МХК I класса
| МХК I класса | |
|---|---|
 Схематическое изображение MHC класса I | |
| Идентификаторы | |
| Символ | МХК I класса |
| Мембраном | 63 |
Молекулы MHC класса I являются одним из двух основных классов молекул главного комплекса гистосовместимости (MHC) (второй — MHC класса II ) и обнаруживаются на клеточной поверхности всех ядросодержащих клеток в телах позвоночных . [ 1 ] [ 2 ] Они также встречаются на тромбоцитах , но не на эритроцитах . Их функция состоит в том, чтобы отображать пептидные фрагменты белков изнутри клетки цитотоксическим Т-клеткам ; это вызовет немедленный ответ иммунной системы против определенного чужеродного антигена, отображаемого с помощью белка MHC класса I. Поскольку молекулы MHC класса I представляют собой пептиды, полученные из цитозольных белков, путь презентации MHC класса I часто называют цитозольным или эндогенным путем . [ 3 ]
У людей HLA , соответствующие классу I MHC, представляют собой HLA-A , HLA-B и HLA-C .
Функция
[ редактировать ]Молекулы MHC класса I связывают пептиды, образующиеся в основном в результате деградации цитозольных белков протеасомой . Затем комплекс MHC I:пептид встраивается через эндоплазматический ретикулум во внешнюю плазматическую мембрану клетки. Эпитопный пептид связан с внеклеточными частями молекулы MHC класса I. Таким образом, функция MHC класса I заключается в отображении внутриклеточных белков цитотоксическим Т-клеткам (CTL). Однако MHC класса I может также представлять пептиды, полученные из экзогенных белков, в процессе, известном как перекрестная презентация .
Нормальная клетка будет отображать пептиды нормального клеточного белкового обмена на своем MHC класса I, и CTL не будут активироваться в ответ на них из-за механизмов центральной и периферической толерантности. Когда клетка экспрессирует чужеродные белки, например, после вирусной инфекции, часть MHC класса I будет отображать эти пептиды на поверхности клетки. Следовательно, CTL, специфичные для комплекса MHC:пептид, будут распознавать и убивать представленные клетки.
Альтернативно, сам MHC класса I может служить ингибирующим лигандом для естественных клеток-киллеров (NK). Снижение нормального уровня MHC поверхностного класса I - механизм, используемый некоторыми вирусами. [ 4 ] и некоторые опухоли, уклоняющиеся от реакции CTL, активируют уничтожение NK-клеток.
ПирБ и визуальная пластичность
[ редактировать ]Парный иммуноглобулиноподобный рецептор B (PirB), MHCI-связывающий рецептор, участвует в регуляции зрительной пластичности . [ 5 ] PirB экспрессируется в центральной нервной системе и снижает глазного доминирования пластичность развития в критический период и во взрослом возрасте. [ 5 ] Когда функция PirB была отменена у мутантных мышей, глазного доминирования пластичность стала более выраженной во всех возрастах. [ 5 ] Мутантные мыши с потерей функции PirB также демонстрировали повышенную пластичность после монокулярной депривации в течение критического периода . [ 5 ] Эти результаты позволяют предположить, что PirB может участвовать в модуляции синаптической пластичности в зрительной коре .
Структура
[ редактировать ]Молекулы MHC класса I представляют собой гетеродимеры, состоящие из двух полипептидных цепей: α и β 2 -микроглобулина (В2М). Две цепи связаны нековалентно посредством взаимодействия B2M и домена α3 . Только α-цепь полиморфна и кодируется геном HLA , тогда как субъединица B2M не является полиморфной и кодируется геном микроглобулина бета-2 . Домен α3 - охватывает плазматическую мембрану и взаимодействует с корецептором CD8 Т клеток . Взаимодействие α 3 -CD8 удерживает молекулу MHC I на месте, в то время как рецептор Т-клеток (TCR) на поверхности цитотоксической Т-клетки связывает ее гетеродимерный лиганд α 1 -α 2 и проверяет связанный пептид на антигенность. Домены α1 складываются и α2 , образуя бороздку для связывания пептидов. Молекулы MHC класса I связывают пептиды, длина которых преимущественно состоит из 8-10 аминокислот (Parham 87), но также сообщалось о связывании более длинных пептидов. [ 6 ]
между пептидом, MHC I и B2M обычно необходимы пептид с высоким сродством и субъединица B2M Хотя для поддержания стабильного тройного комплекса MHC I/B2M с дефицитом пептида . стабильные гетеродимеры , при субфизиологических температурах наблюдались [ 7 ] [ 8 ] Синтетические стабильные, пептид-рецепторные молекулы MHC I были созданы с использованием дисульфидной связи между MHC I и B2M, получившей название «открытый MHC-I». [ 9 ]
Синтез
[ редактировать ]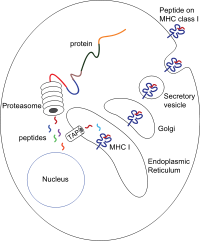
Пептиды генерируются в основном цитозоле протеасомой в . Протеасома – макромолекула, состоящая из 28 субъединиц, половина из которых оказывает протеолитическую активность. Протеасома расщепляет внутриклеточные белки на небольшие пептиды, которые затем высвобождаются в цитозоль. Протеасомы также могут лигировать отдельные пептидные фрагменты (так называемые сплайсированные пептиды), создавая несмежные последовательности и, следовательно, не образующие линейного шаблона в геноме. Происхождение сплайсированных пептидных сегментов может происходить из одного и того же белка (цис-сплайсинг) или из разных белков (транс-сплайсинг). [ 10 ] [ 11 ] Пептиды должны быть транслоцированы из цитозоля в эндоплазматический ретикулум (ЭР), чтобы встретиться с молекулой MHC класса I, чей сайт связывания пептидов находится в просвете ЭР . У них есть проксимальная к мембране Ig-складка.
Транслокация и загрузка пептидов
[ редактировать ]Транслокация пептида из цитозоля в просвет ЭР осуществляется транспортером, связанным с процессингом антигена (TAP). TAP является членом семейства транспортеров ABC и представляет собой гетеродимерный мультимембранный полипептид, состоящий из TAP1 и TAP2 . Две субъединицы образуют сайт связывания пептида и два сайта связывания АТФ, обращенные к цитозолю. TAP связывает пептиды на цитоплазматической стороне и транслоцирует их при потреблении АТФ в просвет ЭР. Молекула MHC класса I затем, в свою очередь, нагружается пептидами в просвете ЭР.
В процессе загрузки пептидов участвуют несколько других молекул, которые образуют большой мультимерный комплекс, называемый комплексом загрузки пептидов. [ 12 ] состоит из TAP, тапасина , кальретикулина , кальнексина и Erp57 ( PDIA3 ). Кальнексин стабилизирует α-цепи MHC класса I до связывания β2m. После полной сборки молекулы MHC кальнексин диссоциирует. Молекула MHC, лишенная связанного пептида, по своей природе нестабильна и требует связывания шаперонов кальретикулина и Erp57. Кроме того, тапасин связывается с молекулой MHC и служит для связывания ее с белками TAP и облегчает выбор пептида в итеративном процессе, называемом редактированием пептида. [ 13 ] [ 14 ] [ 15 ] тем самым способствуя усиленной загрузке пептидов и колокализации.
Как только пептид загружается в молекулу MHC класса I, комплекс диссоциирует и покидает ЭР секреторным путем , достигая поверхности клетки. Транспорт молекул MHC класса I по секреторному пути включает несколько посттрансляционных модификаций молекулы MHC. Некоторые из посттрансляционных модификаций происходят в ЭР и включают изменения в N-гликановых областях белка с последующими обширными изменениями в N-гликанах аппарата Гольджи . N-гликаны полностью созревают еще до того, как достигают поверхности клетки.
Удаление пептидов
[ редактировать ]Пептиды, которые не способны связывать молекулы MHC класса I в просвете эндоплазматического ретикулума (ЭР), удаляются из ЭР через канал sec61 в цитозоль, [ 16 ] [ 17 ] где они могут подвергнуться дальнейшему уменьшению размера и могут быть транслоцированы с помощью TAP обратно в ER для связывания с молекулой MHC класса I.
взаимодействие sec61 с бычьим альбумином . Например, наблюдалось [ 18 ]
Влияние вирусов
[ редактировать ]Молекулы MHC класса I нагружены пептидами, образующимися в результате деградации убиквитинированных цитозольных белков в протеасомах . Поскольку вирусы индуцируют клеточную экспрессию вирусных белков, некоторые из этих продуктов подвергаются деградации, в результате чего полученные пептидные фрагменты попадают в эндоплазматический ретикулум и связываются с молекулами MHC I. Таким образом, MHC-зависимый путь презентации антигена класса I заключается в том, что инфицированные вирусом клетки сигнализируют Т-клеткам о том, что в результате инфекции вырабатываются аномальные белки.
Судьба инфицированной вирусом клетки почти всегда заключается в индукции апоптоза посредством клеточного иммунитета , снижающем риск заражения соседних клеток. В качестве эволюционного ответа на этот метод иммунного надзора многие вирусы способны подавлять или иным образом предотвращать презентацию молекул MHC класса I на поверхности клеток. В отличие от цитотоксических Т-лимфоцитов, естественные клетки-киллеры (NK) обычно инактивируются после узнавания молекул MHC I на поверхности клеток. Следовательно, в отсутствие молекул MHC I NK-клетки активируются и распознают клетку как аберрантную, что позволяет предположить, что она может быть инфицирована вирусами, пытающимися избежать иммунного разрушения. Некоторые виды рака человека также демонстрируют снижение уровня MHC I, что дает трансформированным клеткам такое же преимущество выживания, что и возможность избежать обычного иммунного надзора, направленного на уничтожение любых инфицированных или трансформированных клеток. [ 19 ]
Гены и изотипы
[ редактировать ]- Очень полиморфный
- Менее полиморфный
Эволюционная история
[ редактировать ]Гены MHC класса I произошли от самого недавнего общего предка всех челюстных позвоночных и были обнаружены у всех ныне живущих челюстных позвоночных, которые были изучены до сих пор. [ 2 ] С момента своего появления у челюстных позвоночных это семейство генов претерпело множество различных эволюционных путей по мере того, как видообразования происходили события . Однако зарегистрированы случаи межвидового полиморфизма в генах MHC класса I, когда определенный аллель в эволюционно родственном гене MHC класса I остается у двух видов, вероятно, из-за сильного патоген-опосредованного балансирующего отбора со стороны патогенов , которые могут инфицировать оба вида. разновидность. [ 20 ] Эволюция рождения и смерти является одним из механистических объяснений размера семейства генов MHC класса I.
Рождение и смерть генов MHC класса I
[ редактировать ]Эволюция рождения и смерти утверждает, что события дупликации генов приводят к тому, что геном содержит несколько копий гена, которые затем могут подвергаться отдельным эволюционным процессам. Иногда эти процессы приводят к псевдогенизации (гибель) одной копии гена, хотя иногда этот процесс приводит к образованию двух новых генов с дивергентными функциями. [ 21 ] Вполне вероятно, что локусы MHC класса Ib человека (HLA-E, -F и -G), а также псевдогены MHC класса I возникли из локусов MHC класса Ia (HLA-A, -B и -C) при этом рождении. и – процесс смерти. [ 22 ]
Ссылки
[ редактировать ]- ^ Хьюитт EW (октябрь 2003 г.). «Путь презентации антигена MHC класса I: стратегии уклонения от вирусного иммунитета» . Иммунология . 110 (2): 163–9. дои : 10.1046/j.1365-2567.2003.01738.x . ПМК 1783040 . ПМИД 14511229 .
- ^ Jump up to: а б Кульски Дж.К., Шиина Т., Анзай Т., Кохара С., Иноко Х. (декабрь 2002 г.). «Сравнительный геномный анализ MHC: эволюция блоков дупликации класса I, разнообразие и сложность от акулы к человеку». Иммунологические обзоры . 190 : 95–122. дои : 10.1034/j.1600-065x.2002.19008.x . ПМИД 12493009 . S2CID 41765680 .
- ^ http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/H/HLA.html#Class_I_Histocompatibility_Molecules . Архивировано 4 февраля 2016 г. на страницах биологии Wayback Machine Кимбалла, Молекулы гистосовместимости.
- ^ Хансен Т.Х., Бувье М. (июль 2009 г.). «Презентация антигена MHC класса I: изучение стратегий уклонения от вирусов». Обзоры природы. Иммунология . 9 (7): 503–13. дои : 10.1038/nri2575 . ПМИД 19498380 . S2CID 9278263 .
- ^ Jump up to: а б с д Сайкен Дж., Грандпре Т., Канольд П.О., Шац С.Дж. (сентябрь 2006 г.). «PirB ограничивает пластичность глазного доминирования в зрительной коре». Наука . 313 (5794): 1795–800. Бибкод : 2006Sci...313.1795S . дои : 10.1126/science.1128232 . ПМИД 16917027 . S2CID 1860730 .
- ^ Берроуз С.Р., Россджон Дж., МакКласки Дж. (январь 2006 г.). «Не слишком ли мы ограничились картированием эпитопов CTL?». Тенденции в иммунологии . 27 (1): 11–6. дои : 10.1016/j.it.2005.11.001 . ПМИД 16297661 .
- ^ Юнггрен Х.Г., Стам Н.Ю., Олен С., Нефьес Дж.Дж., Хёглунд П., Хемелс М.Т., Бастин Дж., Шумахер Т.Н., Таунсенд А., Карре К., Плоег Х.Л. (02.08.1990). «Пустые молекулы MHC класса I выходят на холод» . Природа . 346 (6283): 476–480. Бибкод : 1990Natur.346..476L . дои : 10.1038/346476a0 . ISSN 0028-0836 . ПМИД 2198471 .
- ^ Шумахер Т.Н., Хемелс М.Т., Нифьес Дж.Дж., Каст В., Мелиф С.Дж., Плух Х.Л. (август 1990 г.). «Прямое связывание пептида с пустыми молекулами MHC класса I на интактных клетках и in vitro» . Клетка . 62 (3): 563–567. дои : 10.1016/0092-8674(90)90020-F . ПМИД 2199065 .
- ^ Сан Ю, Янг MC, Вудворд CH, Данон Дж. Н., Труонг Х. В., Гупта С., Уинтерс Т. Дж., Фонт-Бургада Дж., Бурслем Г. М., Сгуракис Н. Г. (20 июня 2023 г.). «Универсальные открытые молекулы MHC-I для быстрой загрузки пептидов и повышения комплексной стабильности во всех аллотипах HLA» . Труды Национальной академии наук . 120 (25): e2304055120. Бибкод : 2023PNAS..12004055S . дои : 10.1073/pnas.2304055120 . ISSN 0027-8424 . ПМЦ 10288639 . ПМИД 37310998 .
- ^ Фариди П., Ли С., Рамаратинам С.Х., Вивиан Дж.П., Иллинг П.Т., Мифсуд Н.А., Аяла Р., Сонг Дж., Гиринг Л.Дж., Герцог П.Дж., Тернетт Н., Россджон Дж., Крофт Н.П., Перселл А.В. (12 октября 2018 г.). «Подмножество пептидов HLA-I не имеет геномной матрицы: данные о цис- и транс-сплайсированных пептидных лигандах» (PDF) . Наука Иммунология . 3 (28): eaar3947. doi : 10.1126/sciimmunol.aar3947 . ПМИД 30315122 .
- ^ Липе Дж., Марино Ф., Сидни Дж., Джеко А., Бантинг Д.Е., Сетте А., Клёцель П.М., Штумпф М.П., Хек А.Дж., Мишто М. (21 октября 2016 г.). «Большая часть лигандов HLA класса I представляет собой сплайсированные пептиды, генерируемые протеасомами» (PDF) . Наука . 354 (6310): 354–358. Бибкод : 2016Sci...354..354L . doi : 10.1126/science.aaf4384 . hdl : 10044/1/42330 . ПМИД 27846572 . S2CID 41095551 .
- ^ Блис А., Янулиен Д., Хофманн Т., Коллер Н., Шмидт С., Тровитч С., Мёллер А., Тампе Р. (ноябрь 2017 г.). «Структура человеческого комплекса загрузки пептида MHC-I». Природа . 551 (7681): 525–528. Бибкод : 2017Natur.551..525B . дои : 10.1038/nature24627 . ПМИД 29107940 . S2CID 4447406 .
- ^ Ховарт М., Уильямс А., Толструп А.Б., Эллиот Т. (август 2004 г.). «Тапасин усиливает презентацию пептида MHC класса I в зависимости от периода полураспада пептида» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (32): 11737–42. Бибкод : 2004PNAS..10111737H . дои : 10.1073/pnas.0306294101 . ПМК 511045 . ПМИД 15286279 .
- ^ Уирш, Пенсильвания, Крессвелл П. (август 2007 г.). «Селективная загрузка высокоаффинных пептидов на молекулы главного комплекса гистосовместимости класса I с помощью гетеродимера тапасин-ERp57». Природная иммунология . 8 (8): 873–81. дои : 10.1038/ni1485 . ПМИД 17603487 . S2CID 29762957 .
- ^ Третьеборо С.М., Роддик Дж.С., Рэдклифф Дж.Н., Ховарт М., Стивенсон Ф.К., Эллиотт Т. (февраль 2008 г.). «Тапасин формирует иерархию иммунодоминирования в зависимости от кинетической стабильности комплексов пептид-MHC класса I» . Европейский журнал иммунологии . 38 (2): 364–9. дои : 10.1002/eji.200737832 . ПМИД 18196518 . S2CID 28659293 .
- ^ Купманн Дж., Альбринг Дж., Хютер Э., Бульбук Н., Спи П., Нефьес Дж., Хеммерлинг Г.Дж., Момбург Ф. и др. (июль 2000 г.). «Экспорт антигенных пептидов из эндоплазматического ретикулума пересекается с ретроградной транслокацией белка через канал Sec61p» . Иммунитет . 13 (1): 117–27. дои : 10.1016/S1074-7613(00)00013-3 . ПМИД 10933400 .
- ^ Альбринг Дж., Купманн Дж.О., Хеммерлинг Г.Дж., Момбург Ф. (январь 2004 г.). «Ретротранслокация тяжелой цепи MHC класса I из эндоплазматического ретикулума в цитозоль зависит от доставки АТФ в просвет ЭР». Молекулярная иммунология . 40 (10): 733–41. дои : 10.1016/j.molimm.2003.08.008 . ПМИД 14644099 .
- ^ Имаи Дж., Хасегава Х., Маруя М., Коясу С., Яхара I (январь 2005 г.). «Экзогенные антигены обрабатываются посредством деградации, связанной с эндоплазматическим ретикулумом (ERAD), при перекрестной презентации дендритными клетками» . Международная иммунология . 17 (1): 45–53. дои : 10.1093/intimm/dxh184 . ПМИД 15546887 .
- ^ Ван З, Чжан Л., Цяо А., Уотсон К., Чжан Дж., Фань Г.Х. (февраль 2008 г.). «Активация CXCR4 вызывает убиквитинирование и подавление главного комплекса гистосовместимости класса I (MHC-I) на клетках эпителиоидной карциномы HeLa» . Журнал биологической химии . 283 (7): 3951–9. дои : 10.1074/jbc.m706848200 . ПМИД 18083706 .
- ^ Азеведо Л., Серрано С., Аморим А., Купер Д.Н. (сентябрь 2015 г.). «Межвидовой полиморфизм у людей и человекообразных обезьян обычно поддерживается путем балансирующего отбора, который модулирует иммунный ответ хозяина» . Геномика человека . 9 (1): 21. дои : 10.1186/s40246-015-0043-1 . ПМК 4559023 . ПМИД 26337052 .
- ^ Ней М, Руни А.П. (14 ноября 2005 г.). «Согласованная эволюция мультигенных семей по принципу рождения и смерти» . Ежегодный обзор генетики . 39 (1): 121–52. дои : 10.1146/annurev.genet.39.073003.112240 . ПМЦ 1464479 . ПМИД 16285855 .
- ^ Хьюз А.Л. (март 1995 г.). «Происхождение и эволюция псевдогенов HLA класса I» . Молекулярная биология и эволюция . 12 (2): 247–58. doi : 10.1093/oxfordjournals.molbev.a040201 . ПМИД 7700152 .
Внешние ссылки
[ редактировать ]- Гистосовместимость + Антигены + Класс + I в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- MHC + Класс + I + Гены в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)