Планктомицетота
| Планктомицетота | |
|---|---|

| |
| Брокадия анаммоксиданс | |
| Научная классификация | |
| Домен: | Бактерии |
| Супертип: | Суперфилум ПВХ |
| Тип: | Планктомицетота Гаррити и Холт 2021 [ 1 ] |
| Занятия и заказы [ 2 ] [ 3 ] | |
| |
| Синонимы | |
| |
Planctomycetota , — это тип широко распространенных бактерий , встречающихся как в водной так и в наземной среде обитания. [ 5 ] Они играют значительную роль в глобальных циклах углерода и азота, причем многие виды этого типа способны к анаэробному окислению аммония, также известному как анаммокс . [ 5 ] [ 6 ] Многие Planctomycetota встречаются в относительно высокой численности в виде биопленок . [ 7 ] часто связываясь с другими организмами, такими как макроводоросли и морские губки . [ 8 ]
Planctomycetota включены в супертип PVC наряду с Verrucomicrobiota , Chlamydiota , Lentisphaerota , Kiritimatiellaeota и Candidatus Omnitropica . [ 9 ] [ 10 ] Тип Planctomycetota состоит из классов Planctomycetia и Phycisphaerae. Впервые описанные в 1924 году, представители рода Planctomycetota были идентифицированы как эукариоты и лишь позже, в 1972 году, были описаны как бактерии. [ 5 ] Раннее исследование представителей Planctomycetota позволило предположить, что клеточный план значительно отличается от других бактерий, хотя теперь подтверждено, что они являются грамотрицательными бактериями , но со многими уникальными характеристиками.
Бактерии Planctomycetota часто представляют собой маленькие сферические клетки, но наблюдается большое количество морфологических вариаций. [ 11 ] Члены Planctomycetota также демонстрируют различные репродуктивные привычки: многие виды делятся путем почкования , в отличие от всех других свободноживущих бактерий, которые делятся путем бинарного деления . [ 5 ] [ 12 ] [ 13 ]
Интерес к Planctomycetota растет с точки зрения биотехнологии и применения человеком, главным образом в качестве источника биоактивных молекул. [ 14 ] Кроме того, некоторые Planctomycetota недавно были описаны как патогены человека. [ 8 ]
Вид Gemmata obscuriglobus был идентифицирован как включающий бактерии с уникальными характеристиками среди Planctomycetota. [ 15 ] [ 16 ] такие как их способность синтезировать стерины . [ 5 ] [ 17 ] [ 15 ]
Структура и морфология
[ редактировать ]Форма клеток и придатки
[ редактировать ]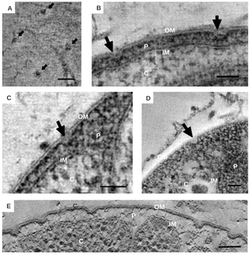
Отличительные морфологические характеристики бактерий Planctomycetota широко обсуждались. [ 6 ] Общая морфология часто представляет собой сферические клетки диаметром примерно 2 мкм, как это наблюдается у вида Aquisphaera giovannonii . Однако разнообразие формы клеток у них часто сильно варьирует. У некоторых видов описаны овоидные и грушевидные клетки, которые часто встречаются в розетках из 3-10 клеток. [ 11 ] Gemmata obscuriglobus — хорошо изученный вид Planctomycetota со сферическими клетками. Напротив, бактерии вида Planctopirus limnophila имеют овальные клетки. [ 15 ]
Многие виды Planctomycetota имеют структуры и придатки на внешней поверхности клетки. Жгутики , распространенные у большинства бактерий, также наблюдались у вида P. limnophila. [ 5 ] [ 11 ] [ 18 ] У многих Planctomycetota также есть опора или ножка, с помощью которой клетка прикрепляется к поверхности или субстрату. [ 5 ] [ 18 ] Однако представителям некоторых видов, таких как Isosphaera pallida, не хватает опоры. [ 5 ]
Были обнаружены уникальные придатки, известные как кратерообразные структуры. [ 5 ] [ 11 ] [ 18 ] у видов Planctomycetota, принадлежащих к классу Planctomycetia. [ 13 ] На внешней поверхности клеток вида P. limnophila наблюдаются как крупные, так и мелкие кратерообразные структуры. Крупные кратерные структуры часто покрывают поверхность клетки, тогда как мелкие кратерные структуры часто располагаются только на конце клетки. Световая микроскопия выявила волокна как стебельчатого, так и пильного типа у P. limnophila и G. obscuriglobus . Волокна пилей у обоих этих видов часто ассоциировались с крупными кратерообразными структурами; напротив, волокна стебля были связаны с небольшими кратерообразными структурами. [ 18 ]
Состав клеточной стенки
[ редактировать ]Раннее изучение Planctomycetota показало, что их клеточный план значительно отличается как от грамположительных, так и от грамотрицательных бактерий. [ 5 ] До недавнего времени считалось, что у бактерий Planctomycetota отсутствуют пептидогликаны в клеточных стенках, и вместо этого предполагалось, что клеточные стенки имеют белковую структуру. Пептидогликан — важный полимер гликанов, присутствующий во всех свободноживущих бактериях, а его жесткость помогает поддерживать целостность клетки. Синтез пептидогликана также важен во время деления клеток . Недавно было обнаружено, что у представителей вида G. obscuriglobus в клеточных стенках имеется пептидогликан. [ 5 ] [ 18 ]
Внутренний клеточный состав
[ редактировать ]Когда-то считалось, что Planctomycetota демонстрирует отчетливую компартментализацию внутри цитозоля . [ 5 ] [ 18 ] трехмерной электронной томографии Реконструкция G. obscuriglobus продемонстрировала различные интерпретации этой предполагаемой компартментализации. [ 16 ] Было предложено разделить цитозоль на компартменты (парифоплазму и пиреллулосому) внутрицитоплазматической мембраной. С тех пор было показано, что эта интерпретация неверна. Фактически, хорошо известно, что интрацитоплазматическая мембрана представляет собой цитоплазматическую мембрану, которая имеет уникальные впячивания , создающие видимость компартментализации внутри цитозоля. [ 5 ] [ 16 ] [ 18 ] Таким образом, Planctomycetota имеет два отдела, типичные для грамотрицательных бактерий: цитоплазму и периплазму .
Избыток мембраны, наблюдаемый у G. obscuriglobus, утрояет площадь поверхности клетки по отношению к ее объему , что, как предполагается, связано с синтезом стеринов. [ 16 ]
Пигменты
[ редактировать ]Многие виды Planctomycetota имеют розовую или оранжевую окраску, что, как предполагается, является результатом выработки каротиноидных пигментов. Каротиноиды производятся растениями и грибами , а также некоторыми гетеротрофными бактериями для защиты от окислительного стресса . Три разных каротиноидных пигмента были идентифицированы в двух разных штаммах Planctomycetota. [ 19 ]
В морской среде Planctomycetota часто находятся во взвешенном состоянии в толще воды или присутствуют в виде биопленок на поверхности макроводорослей и часто подвергаются вредному ультрафиолетовому излучению. Более пигментированные виды Planctomycetota более устойчивы к ультрафиолетовому излучению, хотя это еще не совсем понятно. [ 20 ] С тех пор было показано, что Planctomycetota синтезирует каротиноиды C30 из сквалена и что этот путь сквалена к каротиноидам C30 является наиболее распространенным у прокариот. [ 21 ]
Уникальные характеристики клеток анаммокса
[ редактировать ]Бактерии Planctomycetota, способные к анаммоксу, образуют отряд Brocadiales. [ 22 ] Клетки анаммокс-бактерий часто кокковидные, диаметром около 0,8 мкм. [ 7 ] Предполагается, что они содержат три отсека, каждый из которых окружен мембраной. Наружная мембрана окружает клетку и протоплазму, а самая внутренняя мембрана окружает анаммоксосому, центральную структуру анаммокс-бактерий. [ 18 ] [ 23 ] Мембрана анаммоксосомы в основном состоит из необычных липидов на основе ладдерана . [ 23 ]
История жизни и воспроизводство
[ редактировать ]Рост
[ редактировать ]Виды Planctomycetota растут медленно по сравнению с другими бактериями. [ 5 ] [ 10 ] [ 7 ] [ 24 ] часто образует розетки из 3-5 клеток. [ 5 ] [ 24 ] Предполагается, что вид P. limnophila относительно быстрорастет. [ 5 ] [ 25 ] с периодом удвоения примерно 6-14 дней. Напротив, у некоторых других Planctomycetota время удвоения составляет около 30 дней. [ 25 ] Их высокая численность во многих экосистемах удивляет, учитывая медленные темпы их роста. [ 7 ] [ 10 ]
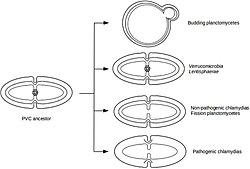
Жизненный цикл
[ редактировать ]Planctomycetota часто переключает образ жизни между стадией сидячего стебля и стадией свободного плавания. [ 24 ] Представители вида P. limnophila меняют образ жизни, что часто связано с делением клеток. Сидячая материнская клетка производит свободноплавающую дочернюю клетку. Затем дочерняя клетка должна прикрепиться к поверхности, прежде чем начать цикл заново. Однако не все Planctomycetota имеют подвижную стадию, и смена образа жизни, наблюдаемая у многих видов, может быть не характерна для всех Planctomycetota. [ 5 ]

Воспроизведение
[ редактировать ]Современное понимание деления бактериальных клеток основано на модельных организмах, таких как Escherichia coli . [ 15 ] Доминирующей формой размножения, наблюдаемой почти у всех бактерий, является деление клеток путем бинарного деления , которое включает синтез как пептидогликанов, так и белков, известных как FtsZ . [ 15 ] [ 26 ] Напротив, многие бактерии Planctomycetota делятся путем почкования . [ 5 ] [ 12 ] [ 13 ]
Предполагается, что белки FtsZ по структуре аналогичны структуре тубулина , белка, присутствующего в эукариотах. [ 27 ] и необходим для формирования перегородок во время деления клеток. [ 5 ] [ 6 ] Отсутствие белков FtsZ часто приводит к летальному исходу. [ 5 ] Пептидогликан также играет значительную роль в делении клеток путем бинарного деления. [ 26 ]
Planctomycetota — один из немногих известных типов, у представителей которого отсутствуют белки FtsZ. [ 5 ] [ 26 ] [ 27 ] У бактерий Chlamydiales, также принадлежащих к супертипу PVC, также отсутствует FtsZ. [ 27 ] Хотя у бактерий Planctomycetota отсутствует FtsZ, наблюдаются два различных способа деления клеток. [ 5 ] Большинство Planctomycetata делятся путем бинарного деления, в основном виды класса Phycisphaerae. Напротив, виды класса Planctomycetia делятся путем почкования. [ 5 ] [ 12 ] [ 13 ]
Механизмы, участвующие в почковании, подробно описаны для дрожжевых клеток. Однако почкование бактерий, наблюдаемое у Planctomycetota, до сих пор плохо изучено. [ 15 ] Почкование наблюдалось как в радиально-симметричных клетках, таких как бактерии вида P. limnophila , так и в аксиально-симметричных клетках. [ 13 ] Во время деления клеток у представителей P. limnophila дочерние клетки происходят из области, противоположной полюсу с опорой или ножкой.
Значительное разнообразие наблюдается в делении клеток среди бактерий Planctomycetota. [ 12 ] [ 13 ] Во время деления клеток Fuerstia marisgermanicae трубчатая структура соединяется от почки с материнской клеткой. [ 5 ] [ 22 ] Вид Kolteria novifilia образует отдельную кладу Planctomycetota и является единственным известным видом, который делится боковым отпочкованием в середине клетки. Наконец, представители клады Saltatorellus способны переключаться между бинарным делением и почкованием. [ 12 ] [ 13 ]
Генетические характеристики
[ редактировать ]Молекулярные подписи
[ редактировать ]Planctomycetota известны своими необычными клеточными характеристиками, а их отличие от всех других бактерий дополнительно подтверждается общим присутствием двух консервативных сигнатурных инделей (CSI). [ 28 ] Эти CSI отделяют группу от соседних типов внутри группы PVC. [ 29 ] Был обнаружен дополнительный CSI, общий для всех видов Planctomycetota, за исключением Kuenenia stuttgartiensis. Это подтверждает идею о том, что K. stuttgartiensis образует глубокую ветвь внутри типа Planctomycetota.
Также было обнаружено, что CSI присущ всему супертипу PVC, включая Planctomycetota. [ 28 ] [ 29 ] Planctomycetota также содержит важный консервативный сигнатурный белок, который, как было охарактеризовано, выполняет важную функцию домашнего хозяйства, эксклюзивную для членов, принадлежащих к супертипу PVC. [ 30 ]
Общие характеристики
[ редактировать ]Размер генома Rhodopirellula baltica оценивается более чем в 7 миллионов оснований, что делает его одним из крупнейших секвенированных прокариотических геномов . Обширное дублирование генома занимает около 25% последовательности генома. [ 6 ] Это может быть способом организма адаптации к мутациям , допускающим дублирование в случае повреждения части генома. Используемый праймер для полимеразной цепной реакции часто не соответствует генам, что создает трудности при секвенировании генома. [ 9 ]
При сравнении под микроскопом определяющей характеристикой некоторых Planctomycetota является то, что рРНК один несвязанный оперон рядом с началом может быть идентифицирован . Изменения генетического материала происходят за счет внутренней хромосомной инверсии, а не за счет латерального переноса генов. Это создает возможность диверсификации вариантов Planctomycetota, поскольку множественные гены транспозонов в этих регионах имеют обратную ориентацию, которая переходит в перестройки.
Некоторые Planctomycetota процветают в регионах, содержащих высококонцентрированные нитраты . [ 6 ] и имеют гены, необходимые для ферментации гетеротактической кислоты. лактатдегидрогеназа фермент . Ключевую роль в этом процессе играет Генетический процесс также имеет реакцию защиты от ультрафиолетового излучения и связан с генами RecA, lexA, uvrA, uvrB и uvrC , в дополнение к гену фотолиазы , который экспрессируется, когда окружающая среда подвергается чрезмерному стрессу от ультрафиолетового излучения. Другие реакции на стресс включают разложение перекиси водорода и окисление .
Многие Planctomycetota также экспрессируют сульфатазы гены . Геном Pirellula sp. Штамм 1 включает 110 генов, которые способствуют кодированию белков, продуцирующих ферменты сульфатазы. По сравнению с другим видом прокариот, Pseudomonas aeruginosa, встречается только 6 сульфатаз, а гены, экспрессирующие эти белки, содержатся в виде двух-пяти пар, обычно сгруппированных в 22 группы. [ 6 ]
Молекулярная эволюция
[ редактировать ]Planctomycetota происходят из бактерий, и эти сходства между белками Planctomycetales и эукариот отражают конвергентную эволюцию . Полученные семейства белков Gemmataceae , подгруппы Planctomycetota, имеют низкое сходство последовательностей с эукариотическими белками; однако они демонстрируют наибольшее сходство последовательностей с другими семействами белков Gemmataceae. [ 31 ]
происходит массовое появление новых семейств белков Внутри Gemmataceae . Путем дупликаций и реаранжировок доменов было получено более тысячи семейств белков. Новые паралоги участвуют в передаче сигналов , регуляторных системах и путях взаимодействия белков. Они связаны с функциональной организацией клетки, которую можно интерпретировать как адаптацию к более сложному образу жизни. [ 31 ] Длина белка у Gemmataceae больше, чем у большинства других бактерий, и гены имеют линкеры. Существует перекрытие между самыми длинными белками у Planctomycetales и самыми короткими белками у эукариот. С точки зрения паралогии генов, длины белка и структуры белковых доменов прокариоты и эукариоты не имеют резких границ. [ 31 ]
Филогения
[ редактировать ]Первоначально классифицированный как эукариот из-за морфологии, появление генетического секвенирования позволило исследователям согласиться с тем, что Planctomycetota принадлежит к домену Bacteria. [ 5 ] В этом домене Planctomycetota классифицируются как отдельный тип, однако другие исследователи утверждают, что их также можно отнести к более крупному супертипу под названием PVC, который будет включать типы Verrucomicrobia, Chlamydiae и Lentisphaerae, а также тип-кандидат Candidatus Omnitropica. ". [ 9 ] Было обнаружено, что внутри этого супертипа его члены тесно связаны посредством создания деревьев 16S рРНК. И Planctomycetota, и Chlamydiota кодируют белки- переносчики нуклеотидов , а также было обнаружено, что Verrucomicrobiota имеет особенности, общие для эукариотических клеток. Таким образом, общий предок этого супертипа мог быть началом эукариотической линии. [ 9 ] Хотя это одно из возможных объяснений, поскольку ПВХ не является началом бактериального дерева. [ 32 ] существование эукариотических признаков и генов, скорее всего, объясняется латеральным переносом генов, а не более поздним предком эукариот. [ 9 ]
| на основе 16S рРНК ДП _12_2021 г. [ 33 ] [ 34 ] [ 35 ] | 120 однокопийных маркерных белков на основе GTDB 08-RS214 [ 36 ] [ 37 ] [ 38 ] |
|---|---|
Экология
[ редактировать ]Распространение и численность
[ редактировать ]Представители Planctomycetota встречаются в самых разных средах, как географически, так и экологически. [ 39 ] и встречаются как в водной, так и в наземной среде обитания. [ 5 ] В водной среде они встречаются как в пресноводных, так и в морских системах. [ 39 ] Первоначально считалось, что Planctomycetota существуют исключительно в водной среде, но теперь известно, что они также в изобилии обитают в почвах. [ 40 ] и гиперсоленой среды. [ 41 ] Они распространены на пяти континентах, включая Антарктиду и Австралию . [ 40 ] [ 39 ]
Флуоресцентная in situ гибридизация использовалась для обнаружения Planctomycetota в различных средах, а Planctomycetota в изобилии обнаружены в сфагновых болотах. Некоторые Planctomycetota были обнаружены в пищеварительных системах морских форм жизни, тогда как другие, как правило, обитают среди эукариот. [ 9 ]
Влияние окружающей среды на распространение
[ редактировать ]Planctomycetota составляют примерно 11% прокариотических сообществ в морских системах, и их обширное распространение демонстрирует их способность обитать в самых разных средах. Они также могут адаптироваться как к аэробным, так и к анаэробным условиям. На их распределение могут влиять многие факторы, такие как влажность, уровень кислорода и уровень pH. Разнообразие и численность Planctomycetota тесно связаны с относительной влажностью. Влияние уровня кислорода демонстрирует энергетические потребности человека. Многие виды Planctomycetota хемогетеротрофны, в том числе G. obscuriglobus . Thermostilla marina , термофильный анаэробный вид, населяющий регионы гидротермальных источников , может использовать элементарную серу для образования сульфида и дышать нитратом . Planctomycetota также может обитать в регионах с уровнем pH от 4,2 до 11,6. [ 8 ]
Экологические последствия и глобальный углеродный цикл
[ редактировать ]Planctomycetota оказывают значительное влияние на глобальную биогеохимию и климат благодаря своей способности минерализовать и расщеплять частицы детрита в толще воды. [ 6 ] [ 20 ]
Planctomycetota играют значительную роль в глобальном углеродном цикле . [ 5 ] [ 6 ] [ 13 ] [ 42 ] Как облигатные, так и факультативные аэробные хемогетеротрофы , основным источником углерода, используемым Planctomycetota, являются углеводы . Многие Planctomycetota обладают способностью расщеплять чрезвычайно сложные углеводы, делая эти питательные вещества доступными для других организмов. Эта способность перерабатывать углерод связана со специфическими генами метаболизма C1, наблюдаемыми у многих Planctomycetota, и предполагается, что она играет значительную роль, но эта область исследований все еще плохо изучена.
Planctomycetota также содержит множество сульфатаз ферментов , которые способны расщеплять сульфатированные гетерополисахариды, вырабатываемые многими группами макроводорослей. Расщепление этих сульфатированных гетерополисахаридов Planctomycetota затем используется в качестве источника энергии. Предполагается, что некоторые Planctomycetota способны расщеплять каррагинан . [ 42 ]

Ассоциация с другими организмами
[ редактировать ]Planctomycetota часто наблюдалась в сочетании со многими организмами, включая макроводоросли, микроводоросли, морские губки и растения, такие как лишайники и мохообразные . [ 8 ] Их также наблюдали в глубоководных холодных просачиваниях , где они являются доминирующими организмами, живущими на трубчатых червях . [ 5 ]
Макроводоросли
[ редактировать ]
Planctomycetota часто обитают на морской поверхности с высоким содержанием питательных веществ. Они встречаются в виде биопленок на поверхности водорослей в относительно большом количестве. [ 7 ] Предполагается, что макроводоросли, такие как водоросли Laminaria Hyperborea и Ecklonia radiata, являются важной средой обитания для Planctomycetota. [ 5 ] [ 43 ] Примерно 70% бактериального сообщества Ecklonia radiata составляли Planctomycetota. [ 5 ] [ 10 ] Из биопленок макроводорослей выделено почти 150 видов Planctomycetota, и эти сообщества, связанные с макроводорослями, в основном не зависят от изменений в географическом распространении. Это предполагает симбиотические отношения. [ 8 ]
Леса водорослей доминируют на скалистых побережьях температурных регионов и обеспечивают среду обитания, укрытие и пищу для многих организмов, включая Planctomycetota. [ 5 ] Учитывая значительную роль лесов ламинарии в первичной продуктивности прибрежных районов , ассоциация Planctomycetota с ламинарией может указывать на их значительную роль в прибрежных местообитаниях. [ 44 ] Planctomycetota также играют важную роль в качестве компонентов детрита в толще воды, также известного как морской снег . [ 5 ] [ 44 ] учитывая их способность прикрепляться к поверхностям. [ 45 ]
Поскольку климат продолжает нагреваться, численность Planctomycetota, связанной с макроводорослями, может увеличиться. Водоросли Caulerpa Taxifolia инкубировали в условиях более высокого содержания CO 2 , и численность Planctomycetota существенно увеличилась, у некоторых видов до 10 раз. [ 5 ]
Цветение микроводорослей и диатомей
[ редактировать ]Хотя макроводоросли являются хорошо известными субстратами для сообществ Planctomycetota, также известно, что их численность коррелирует с цветением микроводорослей , таких как диатомеи. [ 44 ] [ 5 ] Цветение цианобактерий , диатомей и динофлагеллят обеспечивает питательными веществами Planctomycetota, что может объяснить эту связь. [ 8 ]
Морские губки
[ редактировать ]Виды Planctomycetota часто обитают на поверхности морских губок. [ 8 ] [ 45 ] Они взаимодействуют с губками либо путем прикрепления к опоре, либо посредством симбиотических отношений. Высокое разнообразие Planctomycetota присутствует в виде биопленок на губках. Симбиотические отношения между губками и Planctomycetota способствуют здоровью губки, а губка часто обеспечивает подходящую среду обитания и питательные вещества для Planctomycetota. [ 8 ]
Лишайниковые сообщества и сфагновые болота
[ редактировать ]Было обнаружено, что Planctomycetota широко распространены в сообществах лишайников на всей территории Северо-Западной Сибири и демонстрируют чрезвычайно высокое разнообразие. Planctomycetota также была связана с сообществами лишайников и водно-болотными угодьями Sphagnum . Сфагновые водно-болотные угодья хранят большое количество углерода, внося свой вклад в глобальный углеродный цикл. Planctomycetota играют значительную роль в деградации сфагнума, составляя примерно 15% бактериального сообщества. [ 8 ]
Другие бактериальные сообщества
[ редактировать ]Planctomycetota демонстрирует ассоциации с другими бактериальными сообществами, главным образом с Alphaproteobacteria , Bacteroidota , Gemmatimonadota и Verrucomicrobiota . Рост многих Planctomycetota часто поддерживается необходимыми питательными веществами, предоставляемыми другими бактериями сообщества, а некоторые Planctomycetota в значительной степени полагаются на симбиотические отношения с другими бактериями. [ 8 ]
Физиология
[ редактировать ]Эндоцитоз
[ редактировать ]Существование белков мембранной оболочки вблизи интрацитоплазматической мембраны может быть использовано для системы поглощения, подобной эндоцитозу , что будет первым случаем, когда эта функция будет обнаружена за пределами эукариотического домена. Однако теперь, когда существование жесткой клеточной стенки пептидогликана подтверждено, способность этих везикул проходить через эту клеточную стенку кажется маловероятной. Кроме того, удаление одного из этих белков мембранной оболочки у P. limnophila не выявило снижения поглощения макромолекул . [ 18 ] Кроме того, с помощью трехмерной реконструкции Planctomycetota на основе криоэлектронной томографии было обнаружено, что то, что первоначально считалось везикулами, удерживаемыми в периплазме, на самом деле является всего лишь складками цитоплазматической мембраны. [ 5 ] Тем не менее, было продемонстрировано, что Planctomycetota могут выжить на полисахаридах с высокой молекулярной массой в качестве единственного источника углерода, а это означает, что они должны обладать способностью включать сложные углеродные субстраты в свою цитоплазму. Были выдвинуты три гипотезы: во-первых, Planctomycetota выделяют фермент, который за пределами клеточной стенки расщепляет сложные субстраты на более мелкие моносахариды, которые легче транспортируются через различные мембраны. Во-вторых, сложные субстраты закрепляются снаружи Planctomycetota, которые затем способны медленно расщеплять эти субстраты на олигосахариды, которые могут транспортироваться в периплазму Planctomycetota с помощью специализированных белков. Третья гипотеза связана с кратерообразными структурами, обнаруженными на внешней стороне клеточных стенок Planctomycetota. Эти структуры имеют волокна, выстилающие их ямки, которые могут поглощать целые полисахариды в периплазму, где они затем перевариваются. [ 18 ]
Осмотическая регуляция
[ редактировать ]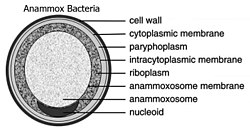
Почти все бактерии имеют цитозоль, повторяющий внешнюю форму клеточной стенки пептидогликана. Эукариоты отличаются тем, что их цитозоль разделен на несколько отсеков для создания органелл, таких как ядро . Planctomycetota уникальны тем, что у них есть большие инвагинации цитоплазматической мембраны, отрывающиеся от клеточной стенки пептидогликана и оставляющие место для периплазмы. Традиционно считалось, что цитоплазматическая мембрана отвечает за контроль осмотического давления бактериальных клеток. Тем не менее, из-за складок цитоплазматической мембраны и существования больших пространств периплазмы внутри Planctomycetota их пептидогликан действует как осмотический барьер, при этом периплазма изотонична цитозолю. [ 5 ]
Анаэробное окисление аммония (анаммокс)
[ редактировать ]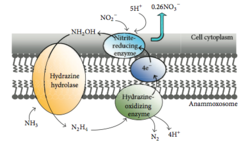
Анаммокс — это процесс окисления аммония, при котором нитрит выступает в качестве акцептора электронов. Этот процесс создает энергию для организма, выполняющего реакцию, точно так же, как люди получают энергию от окисления глюкозы. [ 46 ] В морской среде это в конечном итоге удаляет азот из воды, поскольку газ N 2 не может использоваться фитопланктоном и выбрасывается в атмосферу . До 67% добычи динитрогенного газа в океане приходится на анаммокс. [ 47 ] Считается, что около 50% газообразного азота в атмосфере производится из анаммокса. [ 48 ] Planctomycetota являются наиболее доминирующим типом бактерий, способных осуществлять анаммокс, поэтому Planctomycetota, способные осуществлять анаммокс, играют важную роль в глобальном круговороте азота. [ 49 ]
Синтез стерола
[ редактировать ]Синтез стеринов, часто наблюдаемый у эукариот и редко встречающийся у бактерий, у Planctomycetota наблюдался очень редко. [ 5 ] [ 15 ] Синтез стеринов, таких как ланостерин, наблюдался у G. obscuriglobus . Ланостерин распространен у эукариот и двух других групп бактерий, как метилотрофных Pseudomonadota , так и миксобактерий . Синтез стеринов, наблюдаемый у G. obscuriglobus, уникален для Planctomycetota. Предполагается, что синтез стеринов связан с регуляцией текучести мембран у Planctomycetota. [ 15 ] и был описан как необходимый для правильного роста и размножения G. obscuriglobus . [ 17 ]
Биотехнология и применение на людях
[ редактировать ]В последнее время возник интерес к изучению Planctomycetota на предмет их потенциальной роли в биотехнологии, главным образом как источника биоактивных молекул. [ 8 ] [ 14 ] представляют интерес главным образом для фармацевтической промышленности. Биологически активные соединения присутствуют в основном в виде вторичных метаболитов. [ 14 ] хотя мало что известно о вторичных метаболитах Planctomycetota. [ 50 ] Это неожиданно, поскольку Planctomycetota имеют несколько ключевых особенностей, присущих другим известным производителям биоактивных молекул, таким как миксобактерии. [ 50 ] Тем не менее, ряд текущих исследований служат различными первыми шагами по включению Planctomycetota в разработку низкомолекулярных лекарств для человека.
Виды Planctomycetota заслуживают внимания при оспаривании существующих моделей происхождения ядра, а также других аспектов происхождения и эволюции эукариотической эндомембранной системы. [ 41 ]
Изменение климата
[ редактировать ]Влияние исследований Planctomycetota и их использования может иметь глобальное значение в отношении процессов круговорота питательных веществ и способствовать дальнейшему пониманию глобальной морской биогеохимии. Однако, учитывая растущее влияние Planctomycetota на метаболические процессы, связанные с водой и воздухом, она также может играть роль в обмене между океанами и атмосферой, потенциально влияя на изменение климата. [ 41 ]
Планктомицетоты как возбудители
[ редактировать ]Виды Planctomycetota недавно были идентифицированы как условно-патогенные микроорганизмы человека. [ нужна ссылка ] но нехватка питательных сред ограничивает исследования бактерий Planctomycetota как патогенов человека. [ 8 ]
Ссылки
[ редактировать ]- ^ Орен А., генеральный менеджер Гаррити (2021 г.). «Действительная публикация названий сорока двух типов прокариот» . Int J Syst Evol Microbiol . 71 (10): 5056. doi : 10.1099/ijsem.0.005056 . ПМИД 34694987 . S2CID 239887308 .
- ^ Эзеби Ж.П., Парте AC. "" Планктомицеты "" . Список названий прокариот, имеющих номенклатуру (LPSN) . Проверено 26 августа 2021 г.
- ^ Сэйерс; и др. «Планктомицеты» . База данных таксономии Национального центра биотехнологической информации (NCBI) . Проверено 20 марта 2016 г.
- ^ Гэррити, Холт Дж.Г. (2001). «Дорожная карта к руководству». В составе Бун Д.Р., Кастенхольц Р.В., Гаррити ГМ. (ред.). Руководство Берджи по систематической бактериологии . Том. 1 ( Археи и глубоко ветвящиеся и фототрофные бактерии ) (2-е изд.). Нью-Йорк, штат Нью-Йорк: Springer-Verlag. стр. 119–166.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и Виганд С., Джоглер М., Джоглер С. (ноябрь 2018 г.). «Об индивидуальных планктомицетах» . Обзоры микробиологии FEMS . 42 (6): 739–760. дои : 10.1093/femsre/fuy029 . ПМИД 30052954 .
- ^ Jump up to: а б с д и ж г час Глёкнер Ф.О., Кубе М., Бауэр М., Тилинг Х., Ломбардот Т., Людвиг В. и др. (июль 2003 г.). «Полная последовательность генома морского планктомицета Pirellula sp., штамм 1» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (14): 8298–303. Бибкод : 2003PNAS..100.8298G . дои : 10.1073/pnas.1431443100 . ПМК 166223 . ПМИД 12835416 .
- ^ Jump up to: а б с д и Боерсма А.С., Каллшойер Н., Виганд С., Раст П., Петерс Ш., Месман Р.Дж. и др. (декабрь 2020 г.). «Alienimonas Californiensis gen. nov. sp. nov., новый планктомицет, выделенный из леса водорослей в заливе Монтерей». Антоний Левенгук . 113 (12): 1751–1766. дои : 10.1007/s10482-019-01367-4 . hdl : 10033/623182 . ПМИД 31802338 . S2CID 208641991 .
- ^ Jump up to: а б с д и ж г час я дж к л Каборе О.Д., Годрей С., Дранкур М. (2020). «Планктомицеты как бактерии, ассоциированные с хозяином: многообещающая перспектива для их будущей изоляции путем имитации их естественных экологических ниш в лабораториях клинической микробиологии» . Границы клеточной и инфекционной микробиологии . 10 : 519301. дои : 10.3389/fcimb.2020.519301 . ПМЦ 7734314 . ПМИД 33330115 .
- ^ Jump up to: а б с д и ж Вагнер, Майкл; Хорн, Матиас (01 июня 2006 г.). «Планктомицеты, Verrucomicrobia, Chlamydiae и сестринские типы составляют супертип, имеющий биотехнологическое и медицинское значение» . Современное мнение в области биотехнологии . Экологическая биотехнология/Энергетическая биотехнология. 17 (3): 241–249. doi : 10.1016/j.copbio.2006.05.005 . ISSN 0958-1669 . ПМИД 16704931 .
- ^ Jump up to: а б с д Кон, Тимо; Раст, Патрик; Кальшойер, Николай; Виганд, Сандра; Бедекер, Кристиан; Джеттен, Майк С.М.; Йеске, Ольга; Воллмерс, Джон; Кастер, Анн-Кристин; Роде, Манфред; Джоглер, Марейке (2020). «В микробиоме листьев морских водорослей Posidonia Oceanica могут доминировать планктомицеты» . Границы микробиологии . 11 : 1458. дои : 10.3389/fmicb.2020.01458 . ISSN 1664-302X . ПМЦ 7366357 . ПМИД 32754127 .
- ^ Jump up to: а б с д Лаге О.М., Бондосо Дж., Волк из клина А (октябрь 2013 г.). «Взгляд на ультраструктурную морфологию новых планктомицетов». Антони ван Левенгук . 104 (4): 467–76. дои : 10.1007/s10482-013-9969-2 . ПМИД 23857394 . S2CID 17003623 .
- ^ Jump up to: а б с д и Кумар Д., Кумар Г., Джагадишвари У., Сасикала С., Рамана К.В. (апрель 2021 г.). «» Candidatus Laterigemmans baculatus» рода nov. sp. nov., первый представитель палочковидных планктомицетов с боковым почкованием в семействе Pirellulaceae». Систематическая и прикладная микробиология . 44 (2): 126188. doi : 10.1016/j.syapm.2021.126188 . ПМИД 33647766 . S2CID 232091472 .
- ^ Jump up to: а б с д и ж г час Виганд С., Джоглер М., Бедекер С., Пинто Д., Фоллмерс Дж., Ривас-Марин Э. и др. (январь 2020 г.). «Культивирование и функциональная характеристика 79 планктомицетов раскрывают их уникальную биологию» . Природная микробиология . 5 (1): 126–140. дои : 10.1038/s41564-019-0588-1 . ПМЦ 7286433 . ПМИД 31740763 .
- ^ Jump up to: а б с Граса А.П., Калисто Р., Лаге ОМ (2016). «Планктомицеты как новый источник биоактивных молекул» . Границы микробиологии . 7 : 1241. дои : 10.3389/fmicb.2016.01241 . ПМЦ 4982196 . ПМИД 27570520 .
- ^ Jump up to: а б с д и ж г час Ривас-Марин Э., Петерс Ш., Кларет Фернандес Л., Джоглер С., ван Нифтрик Л., Виганд С., Девос Д.П. (январь 2020 г.). «Несущественность генов канонического клеточного деления у планктомицета Planctopirus limnophila» . Научные отчеты . 10 (1): 66. Бибкод : 2020НацСР..10...66Р . дои : 10.1038/s41598-019-56978-8 . ПМЦ 6952346 . ПМИД 31919386 .
- ^ Jump up to: а б с д и Сантарелла-Меллвиг Р., Пругналлер С., Роос Н., Маттай И.В., Девос Д.П. (21 мая 2013 г.). «Трехмерная реконструкция бактерий со сложной эндомембранной системой» . ПЛОС Биология . 11 (5): e1001565. дои : 10.1371/journal.pbio.1001565 . ПМК 3660258 . ПМИД 23700385 .
- ^ Jump up to: а б Ривас-Марин, Елена; Стеттнер, Шон; Готтшалл, Екатерина Ю.; Сантана-Милл, Чарльз; Хеллинг, Митч; Бэзил, Франко; Уорд, Наоми Л.; Девос, Дэмиен П. (2 июля 2019 г.). «Необходимость генов синтеза стеринов у планктомицетной бактерии Gemmata obscuriglobus » . Природные коммуникации . 10 (1): Бибкод : 2019NatCo..10.2916R. 2916. дои : 10.1038/s41467-019-10983-7 . ISSN 2041-1723 . ПМК 6606645 . ПМИД 31266954 .
- ^ Jump up to: а б с д и ж г час я дж Бодекер С., Шулер М., Рейнтьес Г., Йеске О., ван Тиселинг М.К., Джоглер М. и др. (апрель 2017 г.). «Определение биологии бактериальных клеток планктомицетов» . Природные коммуникации . 8 (1): 14853. Бибкод : 2017NatCo...814853B . дои : 10.1038/ncomms14853 . ПМЦ 5394234 . ПМИД 28393831 .
- ^ Каллшойер Н., Морейра К., Эйрс Р., Ллевеллин К.А., Виганд С., Джоглер К., Лаге О.М. (декабрь 2019 г.). «Планктомицеты с розовым и оранжевым пигментом производят каротиноиды типа сапроксантина, включая редкий каротиноид C 45 » (PDF) . Отчеты по экологической микробиологии . 11 (6): 741–748. дои : 10.1111/1758-2229.12796 . ПМИД 31600855 . S2CID 204244301 .
- ^ Jump up to: а б Виана Ф, Лаге ОМ, Оливейра Р (октябрь 2013 г.). «Высокая устойчивость морских планктомицетов к ультрафиолету C». Антони ван Левенгук . 104 (4): 585–95. дои : 10.1007/s10482-013-0027-x . hdl : 1822/50752 . ПМИД 24052365 . S2CID 13153498 .
- ^ Сантана-Молина К., Энрикес В., Хорнеро-Мендес Д., Девос Д.П., Ривас-Марин Э. (декабрь 2022 г.). «Путь сквалена к биосинтезу каротиноидов C30 и происхождение путей биосинтеза каротиноидов» . Proc Natl Acad Sci США . 119 (52): e2210081119. дои : 10.1073/pnas.2210081119 . ПМК 9907078 . ПМИД 36534808 . S2CID 254909480 .
- ^ Jump up to: а б Кон Т., Хойер А., Джоглер М., Фоллмерс Дж., Бодекер С., Банк Б. и др. (2016). « Fuerstia marisgermanicae gen. nov., sp. nov., необычный представитель типа Planctomycetes из немецкого Ваттового моря» . Границы микробиологии . 7 : 2079. дои : 10.3389/fmicb.2016.02079 . ПМК 5177795 . ПМИД 28066393 .
- ^ Jump up to: а б Картал, Боран; де Алмейда, Наоми М.; Маальке, Воутер Дж.; Оп ден Кэмп, Хууб Дж.М.; Джеттен, Майк С.М.; Келтьенс, Ян Т. (1 мая 2013 г.). «Как зарабатывать на жизнь анаэробным окислением аммония» . Обзоры микробиологии FEMS . 37 (3): 428–461. дои : 10.1111/1574-6976.12014 . hdl : 2066/103425 . ISSN 0168-6445 . ПМИД 23210799 .
- ^ Jump up to: а б с Петерс Ш., Виганд С., Каллшойер Н., Джоглер М., Хойер А., Джеттен М.С. и др. (декабрь 2020 г.). «Три морских штамма составляют новый род и вид Crateriforma conspicua из типа Planctomycetes». Антони ван Левенгук . 113 (12): 1797–1809. дои : 10.1007/s10482-019-01375-4 . hdl : 10033/623180 . ПМИД 31894495 . S2CID 209516837 .
- ^ Jump up to: а б Джоглер К., Глёкнер Ф.О., Колтер Р. (август 2011 г.). «Характеристика Planctomyces limnophilus и разработка генетических инструментов для манипулирования им делают его модельным видом для типа Planctomycetes» . Прикладная и экологическая микробиология . 77 (16): 5826–9. Бибкод : 2011ApEnM..77.5826J . дои : 10.1128/АЕМ.05132-11 . ПМК 3165242 . ПМИД 21724885 .
- ^ Jump up to: а б с д и Ривас-Марин Э, Каноса I, Девос ДП (2016). «Эволюционная клеточная биология способа деления бактериальных планктомицетов - Verrucomicrobia - Chlamydiae Superphylum» . Границы микробиологии . 7 : 1964. doi : 10.3389/fmicb.2016.01964 . ПМК 5147048 . ПМИД 28018303 .
- ^ Jump up to: а б с Бернандер Р., Эттема Т.Дж. (декабрь 2010 г.). «Деление клеток без FtsZ у архей и бактерий». Современное мнение в микробиологии . Рост и развитие: эукариоты/прокариоты. 13 (6): 747–52. дои : 10.1016/j.mib.2010.10.005 . ПМИД 21050804 .
- ^ Jump up to: а б Гупта Р.С., Бхандари В., Наушад Х.С. (2012). «Молекулярные сигнатуры группы ПВХ (планктомицеты, веррукомикробии, хламидии и лентисферы) бактерий дают представление об их эволюционных взаимоотношениях» . Границы микробиологии . 3 : 327. дои : 10.3389/fmicb.2012.00327 . ПМЦ 3444138 . ПМИД 23060863 .
- ^ Jump up to: а б Гупта РС (июль 2016 г.). «Влияние геномики на понимание эволюции и классификации микробов: важность взглядов Дарвина на классификацию» . Обзоры микробиологии FEMS . 40 (4): 520–53. дои : 10.1093/femsre/fuw011 . ПМИД 27279642 .
- ^ Лагкувардос И., Джел М.А., Раттей Т., Хорн М. (январь 2014 г.). «Сигнатурный белок супертипа ПВХ» . Прикладная и экологическая микробиология . 80 (2): 440–5. Бибкод : 2014ApEnM..80..440L . дои : 10.1128/АЕМ.02655-13 . ПМЦ 3911108 . ПМИД 24185849 .
- ^ Jump up to: а б с Махаджан, Маянк; Да, Бенджамин; Хэгглунд, Эмиль; Гай, Лайонел; Фюрст, Джон А; Андерссон, Сив Г.Е. (11 декабря 2019 г.). «Паралогизация и новая белковая архитектура у бактерий-планктомицетов со сложной клеточной структурой» . Молекулярная биология и эволюция . 37 (4): 1020–1040. дои : 10.1093/molbev/msz287 . ISSN 0737-4038 . ПМИД 31808939 .
- ^ Строус М., Пеллетье Э., Манжено С. (6 апреля 2006 г.). «Расшифровка эволюции и метаболизма анаммокс-бактерии на основе генома сообщества». Природа . 440 (7085): 790–794. Бибкод : 2006Natur.440..790S . дои : 10.1038/nature04647 . hdl : 2066/35981 . ПМИД 16598256 . S2CID 4402553 .
- ^ «ЛТП» . Проверено 23 февраля 2021 г.
- ^ «Дерево LTP_all в формате Ньюика» . Проверено 23 февраля 2021 г.
- ^ «Примечания к выпуску LTP_12_2021» (PDF) . Проверено 23 февраля 2021 г.
- ^ «Выпуск GTDB 08-RS214» . База данных геномной таксономии . Проверено 10 мая 2023 г.
- ^ "bac120_r214.sp_label" . База данных геномной таксономии . Проверено 10 мая 2023 г.
- ^ «История таксонов» . База данных геномной таксономии . Проверено 10 мая 2023 г.
- ^ Jump up to: а б с Фюрст, Джон (2004). «Планктомицеты: тип возникающего интереса к микробной эволюции и экологии». Информационный бюллетень Всемирной федерации коллекций культуры . 38 . CiteSeerX 10.1.1.538.2883 .
- ^ Jump up to: а б Бакли, Дэниел Х.; Хуангьютитхам, Вариса; Нельсон, Тиррелл А.; Румбергер, Анжелика; Тис, Дженис Э. (1 июля 2006 г.). «Разнообразие планктомицетов в почве в связи с историей почвы и неоднородностью окружающей среды» . Прикладная и экологическая микробиология . 72 (7): 4522–4531. Бибкод : 2006ApEnM..72.4522B . дои : 10.1128/АЕМ.00149-06 . ISSN 0099-2240 . ПМЦ 1489350 . ПМИД 16820439 .
- ^ Jump up to: а б с Фюрст, Дж. А. (1 июля 1995 г.). «Планктомицеты: новые модели микробной экологии, эволюции и клеточной биологии» . Микробиология . 141 (7): 1493–1506. дои : 10.1099/13500872-141-7-1493 . ISSN 1350-0872 . ПМИД 7551018 .
- ^ Jump up to: а б Вёбкен Д., Тилинг Х., Векер П., Думитриу А., Костадинов И., Делонг Э.Ф. и др. (сентябрь 2007 г.). «Фосмиды новых морских планктомицетов из систем апвеллинга побережья Намибии и Орегона и их перекрестное сравнение с геномами планктомицетов» . Журнал ISME . 1 (5): 419–35. дои : 10.1038/ismej.2007.63 . ПМИД 18043661 . S2CID 23859244 .
- ^ Бондосо Дж., Балаге В., Газоль Ж.М., Лаге О.М. (июнь 2014 г.). «Состав сообщества Planctomycetota, связанного с различными макроводорослями» . ФЭМС Микробиология Экология . 88 (3): 445–56. дои : 10.1111/1574-6941.12258 . ПМИД 24266389 .
- ^ Jump up to: а б с Фюрст Дж. А. (октябрь 2005 г.). «Внутриклеточная компартментация планктомицетов». Ежегодный обзор микробиологии . 59 (1): 299–328. дои : 10.1146/annurev.micro.59.030804.121258 . ПМИД 15910279 .
- ^ Jump up to: а б Изуми Х., Сагуленко Е., Уэбб Р.И., Фюрст Дж.А. (октябрь 2013 г.). «Выделение и разнообразие планктомицетов из губки Niphates sp., морской воды и отложений залива Мортон, Австралия». Антони ван Левенгук . 104 (4): 533–46. дои : 10.1007/s10482-013-0003-5 . ПМИД 23959164 . S2CID 12315225 .
- ^ Картал, Боран (2011). Культивирование, обнаружение и экофизиология анаэробных аммонийокисляющих бактерий . Сан-Диего, Калифорния: Академическая пресса. стр. 89–108. ISBN 978-0-12-381294-0 .
- ^ Цянь; и др. (2018). «Разнообразие и распространение анаммокс-бактерий в толще воды и отложениях восточной части Индийского океана». Международная биопорча и биодеградация . 133 : 52–62. дои : 10.1016/j.ibiod.2018.05.015 . S2CID 90491277 .
- ^ Тислинг; и др. (2015). «Анаммокс-планктомицеты имеют пептидогликановую клеточную стенку» . Природные коммуникации . 6 : 6878. Бибкод : 2015NatCo...6.6878V . дои : 10.1038/ncomms7878 . ПМЦ 4432595 . ПМИД 25962786 .
- ^ Цзин; и др. (2015). «Снимок пространственного и вертикального распределения бактериальных сообществ в восточной части Индийского океана». Acta Oceanologica Sinica . 35 (6): 85–93. дои : 10.1007/s13131-016-0871-4 . S2CID 89295982 .
- ^ Jump up to: а б Йеске, Ольга; Джоглер, Марейке; Петерсен, Йорн; Сикорский, Йоханнес; Джоглер, Кристиан (1 октября 2013 г.). «От геномного анализа к фенотипическим микрочипам: планктомицеты как источник новых биоактивных молекул» . Антони ван Левенгук . 104 (4): 551–567. дои : 10.1007/s10482-013-0007-1 . ISSN 1572-9699 . ПМИД 23982431 . S2CID 18752686 .
Внешние ссылки
[ редактировать ] Данные, относящиеся к Planctomycetota , в Wikispecies
Данные, относящиеся к Planctomycetota , в Wikispecies - Помимо прокариотов и эукариотов: планктомицеты и клеточная организация.
- Гемата в Microbewiki