Нитчатый бактериофаг
| Иновириды | |
|---|---|

| |
| Электронная микрофотография затененного нитчатого бактериофага (иновируса) | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Моноднавирия |
| Королевство: | Лебвира |
| Тип: | Хофнейвирикота |
| Сорт: | Фасервирицетес |
| Заказ: | Тубулавирусы |
| Семья: | Иновириды |
| Роды | |
Нитчатые бактериофаги — семейство вирусов ( Inoviridae ), поражающих бактерии , или бактериофаги . Они названы в честь своей нитевидной формы, червеобразной цепочки (длинной, тонкой и гибкой, напоминающей длину приготовленных спагетти), диаметром около 6 нм и длиной около 1000-2000 нм. [ 1 ] [ 2 ] [ 3 ] [ 4 ] [ 5 ] Эта отличительная форма отражает их метод репликации : оболочка вириона состоит из пяти типов вирусных белков, которые располагаются во внутренней мембране бактерии-хозяина во время сборки фага, и эти белки добавляются к ДНК зарождающегося вириона по мере его выдавливания. через мембрану. Простота нитчатых фагов делает их привлекательными модельными организмами для исследований в области молекулярной биологии , а также они показали себя многообещающими в качестве инструментов в нанотехнологиях и иммунологии .
Характеристики
[ редактировать ]
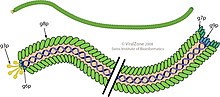
Нитчатые бактериофаги являются одними из самых простых известных живых организмов, у них гораздо меньше генов, чем у классических хвостатых бактериофагов, изученных группой фагов в середине 20 века. Семейство включает 29 определенных видов, разделенных на 23 рода. [ 6 ] [ 7 ] Однако анализ наборов геномных и метагеномных данных с использованием подхода машинного обучения привел к открытию 10 295 иновирусоподобных последовательностей почти во всех бактериальных типах практически во всех экосистемах, что указывает на то, что эта группа вирусов гораздо более разнообразна и широко распространена, чем первоначально предполагалось. [ 5 ]
Три нитчатых бактериофага fd, f1 и M13 были выделены и охарактеризованы тремя разными исследовательскими группами в начале 1960-х годов, но они настолько похожи, что их иногда объединяют под общим названием «Ff», которые являются представителями рода Inovirus. , как признано Международным комитетом по таксономии вирусов (ICTV). [ 8 ] [ 9 ] Молекулярная структура фагов Ff была определена с использованием ряда физических методов, в частности дифракции рентгеновских лучей . [ 2 ] [ 6 ] твердотельный ЯМР и криоэлектронная микроскопия . [ 10 ] Структуры фагового капсида и некоторых других фаговых белков доступны в Банке данных белков. [ 6 ] Одноцепочечная ДНК фага Ff проходит по центральному ядру фага и защищена цилиндрической белковой оболочкой, построенной из тысяч идентичных субъединиц α-спирального основного белка оболочки, кодируемых фаговым геном 8. Белок гена 8 вставляется в Плазматическая мембрана как ранний этап сборки фага. [ 2 ] Некоторые штаммы фага имеют «лидерную последовательность» в белке гена 8, способствующую встраиванию в мембрану, но другим, похоже, лидерная последовательность не нужна. Два конца фага покрыты несколькими копиями белков, которые важны для заражения бактерий-хозяев, а также для сборки зарождающихся фаговых частиц. Эти белки являются продуктами фаговых генов 3 и 6 на одном конце фага и фаговых генов 7 и 9 на другом конце. Исследования дифракции волокон выявили два структурных класса фагов, различающихся деталями расположения белка гена 8. Класс I имеет ось вращения, связанную с белками оболочки гена 8, тогда как для класса II эта ось вращения заменена осью спирали. Это техническое различие мало заметно влияет на общую структуру фага, но объем независимых дифракционных данных больше для класса симметрии II, чем для класса I. Это помогло определить структуру Pf1 фага класса II и, как следствие, структуру класса I. . [ 2 ] [ 6 ]
Структурный класс I включает штаммы fd, f1, M13 рода Inovirus , а также If1 (вида ICTV вируса Escherichia If1 , рода Infulavirus ). [ 11 ] и IKe (из вида ICTV вируса сальмонеллы IKe , рода Lineavirus ), [ 12 ] тогда как класс II включает штаммы Pf1 (вида ICTV вируса Pseudomonas Pf1 рода Primolicivirus ), [ 13 ] вида ICTV и, возможно, также Pf3 ( вирус Pseudomonas Pf3 рода Tertilicivirus ), [ 14 ] Пф4 [ 15 ] и PH75 (предложенный NCBI вид фага Thermus PH75 , неопределенного размещения в Inoviridae ). [ 16 ]
ДНК, выделенная из фага fd (рода Inovirus ), является одноцепочечной и топологически представляет собой круг. То есть одна цепь ДНК простирается от одного конца фаговой частицы к другому, а затем снова обратно, замыкая круг, хотя две цепи не спарены основаниями. Предполагалось, что эта топология распространяется на все другие нитчатые фаги, но это не относится к фагу Pf4. [ 15 ] для которого ДНК в фаге одноцепочечная, но топологически линейная, а не кольцевая. [ 10 ] Во время сборки фага fd ДНК фага сначала упаковывается в линейный внутриклеточный нуклеопротеиновый комплекс со многими копиями белка репликации/сборки фагового гена 5. Затем белок гена 5 заменяется оболочочным белком гена 8, когда возникающий фаг экструдируется через бактериальную плазматическую мембрану, не убивая бактериального хозяина. [ 17 ] [ 18 ] [ 2 ] [ 19 ] Этот белок также с высоким сродством связывается со структурами G-квадруплекса (хотя они не присутствуют в фаговой ДНК) и с аналогичными шпилечными структурами в фаговой ДНК. [ 20 ]
Белок p1 фага Ff (т.е. рода Inovirus ), который необходим для сборки фага на мембране, имеет трансмембранный гидрофобный домен с N-концевой частью в цитоплазме и С-концевой частью в периплазме (обратный ориентации белка оболочки гена 8). К цитоплазматической стороне трансмембранного домена примыкает последовательность из 13 остатков р1, имеющая структуру основных остатков, близко соответствующую структуре основных остатков вблизи С-конца р8, но инвертированную по отношению к последовательности. Этот механизм сборки делает этот фаг ценной системой для изучения трансмембранных белков . [ 2 ] [ 21 ] [ 4 ] Ген 1, кодирующий АТФазу, [ 22 ] представляет собой консервативный маркерный ген, который (наряду с тремя дополнительными генетическими признаками) использовался для автоматического обнаружения последовательностей иновируса. [ 5 ]
Жизненный цикл
[ редактировать ]Репликация вируса цитоплазматическая. Проникновение в клетку-хозяина достигается путем опосредованной пилусами адсорбции в клетке-хозяине. Репликация следует модели вращающегося круга оцДНК. Транскрипция с помощью ДНК-матрицы - это метод транскрипции. Вирус покидает клетку-хозяина путем вирусной экструзии. [ 23 ] Сборка вируса происходит на внутренней мембране (в случае грамотрицательных бактерий) при помощи встроенного в мембрану комплекса моторных белков. [ 23 ] Этот мультимерный комплекс сборки, включающий p1, кодируемый геном 1 (называемый исследователями фага холерного вибриона CTXΦ ZOT, токсином zonula occludens), представляет собой АТФазу, содержащую функциональные и существенные мотивы Уокера. [ 22 ] Считается, что они опосредуют гидролиз АТФ, обеспечивая энергию для сборки фаговой нити. Нитчатый фаг Cf1t из Xanthomonas Campestris (предложенного NCBI вида Xanthomonas фаг Cf1t , incertae sedis внутри Inoviridae , вероятно, неправильно написанный как Cflt), [ 24 ] в 1987 году было показано, что они интегрируются в бактериальный геном хозяина, и с тех пор сообщалось о таких умеренных нитчатых фагах, многие из которых участвуют в патогенезе. [ 1 ]
Таксономия
[ редактировать ]Выделяют следующие роды: [ 7 ]
Филогенетические деревья и клады все чаще используются для изучения таксономии. [ 25 ] из иновирусид. [ 1 ] [ 3 ] [ 5 ] [ 26 ]
На основании метагеномных данных семейство было предложено разделить на новые семейства. Amplinoviridae , Protoinoviridae , Photinoviridae , Vespertilinoviridae , Densinoviridae и Paulinoviridae , все, конечно, относятся к отряду Tubulavirales . [ 27 ]
Известные участники
[ редактировать ]- род Inovirus, также известный как фаги Ff [ 8 ] [ 9 ] – они заражают Escherichia coli, несущую F-эпизому .
- вид вируса эшерихии M13
- вид Нитчатый бактериофаг fd (предложение)
- ФД фаг
- вид Вибрион вирус CTXphi
- Вибриофаг CTXphi (он же бактериофаг CTXφ)
- род Инфулавирус ) [ 11 ]
- вид вируса Escherichia If1
- Фаг If1
- род Lineavirus [ 12 ]
- вид вируса сальмонеллы IKe
- Айк фаг
- род Primolicivirus ) [ 13 ]
- вид Pseudomonas вирус Pf1
- Фаг Pf1
- род Tertilicivirus ) [ 14 ]
- вирус вида Pseudomonas Pf3 — бактериофаги, инфицирующие Pseudomonas aeruginosa.
- Фаг Pf3
Другими примечательными предлагаемыми видами являются:
- вид Thermus фаг PH75 [ 16 ]
- Фаг PH75
- вид Xanthomonas фаг Cf1t (вероятно, написано с ошибкой Cflt) [ 24 ]
- Фаг Cf1t
История
[ редактировать ]Нитчатая частица, видимая на электронных микрофотографиях, изначально была ошибочно интерпретирована как загрязняющая бактериальный пилус , но ультразвуковая деградация, которая разрывает гибкие нити примерно пополам, [ 29 ] инактивированная инфекционность, предсказанная для морфологии нитчатого бактериофага. [ 30 ] Три нитчатых бактериофага fd, f1 и M13 были выделены и охарактеризованы тремя различными исследовательскими группами в начале 1960-х годов. Поскольку эти три фага различаются менее чем на 2 процента по последовательностям ДНК, что соответствует изменениям всего в нескольких десятках кодонов во всем геноме, для многих целей их можно считать идентичными. [ 31 ] Дальнейшая независимая характеристика в последующие полвека определялась интересами этих исследовательских групп и их последователей. [ 2 ]
Нитчатые фаги, в отличие от большинства других фагов, постоянно проникают через бактериальную мембрану, не убивая хозяина. [ 19 ] Генетические исследования M13 с использованием условно летальных мутантов, инициированные Дэвидом Праттом и его коллегами, привели к описанию функций генов фага. [ 32 ] [ 33 ] Примечательно, что у инфицированных бактерий в больших количествах образуется белковый продукт гена 5, необходимый для синтеза одноцепочечной ДНК потомства. [ 34 ] [ 35 ] [ 36 ] и он связывается с зарождающейся ДНК, образуя линейный внутриклеточный комплекс. [ 17 ] (Простая нумерация генов арабскими цифрами 1,2,3,4..., введенная группой Пратта, иногда заменяется практикой использования римских цифр I, II, III, IV... но номера генов, определяемые эти две системы одинаковы).
Более длинная (или более короткая) ДНК может быть включена в фаг fd, поскольку во время сборки можно добавить больше (или меньше) белковых субъединиц, что необходимо для защиты ДНК, что делает фаг удобным для генетических исследований. [ 37 ] [ 38 ] На длину фага также влияет положительный заряд на длину внутренней поверхности капсида фага. [ 39 ] Геном fd был одним из первых полных геномов, секвенированных. [ 40 ]
Таксономия нитчатых бактериофагов была определена Андре Львоффом и Полем Турнье как семейство Inophagoviridae, род I. inophagovirus, вид Inophagovirus бактерии (Inos = волокно или нить по-гречески), с фагом fd (Гоффмана-Берлинга) в качестве типового вида. [ 41 ] [ 42 ] «Фаговирус» является тавтологичным , название семейства было изменено на Inoviridae , а типовой род — на Inovirus . Эта номенклатура сохранялась на протяжении многих десятилетий. [ 9 ] хотя определение fd как типового вида было заменено, поскольку M13 стал более широко использоваться для генетических манипуляций, [ 43 ] [ 44 ] и для изучения p8 в средах, имитирующих мембраны. [ 2 ] Число известных нитчатых бактериофагов многократно увеличилось с использованием подхода машинного обучения, и было предложено, чтобы «бывшее семейство Inoviridae было реклассифицировано в отряд, предварительно разделенный на 6 семейств-кандидатов и 212 подсемейств-кандидатов». [ 5 ] Фаги fd, f1, M13 и другие родственные фаги являются фагами Ff , для F- специфичных (они инфицируют Escherichia coli, несущих F-эпизому ) нитевидных фагов, если использовать концепцию народного названия. [ 45 ]
Нитчатые бактериофаги, созданные для отображения иммуногенных пептидов, полезны в иммунологии и более широких биологических приложениях. [ 46 ] [ 47 ] [ 48 ] [ 49 ] Джордж Смит и Грег Винтер использовали f1 и fd в своей работе по фаговому дисплею , за которую они были удостоены части Нобелевской премии по химии 2018 года. занимались созданием и использованием многих производных M13 для самых разных целей, особенно в материаловедении . Анжела Белчер и ее коллеги [ 49 ] [ 50 ] [ 51 ] [ 52 ] Нитчатый бактериофаг может способствовать толерантности к антибиотикам, образуя жидкокристаллические домены. [ 53 ] вокруг бактериальных клеток. [ 54 ] [ 10 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с Hay ID, Литгоу Т. (июнь 2019 г.). «Нитчатые фаги: мастера микробной экономики совместного использования» . Отчеты ЭМБО . 20 (6): e47427. дои : 10.15252/эмбр.201847427 . ПМК 6549030 . ПМИД 30952693 .
- ^ Jump up to: а б с д и ж г час Штраус С.К., Бо Х.Э. (2018). «Белки и сборка нитевидных бактериофагов». В Bhella JR, Харрис Д. (ред.). Вирусные белки и нуклеопротеиновые комплексы . Субклеточная биохимия. Том. 88. Спрингер Сингапур. стр. 261–279. дои : 10.1007/978-981-10-8456-0_12 . ISBN 978-981-10-8455-3 . ПМИД 29900501 .
- ^ Jump up to: а б Май-Прохнов А., Хуэй Дж.Г., Кьеллеберг С., Раконьяк Дж. , Макдугалд Д., Райс С.А. (июль 2015 г.). «Большие вещи в маленьких упаковках: генетика нитчатых фагов и влияние на приспособленность их хозяина» . Обзоры микробиологии FEMS . 39 (4): 465–87. дои : 10.1093/femsre/fuu007 . HDL : 10453/65260 . ПМИД 25670735 .
- ^ Jump up to: а б Раконьяк Дж., Рассел М., Ханум С., Брук С.Дж., Райич М. (2017). «Нитчатый фаг: структура и биология». В Лим Т.С. (ред.). Рекомбинантные антитела к инфекционным заболеваниям . Достижения экспериментальной медицины и биологии. Том. 1053. Международное издательство Springer. стр. 1–20. дои : 10.1007/978-3-319-72077-7_1 . ISBN 978-3-319-72076-0 . ПМИД 29549632 .
- ^ Jump up to: а б с д и Ру С., Крупович М., Дейли Р.А., Борхес А.Л., Найфач С., Шульц Ф. и др. (ноябрь 2019 г.). «Загадочные иновирусы, как выяснилось, широко распространены среди бактерий и архей во всех биомах Земли» . Природная микробиология . 4 (11): 1895–1906. дои : 10.1038/s41564-019-0510-x . ПМК 6813254 . ПМИД 31332386 .
- ^ Jump up to: а б с д «Иновиридиды ~ ViralZone» . www.viralzone.expasy.org . Проверено 31 марта 2021 г.
- ^ Jump up to: а б ICTV. «Таксономия вирусов: выпуск 2019 г.» . Проверено 4 июля 2020 г.
- ^ Jump up to: а б NCBI: иновирус (род)
- ^ Jump up to: а б с ICTV: История таксономии ICTV: Иновирус . 2019 EC 51, Берлин, Германия, июль 2019 г.; Ратификация по электронной почте, март 2020 г. (MSL № 35).
- ^ Jump up to: а б с Тарафдер А.К., фон Кюгельген А., Меллул А.Дж., Шульце У., Аартс Д.Г., Бхарат Т.А. (март 2020 г.). «Жидкокристаллические капли фага образуют окклюзионные оболочки, которые инкапсулируют и защищают инфекционные палочковидные бактерии» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (9): 4724–4731. Бибкод : 2020PNAS..117.4724T . дои : 10.1073/pnas.1917726117 . ПМК 7060675 . ПМИД 32071243 .
- ^ Jump up to: а б НКБИ: Инфлунавирус (род)
- ^ Jump up to: а б NCBI: Lineavirus (род)
- ^ Jump up to: а б NCBI: Примолицивирус (род)
- ^ Jump up to: а б NCBI: Тертилицивирус (род)
- ^ Jump up to: а б Секор, Патрик Р.; Бургенер, Элизабет Б.; Кинерсли, М.; Дженнингс, Лаура К.; Роман-Круз, Валерий; Попеску, Медея; Ван Беллегем, Йонас Д.; Хэддок, Наоми; Коупленд, Коннер; Майклс, Лия А.; де Врис, Кристиан Р. (2020). «Бактериофаг Pf и их влияние на вирулентность Pseudomonas, иммунитет млекопитающих и хронические инфекции» . Границы в иммунологии . 11 : 244. дои : 10.3389/fimmu.2020.00244 . ISSN 1664-3224 . ПМК 7047154 . ПМИД 32153575 .
- ^ Jump up to: а б NCBI: фаг Thermus PH75 (виды)
- ^ Jump up to: а б Пратт Д., Лоуз П., Гриффит Дж. (февраль 1974 г.). «Комплекс одноцепочечной ДНК бактериофага М13 и белка гена 5». Журнал молекулярной биологии . 82 (4): 425–39. дои : 10.1016/0022-2836(74)90239-3 . ПМИД 4594145 .
- ^ Грей CW (июль 1989 г.). «Трехмерная структура комплексов одноцепочечных ДНК-связывающих белков с ДНК. Белки гена IKe и fd 5 образуют левосторонние спирали с одноцепочечной ДНК». Журнал молекулярной биологии . 208 (1): 57–64. дои : 10.1016/0022-2836(89)90087-9 . ПМИД 2671388 .
- ^ Jump up to: а б Хоффманн Берлинг Х., Мейз Р. (март 1964 г.). «Высвобождение мужских бактериофагов из выживших бактерий-хозяев». Вирусология . 22 (3): 305–13. дои : 10.1016/0042-6822(64)90021-2 . ПМИД 14127828 .
- ^ Вэнь Дж. Д., Грей Д. М. (март 2004 г.). «Одноцепочечный ДНК-связывающий белок гена Ff 5 собирается на нуклеотидах, ограниченных шпилькой ДНК». Биохимия . 43 (9): 2622–34. дои : 10.1021/bi030177g . ПМИД 14992600 .
- ^ Рапоза, член парламента; Вебстер, РЛ (1995). «Продукты гена I и перекрывающегося внутрирамочного гена XI необходимы для сборки нитчатого фага». Дж. Мол. Биол . 248 (3): 627–638. дои : 10.1006/jmbi.1995.0247 . ПМИД 7752229 .
- ^ Jump up to: а б Ло, Белинда; Хаазе, Максимилиан; Мюллер, Лукас; Кун, Андреас; Лептин, Себастьян (9 апреля 2017 г.). «Белок трансмембранного морфогенеза gp1 нитчатых фагов содержит мотивы Walker A и Walker B, необходимые для сборки фага» . Вирусы . 9 (4): 73. дои : 10.3390/v9040073 . ПМЦ 5408679 . ПМИД 28397779 .
- ^ Jump up to: а б Ло, Белинда; Кун, Андреас; Лептин, Себастьян (май 2019 г.). «Увлекательная биология фагового дисплея: сборка нитевидных фагов» . Молекулярная микробиология . 111 (5): 1132–1138. дои : 10.1111/mmi.14187 . ПМИД 30556628 .
- ^ Jump up to: а б NCBI: фаг Xanthomonas Cf1t (виды)
- ^ Исполнительный комитет Международного комитета по таксономии вирусов (май 2020 г.). «Новая сфера таксономии вирусов: разделение виросферы на 15 иерархических рангов» . Природная микробиология . 5 (5): 668–674. дои : 10.1038/s41564-020-0709-x . ПМК 7186216 . ПМИД 32341570 .
- ^ Казлаускас Д., Варшани А., Кунин Е.В., Крупович М. (июль 2019 г.). «Множественное происхождение прокариотических и эукариотических одноцепочечных ДНК-вирусов из бактериальных и архейных плазмид» . Природные коммуникации . 10 (1): 3425. Бибкод : 2019NatCo..10.3425K . дои : 10.1038/s41467-019-11433-0 . ПМК 6668415 . PMID 31366885 .
- ^ Саймон Ру, Март Крупович, Ребекка А. Дейли, Адэр Л. Борхес, Стивен Найфак, Фредерик Шульц, Эмили А. Эло-Фадрош и др. : Выявлено, что загадочные иновирусы широко распространены среди бактерий и архей во всех биомах Земли . В: Природная микробиология, 22 июля 2019 г., doi:10.1038/s41564-019-0510-x.
- ^ NCBI: Аффертхолерамвирус (род)
- ^ Фрайфельдер Д., Дэвисон П.Ф. (май 1962 г.). «Исследования звуковой деградации дезоксирибонуклеиновой кислоты» . Биофизический журнал . 2 (3): 235–47. Бибкод : 1962BpJ.....2..235F . дои : 10.1016/S0006-3495(62)86852-0 . ПМЦ 1366369 . ПМИД 13894963 .
- ^ Марвин Д.А., Хоффманн-Берлинг Х (1963). «Физические и химические свойства двух новых малых бактериофагов». Природа . 197 (4866): 517–518. Бибкод : 1963Natur.197..517M . дои : 10.1038/197517b0 . hdl : 11858/00-001M-0000-002D-442F-1 . S2CID 4224468 .
- ^ Мораг О., Абрамов Г., Голдборт А. (декабрь 2011 г.). «Сходства и различия внутри членов семейства Ff вирусов нитчатых бактериофагов» . Журнал физической химии Б. 115 (51): 15370–9. дои : 10.1021/jp2079742 . ПМИД 22085310 .
- ^ Пратт Д., Цаголов Х., Эрдал В.С. (ноябрь 1966 г.). «Условно-летальные мутанты малого нитчатого колифага М13. I. Выделение, комплементация, уничтожение клеток, время действия цистронов». Вирусология . 30 (3): 397–410. дои : 10.1016/0042-6822(66)90118-8 . ПМИД 5921643 .
- ^ Пратт Д., Цаголов Х., Бодуэн Дж. (сентябрь 1969 г.). «Условно-летальные мутанты малого нитчатого колифага M13. II. Два гена белков оболочки». Вирусология . 39 (1): 42–53. дои : 10.1016/0042-6822(69)90346-8 . ПМИД 5807970 .
- ^ Пратт Д., Эрдал В.С. (октябрь 1968 г.). «Генетический контроль синтеза ДНК бактериофага М13». Журнал молекулярной биологии . 37 (1): 181–200. дои : 10.1016/0022-2836(68)90082-X . ПМИД 4939035 .
- ^ Генри Т.Дж., Пратт Д. (март 1969 г.). «Белки бактериофага М13» . Труды Национальной академии наук Соединенных Штатов Америки . 62 (3): 800–7. Бибкод : 1969PNAS...62..800H . дои : 10.1073/pnas.62.3.800 . ПМК 223669 . ПМИД 5257006 .
- ^ Альбертс Б., Фрей Л., Делиус Х. (июль 1972 г.). «Выделение и характеристика белка гена 5 нитчатых бактериальных вирусов». Журнал молекулярной биологии . 68 (1): 139–52. дои : 10.1016/0022-2836(72)90269-0 . ПМИД 4115107 .
- ^ Херрманн Р., Нойгебауэр К., Центграф Х., Шаллер Х. (февраль 1978 г.). «Транспозиция последовательности ДНК, определяющей устойчивость к канамицину, в одноцепочечный геном бактериофага fd». Молекулярная и общая генетика . 159 (2): 171–8. дои : 10.1007/bf00270890 . ПМИД 345091 . S2CID 22923713 .
- ^ Саттар, Садия; Беннетт, Николас Дж.; Вэнь, Уэсли X.; Гатри, Дженнесс М.; Блэквелл, Лен Ф.; Конвей, Джеймс Ф.; Раконьяц, Ясна (2015). «Ff-нано, короткие функционализированные наностержни, полученные из нитчатого бактериофага Ff (f1, fd или M13)» . Границы микробиологии . 6 : 316. дои : 10.3389/fmicb.2015.00316 . ISSN 1664-302X . ПМЦ 4403547 . ПМИД 25941520 .
- ^ Гринвуд Дж., Хантер Дж.Дж., Перхэм Р.Н. (январь 1991 г.). «Регуляция длины нитчатого бактериофага путем модификации электростатических взаимодействий между белком оболочки и ДНК». Журнал молекулярной биологии . 217 (2): 223–7. дои : 10.1016/0022-2836(91)90534-д . ПМИД 1992159 .
- ^ Бек Э., Зоммер Р., Ауэрсвальд Э.А., Курц С., Цинк Б., Остербург Г. и др. (декабрь 1978 г.). «Нуклеотидная последовательность ДНК бактериофага» . Исследования нуклеиновых кислот . 5 (12): 4495–503. дои : 10.1093/нар/5.12.4495 . ПМЦ 342768 . ПМИД 745987 .
- ^ Львофф А, Турнье П (1966). «Классификация вирусов». Ежегодный обзор микробиологии . 20 (1): 45–74. дои : 10.1146/annurev.mi.20.100166.000401 . ПМИД 5330240 .
- ^ Мэтьюз Р.Э. (1982). «Классификация и номенклатура вирусов. Четвертый доклад Международного комитета по таксономии вирусов» . Интервирусология . 17 (1–3): 1–199. дои : 10.1159/000149278 . ПМИД 6811498 .
- ^ Мессинг, Иоахим (1991). «Клонирование фага М13 или как лучше всего использовать биологию» . Джин . 100 : 3–12. дои : 10.1016/0378-1119(91)90344-Б . ISSN 0378-1119 . ПМИД 2055478 .
- ^ Мессинг, Иоахим (1996). «Клонирование одноцепочечной ДНК». Молекулярная биотехнология . 5 (1): 39–47. дои : 10.1007/BF02762411 . ПМИД 8853015 . S2CID 33495106 .
- ^ Гиббс А.Дж., Харрисон Б.Д., Уотсон Д.Х., Уайлди П. (январь 1966 г.). «Что в названии вируса?». Природа . 209 (5022): 450–4. Бибкод : 1966Natur.209..450G . дои : 10.1038/209450a0 . ПМИД 5919575 . S2CID 4288812 .
- ^ Смит ГП (июнь 1985 г.). «Нитевидный слитый фаг: новые векторы экспрессии, которые отображают клонированные антигены на поверхности вириона». Наука . 228 (4705): 1315–7. Бибкод : 1985Sci...228.1315S . дои : 10.1126/science.4001944 . ПМИД 4001944 .
- ^ Приско А., Де Берардинис П. (24 апреля 2012 г.). «Нитчатый бактериофаг ФД как система доставки антигена при вакцинации» . Международный журнал молекулярных наук . 13 (4): 5179–94. дои : 10.3390/ijms13045179 . ПМЦ 3344273 . ПМИД 22606037 .
- ^ Сиуд М (апрель 2019 г.). «Библиотеки фагового дисплея: от связующих к адресной доставке лекарств и терапии человека». Молекулярная биотехнология . 61 (4): 286–303. дои : 10.1007/s12033-019-00156-8 . ПМИД 30729435 . S2CID 73434013 .
- ^ Jump up to: а б Генри К.А., Арбаби-Гахруди М., Скотт Дж.К. (2015). «За пределами фагового дисплея: нетрадиционные применения нитчатого бактериофага в качестве носителя вакцин, терапевтических биологических препаратов и каркаса для биоконъюгации» . Границы микробиологии . 6 : 755. дои : 10.3389/fmicb.2015.00755 . ПМЦ 4523942 . ПМИД 26300850 .
- ^ Кейси Дж.П., Барберо Р.Дж., Хелдман Н., Белчер А.М. (ноябрь 2014 г.). «Универсальная активность ферментов de novo в капсидных белках из сконструированной библиотеки бактериофагов M13». Журнал Американского химического общества . 136 (47): 16508–14. дои : 10.1021/ja506346f . ПМИД 25343220 .
- ^ О Д., Ци Дж., Хан Б., Чжан Дж., Карни Т.Дж., Омура Дж. и др. (август 2014 г.). «Вирусно-направленный синтез наноструктурированных оксидов металлов для литий-кислородных аккумуляторов». Нано-буквы . 14 (8): 4837–45. Бибкод : 2014NanoL..14.4837O . дои : 10.1021/nl502078m . ПМИД 25058851 .
- ^ Дорваль Куршен Н.М., Клуг М.Т., Хуанг К.Дж., Вайдман М.С., Канту В.Дж., Чен П.Ю. и др. (июнь 2015 г.). «Создание многофункциональных нанопористых композитов с вирусным шаблоном для тонкопленочных солнечных элементов: вклад морфологии и оптики в генерацию фототока». Журнал физической химии C. 119 (25): 13987–4000. дои : 10.1021/acs.jpcc.5b00295 . hdl : 1721.1/102981 . ISSN 1932-7447 .
- ^ Догич З (30 июня 2016 г.). «Нитевидные фаги как модельная система в физике мягких веществ» . Границы микробиологии . 7 : 1013. дои : 10.3389/fmicb.2016.01013 . ПМЦ 4927585 . ПМИД 27446051 .
- ^ Secor PR, Дженнингс Л.К., Майклс Л.А., Свир Дж.М., Сингх П.К., Parks WC, Bollyky PL (декабрь 2015 г.). «Матрица биопленки Pseudomonas aeruginosa в жидкий кристалл» . Микробная клетка . 3 (1): 49–52. doi : 10.15698/mic2016.01.475 (неактивен 13 марта 2024 г.). ПМК 5354590 . ПМИД 28357315 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на март 2024 г. ( ссылка )