Нокаутирующая крыса

Нокаутная крыса – это генетически модифицированная крыса , у которой единственный ген отключен посредством целевой мутации ( ловушки гена ), используемая для академических и фармацевтических исследований . Нокаутные крысы могут имитировать человеческие заболевания и являются важным инструментом для изучения функции генов ( функциональная геномика ), а также для открытия и разработки лекарств . Производство нокаутных крыс было экономически и технически нецелесообразно до 2008 года. [1] [2] [3] [4]
Технология разработана благодаря финансированию Национальных институтов здравоохранения (NIH) и работе, проделанной членами Консорциум Knock Out Rat Consortium (KORC) привел к разработке экономически эффективных методов создания нокаутных крыс. О важности развития крыс как более универсального инструмента для исследований в области здоровья человека свидетельствуют инвестиции в размере 120 миллионов долларов, сделанные НИЗ через Консорциум проекта секвенирования генома крысы , результатом которого стал проект последовательности лабораторного штамма коричневой или норвежской крысы ( Rattus norvegicus ). [5] Дополнительные разработки в области нуклеазы с цинковыми пальцами в 2009 году привели к появлению первой нокаутной крысы с целевыми мутациями, передаваемыми по зародышевой линии. [6] нокаутные модели крысиных болезней Паркинсона , Альцгеймера , гипертонии и диабета с использованием технологии нуклеазы с цинковыми пальцами. Лаборатории SAGE Labs коммерциализируют [7] [8]
Исследовательское использование
[ редактировать ]Мыши, крысы и люди имеют общие гены друг друга, кроме примерно 1%. [5] [9] [10] сделать грызунов хорошими модельными организмами для изучения функции генов человека. И мыши, и крысы относительно небольшие, с ними легко обращаться, у них короткий период генерации и они генетически инбредны. Хотя мыши оказались полезной моделью грызунов и были разработаны методы рутинного разрушения их генов, во многих случаях крысы считаются лучшими лабораторными животными для изучения и моделирования заболеваний человека.
Крысы физиологически более похожи на человека, чем мыши. Например, у крыс частота сердечных сокращений больше похожа на человеческую, а у мышей частота сердечных сокращений в пять-десять раз выше. Широко распространено мнение, что крыса является лучшей моделью, чем мышь, для сердечно-сосудистых заболеваний человека , диабета, артрита и многих аутоиммунных , неврологических , поведенческих расстройств и расстройств, связанных с зависимостью. [11] Кроме того, модели на крысах превосходят модели на мышах при тестировании фармакодинамики и токсичности потенциальных терапевтических соединений, отчасти потому, что количество и тип многих из их детоксифицирующих ферментов очень похожи на таковые у человека. [12] Их больший размер делает крыс более удобными для изучения с помощью приборов, а также облегчает такие манипуляции, как забор крови, нервную проводимость и выполнение операций.
На мышах доступны методы генетических манипуляций, которые обычно используются для моделирования заболеваний человека. Хотя опубликованные нокауты существуют примерно для 60% [13] Из мышиных генов подавляющее большинство распространенных заболеваний человека не имеют нокаутной мышиной модели. Нокаутные крысиные модели являются альтернативой мышам, которая может позволить создавать новые нарушения генов, недоступные у мышей. Модели нокаутных крыс также могут дополнять существующие модели трансгенных мышей. Сравнение мутантов мышей и крыс может облегчить различие между специфичными для грызунов и общими млекопитающих фенотипами .
Производственные проблемы
[ редактировать ]Крысиные модели использовались для продвижения многих областей медицинских исследований, включая сердечно-сосудистые заболевания, психические расстройства (исследования поведенческого вмешательства и зависимости), нервную регенерацию , диабет, трансплантацию , аутоиммунные заболевания ( ревматоидный артрит ), рак и заживление ран и костей. Хотя завершение последовательности генома крысы дает очень важную информацию, то, как эти заболевания связаны с функцией гена, требует эффективного метода создания нокаутных моделей крыс, в которых манипулируют конкретными геномными последовательностями.Большинство методов генетических манипуляций, включая случайный мутагенез с генной ловушкой (на основе ретровирусов и не на основе ретровирусов), нокауты/нокины генов и условные мутации, зависят от культуры и манипуляций с эмбриональным стволом (ES). клетки. [14] Крысиные ЭС клетки были выделены совсем недавно, и о каких-либо проявлениях генной модификации в них не сообщалось. Следовательно, многие методы генетических манипуляций, широко используемые на мышах, невозможны на крысах.
Ранние методы
[ редактировать ]До коммерческого развития технологии мобильной ДНК в 2007 году и технологии нуклеазы с цинковыми пальцами в 2009 году существовало только две технологии, которые можно было использовать для создания крысиных моделей заболеваний человека: клонирование и химический мутагенез с использованием N-этил-N-нитрозомочевины ( ЕНУ). ). Хотя клонирование путем переноса ядра соматических клеток (SCNT) теоретически можно использовать для создания крыс со специфическими мутациями путем мутирования соматических клеток и последующего использования этих клеток для SCNT, этот подход не был успешно использован для создания нокаутных крыс. Одна из проблем этой стратегии заключается в том, что SCNT крайне неэффективна. Первая опубликованная попытка имела успех менее 1%. [15] Альтернативно, мутагенез ENU является распространенной стратегией случайного нокаута генов мутагенеза у мышей, которую также можно использовать у крыс. Мутагенез ENU включает использование химического вещества N-этил-N-нитрозомочевины (ENU) для создания изменений отдельных оснований в геноме. ENU переносит свою этильную группу на радикалы кислорода или азота в ДНК, что приводит к неправильному спариванию и замене пар оснований. Мутантных животных можно получить путем инъекции самцу мыши ENU и скрещивания с самкой дикого типа для получения мутантного потомства. Мутагенез ENU создает высокую частоту случайных мутаций, при которой в любом гене меняется примерно одна пара оснований на каждые 200–700 гамет. [16] Несмотря на высокую мутагенность, физическое проникновение ЕНУ ограничено: у каждого самца мутирует только около 500 генов, и очень небольшое количество общих мутаций имеет наблюдаемый фенотип. Обычно у одного животного необходимо создать тысячи мутаций, чтобы создать один новый фенотип.
Несмотря на недавние улучшения в технологии ЕНУ, [17] [18] [19] картирование мутаций, ответственных за определенный фенотип, обычно сложно и требует много времени. Нейтральные мутации необходимо отделять от причинных мутаций посредством обширной селекции. ЕНУ и методы клонирования просто неэффективны для создания и картирования нокаутных генов у крыс для создания новых моделей болезней человека. В течение 2007 года крупнейший на сегодняшний день проект по мутагенезу крыс в ЕНУ, проводимый Медицинским колледжем Висконсина, смог произвести только 9 нокаутных линий крыс за пять лет при средней стоимости 200 000 долларов за нокаутную линию. Хотя некоторые компании все еще придерживаются этой стратегии, Медицинский колледж Висконсина перешел на более эффективный и коммерчески жизнеспособный метод с использованием мобильной ДНК и технологии CompoZr ZFN.
Нуклеазная технология «цинковые пальцы» и TALE
[ редактировать ]Нуклеазы цинковых пальцев (ZFN) и эффекторные нуклеазы, подобные активатору транскрипции (TALEN), представляют собой сконструированные ДНК-связывающие белки, которые облегчают целенаправленное редактирование генома путем создания двухцепочечных разрывов ДНК в указанных пользователем местах. Двухцепочечные разрывы важны для сайт-специфического мутагенеза, поскольку они стимулируют естественные процессы восстановления ДНК клетки, а именно гомологичную рекомбинацию и негомологичное соединение концов. Когда клетка использует негомологичный путь соединения концов для восстановления двухцепочечного разрыва, присущая репарации неточность часто приводит к точно направленным мутациям. В результате появляются эмбрионы с целевым нокаутом генов. [6] [20] Стандартные методы микроинъекции позволяют с помощью этой технологии получить нокаутных крыс через 4–6 месяцев. Основным преимуществом нокаута генов, опосредованного ZFN и TALEN, по сравнению с использованием мобильной ДНК является то, что конкретный ген может быть уникально и специфично нацелен на нокаут. Напротив, нокауты, сделанные с использованием технологии мобильной ДНК, являются случайными и поэтому маловероятно нацелены на интересующий ген.
Технология мобильной ДНК
[ редактировать ]Технология мобильной ДНК (прыгающих генов) использует ретротранспозоны и транспозоны для создания нокаутных моделей крыс. Эта платформенная технология отвечает всем критериям успешного подхода к нокауту генов у млекопитающих, позволяя осуществлять случайный мутагенез непосредственно в зародышевых клетках ( сперматозоидах и ооцитах ) модельных организмов млекопитающих, включая крыс. С помощью этой технологии гены разрушаются полностью и стабильно, выключаются с высокой частотой и случайным образом разрушаются по всему геному. Геномное расположение мутаций можно легко нанести на карту, создав библиотеку нокаутированных крыс для дальнейшего использования. После создания случайных нокаутных мутаций можно создавать более сложные мутации, такие как условные мутации, путем скрещивания нокаутных линий с линиями крыс, экспрессирующими рекомбиназу CRE тканеспецифичным образом. Нокаины могут быть вызваны обменом кассет, опосредованным рекомбинацией.
ДНК-транспозоны piggyBac (PB)
[ редактировать ]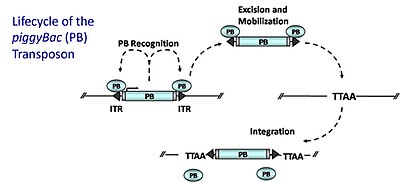
ДНК-транспозоны piggyBac (PB) мобилизуются по механизму «вырезать и вставить», при котором фермент транспозаза (PB-транспозаза), кодируемый самим транспозоном, вырезает и реинтегрирует транспозон в других участках генома. Транспозаза PB специфически распознает инвертированные концевые повторы PB (ITR), которые фланкируют транспозон; он связывается с этими последовательностями и катализирует удаление транспозона. Затем PB интегрируется на площадках TTAA. [21] по всему геному, относительно случайным образом. Для создания мутаций генной ловушки (или адаптации для создания трансгенных животных) транспозазу поставляют в транс-форме на одну плазмиду и котрансфицируют с плазмидой, содержащей донорский транспозон, рекомбинантный транспозон, содержащий генную ловушку, фланкированную сайтами связывания для транспозаза (ITR). Транспозаза будет катализировать удаление транспозона из плазмиды и последующую интеграцию в геном. Интеграция внутри кодирующей области захватывает элементы, необходимые для экспрессии генной ловушки. PB обладает несколькими идеальными свойствами: (1) он преимущественно вставляется внутри генов (от 50 до 67% вставок попадает в гены) (2) он не проявляет локального прыжка (широкий охват генома) (3) он не чувствителен кингибирование чрезмерного производства, при котором повышенные уровни транспозазы вызывают снижение транспозиции. 4) она аккуратно вырезается из донорского участка, не оставляя «следа», в отличие от Спящей красавицы. [22] [23]
Транспозоны Спящей красавицы (SB)
[ редактировать ]Транспозон «Спящая красавица» (SB) является производным суперсемейства ДНК-транспозонов Tc1/mariner, преобладающих в геномах как позвоночных, так и беспозвоночных. Однако эндогенные ДНК-транспозоны этого семейства совершенно неактивны в геномах позвоночных. Активный транспозон Tc1/mariner, синтезированный в результате выравнивания неактивных транспозонов из подсемейства элементов лососевых, «проснулся» и образовал транспозон, названный «Спящая красавица». [24] SB, как и другие ДНК-транспозоны, мобилизуется с помощью механизма вырезания и вставки, при котором фермент транспозаза, кодируемый самим транспозоном, вырезает и реинтегрирует транспозон в других участках генома. Белок SB из 340 аминокислот распознает инвертированные концевые повторы (ITR), фланкирующие транспозон; он связывается с этими последовательностями и катализирует удаление транспозона. Затем SB интегрируется в случайные участки генома, хотя некоторые исследования сообщают об очень незначительных предпочтениях транскрипционных единиц. [25] [26] Существует также простое требование наличия ТА-динуклеотида в целевом сайте, как и у всех транспозонов Tc1/mariner. [27]
Транспозон SB является мощным инструментом инсерционного мутагенеза у многих видов позвоночных. Недавно он продемонстрировал особую полезность для мутагенеза зародышевой линии как у мышей, так и у крыс. [28] [29] [30] [31] [32] [33] [34] Есть несколько преимуществ, которые делают SB очень привлекательным мутагеном, ориентированным на открытие генов: 1) он имеет небольшую предвзятость при вставке в определенные геномные области или в определенные последовательности распознавания, 2) вставки транспозона de novo обеспечивают «маркированный» маркер последовательности для быстрая идентификация конкретной мутации с помощью простых методов ПЦР-клонирования; 3) инсерционный мутагенез SB in vivo позволяет быстро и легко генерировать множественные мутации у одного животного и в одной ткани, такой как аденоматозный полип.
LINE1 (L1) ретротранспозоны
[ редактировать ]Транспозоны и ретротранспозоны являются ценными инструментами для объективного открытия генов, поскольку мобильные фрагменты ДНК используются для разрушения генов. Ретротранспозоны, такие как LINE (длинно вкрапленные ядерные элементы), мобилизуются по механизму «копирования и вставки» и широко распространены у многих видов эукариот. Некоторые ретротранспозоны L1 остались активными у мышей и людей. L1 содержат небольшой внутренний промотор в 5'-нетранслируемой области для управления экспрессией, две открытые рамки считывания (ORF) и 3'-нетранслируемую область, содержащую последовательности для полиаденилирования. Две ORF кодируют белки, необходимые для автономной ретротранспозиции; ORF1 кодирует РНК -связывающий белок, тогда как ORF2 кодирует белок, обладающий активностью эндонуклеазы (EN) и обратной транскриптазы (RT), которые разрывают сайт в ДНК, а затем создают копию посредством RT. Эти белки обладают подавляющей специфичностью связывания и действия на транскрипт, который их кодирует, что обеспечивает почти эксклюзивную мобилизацию родительской РНК L1. Используя RT-активность белка ORF2, транскрибируемая РНК L1 копируется в ДНК с помощью процесса, называемого обратной транскрипцией с целевой праймацией (TPRT). [35] и интегрированы в геном. Интеграция происходит с небольшим смещением для любой конкретной области генома, требуя простой консенсусной последовательности 5'TTTT'A-3' (наряду с незначительными вариациями этой последовательности). Интегрированные последовательности L1 часто усекаются на 5'-конце, их средний общий размер составляет 1 КБ, многие из них содержат только 3'-концевые последовательности.
Природа ретротранспозиции наделяет L1 некоторыми уникальными преимуществами; Ретротранспозоны L1 имеют практически неограниченный запас инсерционного мутагена, поскольку он постоянно транскрибируется с промотора, что было бы полезно для применений, где требуется большое количество мутаций в одной клетке. Элементы L1 также демонстрируют широкий геномный охват с в значительной степени случайным распределением вставок. [36] [37] [38] Вставки L1 в геномные сайты также необратимы, и, таким образом, любое мутагенное событие, вызванное вставкой L1, «помечается» последовательностями L1.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Эбботт А: Лабораторные животные: крыса эпохи Возрождения. Природа 2004, 428:464-466.
- ^ Чжоу К., Ренар Дж.П., Ле Фрик Г., Брошар В., Божан Н., Шерифи Ю., Фрейшар А., Коцци Дж.: Создание плодовитых клонированных крыс путем регуляции активации ооцитов. Наука 2003, 302:1179.
- ^ Джастис М.Дж., Новероске Дж.К., Вебер Дж.С., Чжэн Б., Брэдли А.: Мутагенез мышиного ЕНУ. Хум Мол Женет 1999, 8:1955–1963.
- ^ Китада К., Ишишита С., Тосака К., Такахаши Р., Уэда М., Кенг В.В., Хори К., Такеда Дж. Мутагенез с мечением транспозонов у крыс. Nat Methods 2007, 4:131–133.
- ^ Jump up to: а б Консорциум проекта секвенирования генома крысы. Последовательность генома коричневой крысы дает представление об эволюции млекопитающих. Природа, 2004. 428(6982): с. 493-521.
- ^ Jump up to: а б Гертс, AM и др. др., Нокаутированные крысы посредством микроинъекции в эмбрион нуклеаз с цинковыми пальцами. Наука. Том 325: 433 (24 июля 2009 г.) Гертс, AM; Стоимость, ГДж; Фрейверт, Ю.; Цейтлер, Б.; Миллер, Дж. К.; Цой, В.М.; Дженкинс, СС; Вуд, А.; Кюи, X.; Мэн, X.; Винсент, А.; Лам, С.; Михалкевич, М.; Шиллинг, Р.; Фоклер, Дж.; Каллоуэй, С.; Вейлер, Х.; Менорет, С.; Анегон, И.; Дэвис, Джорджия; Чжан, Л.; Ребар, Э.Дж.; Грегори, доктор медицинских наук; Урнов, ФД; Джейкоб, HJ; Бьюлоу, Р. (2009). «Нокаутные крысы посредством микроинъекции в эмбрион нуклеаз с цинковыми пальцами» . Наука . 325 (5939): 433. Бибкод : 2009Sci...325..433G . дои : 10.1126/science.1172447 . ПМЦ 2831805 . ПМИД 19628861 .
- ^ Вицек, Эндрю. «Год крысы» , BioTechniques , 01.10.2009.
- ^ «Сигма-Олдрич разрабатывает модели болезни Паркинсона». Архивировано 8 октября 2009 г. в Wayback Machine , LaboratoryTalk.
- ^ Международный консорциум по секвенированию генома человека, Первоначальное секвенирование и анализ генома человека. Природа, 2001. 409(6822): с. 860-921.
- ^ Консорциум по секвенированию генома мыши, Первоначальное секвенирование и сравнительный анализ генома мыши. Природа, 2002. 420(6915): с. 520-62.
- ^ Эбботт, А., Лабораторные животные: крыса эпохи Возрождения. Природа, 2004. 428(6982): с. 464-6.
- ^ Линдблад-То, К., Секвенирование генома: компания троих. Природа, 2004. 428(6982): с. 475-6.
- ^ Замбрович, 1998; Скарнес и др.др., 2004; К и др., 2004; Норд и др., 2006 г.
- ^ Коэн-Таннуджи, М. и К. Бабине, Помимо «нокаутных» мышей: новые перспективы запрограммированной модификации генома млекопитающих. Молекулярная репродукция человека, 1998. 4(10): с. 929-38.
- ^ Чжоу, К., Дж. П. Ренар, Г. Ле Фрик, В. Брошар, Н. Божан, Ю. Шерифи, А. Фрейшар и Дж. Коцци, Создание плодовитых клонированных крыс путем регулирования активации ооцитов. Наука, 2003. 302(5648): с. 1179.
- ^ Хитоцумачи, С., Д.А. Карпентер и В.Л. Рассел, Повторение дозы увеличивает мутагенную эффективность N-этил-N-нитрозомочевины в сперматогониях мышей. Труды Национальной академии наук Соединенных Штатов Америки, 1985. 82 (19): с. 6619-21.
- ^ Браун, С.Д. и Р.Э. Хардисти, Стратегии мутагенеза для выявления новых локусов, связанных с фенотипами заболеваний. Семинары по клеточной биологии и биологии развития, 2003. 14 (1): с. 19-24.
- ^ Чен, Ю., Д. Йи, К. Дэйнс, А. Чаттерджи, Дж. Кавальколи, Э. Шнайдер, Дж. Ом, Р. П. Войчик и Т. Магнусон, Скрининг на основе генотипа для выявления мутаций, индуцированных ENU, в эмбриональных мышах. стволовые клетки. Nature Genetics, 2000. 24(3): с. 314-7.
- ^ Зан, Ю., Дж. Д. Хааг, К. С. Чен, Л. А. Шепель, Д. Вигингтон, Ю. Р. Ван, Р. Ху, К. К. Лопес-Гуахардо, Х. Л. Брозе, К. И. Портер, Р. А. Леонард, А. А. Хитт, С. Л. Шоммер, А. Ф. Элегбеде и М. Н. Гулд, Производство нокаутных крыс с использованием мутагенеза ENU и скринингового анализа на основе дрожжей. Природная биотехнология, 2003. 21(6): с. 645-51.
- ^ Тессон и др., Нокаутные крысы, полученные путем микроинъекции эмбрионам TALEN. Природная биотехнология, том 29: 695-96 (5 августа 2011 г.) Тессон, Л.; Усал, К.; Меноре, СВ; Люнг, Э.; Найлз, Би Джей; Реми, СВ; Сантьяго, Ю.; Винсент, AI; Мэн, X.; Чжан, Л.; Грегори, доктор медицинских наук; Анегон, И.; Стоимость, ГДж (2011). «Нокаутные крысы, полученные путем микроинъекции эмбрионов TALEN» . Природная биотехнология . 29 (8): 695–696. дои : 10.1038/nbt.1940 . ПМИД 21822240 .
- ^ Фрейзер, MJ и др., Точное удаление TTAA-специфичных транспозонов чешуекрылых piggyBac (IFP2) и tagalong (TFP3) из генома бакуловируса в клеточных линиях двух видов чешуекрылых. Насекомое Мол Биол, 1996. 5(2): с. 141-51.
- ^ Митра, Р., Дж. Фейн-Торнтон и Н.Л. Крейг, piggyBac может обходить синтез ДНК во время транспозиции вырезания и вставки. ЭМБО Дж, 2008.
- ^ Динг, С. и др., Эффективная транспозиция транспозона piggyBac (PB) в клетках млекопитающих и мышей. Cell, 2005. 122(3): с. 473-83.
- ^ Ивикс З. и др., Молекулярная реконструкция Спящей красавицы, Tc1-подобного транспозона рыбы, и его транспозиция в клетках человека. Cell, 1997. 91(4): с. 501-10.
- ^ Вигдаль, Т.Дж. и др., Общие физические свойства ДНК, влияющие на выбор целевого участка спящей красавицы и других мобильных элементов Tc1/mariner. J Mol Biol, 2002. 323(3): с. 441-52.
- ^ Янт, С.Р. и др., Полногеномное картирование интеграции транспозонов у млекопитающих в высоком разрешении. Мол Селл Биол, 2005. 25(6): с. 2085-94.
- ^ Пластерк, Р.Х., З. Изсвак и З. Ивикс, Постоянные инопланетяне: суперсемейство мобильных элементов Tc1/mariner. Тенденции Жене, 1999. 15(8): с. 326-32.
- ^ Гертс, А.М. и др., Генные мутации и геномные перестройки у мышей в результате мобилизации транспозонов из хромосомных конкатемеров. PLoS Genet, 2006. 2(9): с. е156.
- ^ Хори, К. и др., Характеристика транспозиции «Спящей красавицы» и ее применение для генетического скрининга на мышах. Мол Селл Биол, 2003. 23(24): с. 9189-207.
- ^ Кенг, В.В. и др., Регионально-специфичный насыщающий мутагенез зародышевой линии у мышей с использованием транспозонной системы «Спящая красавица». Nat Methods, 2005. 2(10): с. 763-9.
- ^ Китада, К. и др., Мутагенез с мечением транспозонов у крыс. Nat Methods, 2007. 4(2): с. 131-3
- ^ Гертс, А.М. и др., Условная экспрессия генов у мышей с использованием транспозона-ловушки гена Спящей красавицы. BMC Biotechnol, 2006. 6: с. 30.
- ^ Дюпюи, А.Дж., С. Фриц и Д.А. Ларгаэспада, Транспозиция и нарушение генов в зародышевой линии самцов мышей. Бытие, 2001. 30(2): с. 82-8.
- ^ Дюпюи, А.Дж. и др., Трансгенез зародышевой линии млекопитающих путем транспозиции. Proc Natl Acad Sci USA, 2002. 99(7): с. 4495-9.
- ^ Луан, Д.Д., М.Х. Корман, Дж.Л. Якубчак и Т.Х. Эйкбуш, Обратная транскрипция РНК R2Bm инициируется разрывом в хромосомном целевом сайте: механизм ретротранспозиции без LTR. Cell, 1993. 72(4): с. 595-605.
- ^ Остертаг, Э.М. и др., Мышиная модель ретротранспозиции L1 человека. Нат Женет, 2002. 32(4): с. 655-60.
- ^ Babushok, D.V., et al., L1 integration in a transgenic mouse model. Genome Res, 2006. 16(2): p. 240-50.
- ^ Стоимость, Г.Дж. и Дж.Д. Буке, Нацеливание на интеграцию ретротранспозонов человека определяется специфичностью эндонуклеазы L1 к областям необычной структуры ДНК. Биохимия, 1998. 37(51): с. 18081-93.