступления менопаузы .
Истощение овариального резерва
[ редактировать ]С возрастом у женщин (и мышей) в резерве примордиальных фолликулов накапливаются двухцепочечные разрывы. Эти фолликулы содержат первичные ооциты, которые задерживаются в профазе первого клеточного деления мейоза. Двухцепочечные разрывы аккуратно восстанавливаются во время мейоза путем поиска и построения соответствующей цепи (так называемая «гомологичная рекомбинационная репарация»). Титус и др. [13] (2013) обнаружили, что с возрастом у людей (и мышей) экспрессия четырех ключевых генов репарации ДНК, необходимых для гомологичной рекомбинационной репарации, в ооцитах снижается. Они предположили, что репарация двухцепочечных разрывов ДНК жизненно важна для поддержания резерва ооцитов и что снижение эффективности репарации с возрастом играет ключевую роль в истощении резерва яичников (старении яичников).
См. также
[ редактировать ]Дополнительные изображения
[ редактировать ] Раздел яичника. (№5–9 представляют стадии фолликулогенеза)
Раздел яичника. (№5–9 представляют стадии фолликулогенеза) переходный первичный фолликул.
переходный первичный фолликул.


Ссылки
[ редактировать ]- ^ Бервальд, Анджела Р.; Адамс, Грегг П.; Пирсон, Роджер А. (2012). «Антральный фолликулогенез яичников во время менструального цикла человека: обзор» . Обновление репродукции человека . 18 (1): 73–91. дои : 10.1093/humupd/dmr039 . ISSN 1460-2369 . ПМИД 22068695 .
- ^ Jump up to: а б Страница 769 , раздел «Формирование антрального отдела» в: Шервуд, Лорали. (2010). Физиология человека: от клеток к системе . Австралия; США: Брукс/Коул. ISBN 978-0-495-39184-5 .
- ^ Jump up to: а б Страница 76 в: Ванденхурк, Р.; Беверс, М.; Беккерс, Дж. (1997). «Развитие преантральных фолликулов in vivo и in vitro». Териогенология . 47 : 73–82. дои : 10.1016/S0093-691X(96)00341-X .
- ^ Jump up to: а б с д Уоллес, WHB; Келси, ТВ (2010). «Овариальный резерв человека от зачатия до менопаузы» . ПЛОС ОДИН . 5 (1): е8772. arXiv : 1106.1382 . Бибкод : 2010PLoSO...5.8772W . дои : 10.1371/journal.pone.0008772 . ПМК 2811725 . ПМИД 20111701 .
- ^ Форчун Дж., Кушман Р., Валь С., Кито С. (2000). «Переход из первичного фолликула в первичный» . Мол клеточный эндокринол . 163 (1–2): 53–60. дои : 10.1016/S0303-7207(99)00240-3 . ПМИД 10963874 . S2CID 8746207 .
- ^ «Антральный фолликулогенез яичников во время менструального цикла человека: обзор» . Исследовательские ворота . Проверено 23 января 2019 г.
- ^ де Зиглер Д. (2007), «Роль ФСГ и ЛГ во время фолликулярной фазы: понимание ЭКО в естественном цикле», RBM Online, том 15, № 5, стр. 508
- ^ Jump up to: а б Бервальд, Анджела Р.; Уокер, Рэнди А.; Пирсон, Роджер А. (2009). «Темпы роста фолликулов яичников во время естественных менструальных циклов, циклов оральной контрацепции и циклов стимуляции яичников» . Фертильность и бесплодие . 91 (2): 440–449. doi : 10.1016/j.fertnstert.2007.11.054 . ISSN 0015-0282 . ПМИД 18249401 .
- ^ Страница 34 в: Майкл К. Скиннер (2018). Энциклопедия репродукции (2-е изд.). Академическая пресса. ISBN 9780128151457 .
- ^ Jump up to: а б Уоллес, В. Хэмиш Б.; Томас В. Келси (27 января 2010 г.). «Овариальный резерв человека от зачатия до менопаузы» . ПЛОС ОДИН . 5 (1): е8772. arXiv : 1106.1382 . Бибкод : 2010PLoSO...5.8772W . дои : 10.1371/journal.pone.0008772 . ПМК 2811725 . ПМИД 20111701 .
- ^ Джонсон Дж., Бэгли Дж., Сказник-Викиэль М., Ли Х., Адамс Г., Ниикура Ю., Чуди К., Тилли Дж., Кортес М., Форкерт Р., Спитцер Т., Якомини Дж., Скадден Д., Тилли Дж. (2005). «Генерация ооцитов в яичниках взрослых млекопитающих предполагаемыми половыми клетками в костном мозге и периферической крови» . Клетка . 122 (2): 303–15. дои : 10.1016/j.cell.2005.06.031 . ПМИД 16051153 . S2CID 19006732 .
- ^ Джонсон Дж., Каннинг Дж., Канеко Т., Пру Дж., Тилли Дж. (2004). «Зародышевые стволовые клетки и обновление фолликулов в постнатальных яичниках млекопитающих». Природа . 428 (6979): 145–50. Бибкод : 2004Natur.428..145J . дои : 10.1038/nature02316 . ПМИД 15014492 . S2CID 1124530 .
- ^ Титус, С; Ли, Ф; Стобезки, Р; Акула, К; Унсал, Э; Чон, К; Диклер, М; Робсон, М; Мой, Ф; Госвами, С; Октай, К (2013). «Нарушение репарации двухцепочечных разрывов ДНК, связанных с BRCA1, приводит к старению яичников у мышей и людей» . Научный перевод Мед . 5 (172): 172. doi : 10.1126/scitranslmed.3004925 . ПМК 5130338 . ПМИД 23408054 .
- Чаглар Г., Асимакопулос Б., Николеттос Н., Дидрих К., Аль-Хасани С. (2005). «Рекомбинантный ЛГ при стимуляции яичников» . Репрод Биомед Онлайн . 10 (6): 774–85. дои : 10.1016/S1472-6483(10)61123-6 . ПМИД 15970010 .
- Форчун, Дж. Э.; Ян, МОЙ; Муруви, В. (2010). Люси, MC; Пейт, Дж.Л.; Смит, МФ; Спенсер, Т.Э. (ред.). «Самые ранние стадии развития фолликула: образование и активация фолликула» . Soc Reprod Fertil Suppl . 67 : 203–16. doi : 10.7313/upo9781907284991.018 (неактивен 31 марта 2024 г.). ISBN 9781907284991 . ПМК 5316466 . ПМИД 21755674 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на март 2024 г. ( ссылка ) - Гужон А (1996). «Регуляция развития фолликулов яичников у приматов: факты и гипотезы» . Эндокр преп . 17 (2): 121–55. дои : 10.1210/edrv-17-2-121 . ПМИД 8706629 .
- Гужон А (1986). «Динамика роста фолликулов у человека: модель на основе предварительных результатов». Хум Репрод . 1 (2): 81–7. doi : 10.1093/oxfordjournals.humrep.a136365 . ПМИД 3558758 .
- ван ден Хурк Р., Чжао Дж (2005). «Формирование ооцитов млекопитающих, их рост, дифференциация и созревание в фолликулах яичников» . Териогенология . 63 (6): 1717–51. doi : 10.1016/j.theriogenology.2004.08.005 . ПМИД 15763114 .
- Узюмджу, Мехмет; Зачоу, Роб (2007). «Воздействие на развитие эндокринных разрушителей окружающей среды: последствия для яичников и женской репродуктивной функции» . Репродуктивная токсикология . 23 (3): 337–352. Бибкод : 2007RepTx..23..337U . дои : 10.1016/j.reprotox.2006.10.006 . ПМЦ 1950429 . ПМИД 17140764 .