Полиаденилирование

Полиаденилирование - это добавление поли (a) хвоста к транскрипту РНК, как правило, РНК мессенджера (мРНК). Поли (а) хвост состоит из множества аденозиновых монофосфатов ; Другими словами, это участок РНК, которая имеет только адениновые основания. У эукариот полиаденилирование является частью процесса, который продуцирует зрелую мРНК для трансляции . Во многих бактериях поли (а) хвост способствует деградации мРНК. Следовательно, он является частью более широкого процесса экспрессии генов .
Процесс полиаденилирования начинается транскрипция гена как завершается . сегмент 3'-самый вновь изготовленной пре-мРНК сначала расщепляется набором белков ; Эти белки затем синтезируют поли (A) хвост на 3 'конец РНК. В некоторых генах эти белки добавляют поли (а) хвост в одном из нескольких возможных сайтов. Следовательно, полиаденилирование может создавать более одного транскрипта из одного гена ( альтернативное полиаденилирование ), аналогичное альтернативному сплайсингу . [ 1 ]
Поли (а) хвост важен для ядерного экспорта, трансляции и стабильности мРНК. Хвост сокращается с течением времени, и, когда он достаточно короткий, мРНК ферментативно разлагается. [ 2 ] Однако в нескольких типах клеток мРНК с короткими поли (а) хвостами хранятся для последующей активации путем повторного полиаденилирования в цитозоле. [ 3 ] Напротив, когда полиаденилирование происходит у бактерий, оно способствует деградации РНК. [ 4 ] Это также иногда случается с эукариотическими некодирующими РНК . [ 5 ] [ 6 ]
Молекулы мРНК как у прокариот, так и у эукариот имеют полиаденилированные 3'-коне, с прокариотическими поли (а) хвостами, как правило, более короче и меньше молекул мРНК, полиаденилированные. [ 7 ]
Фон на РНК
[ редактировать ]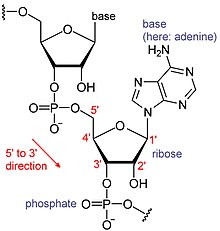
РНК - это тип крупных биологических молекул, отдельные строительные блоки которых называются нуклеотидами. Название поли (а) хвост (для хвоста полиадениловой кислоты) [ 8 ] Отражает способ сокращения нуклеотидов РНК, с буквой для основания, которое содержит нуклеотид (для аденина , C для цитозина , G для гуанина и U для урацила ). РНК продуцируются ( транскрибируются ) из матрицы ДНК . По соглашению, РНК -последовательности записываются в направлении от 5 'до 3'. 5' -конец является частью молекулы РНК, которая сначала транскрибируется, а 3' -конец транскрибируется последним. 3' -конец также, где поли (а) хвост обнаруживается на полиаденилированных РНК. [ 1 ] [ 9 ]
РНКсенджер (мРНК) представляет собой РНК, которая имеет кодирующую область, которая действует как матрица для синтеза белка ( трансляция ). Остальная часть мРНК, нетранслируемые области , настраивают, насколько активна мРНК. [ 10 ] Есть также много РНК, которые не переведены, называемые некодирующими РНК. Как и нетранслируемые регионы, многие из этих некодирующих РНК имеют регуляторные роли. [ 11 ]
Ядерное полиаденилирование
[ редактировать ]Функция
[ редактировать ]При ядерном полиаденилировании поли (A) хвост добавляется в РНК в конце транскрипции. На мРНК хвост поли (A) защищает молекулу мРНК от ферментативной деградации в цитоплазме и помогает в прекращении транскрипции, экспорте мРНК из ядра и трансляции. [ 2 ] Почти все эукариотические мРНК полиаденилированы, [ 12 ] За исключением того, что животные, зависящие от репликации, гистоновые мРНК. [ 13 ] Это единственные мРНК у эукариот, в которых отсутствует поли (а) хвоста, заканчивая вместо этого в структуре ствола, за которой следует последовательность, богатую пурином, называемая гистоновым нижним элементом, которая направляет, где РНК вырезана так, чтобы 3'-конец гистоновой мРНК образуется. [ 14 ]
Многие эукариотические некодирующие РНК всегда полиаденилированы в конце транскрипции. Существуют небольшие РНК, в которых поли (а) хвост наблюдается только в промежуточных формах, а не в зрелой РНК, поскольку концы удаляются во время обработки, примечательными являются микроРНК . [ 15 ] [ 16 ] Но для многих длинных некодирующих РНК - казалось бы, большой группы регуляторных РНК, которые, например, включают РНК -XIST , которая опосредует инактивацию Х -хромосомы - поли (а) хвост является частью зрелой РНК. [ 17 ]
Механизм
[ редактировать ]| Белки вовлечены: [ 12 ] [ 18 ] CPSF : фактор специфичности расщепления/полиаденилирования |
Процессовный , комплекс полиаденилирования в ядре эукариот работает на продуктах РНК -полимеразы II таких как мРНК -предшественника . Здесь многопротеиновый комплекс (см. Компоненты справа) [ 18 ] расщепляет 3'-самую часть вновь продуцированной РНК и полиаденилирует конец, полученный этим расщеплением. Расщепление катализируется ферментом CPSF [ 13 ] [ 18 ] и происходит 10–30 нуклеотидов ниже по течению от сайта связывания. [ 19 ] Этот сайт часто имеет последовательность сигнала полиаденилирования AAUAAA на РНК, но существуют его варианты, которые связываются более слабо с CPSF . [ 18 ] [ 20 ] Два других белка добавляют специфичность к связыванию с РНК: CSTF и CFI. CSTF связывается с областью, богатой GU, в дальнейшем по течению от сайта CPSF. [ 21 ] CFI распознает третий сайт на РНК (набор последовательностей Uguaa у млекопитающих [ 22 ] [ 23 ] [ 24 ] ) и может рекрутировать CPSF, даже если последовательность AAUAAA отсутствует. [ 25 ] [ 26 ] Сигнал полиаденилирования - мотив последовательности, распознаваемый комплексом расщепления РНК - варьируется между группами эукариот. Большинство участков полиаденилирования человека содержат последовательность AAUAAA, [ 21 ] Но эта последовательность встречается реже в растениях и грибах. [ 27 ]
РНК обычно расщепляется перед завершением транскрипции, поскольку CSTF также связывается с РНК -полимеразой II. [ 28 ] Благодаря плохо изученному механизму (по состоянию на 2002 г.) он сигнализирует РНК -полимеразу II, чтобы ускользнуть от транскрипта. [ 29 ] Расчет также включает в себя белок CFII, хотя это неизвестно как. [ 30 ] Сайт расщепления, связанный с сигналом полиаденилирования, может варьироваться до 50 нуклеотидов. [ 31 ]
Когда РНК расщепляется, начинается полиаденилирование, катализируется полиаденилат -полимеразой. Полиаденилат -полимераза строит поли (а) хвост, добавляя аденозин -монофосфатные единицы из аденозинтрифосфата в РНК, расщепляя пирофосфат . [ 32 ] Другой белок, PAB2, связывается с новым коротким поли (A) хвостом и увеличивает аффинность полиаденилат -полимеразы к РНК. Когда поли (a) хвост составляет приблизительно 250 нуклеотидов длиной, фермент больше не может связываться с CPSF, а полиаденилирование остановки, таким образом определяя длину поли (а) хвоста. [ 33 ] [ 34 ] CPSF находится в контакте с РНК -полимеразой II, что позволяет ему сигнализировать полимеразу для прекращения транскрипции. [ 35 ] [ 36 ] Когда РНК -полимераза II достигает «последовательности завершения» (⁵'tttatt 3 'На шаблоне ДНК и ⁵aauaaa 3 «На первичной транскрипции) конец транскрипции сигнализируется. [ 37 ] Механизм полиаденилирования также физически связан с сплайсосомой , комплексом, который удаляет интроны из РНК. [ 26 ]
Последующие эффекты
[ редактировать ]Поли (а) хвост действует как сайт связывания для поли (а)-связывающего белка . Поли (а) -связывающий белок способствует экспорту из ядра и трансляции и ингибирует деградацию. [ 38 ] Этот белок связывается с поли (а) хвостом перед экспортом мРНК из ядра, а в дрожжах также рекрутирует поли (а) нуклеаза, фермент, который сокращает поли (а) хвост и позволяет экспортировать мРНК. Поли (A) -связывающий белок экспортируется в цитоплазму с РНК. МРНК, которые не экспортируются, деградируются экзосомой . [ 39 ] [ 40 ] Поли (а) -связывающий белок также может связываться с несколькими белками, которые влияют на трансляцию, [ 39 ] Одним из них является коэффициент инициации -4G, который, в свою очередь, набирает рибосомную субъединицу 40 -х годов . [ 41 ] Тем не менее, поли (а) хвост не требуется для перевода всех мРНК. [ 42 ] Кроме того, поли (а) хвостовой (олигоаденилирование) может определить судьбу молекул РНК, которые обычно не являются поли (а) хвост (такие как (маленькие) некодирующие (SN) РНК и т. Д.) и тем самым индуцируют их РНК разлагаться. [ 43 ]
Дребетяция
[ редактировать ]В эукариотических соматических клетках поли (а) хвостов большинства мРНК в цитоплазме постепенно становятся короче, а мРНК с более коротким поли (а) хвостом переводится меньше и разлагаются раньше. [ 44 ] Тем не менее, это может занять много часов, прежде чем мРНК разлагается. [ 45 ] Этот процесс поэтапления и деградации может быть ускорен с помощью микроРНК, дополненных 3' -нетранслируемой области мРНК. [ 46 ] В незрелых яичных клетках мРНК с укороченными поли (а) хвостами не разлагаются, а вместо этого хранятся и трансляционно неактивны. Эти короткие хвостовые мРНК активируются цитоплазматическим полиаденилированием после оплодотворения во время активации яиц . [ 47 ]
У животных поли (а) рибонуклеаза ( Парн ) может связываться с 5' -крышей и удалять нуклеотиды из поли (а) хвоста. Уровень доступа к 5' -шапке и поли (а) хвосту важен для контроля того, как скоро мРНК разлагается. Парн детеенилат меньше, если РНК связана факторами инициации 4e (в 5' -шапке) и 4G (при поли (а) хвосте), поэтому трансляция уменьшает деветенилирование. Скорость детеенилирования также может регулироваться РНК-связывающими белками. Кроме того, РНК -тройные структуры спирали и мотивы РНК, такие как поли (а) хвоста 3 'Конечный карманный карманный процесс замедления мехенилирования и ингибируют удаление поли (а) хвоста. [ 48 ] Как только поли (а) хвост удаляется, комплекс окупания удаляет 5' -шаблон, что приводит к деградации РНК. Несколько других белков участвуют в дегенилировании в почкующихся дрожжах и клетках человека, особенно в комплексе CCR4-NOT . [ 49 ]
Цитоплазматическое полиаденилирование
[ редактировать ]Существует полиаденилирование в цитозоле некоторых типов животных клеток, а именно в зародышевой линии , во время раннего эмбриогенеза и в пост- синаптических местах нервных клеток . Это удлиняет поли (а) хвост мРНК с укороченным поли (а) хвостом, так что мРНК будет переведена . [ 44 ] [ 50 ] Эти укороченные поли (а) хвосты часто менее 20 нуклеотидов и удлиняются до около 80–150 нуклеотидов. [ 3 ]
У раннего эмбриона мыши цитоплазматическое полиаденилирование материнских РНК из яичной клетки позволяет клетке выживать и расти, даже если транскрипция не начинается до середины 2-клеточной стадии (4-клеточная стадия у человека). [ 51 ] [ 52 ] В мозге цитоплазматическое полиаденилирование является активным во время обучения и может играть роль в долгосрочном потенцировании , что является укреплением передачи сигнала от нервной клетки к другой в ответ на нервные импульсы и важно для обучения и формирования памяти. [ 3 ] [ 53 ]
Цитоплазматическое полиаденилирование требует РНК-связывающих белков CPSF и CPEB , и может включать другие РНК-связывающие белки, такие как Pumilio . [ 54 ] В зависимости от типа клетки полимераза может быть тем же типом полиаденилат-полимеразы (PAP), которая используется в ядерном процессе, или цитоплазматической полимеразы GLD-2 . [ 55 ]
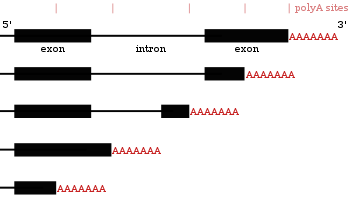
Альтернативное полиаденилирование
[ редактировать ]Многие гены, кодирующие белок, имеют более одного сайта полиаденилирования, поэтому ген может кодировать для нескольких мРНК, которые отличаются по их 3'-конце . [ 27 ] [ 56 ] [ 57 ] 3 'область транскрипта содержит множество сигналов полиаденилирования (PAS). Когда используются более проксимальные (ближе к 5 'конечному) сайтам PAS, это сокращает длину 3' нетранслируемой области (3 'UTR) транскрипта. [ 58 ] Исследования как у людей, так и у мух показали, что APA специфична для тканей. С нейрональными тканями предпочитают дистальное использование PAS, что приводит к более длинным 3 'тканям UTR и яичкам, предпочитающим проксимальную PAS, что приводит к более коротким 3' UTR. [ 59 ] [ 60 ] Исследования показали, что существует корреляция между уровнем сохранения гена и его тенденцией к альтернативному полиаденилированию с высоко консервативными генами, демонстрирующими больше APA. Точно так же высоко экспрессируемые гены следуют этой же паттерне. [ 61 ] Данные рибо-последовательности (секвенирование только мРНК внутри рибосомов) показали, что изоформы мРНК с более коротким 3 'UTR с большей вероятностью будут переведены. [ 58 ]
Поскольку альтернативное полиаденилирование изменяет длину 3 'UTR , [ 62 ] Это также может изменить, какие сайты связывания доступны для микроРНК в 3' -UTR. [ 19 ] [ 63 ] МикроРНК, как правило, подавляют трансляцию и способствуют деградации мРНК, с которыми они связываются, хотя есть примеры микроРНК, которые стабилизируют транскрипты. [ 64 ] [ 65 ] Альтернативное полиаденилирование также может сократить кодирующую область, что делает код мРНК для другого белка, [ 66 ] [ 67 ] Но это гораздо реже, чем просто сокращение 3' -нетранслируемой области. [ 27 ]
На выбор поли (а) сайта может влиять внеклеточные стимулы и зависит от экспрессии белков, которые принимают участие в полиаденилировании. [ 68 ] [ 69 ] Например, экспрессия CSTF-64 , субъединицы стимулирующего фактора расщепления (CSTF), увеличивается в макрофагах в ответ на липополисахариды (группа бактериальных соединений, которые вызывают иммунный ответ). Это приводит к выбору слабых поли (а) сайтов и, следовательно, более коротких транскриптов. Это удаляет регуляторные элементы в 3'-нетранслируемых областях мРНК для продуктов, связанных с защитой, таких как лизоцим и TNF-α . Эти мРНК затем имеют более длительный период полураспада и производят больше этих белков. [ 68 ] РНК-связывающие белки, отличные от белков в машине полиаденилирования, также могут влиять на то, используется ли сайт полиаденилирования, [ 70 ] [ 71 ] [ 72 ] [ 73 ] как может метилирование ДНК вблизи сигнала полиаденилирования. [ 74 ] Кроме того, многочисленные другие компоненты, участвующие в транскрипции, сплайсинге или других механизмах, регулирующих биологию РНК, могут влиять на APA. [ 75 ]
Метка для деградации у эукариот
[ редактировать ]Для многих некодирующих РНК , включая тРНК , рРНК , SNRNA и Snorna , полиаденилирование-это способ отметить РНК для деградации, по крайней мере, у дрожжей . [ 76 ] Это полиаденилирование выполняется в ядре комплексом бродяги , который поддерживает хвост, который составляет около 4 нуклеотидов длиной до 3' -конца. [ 77 ] [ 78 ] Затем РНК деградируется экзосомой . [ 79 ] Поли (а) хвосты также были обнаружены на фрагментах рРНК человека, как в форме гомополимерных (единственных), так и гетерополимерных (в основном а) хвостов. [ 80 ]
В прокариотах и органеллах
[ редактировать ]
Во многих бактериях как мРНК, так и некодирующие РНК могут быть полиаденилированы. Этот поли (a) хвост способствует деградации дегросомой , которая содержит два фермента, разрушающие РНК: полинуклеотидфосфорилазу и РНКазу E. Полинуклеотидфосфорилаза связывается с 3' -концом РНК, а 3' -расширение, обеспечиваемое поли (а) хвостом, позволяет ему связываться с РНК, вторичная структура , в противном случае блокировала бы 3' -конец. Последовательные раунды полиаденилирования и деградации 3' -конца полинуклеотидфосфорилазой позволяют дегложосоме преодолевать эти вторичные структуры. Поли (а) хвост также может рекрутировать РНКазы, которые разрезают РНК в двух. [ 81 ] Эти бактериальные поли (а) хвосты имеют длину около 30 нуклеотидов. [ 82 ]
В таких разных группах, как животные и трипаносомы , митохондрии содержат как стабилизирующие, так и дестабилизирующие поли (а) хвосты. Дестабилизация полиаденилирования нацелен как на мРНК, так и некодирующие РНК. Поли (а) хвосты в среднем составляют 43 нуклеотида. Стабилизирующие начинаются у стоп -кодона, и без них стоп -кодон (UAA) не является полным, поскольку геном только кодирует часть U или UA. Растительные митохондрии имеют только дестабилизирующее полиаденилирование. Митохондриальное полиаденилирование никогда не наблюдалось ни в яростях, ни в дрожжах деления. [ 83 ] [ 84 ]
В то время как многие бактерии и митохондрии имеют полиаденилат -полимеразы, они также имеют другой тип полиаденилирования, выполняемый самим полинуклеотидфосфорилазой . Этот фермент находится в бактериях, [ 85 ] митохондрия, [ 86 ] пластиды [ 87 ] и как составляющая архаальной экзосомы (в тех археи , которые имеют экзосому ). [ 88 ] Он может синтезировать 3 'расширение, где подавляющее большинство оснований являются аденинами. Как и у бактерий, полиаденилирование полинуклеотидфосфорилазой способствует деградации РНК в пластидах [ 89 ] и, вероятно, также архаи. [ 83 ]
Эволюция
[ редактировать ]Хотя полиаденилирование наблюдается почти во всех организмах, оно не универсально. [ 7 ] [ 90 ] Тем не менее, широкое распределение этой модификации и тот факт, что она присутствует в организмах из всех трех доменов жизни, подразумевает, что последний универсальный общий предок всех живых организмов, как предполагается, имел некоторую форму системы полиаденилирования. [ 82 ] Несколько организмов не имеют полиаденилатной мРНК, что подразумевает, что они потеряли свои полиаденилиционные механизмы во время эволюции. Хотя никаких примеров эукариот, в которых отсутствует полиаденилирование, не известно, мРНК из бактерии Mycoplasma gallisepticum и соленой архейской вулкани Haloferax не хватает этой модификации. [ 91 ] [ 92 ]
Самым древним полиаденилирующим ферментом является полинуклеотидфосфорилаза . Этот фермент является частью как бактериальной деградзомы , так и архаальной экзосомы , [ 93 ] два тесно связанных комплекса, которые перерабатывают РНК в нуклеотиды. Этот фермент разрушает РНК, атакуя связь между 3'-самыми нуклеотидами с фосфатом, разбивая дифхосфатный нуклеотид. Эта реакция обратима, и поэтому фермент также может расширять РНК с большим количеством нуклеотидов. Гетерополимерный хвост, добавленный полинуклеотидфосфорилазой, очень богат аденином. Выбор аденина, скорее всего, является результатом более высоких концентраций ADP , чем другие нуклеотиды в результате использования АТФ в качестве энергетической валюты, что делает его более вероятностью включать в этот хвост в ранние формы жизни. Было высказано предположение, что участие богатых аденином хвостов в деградации РНК вызвало более позднюю эволюцию полиаденилат-полимераз (ферменты, которые продуцируют поли (а) хвосты без других нуклеотидов в них). [ 94 ]
Полиаденилат -полимеразы не такие древние. Они отдельно развивались как у бактерий, так и в эукариотах из фермента, добавляющего CCA , который является ферментом, который завершает 3'-концы тРНК . Его каталитический домен гомологичен таковым другим полимеразам . [ 79 ] Предполагается, что горизонтальный перенос бактериального фермента CCA-добавления к эукариотам позволил архейноподобному ферменту CCA-добавления для переключения функции на поли (а) полимеразу. [ 82 ] Некоторые линии, такие как археи и цианобактерии , никогда не эволюционировали полиаденилат -полимеразу. [ 94 ]
Полиаденилатные хвосты наблюдаются в нескольких РНК -вирусах , включая грипп А , [ 95 ] Coronavirus , [ 96 ] Вирус мозаики люцерны , [ 97 ] и уток гепатит А. [ 98 ] Некоторые вирусы, такие как ВИЧ-1 и полиовирус , ингибируют полити-белок, связывающий клетку ( PABPC1 ), чтобы подчеркнуть экспрессию их собственных генов над клеткой хозяина. [ 99 ]
История
[ редактировать ]Поли (а) полимераза была впервые идентифицирована в 1960 году как ферментативную активность в экстрактах, изготовленных из ядер клеток, которые могут полимеризировать АТФ, но не ADP, в полиаденин. [ 100 ] [ 101 ] Хотя это идентифицировано во многих типах клеток, эта активность не имела известной функции до 1971 года, когда в мРНК были обнаружены поли (а) последовательности. [ 102 ] [ 103 ] Считалось, что единственная функция этих последовательностей - это защита 3' -конца РНК от нуклеаз, но позже были идентифицированы специфические роли полиаденилирования в ядерном экспорте и трансляции. Полимеразы, ответственные за полиаденилирование, были впервые очищены и охарактеризованы в 1960 -х и 1970 -х годах, но большое количество вспомогательных белков, которые контролировали этот процесс, было обнаружено только в начале 1990 -х годов. [ 102 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный Proudfoot NJ, Furger A, Dye MJ (февраль 2002 г.). «Интеграция обработки мРНК с транскрипцией» . Клетка . 108 (4): 501–12. doi : 10.1016/s0092-8674 (02) 00617-7 . PMID 11909521 . S2CID 478260 .
- ^ Jump up to: а беременный Guhaniyogi J, Brewer G (март 2001 г.). «Регуляция стабильности мРНК в клетках млекопитающих» . Ген . 265 (1–2): 11–23. doi : 10.1016/s0378-1119 (01) 00350-x . PMC 3340483 . PMID 11255003 .
- ^ Jump up to: а беременный в Рихтер Д.Д. (июнь 1999 г.). «Цитоплазматическое полиаденилирование в развитии и за его пределами» . Микробиология и молекулярная биология обзоры . 63 (2): 446–56. doi : 10.1128/mmbr.63.2.446-456.1999 . PMC 98972 . PMID 10357857 .
- ^ Steege da (август 2000 г.). «Новые особенности распада мРНК у бактерий» . РНК . 6 (8): 1079–90. doi : 10.1017/s135583820000001023 . PMC 1369983 . PMID 10943888 .
- ^ Zhuang Y, Zhang H, Lin S (июнь 2013 г.). «Полиаденилирование 18S рРНК в водорослях (1)». Журнал Phycology . 49 (3): 570–9. Bibcode : 2013jpcgy..49..570z . doi : 10.1111/jpy.12068 . PMID 27007045 . S2CID 19863143 .
- ^ Андерсон JT (август 2005 г.). «Оборот РНК: неожиданные последствия хвоста» . Текущая биология . 15 (16): R635-8. Bibcode : 2005cbio ... 15.r635a . doi : 10.1016/j.cub.2005.08.002 . PMID 16111937 . S2CID 19003617 .
- ^ Jump up to: а беременный Саркар Н (июнь 1997 г.). «Полиаденилирование мРНК у прокариот». Ежегодный обзор биохимии . 66 (1): 173–97. doi : 10.1146/annurev.biochem.66.1.173 . PMID 9242905 .
- ^ Стивенс А (1963). «Биозинтез рибонуклеиновых кислот и деградация». Ежегодный обзор биохимии . 32 : 15–42. doi : 10.1146/annurev.bi.32.070163.000311 . PMID 14140701 .
- ^ Lehninger AL, Nelson DL, Cox MM, Eds. (1993). Принципы биохимии (2 -е изд.). Нью -Йорк: Стоит. ISBN 978-0-87901-500-8 . [ страница необходима ]
- ^ Abaza I, Gebauer F (март 2008 г.). «Торговый перевод с РНК-связывающими белками» . РНК . 14 (3): 404–9. doi : 10.1261/rna.848208 . PMC 2248257 . PMID 18212021 .
- ^ Mattick JS, Makunin IV (апрель 2006 г.). «Некодирующая РНК» . Молекулярная генетика человека . 15 Spec № 1 (90001): R17-29. doi : 10.1093/hmg/ddl046 . PMID 16651366 .
- ^ Jump up to: а беременный Hunt AG, Xu R, Addepalli B, Rao S, Forbes KP, Meeks LR, Xing D, Mo M, Zhao H, Bandyopadhyay A, Mampanaboina L, Marion A, Von Lanken C, Li QQ (май 2008 г.). «Механизм полиаденилирования мРНК арабидопсиса: всесторонний анализ взаимодействия белкового белка и профилирования экспрессии генов» . BMC Genomics . 9 : 220. DOI : 10.1186/1471-2164-9-220 . PMC 2391170 . PMID 18479511 .
- ^ Jump up to: а беременный Dávila López M, Samuelsson T (январь 2008 г.). «Ранняя эволюция гистон -мРНК 3' -обработки» . РНК . 14 (1): 1–10. doi : 10.1261/rna.782308 . PMC 2151031 . PMID 17998288 .
- ^ Marzluff WF, Gongidi P, Woods KR, Jin J, Maltais LJ (ноябрь 2002 г.). «Гены гистонов, зависящие от репликации человека и мыши». Геномика . 80 (5): 487–98. doi : 10.1016/s0888-7543 (02) 96850-3 . PMID 12408966 .
- ^ Saini HK, Griffiths-Jones S, Enright AJ (ноябрь 2007 г.). «Геномный анализ транскриптов микроРНК человека» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (45): 17719–24. Bibcode : 2007pnas..10417719S . doi : 10.1073/pnas.0703890104 . PMC 2077053 . PMID 17965236 .
- ^ Йошикава М., Перагин А., Парк Мей, Поэтиг Р.С. (сентябрь 2005 г.). «Путь для биогенеза транс-действующих миРНК у арабидопсиса» . Гены и развитие . 19 (18): 2164–75. doi : 10.1101/gad.1352605 . PMC 1221887 . PMID 16131612 .
- ^ Amaral PP, Mattick JS (август 2008 г.). «Некодирующая РНК в разработке». Геном млекопитающих . 19 (7–8): 454–92. doi : 10.1007/s00335-008-9136-7 . PMID 18839252 . S2CID 206956408 .
- ^ Jump up to: а беременный в дюймовый Bienroth S, Keller W, Wahle E (февраль 1993 г.). «Сборка комплекса полиаденилирования РНК процесса» . Embo Journal . 12 (2): 585–94. doi : 10.1002/j.1460-2075.1993.tb05690.x . PMC 413241 . PMID 8440247 .
- ^ Jump up to: а беременный Liu D, Brockman JM, Dass B, Hutchins LN, Singh P, McCarrey JR, MacDonald CC, Graber JH (2006). «Систематические изменения в 3'-обрабатывающих сигналах мРНК во время сперматогенеза мыши» . Исследование нуклеиновых кислот . 35 (1): 234–46. doi : 10.1093/nar/gkl919 . PMC 1802579 . PMID 17158511 .
- ^ Lutz CS (октябрь 2008 г.). «Альтернативное полиаденилирование: поворот на 3' -формации мРНК». ACS Химическая биология . 3 (10): 609–17. doi : 10.1021/cb800138w . PMID 18817380 .
- ^ Jump up to: а беременный Beaudoing E, Freier S, Wyatt JR, Claverie JM, Gautheret D (июль 2000 г.). «Паттерны варианта использования сигналов полиаденилирования в генах человека» . Исследование генома . 10 (7): 1001–10. doi : 10.1101/gr.10.7.1001 . PMC 310884 . PMID 10899149 .
- ^ Браун К.М., Гилмартин Г.М. (декабрь 2003 г.). «Механизм регуляции пре-мРНК 3'-обработки с помощью фактора расщепления человека IM» . Молекулярная клетка . 12 (6): 1467–76. doi : 10.1016/s1097-2765 (03) 00453-2 . PMID 14690600 .
- ^ Ян Q, Гилмартин GM, Doublié S (июнь 2010 г.). «Структурная основа распознавания Угуа с помощью белка NUDIX CFI (M) 25 и последствия для регуляторной роли в обработке 3' -мРНК» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (22): 10062–7. Bibcode : 2010pnas..10710062Y . doi : 10.1073/pnas.1000848107 . PMC 2890493 . PMID 20479262 .
- ^ Ян Q, Косено М., Гилмартин Г.М., Doublié S (март 2011 г.). «Кристаллическая структура коэффициента расщепления человека CFI (M) 25/CFI (M) 68/РНК дает представление о распознавании поли (A) сайта и петле РНК» . Структура 19 (3): 368–77. doi : 10.1016/j.str.2010.12.021 . PMC 3056899 . PMID 21295486 .
- ^ Венкатараман К., Браун К.М., Гилмартин Г.М. (июнь 2005 г.). «Анализ неканонического поли (а) сайта выявляет трехсторонний механизм для распознавания поли (а) сайта позвоночных» . Гены и развитие . 19 (11): 1315–27. doi : 10.1101/gad.1298605 . PMC 1142555 . PMID 15937220 .
- ^ Jump up to: а беременный Millevoi S, Loulergue C, Dettwiler S, Karaa SZ, Keller W, Antoniou M, Vagner S (октябрь 2006 г.). «Взаимодействие между U2AF 65 и CF I (M) связывает сплайсинг и 3 'конечную обработку» . Embo Journal . 25 (20): 4854–64. doi : 10.1038/sj.emboj.7601331 . PMC 1618107 . PMID 17024186 .
- ^ Jump up to: а беременный в Shen Y, Ji G, Haas BJ, Wu X, Zheng J, Reese GJ, Li QQ (май 2008 г.). «Анализ уровня генома рисовых мРНК 3'-энд-сигналов и альтернативного полиаденилирования» . Исследование нуклеиновых кислот . 36 (9): 3150–61. doi : 10.1093/nar/gkn158 . PMC 2396415 . PMID 18411206 .
- ^ Glover-Cutter K, Kim S, Espinosa J, Bentley DL (январь 2008 г.). «РНК-полимераза II пауза и ассоциируется с факторами обработки пре-мРНК на обоих концах генов» . Природа структурная и молекулярная биология . 15 (1): 71–8. doi : 10.1038/nsmb1352 . PMC 2836588 . PMID 18157150 .
- ^ Молекулярная биология клетки, глава 6, «От ДНК до РНК». 4 -е издание. Alberts B, Johnson A, Lewis J, et al. Нью -Йорк: Гарлендская наука; 2002.
- ^ Stumpf G, Domdey H (ноябрь 1996 г.). «Зависимость до-мРНК-3'-энды дрожжей от 3'-энда от CFT1: гомолог последовательности коэффициента связывания AAUAAA млекопитающего». Наука . 274 (5292): 1517–20. Bibcode : 1996sci ... 274.1517s . doi : 10.1126/science.274.5292.1517 . PMID 8929410 . S2CID 34840144 .
- ^ Исели С., Стивенсон Б.Дж., Де Соуза С.Дж., Самайя Х.Б., Камарго А.А., Буэтоу К.Х., Страусберг Р.Л., Симпсон А.Дж., Бухер П., Чоннил С.В. (июль 2002 г.). «Большое расстояние неоднородности на 3 'концах человеческих мРНК» . Исследование генома . 12 (7): 1068–74. doi : 10.1101/gr.62002 . PMC 186619 . PMID 12097343 .
- ^ Balbo PB, Bohm A (сентябрь 2007 г.). «Механизм поли (а) полимеразы: структура фермента-MGATP-РНК-тройного комплекса и кинетический анализ» . Структура 15 (9): 1117–31. doi : 10.1016/j.str.2007.07.010 . PMC 2032019 . PMID 17850751 .
- ^ Viphakone N, Voisinet-Hakil F, Minvielle-Sebastia L (апрель 2008 г.). «Молекулярное рассечение МРНК поли (а) контроля длины хвоста у дрожжей» . Исследование нуклеиновых кислот . 36 (7): 2418–33. doi : 10.1093/nar/gkn080 . PMC 2367721 . PMID 18304944 .
- ^ Wahle E (февраль 1995 г.). «Поли (а) контроль длины хвоста вызвано прекращением обработчивого синтеза» . Журнал биологической химии . 270 (6): 2800–8. doi : 10.1074/jbc.270.6.2800 . PMID 7852352 .
- ^ Dichtl B, Blank D, Sadowski M, Hübner W, Weiser S, Keller W (август 2002 г.). «YHH1P/CFT1P непосредственно связывает поли (A) распознавание сайта и прекращение транскрипции RNA -полимеразы II» . Embo Journal . 21 (15): 4125–35. doi : 10.1093/emboj/cdf390 . PMC 126137 . PMID 12145212 .
- ^ Наг А., Нарсин К, Мартинсон Х.Г. (июль 2007 г.). «Поли (A) -зависимая транскрипционная пауза опосредуется CPSF, действующим на организм полимеразы». Природа структурная и молекулярная биология . 14 (7): 662–9. doi : 10.1038/nsmb1253 . PMID 17572685 . S2CID 5777074 .
- ^ Tefferi A, Wieben ED, Dewald GW, Whiteman DA, Bernard ME, Spelsberg TC (август 2002 г.). «Праймер по медицинской геномике, часть II: фоновые принципы и методы в молекулярной генетике». Майо -клиника . 77 (8): 785–808. doi : 10.4065/77.8.785 . PMID 12173714 . S2CID 2237085 .
- ^ Coller JM, Grey NK, MP Wickens (октябрь 1998 г.). «Стабилизация мРНК с помощью поли (а) связывающего белка не зависит от поли (а) и требует трансляции» . Гены и развитие . 12 (20): 3226–35. doi : 10.1101/gad.12.20.3226 . PMC 317214 . PMID 9784497 .
- ^ Jump up to: а беременный Сиддики Н., Мангус Д.А., Чанг Т.К., Палермино Дж.М., Шью А.Б., Геринг К (август 2007 г.). «Поли (а) нуклеаза взаимодействует с С-концевым доменом полиаденилат-связывающего белкового домена из поли (a)-связывающего белка» . Журнал биологической химии . 282 (34): 25067–75. doi : 10.1074/jbc.m701256200 . PMID 17595167 .
- ^ Vinciguerra P, Stutz F (июнь 2004 г.). «Экспорт мРНК: сборочная линия от генов до ядерных пор». Современное мнение в клеточной биологии . 16 (3): 285–92. doi : 10.1016/j.ceb.2004.03.013 . PMID 15145353 .
- ^ Грей Н.К., Коллер Дж. М., Диксон К.С., Викенс М (сентябрь 2000 г.). «Многочисленные порции поли (A) -связывающего белка стимулируют трансляцию in vivo» . Embo Journal . 19 (17): 4723–33. doi : 10.1093/emboj/19.17.4723 . PMC 302064 . PMID 10970864 .
- ^ Meaux S, Van Hoof A (июль 2006 г.). «Транскрипты дрожжей, расщепленные внутренним рибозимом, дают новое понимание роли крышки и поли (а) хвоста в трансляции и распаде мРНК» . РНК . 12 (7): 1323–37. doi : 10.1261/rna.46306 . PMC 1484436 . PMID 16714281 .
- ^ Каргаполова Y, Левин М., Лакнер К., Данквардт С. (июнь 2017 г.). «Интегрированная платформа SCLIP-AN для изучения РНК-белковых взаимодействий в биомедицинских исследованиях: идентификация CSTF2TAU в альтернативной обработке малых ядерных РНК» . Исследование нуклеиновых кислот . 45 (10): 6074–6086. doi : 10.1093/nar/gkx152 . PMC 5449641 . PMID 28334977 .
- ^ Jump up to: а беременный Meijer HA, Bushell M, Hill K, Gant TW, Willis AE, Jones P, De Moor CH (2007). «Новый метод поли (а) фракционирования выявляет большую популяцию мРНК с коротким поли (а) хвостом в клетках млекопитающих» . Исследование нуклеиновых кислот . 35 (19): E132. doi : 10.1093/nar/gkm830 . PMC 2095794 . PMID 17933768 .
- ^ Lehner B, Sanderson CM (июль 2004 г.). «Структура взаимодействия белка для деградации мРНК человека» . Исследование генома . 14 (7): 1315–23. doi : 10.1101/gr.2122004 . PMC 442147 . PMID 15231747 .
- ^ Wu L, Fan J, Belasco JG (март 2006 г.). «МикроРНК прямое быстрое детеенилирование мРНК» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (11): 4034–9. Bibcode : 2006pnas..103.4034W . doi : 10.1073/pnas.0510928103 . PMC 1449641 . PMID 16495412 .
- ^ Cui J, Sackton KL, Horner VL, Kumar Ke, Wolfner MF (апрель 2008 г.). «Wispy, гомолог Drosophila GLD-2, требуется во время оогенеза и активации яиц» . Генетика . 178 (4): 2017–29. doi : 10.1534/Genetics.107.084558 . PMC 2323793 . PMID 18430932 .
- ^ Тораби, Сейед-Фахреддин; Вайдья, Ананд Т.; Tycowski, Kazimierz T.; Degregorio, Suzanne J.; Ван, Джимин; Шу, Мей-ди; Стейц, Томас А.; Стейц, Джоан А. (2021-02-05). «Стабилизация РНК с помощью поли (а) хвоста 3'-энд связывающего кармана и других мод взаимодействия поли (a) -РНК» . Наука . 371 (6529): EABE6523. doi : 10.1126/science.abe6523 . ISSN 0036-8075 . PMC 9491362 . PMID 33414189 . S2CID 231195473 .
- ^ Wilusz CJ, Wormington M, Peltz SW (апрель 2001 г.). «Руководство по обороту мРНК на хвост». Природа обзор молекулярной клеточной биологии . 2 (4): 237–46. doi : 10.1038/35067025 . PMID 11283721 . S2CID 9734550 .
- ^ Юнг Ма, Лоренц Л., Рихтер Д.Д. (июнь 2006 г.). «Трансляционный контроль с помощью нейрогаидина, эукариотического фактора инициации 4E и CPEB -связывающего белка» . Молекулярная и клеточная биология . 26 (11): 4277–87. doi : 10.1128/mcb.02470-05 . PMC 1489097 . PMID 16705177 .
- ^ Сакурай Т., Сато М., Кимура М (ноябрь 2005 г.). «Разнообразные закономерности поли (A) удлинения хвоста и укорочения мышиных материнских мРНК от полностью выращенных ооцитов до 2-клеточных стадий эмбриона». Биохимическая и биофизическая исследовательская коммуникация . 336 (4): 1181–9. doi : 10.1016/j.bbrc.2005.08.250 . PMID 16169522 .
- ^ Тафт Р.А. (январь 2008 г.). «Добродетели и ограничения предамплантационного эмбриона мыши как модельной системы» . Териогенология . 69 (1): 10–6. doi : 10.1016/j.theriogenology.2007.09.032 . PMC 2239213 . PMID 18023855 .
- ^ Рихтер Д.Д. (июнь 2007 г.). «CPEB: жизнь в переводе». Тенденции в биохимических науках . 32 (6): 279–85. doi : 10.1016/j.tibs.2007.04.004 . PMID 17481902 .
- ^ Пике М., Лопес Дж. М., Фоссак С., Гиго Р., Мендес Р. (февраль 2008 г.). «Код комбинаторного кода для CPE-опосредованного трансляционного управления» . Клетка . 132 (3): 434–48. doi : 10.1016/j.cell.2007.12.038 . PMID 18267074 . S2CID 16092673 .
- ^ Бенуа П., Папин С., Квак Дж., Викенс М., Симонелиг М (июнь 2008 г.). «Полиполимеразы типа PAP- и GLD-2-типа необходимы последовательно в цитоплазматическом полиаденилировании и оогенезе у дрозофилы» . Разработка . 135 (11): 1969–79. doi : 10.1242/dev.021444 . PMC 9154023 . PMID 18434412 .
- ^ Tian B, Hu J, Zhang H, Lutz CS (2005). «Крупномасштабный анализ мРНК полиаденилирования генов человека и мыши» . Исследование нуклеиновых кислот . 33 (1): 201–12. doi : 10.1093/nar/gki158 . PMC 546146 . PMID 15647503 .
- ^ Danckwardt S, Hentze MW, Kulozik AE (февраль 2008 г.). «3 'END Обработка мРНК: молекулярные механизмы и последствия для здоровья и заболевания» . Embo Journal . 27 (3): 482–98. doi : 10.1038/sj.emboj.7601932 . PMC 2241648 . PMID 18256699 .
- ^ Jump up to: а беременный Тянь, бин; Мэнли, Джеймс Л. (2017). «Альтернативное полиаденилирование предшественников мРНК» . Природные обзоры. Молекулярная клеточная биология . 18 (1): 18–30. doi : 10.1038/nrm.2016.116 . ISSN 1471-0080 . PMC 5483950 . PMID 27677860 .
- ^ Чжан, Хайбо; Ли, Джу Юн; Tian, Bin (2005). «Предвзятое альтернативное полиаденилирование в тканях человека» . Биология генома . 6 (12): R100. doi : 10.1186/gb-2005-6-12-r100 . ISSN 1474-760x . PMC 1414089 . PMID 16356263 .
- ^ Smibert, Петр; Миура, Педро; Вестхольм, Якуб О.; Шенкер, Сол; Май, Джемма; Дафф, Майкл О.; Чжан, Дейу; Идс, Брайан Д.; Карлсон, Джо; Браун, Джеймс Б.; Эйсман, Роберт С. (2012). «Глобальные паттерны альтернативного полиаденилирования тканеспецифического в дрозофиле» . Сотовые отчеты . 1 (3): 277–289. doi : 10.1016/j.celrep.2012.01.001 . ISSN 2211-1247 . PMC 3368434 . PMID 22685694 .
- ^ Ли, Джу Юн; Джи, Чжэ; Tian, Bin (2008). «Филогенетический анализ сайтов полиаденилирования мРНК выявляет роль переносимых элементов в эволюции 3'-конца генов» . Исследование нуклеиновых кислот . 36 (17): 5581–5590. doi : 10.1093/nar/gkn540 . ISSN 1362-4962 . PMC 2553571 . PMID 18757892 .
- ^ Ogorodnikov A, Kargapolova Y, Danckwardt S (июнь 2016 г.). «Обработка и расширение транскриптома на 3 'мРНК 3' заканчиваются здоровьем и заболеванием: поиск правильного конца» . Pflügers Archiv . 468 (6): 993–1012. doi : 10.1007/s00424-016-1828-3 . PMC 4893057 . PMID 27220521 .
- ^ Сандберг Р., Нилсон младший, Сарма А., Шарп П.А., Бердж К.Б. (июнь 2008 г.). «Пролиферирующие клетки экспрессируют мРНК с укороченными 3' -нетранслируемыми областями и меньшим количеством сайтов микроРНК» . Наука . 320 (5883): 1643–7. Bibcode : 2008Sci ... 320.1643S . doi : 10.1126/science.1155390 . PMC 2587246 . PMID 18566288 .
- ^ Tili E, Michaille JJ, Calin GA (апрель 2008 г.). «Экспрессия и функция микро-РНК в иммунных клетках во время нормального или заболевания» . Международный журнал медицинских наук . 5 (2): 73–9. doi : 10.7150/ijms.5.73 . PMC 2288788 . PMID 18392144 .
- ^ Гош Т., Сони К., Скаррия В., Халимани М., Бхаттачарджи С., Пиллай Б (ноябрь 2008 г.). «МикроРНК-опосредованная активация альтернативно полиаденилированного варианта цитоплазматического гена мыши {бета} -актин» . Исследование нуклеиновых кислот . 36 (19): 6318–32. doi : 10.1093/nar/gkn624 . PMC 2577349 . PMID 18835850 .
- ^ Alt FW, Bothwell AL, Knapp M, Siden E, Mather E, Koshland M, Baltimore D (июнь 1980 г.). «Синтез секретируемых и мембранных иммуноглобулиновых тяжелых цепей MU направлен мРНК, которые различаются на их 3'-концах». Клетка . 20 (2): 293–301. doi : 10.1016/0092-8674 (80) 90615-7 . PMID 6771018 . S2CID 7448467 .
- ^ Tian B, Pan Z, Lee Jy (февраль 2007 г.). «Широко распространенные события полиаденилирования мРНК в интронах указывают на динамическое взаимодействие между полиаденилированием и сплайсингом» . Исследование генома . 17 (2): 156–65. doi : 10.1101/gr.5532707 . PMC 1781347 . PMID 17210931 .
- ^ Jump up to: а беременный Shell SA, Hesse C, Morris SM, Milcarek C (декабрь 2005 г.). «Повышенные уровни стимулирующего фактора расщепления 64 кДа (CSTF-64) в макрофагах, стимулированных липополисахаридами, влияют на экспрессию генов и вызывают альтернативный выбор поли (a) сайта» . Журнал биологической химии . 280 (48): 39950–61. doi : 10.1074/jbc.m508848200 . PMID 16207706 .
- ^ Ogorodnikov A, Levin M, Tattikota S, Tokalov S, Hoque M, Scherzinger D, Marini F, Poetsch A, Binder H, Macher-Göppinger S, Probst HC, Tian B, Schaefer M, Laxner KJ, Westermann F, Danckardt S ( Декабрь 2018 г.). «Транскриптом 3' -конечная организация от PCF11 связывает альтернативное полиаденилирование с формированием и дифференцировкой нейрональной нейробластомы» . Природная связь . 9 (1): 5331. Bibcode : 2018natco ... 9.5331o . doi : 10.1038/s41467-018-07580-5 . PMC 6294251 . PMID 30552333 .
- ^ Licatalosi DD, Mele A, Fak JJ, Ule J, Kayikci M, Chi SW, Clark TA, Schweitzer AC, Blume JE, Wang X, Darnell JC, Darnell RB (ноябрь 2008 г.). «Хиты зажигания дают общегеном понимание альтернативной обработки РНК мозга» . Природа . 456 (7221): 464–9. Bibcode : 2008natur.456..464L . doi : 10.1038/nature07488 . PMC 2597294 . PMID 18978773 .
- ^ Hall-Pogar T, Liang S, Hague LK, Lutz CS (июль 2007 г.). «Специфические транс-действия белки взаимодействуют с вспомогательными элементами полиаденилирования РНК в 3'-UTR COX-2» . РНК . 13 (7): 1103–15. doi : 10.1261/rna.577707 . PMC 1894925 . PMID 17507659 .
- ^ Danckwardt S, Kaufmann I, Gentzel M, Foerstner Ku, Gantzert AS, Gehring NH, Neu-Yilik G, Bork P, Keller W, Wilm M, Hentze MW, Kulozik Ae (июнь 2007 г.). «Факторы сплайсинга стимулируют полиаденилирование посредством использования при неканонических 3'-сигналах конечных формирования» . Embo Journal . 26 (11): 2658–69. doi : 10.1038/sj.emboj.7601699 . PMC 1888663 . PMID 17464285 .
- ^ Danckardt S, Gantzert AS, Macher-Goeppinger S, Probst HC, Gentzel M, Wilm M, Gröne HJ, Schirmacher P, Hentze MW, Kulozik AE (февраль 2011 г.). «P38 MAPK контролирует экспрессию протромбина с помощью регулируемой 3' -обработки РНК 3' -конечной обработки» . Молекулярная клетка . 41 (3): 298–310. doi : 10.1016/j.molcel.2010.12.032 . PMID 21292162 .
- ^ Wood AJ, Schulz R, Woodfine K, Koltowska K, Beechey CV, Peters J, Bourc'his D, Oakey RJ (май 2008). «Регуляция альтернативного полиаденилирования геномным импринтинг» . Гены и развитие . 22 (9): 1141–6. doi : 10.1101/gad.473408 . PMC 2335310 . PMID 18451104 .
- ^ Marini F, Scherzinger D, Danckwardt S (2021). «Атлас Trend-DB-A транскриптома динамического ландшафта альтернативного полиаденилирования» . Исследование нуклеиновых кислот . 49 (D1): D: 243 - D253. doi : 10.1093/nar/gkaa722 . PMC 7778938 . PMID 32976578 .
- ^ Рейниш К.М., Волин С.Л. (апрель 2007 г.). «Новые темы в некодирующем контроле качества РНК». Современное мнение в структурной биологии . 17 (2): 209–14. doi : 10.1016/j.sbi.2007.03.012 . PMID 17395456 .
- ^ Цзя Х., Ван Х, Лю Ф., Гунтер Юп, Шринивасан С., Андерсон Дж. Т., Янковский Е (июнь 2011 г.). «РНК -геликаза Mtr4p модулирует полиаденилирование в комплексе бродяги» . Клетка . 145 (6): 890–901. doi : 10.1016/j.cell.2011.05.010 . PMC 3115544 . PMID 21663793 .
- ^ Лакава Дж., Хаули Дж., Савину С., Петфальски Е., Томпсон Е., Жакер А., Толлерв и (июнь 2005 г.). «Деградация РНК экзосомой способствует ядерному полиаденилирующему комплексу» . Клетка . 121 (5): 713–24. doi : 10.1016/j.cell.2005.04.029 . PMID 15935758 . S2CID 14898055 .
- ^ Jump up to: а беременный Мартин Г., Келлер В. (ноябрь 2007 г.). «РНК-специфические рибонуклеотидилтрансферазы» . РНК . 13 (11): 1834–49. doi : 10.1261/rna.652807 . PMC 2040100 . PMID 17872511 .
- ^ Сломович С., Лауфер Д., Гейгер Д., Шустер Г. (2006). «Полиаденилирование рибосомной РНК в клетках человека» . Исследование нуклеиновых кислот . 34 (10): 2966–75. doi : 10.1093/nar/gkl357 . PMC 1474067 . PMID 16738135 .
- ^ Régnier P, Arraiano CM (март 2000 г.). «Разложение мРНК в бактериях: появление вездесущих особенностей». Биологии . 22 (3): 235–44. doi : 10.1002/(SICI) 1521-1878 (200003) 22: 3 <235 :: AID-bies5> 3.0.co; 2-2 . PMID 10684583 . S2CID 26109164 .
- ^ Jump up to: а беременный в Anantharaman V, Koonin EV, Aravind L (апрель 2002 г.). «Сравнительная геномика и эволюция белков, участвующих в метаболизме РНК» . Исследование нуклеиновых кислот . 30 (7): 1427–64. doi : 10.1093/nar/30.7.1427 . PMC 101826 . PMID 11917006 .
- ^ Jump up to: а беременный Сломович С., Портнай В., Ливеану В., Шустер Г. (2006). «Полиаденилирование РНК у прокариот и органеллов; разные хвосты рассказывают разные сказки». Критические обзоры в области наук о растениях . 25 (1): 65–77. Bibcode : 2006crvps..25 ... 65 с . doi : 10.1080/07352680500391337 . S2CID 86607431 .
- ^ Чанг, Чон Хо; Тонг, Лян (2012). «Митохондриальная поли (а) полимераза и полиаденилирование» . Biochimica et Biophysica Acta (BBA) - механизмы регуляции генов . 1819 (9–10): 992–997. doi : 10.1016/j.bbagrm.2011.10.012 . ISSN 0006-3002 . PMC 3307840 . PMID 22172994 .
- ^ Chang SA, Cozad M, Mackie GA, Jones GH (январь 2008 г.). «Кинетика полинуклеотидфосфорилазы: сравнение ферментов из Streptomyces и Escherichia coli и эффекты нуклеозидфосфатов» . Журнал бактериологии . 190 (1): 98–106. doi : 10.1128/jb.00327-07 . PMC 2223728 . PMID 17965156 .
- ^ Nagaike T, Suzuki T, Ueda T (апрель 2008 г.). «Полиаденилирование в митохондриях млекопитающих: понимание недавних исследований». Biochimica et Biophysica Acta (BBA) - механизмы регуляции генов . 1779 (4): 266–9. doi : 10.1016/j.bbagrm.2008.02.001 . PMID 18312863 .
- ^ Уолтер М., Килиан Дж., Кудла Дж (декабрь 2002 г.). «Активность PNPase определяет эффективность обработки мРНК 3'-конца, деградацию тРНК и степень полиаденилирования в хлоропластах» . Embo Journal . 21 (24): 6905–14. doi : 10.1093/emboj/cdf686 . PMC 139106 . PMID 12486011 .
- ^ Portnoy V, Schuster G (2006). «Полиаденилирование РНК и деградация в разных археи; роли экзосомы и РНКазы R» . Исследование нуклеиновых кислот . 34 (20): 5923–31. doi : 10.1093/nar/gkl763 . PMC 1635327 . PMID 17065466 .
- ^ Yehudai-Resheff S, Portnoy V, Yogev S, Adir N, Schuster G (сентябрь 2003 г.). «Анализ домена полинуклеотидфосфорилазы хлоропласта выявляет дискретные функции в деградации РНК, полиаденилировании и гомологии последовательности с белками экзосом» . Растительная ячейка . 15 (9): 2003–19. doi : 10.1105/tpc.013326 . PMC 181327 . PMID 12953107 .
- ^ Сломович С., Портнай В., Шустер Г. (2008). «Глава 24 Обнаружение и характеристика полиаденилированной РНК у эукарья, бактерий, археи и органелл». Оборот РНК у бактерий, археи и органеллов . Методы в фермере. Тол. 447. С. 501–20. doi : 10.1016/s0076-6879 (08) 02224-6 . ISBN 978-0-12-374377-0 Полем PMID 19161858 .
- ^ Portnoy V, Evguenieva-Hackenberg E, Klein F, Walter P, Lorentzen E, Klug G, Schuster G (декабрь 2005 г.). «Полиаденилирование РНК в археи: не наблюдается в Haloferax, в то время как экзосома полинуклеотидилат РНК в Sulfolobus» . Embo сообщает . 6 (12): 1188–93. doi : 10.1038/sj.embor.7400571 . PMC 1369208 . PMID 16282984 .
- ^ Portnoy V, Schuster G (июнь 2008 г.). «Mycoplasma wallisepticum как первая проанализированная бактерия, в которой РНК не полиаденилирована» . Письма микробиологии FEMS . 283 (1): 97–103. doi : 10.1111/j.1574-6968.2008.01157.x . PMID 18399989 .
- ^ Evguenieva-Hackenberg E, Roppelt V, Finsterseifer P, Klug G (декабрь 2008 г.). «RRP4 и CSL4 необходимы для эффективной деградации, но не для полиаденилирования синтетической и природной РНК архиальной экзосомой». Биохимия . 47 (50): 13158–68. doi : 10.1021/bi8012214 . PMID 19053279 .
- ^ Jump up to: а беременный Сломович С., Портнай В., Иегудай-Ришэфф С., Бронштейн Е., Шустер Г. (апрель 2008 г.). «Полинуклеотидфосфорилаза и архаальная экзосома в виде поли (a) -полимеразы». Biochimica et Biophysica Acta (BBA) - механизмы регуляции генов . 1779 (4): 247–55. doi : 10.1016/j.bbagrm.2007.12.004 . PMID 18177749 .
- ^ Пун, Лео Л.М.; Притлов, Дэвид С.; Фодор, Эрвин; Браунли, Джордж Г. (1 апреля 1999 г.). «Прямые доказательства того, что поли (а) хвоста мРНК вируса гриппа А синтезируется ретеративным копированием дорожки U в матрице РНК вирион» . Журнал вирусологии . 73 (4): 3473–3476. doi : 10.1128/JVI.73.4.3473-3476.1999 . PMC 104115 . PMID 10074205 .
- ^ Ву, Хун-Йи; Ке, Тинг-Юнг; Ляо, Вей-Ю; Чанг, Най-Юн (2013). «Регуляция коронавиральной поли (а) длины хвоста во время инфекции» . Plos один . 8 (7): E70548. BIBCODE : 2013PLOSO ... 870548W . doi : 10.1371/journal.pone.0070548 . PMC 3726627 . PMID 23923003 .
- ^ Нилман, Лида; Olshoorn, René Cl; Линторст, Huub JM; Бол, Джон Ф. (4 декабря 2001 г.). «Трансляция неполиаденилированной вирусной РНК усиливается путем связывания белка вирусного покрытия или полиаденилирования РНК» . Труды Национальной академии наук . 98 (25): 14286–14291. Bibcode : 2001pnas ... 9814286N . doi : 10.1073/pnas.251542798 . PMC 64674 . PMID 11717411 .
- ^ Чен, Джун-Хао; Чжан, Руи-хуа; Лин, Шао-Ли; Ли, Пенг-Фей; Лан, Цзин-Джинг; Песня, Ша-Ша; Гао, Джи-Мин; Ван, Ю; Се, Чжи-Джинг; Ли, Фу-Чанг; Цзян, Ши-Джин (2018). «Функциональная роль 3'-нетранслируемой области и поли (а) хвоста утка гепатита-вируса типа 1 типа 1 типа в репликации вируса и регуляции трансляции опосредованной IRES» . Границы в микробиологии . 9 : 2250. DOI : 10.3389/fmicb.2018.02250 . PMC 6167517 . PMID 30319572 .
- ^ «Ингибирование поли (a)-связывающего белка с помощью вируса ~ вируса» . viralzone.expasy.org .
- ^ Эдмондс, Мэри ; Абрамс, Ричард (апрель 1960 г.). «Полинуклеотидный биосинтез: образование последовательности аденилатных единиц из аденозина трихосфата ферментом из ядер тимуса» . Журнал биологической химии . 235 (4): 1142–1149. doi : 10.1016/s0021-9258 (18) 69494-3 .
- ^ Colgan DF, Manley JL (ноябрь 1997 г.). «Механизм и регуляция полиаденилирования мРНК» . Гены и развитие . 11 (21): 2755–66. doi : 10.1101/gad.11.21.2755 . PMID 9353246 .
- ^ Jump up to: а беременный Эдмондс, М. (2002). История полис -последовательностей: от формирования до факторов для функционирования . Прогресс в исследованиях нуклеиновых кислот и молекулярной биологии. Тол. 71. С. 285–389. doi : 10.1016/s0079-6603 (02) 71046-5 . ISBN 978-0-12-540071-8 Полем PMID 12102557 .
- ^ Эдмондс . Воган, MH; Наказато, Х. (1 июня 1971 года). «Последовательности полиадениловой кислоты в гетерогенной ядерной РНК и быстро меченной полирибосомной РНК клеток HeLa: возможные доказательства отношения предшественника» . Труды Национальной академии наук . 68 (6): 1336–1340. Bibcode : 1971pnas ... 68.1336e . doi : 10.1073/pnas.68.6.1336 . PMC 389184 . PMID 5288383 .
Дальнейшее чтение
[ редактировать ]- Danckwardt S, Hentze MW, Kulozik AE (февраль 2008 г.). «3 'END Обработка мРНК: молекулярные механизмы и последствия для здоровья и заболевания» . Embo Journal . 27 (3): 482–98. doi : 10.1038/sj.emboj.7601932 . PMC 2241648 . PMID 18256699 .
Внешние ссылки
[ редактировать ] СМИ, связанные с полиаденилированием в Wikimedia Commons
СМИ, связанные с полиаденилированием в Wikimedia Commons