Полиаденилирование

Полиаденилирование — это добавление поли (А)-хвоста к транскрипту РНК, обычно к информационной РНК (мРНК). Хвост поли(А) состоит из множества аденозинмонофосфатов ; другими словами, это участок РНК, содержащий только адениновые основания. У эукариот полиаденилирование является частью процесса производства зрелой мРНК для трансляции . У многих бактерий поли(А)-хвост способствует деградации мРНК. Таким образом, это является частью более широкого процесса экспрессии генов .
Процесс полиаденилирования начинается транскрипции гена с прекращения . Самый 3'- сегмент вновь созданной пре-мРНК сначала отщепляется набором белков ; эти белки затем синтезируют поли(А)-хвост на 3'-конце РНК. В некоторых генах эти белки добавляют поли(А)-хвост в одном из нескольких возможных сайтов. Следовательно, полиаденилирование может производить более одного транскрипта из одного гена ( альтернативное полиаденилирование ), аналогично альтернативному сплайсингу . [1]
Поли(А)-хвост важен для ядерного экспорта, трансляции и стабильности мРНК. Хвост со временем укорачивается, и, когда он становится достаточно коротким, мРНК ферментативно расщепляется. [2] Однако в некоторых типах клеток мРНК с короткими поли(А)-хвостами сохраняются для последующей активации путем повторного полиаденилирования в цитозоле. [3] Напротив, когда полиаденилирование происходит у бактерий, оно способствует деградации РНК. [4] Иногда это также относится к эукариотическим некодирующим РНК . [5] [6]
Молекулы мРНК как у прокариот, так и у эукариот имеют полиаденилированные 3'-концы, при этом прокариотические поли(А)-хвосты обычно короче и полиаденилировано меньше молекул мРНК. [7]
Общие сведения о РНК
[ редактировать ]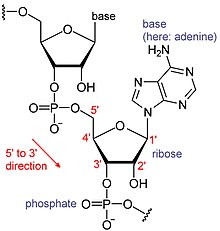
РНК представляют собой тип крупных биологических молекул, отдельные строительные блоки которых называются нуклеотидами. Название хвоста поли(А) (от хвоста полиадениловой кислоты) [8] отражает способ сокращения нуклеотидов РНК: буква обозначает основание, которое содержит нуклеотид (A — аденин , C — цитозин , G — гуанин и U — урацил ). РНК производятся ( транскрибируются ) из матрицы ДНК . По соглашению последовательности РНК записываются в направлении от 5’ к 3’. 5'-конец – это часть молекулы РНК, которая транскрибируется первой, а 3'-конец транскрибируется последней. На 3'-конце также находится поли(А)-хвост полиаденилированных РНК. [1] [9]
Информационная РНК (мРНК) — это РНК, имеющая кодирующую область, которая действует как матрица для синтеза белка ( трансляции ). Остальная часть мРНК, нетранслируемые области , регулирует активность мРНК. [10] Существует также множество РНК, которые не транслируются и называются некодирующими РНК. Как и нетранслируемые области, многие из этих некодирующих РНК выполняют регуляторную роль. [11]
Ядерное полиаденилирование
[ редактировать ]Функция
[ редактировать ]При ядерном полиаденилировании к РНК присоединяется поли(А)-хвост в конце транскрипции. Поли(А)-хвост мРНК защищает молекулу мРНК от ферментативной деградации в цитоплазме и способствует терминации транскрипции, экспорту мРНК из ядра и трансляции. [2] Почти все эукариотические мРНК полиаденилированы. [12] за исключением мРНК гистонов , зависящих от репликации животных . [13] Это единственные мРНК у эукариот, у которых отсутствует поли(А)-хвост, вместо этого оканчивающийся структурой « стебель-петля» , за которой следует богатая пуринами последовательность, называемая нижним элементом гистона, которая определяет место разрезания РНК так, чтобы 3'-конец мРНК гистонов образуется. [14]
Многие эукариотические некодирующие РНК всегда полиаденилированы в конце транскрипции. Существуют небольшие РНК, у которых поли(А)-хвост виден только в промежуточных формах, а не в зрелой РНК, поскольку концы удаляются во время процессинга, причем наиболее заметными из них являются микроРНК . [15] [16] Но для многих длинных некодирующих РНК – казалось бы, большой группы регуляторных РНК, в которую, например, входит РНК Xist , которая опосредует инактивацию Х-хромосомы – поли(А)-хвост является частью зрелой РНК. [17]
Механизм
[ редактировать ]| Участвующие белки: [12] [18] CPSF : фактор специфичности расщепления/полиаденилирования. |
Комплекс процессивного полиаденилирования в ядре эукариот воздействует на продукты РНК-полимеразы II , такие как мРНК-предшественники . Здесь мультибелковый комплекс (см. компоненты справа) [18] расщепляет большую часть 3'-конца вновь полученной РНК и полиаденилирует конец, образовавшийся в результате этого расщепления. Расщепление катализируется ферментом CPSF. [13] [18] и находится на 10–30 нуклеотидов ниже сайта связывания. [19] Этот сайт часто имеет сигнальную последовательность полиаденилирования на РНК AAUAAA, но существуют его варианты, которые более слабо связываются с CPSF . [18] [20] Два других белка добавляют специфичности связыванию с РНК: CstF и CFI. CstF связывается с богатой GU областью, расположенной дальше от сайта CPSF. [21] CFI распознает третий сайт РНК (набор последовательностей UGUAA у млекопитающих). [22] [23] [24] ) и может рекрутировать CPSF, даже если последовательность AAAUAAA отсутствует. [25] [26] Сигнал полиаденилирования – мотив последовательности, распознаваемый комплексом расщепления РНК – варьируется в зависимости от группы эукариот. Большинство сайтов полиаденилирования человека содержат последовательность AAAUAAA, [21] но эта последовательность менее распространена у растений и грибов. [27]
РНК обычно расщепляется до терминации транскрипции, поскольку CstF также связывается с РНК-полимеразой II. [28] Посредством плохо изученного механизма (по состоянию на 2002 год) он сигнализирует РНК-полимеразе II о выскальзывании из транскрипта. [29] В расщеплении также участвует белок CFII, хотя неизвестно, каким образом. [30] Сайт расщепления, связанный с сигналом полиаденилирования, может варьироваться примерно до 50 нуклеотидов. [31]
Когда РНК расщепляется, начинается полиаденилирование, катализируемое полиаденилат-полимеразой. Полиаденилат-полимераза строит поли(А)-хвост, добавляя звенья аденозинмонофосфата из аденозинтрифосфата к РНК, отщепляя пирофосфат . [32] Другой белок, PAB2, связывается с новым коротким поли(А)-хвостом и увеличивает сродство полиаденилат-полимеразы к РНК. Когда длина поли(А)-хвоста составляет примерно 250 нуклеотидов , фермент больше не может связываться с CPSF, и полиаденилирование прекращается, что определяет длину поли(А)-хвоста. [33] [34] CPSF контактирует с РНК-полимеразой II, позволяя ей сигнализировать полимеразе о прекращении транскрипции. [35] [36] Когда РНК-полимераза II достигает «последовательности терминации» (⁵'TTTATT 3 ' на матрице ДНК и ⁵'AAUAAA 3 ' на первичном транскрипте), подается сигнал об окончании транскрипции. [37] Механизм полиаденилирования также физически связан со сплайсосомой — комплексом, который удаляет интроны из РНК. [26]
Последующие эффекты
[ редактировать ]Поли(А)-хвост действует как сайт связывания поли(А)-связывающего белка . Поли(А)-связывающий белок способствует экспорту из ядра и трансляции, а также ингибирует деградацию. [38] Этот белок связывается с поли(А)-хвостом перед экспортом мРНК из ядра, а у дрожжей он также рекрутирует поли(А)-нуклеазу, фермент, который укорачивает поли(А)-хвост и обеспечивает экспорт мРНК. Поли(А)-связывающий белок экспортируется в цитоплазму вместе с РНК. мРНК, которые не экспортируются, разрушаются экзосомой . [39] [40] Поли(А)-связывающий белок также может связываться с несколькими белками, которые влияют на трансляцию, и, таким образом, рекрутировать их. [39] одним из них является фактор инициации -4G, который, в свою очередь, рекрутирует рибосомальную субъединицу 40S . [41] Однако поли(А)-хвост не требуется для трансляции всех мРНК. [42] Кроме того, поли(А)-хвост (олигоаденилирование) может определять судьбу молекул РНК, которые обычно не имеют поли(А)-хвостов (например, (маленькие) некодирующие (sn)РНК и т. д.), и тем самым индуцировать их РНК. разлагаться. [43]
Деаденилирование
[ редактировать ]В эукариотических соматических клетках поли(А)-хвосты большинства мРНК в цитоплазме постепенно становятся короче, а мРНК с более коротким поли(А)-хвостом меньше транслируются и быстрее деградируют. [44] Однако может пройти много часов, прежде чем мРНК деградирует. [45] Этот процесс деаденилирования и деградации может быть ускорен микроРНК, комплементарными 3'-нетранслируемой области мРНК. [46] В незрелых яйцеклетках мРНК с укороченными поли(А)-хвостами не деградируют, а вместо этого сохраняются и трансляционно неактивны. Эти короткохвостые мРНК активируются путем цитоплазматического полиаденилирования после оплодотворения, во время активации яйцеклетки . [47]
У животных поли(А) рибонуклеаза ( PARN ) может связываться с 5'-кэпом и удалять нуклеотиды из поли(А) хвоста. Уровень доступа к 5'-кэпу и поли(А)-хвосту важен для контроля скорости деградации мРНК. PARN деаденилируется меньше, если РНК связана факторами инициации 4E (на 5'-кэпе) и 4G (на поли(А)-хвосте), поэтому трансляция уменьшает деаденилирование. Скорость деаденилирования также может регулироваться РНК-связывающими белками. Кроме того, структуры тройной спирали РНК и мотивы РНК, такие как 3'-концевой связывающий карман поли(А)-хвоста, замедляют процесс деаденилирования и ингибируют удаление поли(А)-хвоста. [48] Как только поли(А)-хвост удаляется, декэпирующий комплекс удаляет 5'-кэп, что приводит к деградации РНК. Несколько других белков участвуют в деаденилировании в почкующихся дрожжах и клетках человека, в первую очередь комплекс CCR4-Not . [49]
Цитоплазматическое полиаденилирование
[ редактировать ]В цитозоле некоторых типов клеток животных, а именно в зародышевой линии , во время раннего эмбриогенеза и в постсинаптических участках нервных клеток наблюдается полиаденилирование . Это удлиняет поли(А)-хвост мРНК за счет укороченного поли(А)-хвоста, так что мРНК будет транслироваться . [44] [50] Эти укороченные поли(А)-хвосты часто имеют длину менее 20 нуклеотидов и удлиняются примерно до 80–150 нуклеотидов. [3]
У ранних эмбрионов мышей цитоплазматическое полиаденилирование материнских РНК из яйцеклетки позволяет клетке выживать и расти, даже если транскрипция не начинается до середины 2-клеточной стадии (4-клеточной стадии у человека). [51] [52] В мозге цитоплазматическое полиаденилирование активно во время обучения и может играть роль в долговременной потенциации , которая представляет собой усиление передачи сигнала от одной нервной клетки к другой в ответ на нервные импульсы и важна для обучения и формирования памяти. [3] [53]
Цитоплазматическое полиаденилирование требует РНК-связывающих белков CPSF и CPEB и может включать в себя другие РНК-связывающие белки, такие как Pumilio . [54] В зависимости от типа клеток полимеразой может быть тот же тип полиаденилат-полимеразы (PAP), который используется в ядерном процессе, или цитоплазматическая полимераза GLD-2 . [55]
Альтернативное полиаденилирование
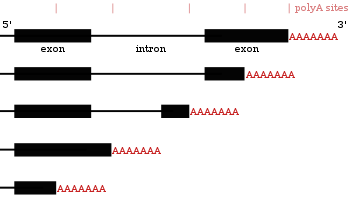
[ редактировать ]Многие гены, кодирующие белки, имеют более одного сайта полиаденилирования, поэтому ген может кодировать несколько мРНК, которые различаются по своему 3'-концу . [27] [56] [57] 3'-область транскрипта содержит множество сигналов полиаденилирования (PAS). Когда используются более проксимальные (ближе к 5'-концу) сайты PAS, это укорачивает длину 3'-нетранслируемой области (3'-UTR) транскрипта. [58] Исследования на людях и мухх показали тканеспецифическую APA. Нейрональные ткани предпочитают использование дистальной PAS, что приводит к более длинным 3'-UTR, а ткани семенников предпочитают проксимальную PAS, что приводит к более коротким 3'-UTR. [59] [60] Исследования показали, что существует корреляция между уровнем консервативности гена и его тенденцией к альтернативному полиаденилированию, при этом высококонсервативные гены демонстрируют больше APA. Точно так же высокоэкспрессированные гены следуют той же схеме. [61] Данные рибосеквенирования (секвенирование только мРНК внутри рибосом) показали, что изоформы мРНК с более короткими 3'-НТО с большей вероятностью будут транслироваться. [58]
Поскольку альтернативное полиаденилирование изменяет длину 3'-UTR , [62] он также может изменить то, какие сайты связывания доступны для микроРНК в 3'-UTR. [19] [63] МикроРНК имеют тенденцию подавлять трансляцию и способствовать деградации мРНК, с которыми они связываются, хотя существуют примеры микроРНК, которые стабилизируют транскрипты. [64] [65] Альтернативное полиаденилирование также может сократить кодирующую область, в результате чего мРНК будет кодировать другой белок. [66] [67] но это встречается гораздо реже, чем просто укорочение 3'-нетранслируемой области. [27]
На выбор сайта поли(А) могут влиять внеклеточные стимулы и зависит от экспрессии белков, участвующих в полиаденилировании. [68] [69] Например, экспрессия CstF-64 , субъединицы фактора, стимулирующего расщепление (CstF), увеличивается в макрофагах в ответ на липополисахариды (группа бактериальных соединений, запускающих иммунный ответ). Это приводит к отбору слабых поли(А)-сайтов и, следовательно, к более коротким транскриптам. Это удаляет регуляторные элементы в 3'-нетранслируемых областях мРНК для продуктов, связанных с защитой, таких как лизоцим и TNF-α . Эти мРНК имеют более длительный период полураспада и производят больше этих белков. [68] РНК-связывающие белки, отличные от тех, которые участвуют в механизме полиаденилирования, также могут влиять на использование сайта полиаденилирования. [70] [71] [72] [73] как и метилирование ДНК вблизи сигнала полиаденилирования. [74] Кроме того, на APA могут влиять многие другие компоненты, участвующие в транскрипции, сплайсинге или других механизмах, регулирующих биологию РНК. [75]
Маркировка деградации эукариот
[ редактировать ]Для многих некодирующих РНК , включая тРНК , рРНК , мяРНК и мяРНК , полиаденилирование является способом маркировки РНК для деградации, по крайней мере, у дрожжей . [76] Это полиаденилирование осуществляется в ядре с помощью комплекса TRAMP , который сохраняет хвост длиной около 4 нуклеотидов до 3'-конца. [77] [78] Затем РНК разрушается экзосомой . [79] Поли(А)-хвосты также были обнаружены на фрагментах рРНК человека, как в форме гомополимерных (только А), так и в форме гетерополимерных (в основном А) хвостов. [80]
В прокариотах и органеллах
[ редактировать ]
У многих бактерий как мРНК, так и некодирующие РНК могут быть полиаденилированы. Этот хвост поли(А) способствует деградации деградосомой , которая содержит два фермента, расщепляющих РНК: полинуклеотидфосфорилазу и РНКазу Е. Полинуклеотидфосфорилаза связывается с 3'-концом РНК, а удлинение 3'-конца, обеспечиваемое поли(А)-хвостом, позволяет ей связываться с РНК, вторичная структура которых в противном случае блокировала бы 3'-конец. Последовательные циклы полиаденилирования и деградации 3'-конца полинуклеотидфосфорилазой позволяют деградосоме преодолеть эти вторичные структуры. Поли(А)-хвост также может рекрутировать РНКазы, которые разрезают РНК на две части. [81] Эти бактериальные поли(А)-хвосты имеют длину около 30 нуклеотидов. [82]
В таких разных группах, как животные и трипаносомы , митохондрии содержат как стабилизирующие, так и дестабилизирующие поли(А) хвосты. Дестабилизирующее полиаденилирование нацелено как на мРНК, так и на некодирующие РНК. Длина поли(А)-хвостов в среднем составляет 43 нуклеотида. Стабилизирующие начинаются со стоп-кодона, и без них стоп-кодон (UAA) не является полным, поскольку геном кодирует только часть U или UA. В митохондриях растений наблюдается только дестабилизирующее полиаденилирование. Митохондриальное полиаденилирование никогда не наблюдалось ни у почкующихся, ни у делящихся дрожжей. [83] [84]
Хотя многие бактерии и митохондрии имеют полиаденилат-полимеразы, у них также есть другой тип полиаденилирования, осуществляемый самой полинуклеотидфосфорилазой . Этот фермент имеется у бактерий, [85] митохондрии, [86] пластиды [87] и как составная часть экзосомы архей (у тех архей , у которых есть экзосома ). [88] Он может синтезировать 3'-удлинение, в котором подавляющее большинство оснований представляют собой аденины. Как и у бактерий, полиаденилирование полинуклеотидфосфорилазой способствует деградации РНК в пластидах. [89] и, вероятно, также археи. [83]
Эволюция
[ редактировать ]Хотя полиаденилирование наблюдается почти у всех организмов, оно не является универсальным. [7] [90] Однако широкое распространение этой модификации и тот факт, что она присутствует в организмах из всех трех областей жизни, предполагают, что последний универсальный общий предок всех живых организмов, как предполагается, имел некоторую форму системы полиаденилирования. [82] Некоторые организмы не полиаденилируют мРНК, а это означает, что они утратили свои механизмы полиаденилирования в ходе эволюции. Хотя не известны примеры эукариот, у которых отсутствует полиаденилирование, мРНК бактерии Mycoplasma Gallisepticum и солеустойчивых архей Haloferax volcanii лишены этой модификации. [91] [92]
Самый древний полиаденилирующий фермент — полинуклеотидфосфорилаза . Этот фермент входит в состав как бактериальной деградосомы , так и архейной экзосомы . [93] два близкородственных комплекса, которые перерабатывают РНК в нуклеотиды. Этот фермент разрушает РНК, атакуя связь между большинством 3'-нуклеотидов фосфатом, разрывая дифосфатный нуклеотид. Эта реакция обратима, поэтому фермент также может удлинять РНК большим количеством нуклеотидов. Гетерополимерный хвост, добавленный полинуклеотидфосфорилазой, очень богат аденином. Выбор аденина, скорее всего, является результатом более высоких концентраций АДФ , чем других нуклеотидов, в результате использования АТФ в качестве энергетической валюты, что повышает вероятность его включения в этот хвост у ранних форм жизни. Было высказано предположение, что участие богатых аденином хвостов в деградации РНК спровоцировало более позднюю эволюцию полиаденилат-полимераз (ферментов, которые производят поли(А)-хвосты без других нуклеотидов в них). [94]
Полиаденилат-полимеразы не столь древние. Они отдельно эволюционировали как у бактерий, так и у эукариот из фермента, добавляющего CCA , который является ферментом, который достраивает 3'-концы тРНК . Его каталитический домен гомологичен домену других полимераз . [79] Предполагается, что горизонтальный перенос бактериального фермента, добавляющего CCA, к эукариотам позволил архейноподобному ферменту, добавляющему CCA, переключить функцию на поли(А)-полимеразу. [82] Некоторые линии, такие как археи и цианобактерии , так и не развили полиаденилат-полимеразу. [94]
Полиаденилатные хвосты наблюдаются у нескольких РНК-вирусов , включая грипп А , [95] Coronavirus , [96] вирус мозаики люцерны , [97] и утиный гепатит А. [98] Некоторые вирусы, такие как ВИЧ-1 и полиовирус , ингибируют поли-А-связывающий белок клетки ( PABPC1 ), чтобы усилить экспрессию своих собственных генов по сравнению с клеткой-хозяином. [99]
История
[ редактировать ]Поли(А)полимераза была впервые идентифицирована в 1960 году как ферментативная активность экстрактов клеточных ядер, способная полимеризовать АТФ, но не АДФ, в полиаденин. [100] [101] Хотя эта активность была обнаружена во многих типах клеток, ее функция не была известна до 1971 года, когда в мРНК были обнаружены последовательности поли(А). [102] [103] Сначала считалось, что единственной функцией этих последовательностей является защита 3'-конца РНК от нуклеаз, но позже была идентифицирована специфическая роль полиаденилирования в ядерном экспорте и трансляции. Полимеразы, ответственные за полиаденилирование, были впервые очищены и охарактеризованы в 1960-х и 1970-х годах, но большое количество вспомогательных белков, контролирующих этот процесс, было обнаружено только в начале 1990-х годов. [102]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Праудфут, Нью-Джерси, Фургер А., Дай М.Дж. (февраль 2002 г.). «Интеграция процессинга мРНК с транскрипцией» . Клетка . 108 (4): 501–12. дои : 10.1016/S0092-8674(02)00617-7 . ПМИД 11909521 . S2CID 478260 .
- ^ Jump up to: а б Гуханиеги Дж., Брюэр Дж. (март 2001 г.). «Регуляция стабильности мРНК в клетках млекопитающих» . Джин . 265 (1–2): 11–23. дои : 10.1016/S0378-1119(01)00350-X . ПМК 3340483 . ПМИД 11255003 .
- ^ Jump up to: а б с Рихтер Дж. Д. (июнь 1999 г.). «Цитоплазматическое полиаденилирование в развитии и за его пределами» . Обзоры микробиологии и молекулярной биологии . 63 (2): 446–56. дои : 10.1128/MMBR.63.2.446-456.1999 . ПМК 98972 . ПМИД 10357857 .
- ^ Стидж Д.А. (август 2000 г.). «Новые особенности распада мРНК у бактерий» . РНК . 6 (8): 1079–90. дои : 10.1017/S1355838200001023 . ПМК 1369983 . ПМИД 10943888 .
- ^ Чжуан Ю, Чжан Х, Линь С (июнь 2013 г.). «Полиаденилирование 18S рРНК в водорослях (1)». Журнал психологии . 49 (3): 570–9. Бибкод : 2013JPcgy..49..570Z . дои : 10.1111/jpy.12068 . ПМИД 27007045 . S2CID 19863143 .
- ^ Андерсон Дж. Т. (август 2005 г.). «Обмен РНК: неожиданные последствия слежки» . Современная биология . 15 (16): Р635-8. Бибкод : 2005CBio...15.R635A . дои : 10.1016/j.cub.2005.08.002 . ПМИД 16111937 . S2CID 19003617 .
- ^ Jump up to: а б Саркар Н. (июнь 1997 г.). «Полиаденилирование мРНК у прокариот». Ежегодный обзор биохимии . 66 (1): 173–97. doi : 10.1146/annurev.biochem.66.1.173 . ПМИД 9242905 .
- ^ Стивенс А. (1963). «Рибонуклеиновые кислоты-биосинтез и деградация». Ежегодный обзор биохимии . 32 : 15–42. дои : 10.1146/annurev.bi.32.070163.000311 . ПМИД 14140701 .
- ^ Ленинджер А.Л., Нельсон Д.Л., Кокс М.М., ред. (1993). Основы биохимии (2-е изд.). Нью-Йорк: Стоит. ISBN 978-0-87901-500-8 . [ нужна страница ]
- ^ Абаза I, Гебауэр Ф (март 2008 г.). «Торговая трансляция с помощью РНК-связывающих белков» . РНК . 14 (3): 404–9. дои : 10.1261/rna.848208 . ПМК 2248257 . ПМИД 18212021 .
- ^ Маттик Дж.С., Макунин И.В. (апрель 2006 г.). «Некодирующая РНК» . Молекулярная генетика человека . 15 Спецификация № 1 (90001): R17-29. дои : 10.1093/hmg/ddl046 . ПМИД 16651366 .
- ^ Jump up to: а б Хант А.Г., Сюй Р., Аддепалли Б., Рао С., Форбс К.П., Микс Л.Р., Син Д., Мо М., Чжао Х., Бандиопадьяй А., Дампанабойна Л., Марион А., Фон Ланкен С., Ли QQ (май 2008 г.). «Машина полиаденилирования мРНК арабидопсиса: комплексный анализ белок-белковых взаимодействий и профилирование экспрессии генов» . БМК Геномика . 9 :220. дои : 10.1186/1471-2164-9-220 . ПМК 2391170 . ПМИД 18479511 .
- ^ Jump up to: а б Давила Лопес М., Самуэльссон Т. (январь 2008 г.). «Ранняя эволюция процессинга 3'-конца мРНК гистонов» . РНК . 14 (1): 1–10. дои : 10.1261/rna.782308 . ПМК 2151031 . ПМИД 17998288 .
- ^ Марзлафф В.Ф., Гонгиди П., Вудс К.Р., Джин Дж., Малтаис Л.Дж. (ноябрь 2002 г.). «Гистоны человека и мыши, зависимые от репликации». Геномика . 80 (5): 487–98. дои : 10.1016/S0888-7543(02)96850-3 . ПМИД 12408966 .
- ^ Сайни Х.К., Гриффитс-Джонс С., Энрайт А.Дж. (ноябрь 2007 г.). «Геномный анализ транскриптов микроРНК человека» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (45): 17719–24. Бибкод : 2007PNAS..10417719S . дои : 10.1073/pnas.0703890104 . ПМК 2077053 . ПМИД 17965236 .
- ^ Ёсикава М., Перагин А., Пак М.Ю., Поетиг Р.С. (сентябрь 2005 г.). «Путь биогенеза транс-действующих миРНК у арабидопсиса» . Гены и развитие . 19 (18): 2164–75. дои : 10.1101/gad.1352605 . ПМЦ 1221887 . ПМИД 16131612 .
- ^ Амарал П.П., Мэттик Дж.С. (август 2008 г.). «Некодирующая РНК в развитии». Геном млекопитающих . 19 (7–8): 454–92. дои : 10.1007/s00335-008-9136-7 . ПМИД 18839252 . S2CID 206956408 .
- ^ Jump up to: а б с д Бьенрот С., Келлер В., Вале Э. (февраль 1993 г.). «Сборка комплекса полиаденилирования процессивной информационной РНК» . Журнал ЭМБО . 12 (2): 585–94. дои : 10.1002/j.1460-2075.1993.tb05690.x . ПМК 413241 . ПМИД 8440247 .
- ^ Jump up to: а б Лю Д., Брокман Дж. М., Дасс Б., Хатчинс Л. Н., Сингх П., МакКерри Дж. Р., Макдональд CC, Грабер Дж. Х. (2006). «Систематические изменения сигналов 3'-процессинга мРНК во время сперматогенеза у мышей» . Исследования нуклеиновых кислот . 35 (1): 234–46. дои : 10.1093/нар/gkl919 . ПМК 1802579 . ПМИД 17158511 .
- ^ Лутц CS (октябрь 2008 г.). «Альтернативное полиаденилирование: поворот в формировании 3'-конца мРНК». АКС Химическая биология . 3 (10): 609–17. дои : 10.1021/cb800138w . ПМИД 18817380 .
- ^ Jump up to: а б Бодуэн Э., Фрейер С., Вятт-младший, Клавери Дж.М., Готре Д. (июль 2000 г.). «Закономерности использования варианта сигнала полиаденилирования в генах человека» . Геномные исследования . 10 (7): 1001–10. дои : 10.1101/гр.10.7.1001 . ПМК 310884 . ПМИД 10899149 .
- ^ Браун К.М., Гилмартин Г.М. (декабрь 2003 г.). «Механизм регуляции процессинга 3' пре-мРНК человеческим фактором расщепления Im» . Молекулярная клетка . 12 (6): 1467–76. дои : 10.1016/S1097-2765(03)00453-2 . ПМИД 14690600 .
- ^ Ян Кью, Гилмартин ГМ, Дубли С (июнь 2010 г.). «Структурная основа распознавания UGUA белком Nudix CFI (m) 25 и значение регуляторной роли в 3'-процессинге мРНК» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (22): 10062–7. Бибкод : 2010PNAS..10710062Y . дои : 10.1073/pnas.1000848107 . ПМК 2890493 . ПМИД 20479262 .
- ^ Ян Кью, Косено М., Гилмартин ГМ, Дубли С. (март 2011 г.). «Кристаллическая структура комплекса человеческого фактора расщепления CFI(m)25/CFI(m)68/РНК дает представление о распознавании сайта поли(А) и зацикливании РНК» . Структура . 19 (3): 368–77. дои : 10.1016/j.str.2010.12.021 . ПМК 3056899 . ПМИД 21295486 .
- ^ Венкатараман К., Браун К.М., Гилмартин Г.М. (июнь 2005 г.). «Анализ неканонического поли(А)-сайта обнаруживает трехсторонний механизм распознавания поли(А)-сайта позвоночных» . Гены и развитие . 19 (11): 1315–27. дои : 10.1101/gad.1298605 . ПМЦ 1142555 . ПМИД 15937220 .
- ^ Jump up to: а б Миллевуа С., Лулерг С., Деттвайлер С., Караа С.З., Келлер В., Антониу М., Вагнер С. (октябрь 2006 г.). «Взаимодействие между U2AF 65 и CF I(m) связывает механизмы сращивания и обработки 3'-конца» . Журнал ЭМБО . 25 (20): 4854–64. дои : 10.1038/sj.emboj.7601331 . ПМК 1618107 . ПМИД 17024186 .
- ^ Jump up to: а б с Шен Ю, Джи Джи, Хаас Б.Дж., Ву X, Чжэн Дж., Риз Дж.Дж., Ли QQ (май 2008 г.). «Анализ на уровне генома сигналов обработки 3'-конца мРНК риса и альтернативного полиаденилирования» . Исследования нуклеиновых кислот . 36 (9): 3150–61. дои : 10.1093/нар/gkn158 . ПМК 2396415 . ПМИД 18411206 .
- ^ Гловер-Каттер К., Ким С., Эспиноза Дж., Бентли Д.Л. (январь 2008 г.). «РНК-полимераза II останавливается и связывается с факторами процессинга пре-мРНК на обоих концах генов» . Структурная и молекулярная биология природы . 15 (1): 71–8. дои : 10.1038/nsmb1352 . ПМЦ 2836588 . ПМИД 18157150 .
- ^ Молекулярная биология клетки, Глава 6, «От ДНК к РНК». 4-е издание. Альбертс Б., Джонсон А., Льюис Дж. и др. Нью-Йорк: Garland Science; 2002.
- ^ Стампф Г., Домдей Х. (ноябрь 1996 г.). «Зависимость процессинга 3'-конца пре-мРНК дрожжей от CFT1: гомолог последовательности фактора связывания AAUAAA млекопитающих». Наука . 274 (5292): 1517–20. Бибкод : 1996Sci...274.1517S . дои : 10.1126/science.274.5292.1517 . ПМИД 8929410 . S2CID 34840144 .
- ^ Исели С., Стивенсон Б.Дж., де Соуза С.Дж., Самайя Х.Б., Камарго А.А., Бютов К.Х., Штраусберг Р.Л., Симпсон А.Дж., Бучер П., Джонджинил К.В. (июль 2002 г.). «Дальняя гетерогенность на 3'-концах мРНК человека» . Геномные исследования . 12 (7): 1068–74. дои : 10.1101/гр.62002 . ЧВК 186619 . ПМИД 12097343 .
- ^ Бальбо П.Б., Бом А. (сентябрь 2007 г.). «Механизм поли(А)-полимеразы: структура тройного комплекса фермент-MgATP-РНК и кинетический анализ» . Структура . 15 (9): 1117–31. дои : 10.1016/j.str.2007.07.010 . ПМК 2032019 . ПМИД 17850751 .
- ^ Випхаконе Н., Вуазине-Хакил Ф., Минвьель-Себастия Л. (апрель 2008 г.). «Молекулярное рассечение контроля длины хвоста поли(А) мРНК у дрожжей» . Исследования нуклеиновых кислот . 36 (7): 2418–33. дои : 10.1093/нар/gkn080 . ПМК 2367721 . ПМИД 18304944 .
- ^ Вале Э. (февраль 1995 г.). «Контроль длины хвоста поли(А) обусловлен прекращением процессивного синтеза» . Журнал биологической химии . 270 (6): 2800–8. дои : 10.1074/jbc.270.6.2800 . ПМИД 7852352 .
- ^ Дихтл Б., Бланк Д., Садовски М., Хюбнер В., Вайзер С., Келлер В. (август 2002 г.). «Yhh1p/Cft1p напрямую связывает распознавание сайта поли(А) и терминацию транскрипции РНК-полимеразы II» . Журнал ЭМБО . 21 (15): 4125–35. дои : 10.1093/emboj/cdf390 . ПМК 126137 . ПМИД 12145212 .
- ^ Наг А., Нарсин К., Мартинсон Х.Г. (июль 2007 г.). «Поли(А)-зависимая пауза транскрипции опосредуется CPSF, действующим на тело полимеразы». Структурная и молекулярная биология природы . 14 (7): 662–9. дои : 10.1038/nsmb1253 . ПМИД 17572685 . S2CID 5777074 .
- ^ Теффери А., Вибен Э.Д., Девальд Г.В., Уайтман Д.А., Бернард М.Е., Спелсберг Т.К. (август 2002 г.). «Букварь по медицинской геномике, часть II: Основные принципы и методы молекулярной генетики». Труды клиники Мэйо . 77 (8): 785–808. дои : 10.4065/77.8.785 . ПМИД 12173714 . S2CID 2237085 .
- ^ Коллер Дж. М., Грей Н. К., член парламента Уикенса (октябрь 1998 г.). «Стабилизация мРНК поли(А)-связывающим белком не зависит от поли(А) и требует трансляции» . Гены и развитие . 12 (20): 3226–35. дои : 10.1101/gad.12.20.3226 . ПМК 317214 . ПМИД 9784497 .
- ^ Jump up to: а б Сиддики Н., Мангус Д.А., Чанг Т.К., Палермино Дж.М., Шью А.Б., Геринг К. (август 2007 г.). «Поли(А)-нуклеаза взаимодействует с С-концевым доменом полиаденилат-связывающего белка, доменом поли(А)-связывающего белка» . Журнал биологической химии . 282 (34): 25067–75. дои : 10.1074/jbc.M701256200 . ПМИД 17595167 .
- ^ Винчигерра П., Штутц Ф. (июнь 2004 г.). «Экспорт мРНК: конвейер от генов к ядерным порам». Современное мнение в области клеточной биологии . 16 (3): 285–92. дои : 10.1016/j.ceb.2004.03.013 . ПМИД 15145353 .
- ^ Грей Н.К., Коллер Дж.М., Диксон К.С., Викенс М. (сентябрь 2000 г.). «Множественные части поли(А)-связывающего белка стимулируют трансляцию in vivo» . Журнал ЭМБО . 19 (17): 4723–33. дои : 10.1093/emboj/19.17.4723 . ПМК 302064 . ПМИД 10970864 .
- ^ Мо С., Ван Хуф А. (июль 2006 г.). «Дрожжевые транскрипты, расщепленные внутренним рибозимом, дают новое представление о роли кэпа и поли(А)-хвоста в трансляции и распаде мРНК» . РНК . 12 (7): 1323–37. дои : 10.1261/rna.46306 . ПМЦ 1484436 . ПМИД 16714281 .
- ^ Каргаполова Ю., Левин М., Лакнер К., Данквардт С. (июнь 2017 г.). «sCLIP-интегрированная платформа для изучения РНК-белковых интерактомов в биомедицинских исследованиях: идентификация CSTF2tau при альтернативном процессинге малых ядерных РНК» . Исследования нуклеиновых кислот . 45 (10): 6074–6086. дои : 10.1093/нар/gkx152 . ПМЦ 5449641 . ПМИД 28334977 .
- ^ Jump up to: а б Мейер Х.А., Бушелл М., Хилл К., Гант Т.В., Уиллис А.Е., Джонс П., де Мур CH (2007). «Новый метод фракционирования поли(А) выявил большую популяцию мРНК с коротким поли(А)-хвостом в клетках млекопитающих» . Исследования нуклеиновых кислот . 35 (19): е132. дои : 10.1093/нар/gkm830 . ПМК 2095794 . ПМИД 17933768 .
- ^ Ленер Б., Сандерсон К.М. (июль 2004 г.). «Схема взаимодействия белков для деградации мРНК человека» . Геномные исследования . 14 (7): 1315–23. дои : 10.1101/гр.2122004 . ПМК 442147 . ПМИД 15231747 .
- ^ Ву Л, Фан Дж, Беласко Дж. Г. (март 2006 г.). «МикроРНК направляют быстрое деаденилирование мРНК» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (11): 4034–9. Бибкод : 2006PNAS..103.4034W . дои : 10.1073/pnas.0510928103 . ПМЦ 1449641 . ПМИД 16495412 .
- ^ Куи Дж., Сактон К.Л., Хорнер В.Л., Кумар К.Е., Вольфнер М.Ф. (апрель 2008 г.). «Виспи, гомолог GLD-2 дрозофилы, необходим во время оогенеза и активации яиц» . Генетика . 178 (4): 2017–29. дои : 10.1534/genetics.107.084558 . ПМЦ 2323793 . ПМИД 18430932 .
- ^ Тораби, Сейед-Фахреддин; Вайдья, Ананд Т.; Тыковский, Казимеж Т.; ДеГрегорио, Сюзанна Дж.; Ван, Чимин; Шу, Мэй-Ди; Стейтц, Томас А.; Стейтц, Джоан А. (05 февраля 2021 г.). «Стабилизация РНК с помощью кармана связывания 3'-конца поли(А)-хвоста и других способов взаимодействия поли(А)-РНК» . Наука . 371 (6529): eabe6523. дои : 10.1126/science.abe6523 . ISSN 0036-8075 . ПМЦ 9491362 . ПМИД 33414189 . S2CID 231195473 .
- ^ Вилуш С.Дж., Уормингтон М., Пельц С.В. (апрель 2001 г.). «Полное руководство по обороту мРНК». Nature Reviews Молекулярно-клеточная биология . 2 (4): 237–46. дои : 10.1038/35067025 . ПМИД 11283721 . S2CID 9734550 .
- ^ Юнг М.Ю., Лоренц Л., Рихтер Дж.Д. (июнь 2006 г.). «Контроль трансляции с помощью нейрогидина, эукариотического фактора инициации 4E и связывающего белка CPEB» . Молекулярная и клеточная биология . 26 (11): 4277–87. дои : 10.1128/MCB.02470-05 . ПМК 1489097 . ПМИД 16705177 .
- ^ Сакураи Т., Сато М., Кимура М. (ноябрь 2005 г.). «Различные закономерности удлинения хвоста поли(А) и укорочения мышиных материнских мРНК от полностью выросшего ооцита до стадии двухклеточного эмбриона». Связь с биохимическими и биофизическими исследованиями . 336 (4): 1181–9. дои : 10.1016/j.bbrc.2005.08.250 . ПМИД 16169522 .
- ^ Тафт РА (январь 2008 г.). «Достоинства и ограничения преимплантационного эмбриона мыши как модельной системы» . Териогенология . 69 (1): 10–6. doi : 10.1016/j.theriogenology.2007.09.032 . ПМК 2239213 . ПМИД 18023855 .
- ^ Рихтер Дж. Д. (июнь 2007 г.). «CPEB: жизнь в переводе». Тенденции биохимических наук . 32 (6): 279–85. дои : 10.1016/j.tibs.2007.04.004 . ПМИД 17481902 .
- ^ Пике М., Лопес Х.М., Фуассак С., Гиго Р., Мендес Р. (февраль 2008 г.). «Комбинаторный код для управления трансляцией, опосредованного CPE» . Клетка . 132 (3): 434–48. дои : 10.1016/j.cell.2007.12.038 . ПМИД 18267074 . S2CID 16092673 .
- ^ Бенуа П., Папен С., Квак Дж.Э., Викенс М., Симонелиг М. (июнь 2008 г.). «Поли(А)-полимеразы PAP- и GLD-2-типа последовательно необходимы для цитоплазматического полиаденилирования и оогенеза у дрозофилы» . Разработка . 135 (11): 1969–79. дои : 10.1242/dev.021444 . ПМЦ 9154023 . ПМИД 18434412 .
- ^ Тянь Б., Ху Дж., Чжан Х., Лутц К.С. (2005). «Масштабный анализ полиаденилирования мРНК генов человека и мыши» . Исследования нуклеиновых кислот . 33 (1): 201–12. дои : 10.1093/nar/gki158 . ПМК 546146 . ПМИД 15647503 .
- ^ Данквардт С., Хентце М.В., Кулозик А.Е. (февраль 2008 г.). «Обработка 3'-конца мРНК: молекулярные механизмы и последствия для здоровья и болезней» . Журнал ЭМБО . 27 (3): 482–98. дои : 10.1038/sj.emboj.7601932 . ПМК 2241648 . ПМИД 18256699 .
- ^ Jump up to: а б Тиан, Бин; Мэнли, Джеймс Л. (2017). «Альтернативное полиаденилирование предшественников мРНК» . Обзоры природы. Молекулярно-клеточная биология . 18 (1): 18–30. дои : 10.1038/номер.2016.116 . ISSN 1471-0080 . ПМК 5483950 . ПМИД 27677860 .
- ^ Чжан, Хайбо; Ли, Джу Юн; Тиан, Бин (2005). «Предвзятое альтернативное полиаденилирование в тканях человека» . Геномная биология . 6 (12): 100 рандов. дои : 10.1186/gb-2005-6-12-r100 . ISSN 1474-760X . ПМЦ 1414089 . ПМИД 16356263 .
- ^ Смайберт, Питер; Миура, Педро; Вестхольм, Якуб О.; Шенкер, Сол; Мэй, Джемма; Дафф, Майкл О.; Чжан, Дайю; Идс, Брайан Д.; Карлсон, Джо; Браун, Джеймс Б.; Эйсман, Роберт С. (2012). «Глобальные закономерности тканеспецифического альтернативного полиаденилирования у дрозофилы» . Отчеты по ячейкам . 1 (3): 277–289. дои : 10.1016/j.celrep.2012.01.001 . ISSN 2211-1247 . ПМЦ 3368434 . ПМИД 22685694 .
- ^ Ли, Джу Юн; Цзи, Чжэ; Тиан, Бин (2008). «Филогенетический анализ сайтов полиаденилирования мРНК выявил роль мобильных элементов в эволюции 3'-конца генов» . Исследования нуклеиновых кислот . 36 (17): 5581–5590. дои : 10.1093/нар/gkn540 . ISSN 1362-4962 . ПМЦ 2553571 . ПМИД 18757892 .
- ^ Огородников А, Каргаполова Ю, Данквардт С (июнь 2016 г.). «Процессинг и расширение транскриптома на 3'-конце мРНК в норме и при болезни: поиск правильного конца» . Архив Пфлюгерса . 468 (6): 993–1012. дои : 10.1007/s00424-016-1828-3 . ПМЦ 4893057 . ПМИД 27220521 .
- ^ Сэндберг Р., Нилсон Дж.Р., Сарма А., Шарп П.А., Бердж CB (июнь 2008 г.). «Пролиферирующие клетки экспрессируют мРНК с укороченными 3'-нетранслируемыми областями и меньшим количеством целевых сайтов микроРНК» . Наука . 320 (5883): 1643–7. Бибкод : 2008Sci...320.1643S . дои : 10.1126/science.1155390 . ПМЦ 2587246 . ПМИД 18566288 .
- ^ Тили Э., Мишель Дж.Дж., Кэлин Г.А. (апрель 2008 г.). «Экспрессия и функция микроРНК в иммунных клетках в нормальном или болезненном состоянии» . Международный журнал медицинских наук . 5 (2): 73–9. дои : 10.7150/ijms.5.73 . ПМК 2288788 . ПМИД 18392144 .
- ^ Гош Т., Сони К., Скариа В., Халимани М., Бхаттачарджи С., Пиллаи Б. (ноябрь 2008 г.). «МикроРНК-опосредованная активация альтернативно полиаденилированного варианта цитоплазматического гена {бета}-актина мыши» . Исследования нуклеиновых кислот . 36 (19): 6318–32. дои : 10.1093/нар/gkn624 . ПМЦ 2577349 . ПМИД 18835850 .
- ^ Альт Ф.В., Ботвелл А.Л., Кнапп М., Сайден Э., Мазер Э., Кошланд М., Балтимор Д. (июнь 1980 г.). «Синтез секретируемых и мембраносвязанных тяжелых цепей иммуноглобулина мю управляется мРНК, которые различаются на своих 3'-концах». Клетка . 20 (2): 293–301. дои : 10.1016/0092-8674(80)90615-7 . ПМИД 6771018 . S2CID 7448467 .
- ^ Тиан Б., Пан З., Ли Дж. Ю. (февраль 2007 г.). «Широко распространенные события полиаденилирования мРНК в интронах указывают на динамическое взаимодействие между полиаденилированием и сплайсингом» . Геномные исследования . 17 (2): 156–65. дои : 10.1101/гр.5532707 . ПМЦ 1781347 . ПМИД 17210931 .
- ^ Jump up to: а б Shell SA, Hesse C, Моррис С.М., Милкарек C (декабрь 2005 г.). «Повышенные уровни фактора, стимулирующего расщепление 64-кДа (CstF-64) в стимулированных липополисахаридами макрофагах, влияют на экспрессию генов и индуцируют выбор альтернативного поли(А)-сайта» . Журнал биологической химии . 280 (48): 39950–61. дои : 10.1074/jbc.M508848200 . ПМИД 16207706 .
- ^ Огородников А, Левин М, Таттикота С, Токалов С, Хоке М, Шерзингер Д, Марини Ф, Поэтш А, Биндер Х, Мачер-Гёппингер С, Пробст ХК, Тиан Б, Шефер М, Лакнер КДж, Вестерманн Ф, Данквардт С ( декабрь 2018 г.). «Организация 3'-конца транскриптома с помощью PCF11 связывает альтернативное полиаденилирование с образованием и нейрональной дифференцировкой нейробластомы» . Природные коммуникации . 9 (1): 5331. Бибкод : 2018NatCo...9.5331O . дои : 10.1038/s41467-018-07580-5 . ПМК 6294251 . ПМИД 30552333 .
- ^ Ликаталоси Д.Д., Меле А., Фак Дж.Дж., Уле Дж., Кайикчи М., Чи С.В., Кларк Т.А., Швейцер А.С., Блюм Дж.Э., Ван Х, Дарнелл Дж.К., Дарнелл Р.Б. (ноябрь 2008 г.). «HITS-CLIP дает общегеномное представление об альтернативной обработке РНК в мозге» . Природа . 456 (7221): 464–9. Бибкод : 2008Natur.456..464L . дои : 10.1038/nature07488 . ПМК 2597294 . ПМИД 18978773 .
- ^ Холл-Погар Т., Лян С., Гаага Л.К., Лутц К.С. (июль 2007 г.). «Специфические транс-действующие белки взаимодействуют со вспомогательными элементами полиаденилирования РНК в 3'-UTR ЦОГ-2» . РНК . 13 (7): 1103–15. дои : 10.1261/rna.577707 . ЧВК 1894925 . ПМИД 17507659 .
- ^ Данквардт С., Кауфманн И., Генцель М., Ферстнер К.У., Ганцерт А.С., Геринг Н.Х., Ной-Йилик Г., Борк П., Келлер В., Вильм М., Хентце М.В., Кулозик А.Е. (июнь 2007 г.). «Факторы сплайсинга стимулируют полиаденилирование посредством USE по неканоническим сигналам образования 3'-конца» . Журнал ЭМБО . 26 (11): 2658–69. дои : 10.1038/sj.emboj.7601699 . ПМЦ 1888663 . ПМИД 17464285 .
- ^ Данквардт С., Ганцерт А.С., Мачер-Геппингер С., Пробст Х.К., Генцель М., Вильм М., Грёне Х.Дж., Ширмахер П., Хентце М.В., Кулозик А.Е. (февраль 2011 г.). «p38 MAPK контролирует экспрессию протромбина посредством регулируемого процессинга 3'-конца РНК» . Молекулярная клетка . 41 (3): 298–310. doi : 10.1016/j.molcel.2010.12.032 . ПМИД 21292162 .
- ^ Вуд А.Дж., Шульц Р., Вудфайн К., Колтовска К., Бичи К.В., Питерс Дж., Бурчис Д., Оки Р.Дж. (май 2008 г.). «Регуляция альтернативного полиаденилирования посредством геномного импринтинга» . Гены и развитие . 22 (9): 1141–6. дои : 10.1101/gad.473408 . ПМЦ 2335310 . ПМИД 18451104 .
- ^ Марини Ф, Шерзингер Д, Данквардт С (2021). «TREND-DB - полный транскриптомный атлас динамического ландшафта альтернативного полиаденилирования» . Исследования нуклеиновых кислот . 49 (Д1): Д:243–Д253. дои : 10.1093/nar/gkaa722 . ПМЦ 7778938 . ПМИД 32976578 .
- ^ Райниш К.М., Волин С.Л. (апрель 2007 г.). «Новые темы контроля качества некодирующих РНК». Современное мнение в области структурной биологии . 17 (2): 209–14. дои : 10.1016/j.sbi.2007.03.012 . ПМИД 17395456 .
- ^ Цзя Х, Ван Х, Лю Ф, Гюнтер УП, Шринивасан С, Андерсон Дж. Т., Янковски Э (июнь 2011 г.). «РНК-хеликаза Mtr4p модулирует полиаденилирование в комплексе TRAMP» . Клетка . 145 (6): 890–901. дои : 10.1016/j.cell.2011.05.010 . ПМК 3115544 . ПМИД 21663793 .
- ^ ЛаКава Дж., Хаусли Дж., Савану С., Петфальски Е., Томпсон Е., Жакье А., Толлерви Д. (июнь 2005 г.). «Деградации РНК экзосомой способствует ядерный комплекс полиаденилирования» . Клетка . 121 (5): 713–24. дои : 10.1016/j.cell.2005.04.029 . ПМИД 15935758 . S2CID 14898055 .
- ^ Jump up to: а б Мартин Дж., Келлер В. (ноябрь 2007 г.). «РНК-специфические рибонуклеотидилтрансферазы» . РНК . 13 (11): 1834–49. дои : 10.1261/rna.652807 . ПМК 2040100 . ПМИД 17872511 .
- ^ Сломович С., Лауфер Д., Гейгер Д., Шустер Г. (2006). «Полиаденилирование рибосомальной РНК в клетках человека» . Исследования нуклеиновых кислот . 34 (10): 2966–75. дои : 10.1093/нар/gkl357 . ПМК 1474067 . ПМИД 16738135 .
- ^ Ренье П., Аррайано CM (март 2000 г.). «Деградация мРНК у бактерий: появление повсеместных особенностей». Биоэссе . 22 (3): 235–44. doi : 10.1002/(SICI)1521-1878(200003)22:3<235::AID-BIES5>3.0.CO;2-2 . ПМИД 10684583 . S2CID 26109164 .
- ^ Jump up to: а б с Анантараман В., Кунин Е.В., Аравинд Л. (апрель 2002 г.). «Сравнительная геномика и эволюция белков, участвующих в метаболизме РНК» . Исследования нуклеиновых кислот . 30 (7): 1427–64. дои : 10.1093/нар/30.7.1427 . ПМК 101826 . ПМИД 11917006 .
- ^ Jump up to: а б Сломович С, Портной В, Ливану В, Шустер Г (2006). «Полиаденилирование РНК в прокариотах и органеллах; разные хвосты рассказывают разные истории». Критические обзоры по наукам о растениях . 25 (1): 65–77. Бибкод : 2006CRvPS..25...65S . дои : 10.1080/07352680500391337 . S2CID 86607431 .
- ^ Чанг, Чон Хо; Тонг, Лян (2012). «Митохондриальная поли(А)-полимераза и полиаденилирование» . Biochimica et Biophysica Acta (BBA) - Механизмы регуляции генов . 1819 (9–10): 992–997. дои : 10.1016/j.bbagrm.2011.10.012 . ISSN 0006-3002 . ПМК 3307840 . ПМИД 22172994 .
- ^ Чанг С.А., Козад М., Маки Г.А., Джонс Г.Х. (январь 2008 г.). «Кинетика полинуклеотидфосфорилазы: сравнение ферментов Streptomyces и Escherichia coli и эффектов нуклеозиддифосфатов» . Журнал бактериологии . 190 (1): 98–106. дои : 10.1128/JB.00327-07 . ПМК 2223728 . ПМИД 17965156 .
- ^ Нагаике Т., Сузуки Т., Уэда Т. (апрель 2008 г.). «Полиаденилирование в митохондриях млекопитающих: результаты недавних исследований». Biochimica et Biophysica Acta (BBA) - Механизмы регуляции генов . 1779 (4): 266–9. дои : 10.1016/j.bbagrm.2008.02.001 . ПМИД 18312863 .
- ^ Уолтер М., Килиан Дж., Кудла Дж. (декабрь 2002 г.). «Активность ПНПазы определяет эффективность процессинга 3'-конца мРНК, деградацию тРНК и степень полиаденилирования в хлоропластах» . Журнал ЭМБО . 21 (24): 6905–14. дои : 10.1093/emboj/cdf686 . ПМК 139106 . ПМИД 12486011 .
- ^ Портной В., Шустер Г. (2006). «Полиаденилирование и деградация РНК у разных архей; роль экзосомы и РНКазы R» . Исследования нуклеиновых кислот . 34 (20): 5923–31. дои : 10.1093/нар/gkl763 . ПМЦ 1635327 . ПМИД 17065466 .
- ^ Иегудай-Решефф С., Портной В., Йогев С., Адир Н., Шустер Г. (сентябрь 2003 г.). «Анализ доменов полинуклеотидфосфорилазы хлоропластов выявляет дискретные функции в деградации РНК, полиаденилировании и гомологии последовательностей с белками экзосом» . Растительная клетка . 15 (9): 2003–19. дои : 10.1105/tpc.013326 . ПМК 181327 . ПМИД 12953107 .
- ^ Сломович С., Портной В., Шустер Г. (2008). «Глава 24. Обнаружение и характеристика полиаденилированной РНК у эукариев, бактерий, архей и органелл». Оборот РНК в бактериях, археях и органеллах . Методы энзимологии. Том. 447. стр. 501–20. дои : 10.1016/S0076-6879(08)02224-6 . ISBN 978-0-12-374377-0 . ПМИД 19161858 .
- ^ Портной В., Евгеньева-Хакенберг Е., Кляйн Ф., Вальтер П., Лоренцен Е., Клюг Г., Шустер Г. (декабрь 2005 г.). «Полиаденилирование РНК у архей: не наблюдается у Haloferax, в то время как экзосома полинуклеотидилирует РНК у Sulfolobus» . Отчеты ЭМБО . 6 (12): 1188–93. дои : 10.1038/sj.embor.7400571 . ПМЦ 1369208 . ПМИД 16282984 .
- ^ Портной В., Шустер Г. (июнь 2008 г.). «Mycoplasma Gallisepticum как первая проанализированная бактерия, у которой РНК не полиаденилирована» . Письма FEMS по микробиологии . 283 (1): 97–103. дои : 10.1111/j.1574-6968.2008.01157.x . ПМИД 18399989 .
- ^ Евгеньева-Хакенберг Е., Роппельт В., Финстерзайфер П., Клюг Г. (декабрь 2008 г.). «Rrp4 и Csl4 необходимы для эффективной деградации, но не для полиаденилирования синтетической и природной РНК экзосомой архей». Биохимия . 47 (50): 13158–68. дои : 10.1021/bi8012214 . ПМИД 19053279 .
- ^ Jump up to: а б Сломович С., Портной В., Иегудай-Решефф С., Бронштейн Е., Шустер Г. (апрель 2008 г.). «Полинуклеотидфосфорилаза и экзосома архей как поли(А)-полимеразы». Biochimica et Biophysica Acta (BBA) - Механизмы регуляции генов . 1779 (4): 247–55. дои : 10.1016/j.bbagrm.2007.12.004 . ПМИД 18177749 .
- ^ Пун, Лео Л.М.; Притлав, Дэвид С.; Фодор, Эрвин; Браунли, Джордж Г. (1 апреля 1999 г.). «Прямое доказательство того, что поли(А)-хвост мРНК вируса гриппа А синтезируется путем многократного копирования U-трека в матрице РНК вириона» . Журнал вирусологии . 73 (4): 3473–3476. doi : 10.1128/JVI.73.4.3473-3476.1999 . ПМЦ 104115 . ПМИД 10074205 .
- ^ Ву, Хун-И; Кэ, Тин-Юнг; Ляо, Вэй-Ю; Чанг, Най-Юн (2013). «Регулирование длины хвоста поли(А) коронавирусного вируса во время инфекции» . ПЛОС ОДИН . 8 (7): е70548. Бибкод : 2013PLoSO...870548W . дои : 10.1371/journal.pone.0070548 . ПМЦ 3726627 . ПМИД 23923003 .
- ^ Нилеман, Лида; Олстхорн, Рене CL; Линторст, Хууб Дж. М.; Бол, Джон Ф. (4 декабря 2001 г.). «Трансляция неполиаденилированной вирусной РНК усиливается за счет связывания белка вирусной оболочки или полиаденилирования РНК» . Труды Национальной академии наук . 98 (25): 14286–14291. Бибкод : 2001PNAS...9814286N . дои : 10.1073/pnas.251542798 . ПМК 64674 . ПМИД 11717411 .
- ^ Чен, Цзюнь-Хао; Чжан, Жуй-Хуа; Линь, Шао-Ли; Ли, Пэн-Фей; Лан, Цзин-Цзин; Песня, Ша-Ша; Гао, Цзи-Мин; Ван, Ю; Се, Чжи-Цзин; Ли, Фу-Чанг; Цзян, Ши-Цзинь (2018). «Функциональная роль 3'-нетранслируемой области и поли(А)-хвоста вируса утиного гепатита А типа 1 в репликации вируса и регуляции IRES-опосредованной трансляции» . Границы микробиологии . 9 : 2250. дои : 10.3389/fmicb.2018.02250 . ПМК 6167517 . ПМИД 30319572 .
- ^ «Ингибирование поли(А)-связывающего белка хозяина вирусом ~ ViralZone» . www.viralzone.expasy.org .
- ^ Эдмондс, Мэри ; Абрамс, Ричард (апрель 1960 г.). «Биосинтез полинуклеотидов: образование последовательности аденилатных единиц из аденозинтрифосфата ферментом из ядер тимуса» . Журнал биологической химии . 235 (4): 1142–1149. дои : 10.1016/S0021-9258(18)69494-3 .
- ^ Колган Д.Ф., Мэнли Дж.Л. (ноябрь 1997 г.). «Механизм и регуляция полиаденилирования мРНК» . Гены и развитие . 11 (21): 2755–66. дои : 10.1101/gad.11.21.2755 . ПМИД 9353246 .
- ^ Jump up to: а б Эдмондс, М. (2002). История поли-А-последовательностей: от формирования до факторов и функционирования . Прогресс в исследованиях нуклеиновых кислот и молекулярной биологии. Том. 71. С. 285–389. дои : 10.1016/S0079-6603(02)71046-5 . ISBN 978-0-12-540071-8 . ПМИД 12102557 .
- ^ Эдмондс, М .; Воган, Миннесота; Наказато, Х. (1 июня 1971 г.). «Последовательности полиадениловой кислоты в гетерогенной ядерной РНК и быстро меченной полирибосомальной РНК клеток HeLa: возможные доказательства связи предшественников» . Труды Национальной академии наук . 68 (6): 1336–1340. Бибкод : 1971PNAS...68.1336E . дои : 10.1073/pnas.68.6.1336 . ПМК 389184 . ПМИД 5288383 .
Дальнейшее чтение
[ редактировать ]- Данквардт С., Хентце М.В., Кулозик А.Е. (февраль 2008 г.). «Обработка 3'-конца мРНК: молекулярные механизмы и последствия для здоровья и болезней» . Журнал ЭМБО . 27 (3): 482–98. дои : 10.1038/sj.emboj.7601932 . ПМК 2241648 . ПМИД 18256699 .
Внешние ссылки
[ редактировать ] СМИ, связанные с полиаденилированием, на Викискладе?
СМИ, связанные с полиаденилированием, на Викискладе?