Цис-регуляторный элемент
Цис -регуляторные элементы ( CRE ) или цис- модули ( CRM ) представляют собой области некодирующей ДНК , которые регулируют транскрипцию соседних генов регуляторные . CREs являются жизненно важными компонентами генетических регуляторных сетей , которые, в свою очередь, контролируют морфогенез , развитие анатомии и другие аспекты эмбрионального развития , изучаемые в эволюционной биологии развития .
CRE находятся вблизи генов, которые они регулируют. CRE обычно регулируют транскрипцию генов путем связывания с факторами транскрипции . Один фактор транскрипции может связываться со многими CRE и, следовательно, контролировать экспрессию многих генов ( плейотропия ). Латинский означает «по эту сторону», то есть на той же молекуле ДНК , префикс цис что и транскрибируемый ген(ы).
CRM представляют собой участки ДНК , обычно длиной 100–1000 пар оснований ДНК. [ 1 ] где ряд факторов транскрипции могут связывать и регулировать экспрессию близлежащих генов , а также регулировать скорость их транскрипции. Их обозначают как цис, потому что они обычно расположены на той же цепи ДНК, что и гены, которые они контролируют, в отличие от транс , что относится к воздействию на гены, не расположенные на той же цепи или дальше, например, на факторы транскрипции. [ 1 ] Один цис -регуляторный элемент может регулировать несколько генов. [ 2 ] и наоборот, один ген может иметь несколько цис -регуляторных модулей. [ 3 ] Цис -регуляторные модули выполняют свою функцию путем интеграции активных факторов транскрипции и связанных с ними кофакторов в определенное время и в определенном месте клетки, где эта информация считывается и выдается выходной сигнал. [ 4 ]
CRE часто, но не всегда, располагаются выше сайта транскрипции. CRE контрастируют с трансрегуляторными элементами (TRE) . TRE кодируют факторы транскрипции. [ нужна ссылка ]
Обзор
[ редактировать ]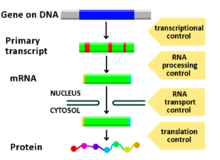
Геном . организма содержит от нескольких сотен до тысяч различных генов, каждый из которых кодирует один или несколько продуктов По многочисленным причинам, включая организационное обеспечение, сохранение энергии и создание фенотипической вариативности, важно, чтобы гены экспрессировались только тогда, когда они необходимы. Самый эффективный способ регулирования экспрессии генов в организме — на уровне транскрипции. CRE контролируют транскрипцию, действуя рядом или внутри гена. Наиболее хорошо охарактеризованными типами CRE являются энхансеры и промоторы . Оба эти элемента последовательности представляют собой структурные области ДНК, которые служат регуляторами транскрипции . [ нужна ссылка ]
Цис -регуляторные модули являются одним из нескольких типов функциональных регуляторных элементов . Регуляторные элементы представляют собой сайты связывания факторов транскрипции, которые участвуют в регуляции генов. [ 1 ] Цис -регуляторные модули выполняют большой объем обработки информации о развитии. [ 1 ] Цис -регуляторные модули представляют собой неслучайные кластеры в указанном сайте-мишени, которые содержат сайты связывания факторов транскрипции. [ 1 ]
Первоначальное определение представляло цис-регуляторные модули как усилители цис-действующей ДНК, которые увеличивали скорость транскрипции со связанного промотора . [ 4 ] Однако это определение было изменено и теперь определяет цис -регуляторные модули как последовательность ДНК с сайтами связывания факторов транскрипции, которые сгруппированы в модульные структуры, включая, помимо прочего, области толок-контроля, промоторы, энхансеры, сайленсеры, элементы пограничного контроля и другие. модуляторы. [ 4 ]
Цис -регуляторные модули можно разделить на три класса; энхансеры , которые положительно регулируют экспрессию генов; [ 1 ] изоляторы , которые действуют опосредованно, взаимодействуя с другими близлежащими цис -регуляторными модулями; и [ 1 ] глушители , которые отключают экспрессию генов. [ 1 ]
Конструкция цис -регуляторных модулей такова, что входными данными служат факторы транскрипции и эпигенетические модификации , а выходным сигналом модуля является команда, подаваемая транскрипционному аппарату, который, в свою очередь, определяет скорость транскрипции генов, включена она или нет. выключенный . [ 1 ] Существует два типа входных факторов транскрипционных факторов: те, которые определяют, когда целевой ген должен экспрессироваться, и те, которые служат функциональными драйверами , которые вступают в игру только в определенных ситуациях во время развития. [ 1 ] Эти входные данные могут поступать из разных моментов времени, могут представлять разные сигнальные лиганды или могут поступать из разных доменов или линий клеток. Однако многое еще остается неизвестным. [ нужна ссылка ]
Кроме того, регуляция структуры хроматина и ядерной организации также играет роль в определении и контроле функции цис-регуляторных модулей. [ 4 ] Таким образом, функции генной регуляции (GRF) обеспечивают уникальную характеристику цис-регуляторного модуля (CRM), связывая концентрации транскрипционных факторов (вход) с активностью промотора (выход). Задача состоит в том, чтобы предсказать GRF. Эта задача до сих пор остается нерешенной. В общем, функции генной регуляции не используют булевую логику . [ 2 ] хотя в некоторых случаях аппроксимация булевой логики все же очень полезна. [ нужна ссылка ]
Предположение булевой логики
[ редактировать ]В рамках булевой логики принципы, определяющие работу этих модулей, включают в себя конструкцию модуля, определяющую регулирующую функцию. Что касается развития, эти модули могут давать как положительные, так и отрицательные результаты. Выходные данные каждого модуля являются продуктом различных операций, выполненных над ним. Общие операции включают в себя логический элемент ИЛИ – эта конструкция указывает, что выходной сигнал будет выдан, когда подан любой входной сигнал [3], и логический элемент И – в этой конструкции необходимы два разных регулирующих фактора, чтобы гарантировать положительный результат на выходе. [ 1 ] «Тумблеры» - такая конструкция возникает, когда сигнальный лиганд отсутствует, а фактор транскрипции присутствует; этот фактор транскрипции в конечном итоге действует как доминантный репрессор. Однако как только сигнальный лиганд присутствует, роль фактора транскрипции как репрессора устраняется и может происходить транскрипция. [ 1 ]
Могут происходить и другие логические логические операции, такие как последовательность специфичных репрессоров транскрипции, которые, когда они связываются с цис -регуляторным модулем, приводят к нулевому выходному сигналу. Кроме того, помимо влияния различных логических операций, на выходные данные «цис»-регулирующего модуля также будут влиять предшествующие события. [ 1 ] 4) Цис -регуляторные модули должны взаимодействовать с другими регуляторными элементами. По большей части, даже при наличии функционального перекрытия между цис -регуляторными модулями гена, входы и выходы модулей, как правило, не совпадают. [ 1 ]
Хотя предположение о булевой логике важно для системной биологии , подробные исследования показывают, что в целом логика регуляции генов не является булевой. [ 2 ] Это означает, например, что в случае цис -регуляторного модуля, регулируемого двумя факторами транскрипции, экспериментально определенные функции генной регуляции не могут быть описаны 16 возможными булевыми функциями двух переменных. Для решения этой проблемы были предложены небулевы расширения логики регулирования генов. [ 2 ]
Классификация
[ редактировать ]Цис -регуляторные модули можно охарактеризовать по обработке информации, которую они кодируют, и организации сайтов связывания их транскрипционных факторов. Кроме того, цис -регуляторные модули характеризуются также тем, как они влияют на вероятность, долю и скорость транскрипции. [ 4 ] Высококооперативные и скоординированные цис- регуляторные модули классифицируются как энхансосомы . [ 4 ] Архитектура и расположение сайтов связывания транскрипционных факторов имеют решающее значение, поскольку нарушение расположения может свести на нет эту функцию. [ 4 ] Функциональные гибкие цис -регуляторные модули называются рекламными щитами. Их транскрипционный результат представляет собой эффект суммирования связанных факторов транскрипции. [ 4 ] Энхансеры влияют на вероятность активации гена, но практически не влияют на скорость. [ 4 ] Модель двоичного ответа действует как переключатель включения/выключения транскрипции. Эта модель увеличит или уменьшит количество клеток, транскрибирующих ген, но не повлияет на скорость транскрипции. [ 4 ] Модель реостатического ответа описывает цис-регуляторные модули как регуляторы скорости инициации транскрипции ассоциированного с ним гена. [ 4 ]
Промоутер
[ редактировать ]Промоторы представляют собой CRE, состоящие из относительно коротких последовательностей ДНК, которые включают сайт, где инициируется транскрипция, и область, расположенную примерно на 35 п.н. выше или ниже сайта инициации (п.н.). [ 5 ] У эукариот промоторы обычно состоят из следующих четырех компонентов: ТАТА-бокс , TFIIB сайт узнавания , инициатор и нижестоящий основной элемент промотора . [ 5 ] Было обнаружено, что один ген может содержать несколько промоторных сайтов. [ 6 ] Чтобы инициировать транскрипцию нижестоящего гена, множество ДНК-связывающих белков, называемых факторами транскрипции (ТФ), должны последовательно связываться с этой областью. [ 5 ] Только после того, как эта область будет связана с соответствующим набором ТФ и в правильном порядке, РНК-полимераза сможет связать и начать транскрипцию гена.
Усилители
[ редактировать ]Энхансеры — это CRE, которые влияют (усиливают) транскрипцию генов на одной и той же молекуле ДНК и могут находиться выше, ниже, внутри интронов или даже относительно далеко от гена, который они регулируют. Множественные энхансеры могут действовать скоординировано, регулируя транскрипцию одного гена. [ 7 ] Ряд проектов полногеномного секвенирования показал, что энхансеры часто транскрибируются в длинную некодирующую РНК (днРНК) или энхансерную РНК (еРНК), изменения уровней которых часто коррелируют с изменениями мРНК целевого гена. [ 8 ]
Глушители
[ редактировать ]Сайленсеры — это CRE, которые могут связывать факторы регуляции транскрипции (белки), называемые репрессорами , тем самым предотвращая транскрипцию гена. Термин «глушитель» также может относиться к участку в 3'-нетранслируемой области информационной РНК, которая связывает белки, подавляющие трансляцию этой молекулы мРНК, но это использование отличается от его использования при описании CRE. [ нужна ссылка ]
Операторы
[ редактировать ]Операторы — это CRE у прокариот и некоторых эукариот, которые существуют внутри оперонов , где они могут связывать белки, называемые репрессорами , чтобы влиять на транскрипцию. [ нужна ссылка ]
Эволюционная роль
[ редактировать ]CRE играют важную эволюционную роль. Кодирующие области генов часто хорошо консервативны среди организмов; тем не менее, разные организмы демонстрируют заметное фенотипическое разнообразие. Было обнаружено, что полиморфизмы , возникающие в некодирующих последовательностях, оказывают глубокое влияние на фенотип, изменяя экспрессию генов . [ 7 ] Мутации, возникающие внутри CRE, могут генерировать вариативность экспрессии за счет изменения способа связывания TF. Более плотное или более слабое связывание регуляторных белков приведет к усилению или понижению регуляции транскрипции.
Цис -регуляторный модуль в регуляторной сети генов
[ редактировать ]Функция генной регуляторной сети зависит от архитектуры узлов , функция которых зависит от множества цис -регуляторных модулей. [ 1 ] Расположение цис -регуляторных модулей может предоставить достаточно информации для создания пространственных и временных паттернов экспрессии генов. [ 1 ] Во время развития каждый домен, где каждый домен представляет собой различные пространственные области эмбриона, экспрессии генов будет находиться под контролем разных цис -регуляторных модулей. [ 1 ] Конструкция регуляторных модулей помогает создавать петли обратной связи , прямой связи и перекрестного регулирования. [ 9 ]
Способ действия
[ редактировать ]Цис -регуляторные модули могут регулировать свои гены-мишени на больших расстояниях. Было предложено несколько моделей для описания того, как эти модули могут взаимодействовать с промотором целевого гена. [ 4 ] К ним относятся модель сканирования ДНК, модель петли последовательности ДНК и модель облегченного отслеживания. В модели сканирования ДНК комплекс транскрипционного фактора и кофактора образуется в цис -регуляторном модуле и затем продолжает двигаться вдоль последовательности ДНК, пока не найдет промотор целевого гена. [ 4 ] В модели с петлей транскрипционный фактор связывается с цис -регуляторным модулем, который затем вызывает закольцовывание последовательности ДНК и обеспечивает взаимодействие с промотором целевого гена. Комплекс транскрипционного фактора- цис -регуляторного модуля вызывает медленное закольцовывание последовательности ДНК по направлению к целевому промотору и образует стабильную петлеобразную конфигурацию. [ 4 ] Модель упрощенного отслеживания объединяет части двух предыдущих моделей.
Идентификация и вычислительное прогнозирование
[ редактировать ]Помимо экспериментального определения CRM, существуют различные биоинформатические алгоритмы для их прогнозирования. Большинство алгоритмов пытаются искать значимые комбинации сайтов связывания транскрипционных факторов ( сайтов связывания ДНК ) в промоторных последовательностях совместно экспрессируемых генов. [ 10 ] Более продвинутые методы сочетают поиск значимых мотивов с корреляцией в наборах данных по экспрессии генов между факторами транскрипции и генами-мишенями. [ 11 ] Оба метода реализованы, например, в ModuleMaster . Другие программы, созданные для идентификации и прогнозирования цис -регуляторных модулей, включают:
НАСЕКОМОЕ 2.0 [ 12 ] представляет собой веб-сервер, позволяющий осуществлять поиск цис-регуляторных модулей по всему геному. Программа опирается на определение строгих ограничений среди сайтов связывания факторов транскрипции (TFBS), составляющих модуль, чтобы снизить уровень ложных срабатываний. INSECT спроектирован так, чтобы быть удобным для пользователя, поскольку он позволяет автоматически извлекать последовательности, а также несколько визуализаций и ссылок на сторонние инструменты, чтобы помочь пользователям находить те экземпляры, которые с большей вероятностью являются настоящими регулирующими сайтами. Алгоритм INSECT 2.0 был ранее опубликован, а алгоритм и теория, лежащие в его основе, объяснены в [ 13 ]
Стабб использует скрытые модели Маркова для идентификации статистически значимых кластеров комбинаций факторов транскрипции. Он также использует второй родственный геном для повышения точности прогнозирования модели. [ 14 ]
Байесовские сети используют алгоритм, который объединяет прогнозы сайтов и данные тканеспецифической экспрессии факторов транскрипции и интересующих генов-мишеней. Эта модель также использует деревья регрессии, чтобы отобразить взаимосвязь между идентифицированным цис -регуляторным модулем и возможным набором связывающих факторов транскрипции. [ 15 ]
CRÈME исследует кластеры сайтов-мишеней на предмет интересующих факторов транскрипции. Эта программа использует базу данных подтвержденных сайтов связывания факторов транскрипции, которые были аннотированы по всему геному человека . поиска К набору данных применяется алгоритм для выявления возможных комбинаций факторов транскрипции, которые имеют сайты связывания, близкие к промотору интересующего набора генов. Затем возможные цис-регуляторные модули статистически анализируются, и значимые комбинации представляются графически. [ 16 ]
Активные цис -регуляторные модули в геномной последовательности идентифицировать сложно. Проблемы с идентификацией возникают потому, что часто ученые сталкиваются с небольшим набором известных факторов транскрипции, что затрудняет идентификацию статистически значимых кластеров сайтов связывания факторов транскрипции. [ 14 ] Кроме того, высокая стоимость ограничивает использование больших массивов мозаики всего генома . [ 15 ]

Примеры
[ редактировать ]Примером цис-действующей регуляторной последовательности является оператор lac-оперона . Эта последовательность ДНК связана с lac-репрессором , который, в свою очередь, предотвращает транскрипцию соседних генов на той же молекуле ДНК. Таким образом, считается, что оператор lac «действует в цис» на регуляцию близлежащих генов. Сам оператор либо не кодирует какой- белок или РНК .
Напротив, трансрегуляторные элементы представляют собой диффузионные факторы, обычно белки, которые могут изменять экспрессию генов, далеких от гена, который был первоначально транскрибирован для их создания. Например, фактор транскрипции , который регулирует ген на хромосоме 6, сам мог быть транскрибирован из гена на хромосоме 11 . Термин трансрегуляторный образован от латинского корня trans , что означает «напротив».
Существуют цис-регуляторные и трансрегуляторные элементы. Цис-регуляторные элементы часто являются сайтами связывания одного или нескольких транс-действующих факторов.
Подводя итог, можно сказать, что цис-регуляторные элементы присутствуют на той же молекуле ДНК, что и ген, который они регулируют, тогда как трансрегуляторные элементы могут регулировать гены, далекие от гена, из которого они были транскрибированы.
Примеры в РНК
[ редактировать ]| Тип | Сокр. | Функция | Распределение | Ссылка. |
|---|---|---|---|---|
| Элемент сдвига рамки | Регулирует использование альтернативной рамки с помощью информационных РНК | Археи , бактерии , эукариоты , РНК-вирусы. | [ 17 ] [ 18 ] [ 19 ] | |
| Внутренний сайт входа в рибосому | ИРЭС | Инициирует трансляцию в середине информационной РНК. | РНК-вирусы , эукариоты | [ 20 ] |
| Железный ответный элемент | ИРЭ | Регулирует экспрессию генов, связанных с железом | Эукариоты | [ 21 ] |
| Лидерный пептид | Регулирует транскрипцию связанных генов и/или оперонов | Бактерии | [ 22 ] | |
| Рибопереключатель | Генная регуляция | Бактерии , Эукариоты | [ 23 ] | |
| РНК-термометр | Генная регуляция | Бактерии | [ 24 ] | |
| Последовательность вставки селеноцистеина | СУХОЙ | Направляет клетку на трансляцию стоп-кодонов UGA в селеноцистеины. | многоклеточные животные | [ 25 ] |
См. также
[ редактировать ]- ДНК
- РНК
- Другой
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д Дэвидсон Э.Х. (2006). Регуляторный геном: регуляторные сети генов в развитии и эволюции . Эльзевир. стр. 1–86.
- ^ Jump up to: а б с д Тейф В.Б. (2010). «Прогнозирование функций генной регуляции: уроки бактериофагов умеренного климата» . Биофизический журнал . 98 (7): 1247–56. Бибкод : 2010BpJ....98.1247T . дои : 10.1016/j.bpj.2009.11.046 . ПМК 2849075 . ПМИД 20371324 .
- ^ Бен-Табу де-Леон С., Дэвидсон Э.Х. (2007). «Генная регуляция: сеть генного контроля в разработке» (PDF) . Annu Rev Biophys Biomol Struct . 36 : 191–212. doi : 10.1146/annurev.biophys.35.040405.102002 . ПМИД 17291181 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Езиорска Д.М., Джордан К.В., Вэнс К.В. (2009). «Подход системной биологии к пониманию функции цис-регуляторного модуля». Семин. Сотовое развитие. Биол . 20 (7): 856–862. дои : 10.1016/j.semcdb.2009.07.007 . ПМИД 19660565 .
- ^ Jump up to: а б с Батлер Дж. Э., Кадонага Дж. Т. (октябрь 2002 г.). «Основной промотор РНК-полимеразы II: ключевой компонент регуляции экспрессии генов» . Гены и развитие . 16 (20): 2583–2592. дои : 10.1101/gad.1026202 . ПМИД 12381658 .
- ^ Чой С (17 мая 2008 г.). Введение в системную биологию . Springer Science & Business Media. п. 78. ИСБН 978-1-59745-531-2 .
- ^ Jump up to: а б Витткопп П.Дж., Калай Дж. (декабрь 2011 г.). «Цис-регуляторные элементы: молекулярные механизмы и эволюционные процессы, лежащие в основе дивергенции». Обзоры природы Генетика . 13 (1): 59–69. дои : 10.1038/nrg3095 . ПМИД 22143240 . S2CID 13513643 .
- ^ Меламед П., Йосефзун Ю. и др. (2 марта 2016 г.). «Усилители транскрипции: транскрипция, функция и гибкость» . Транскрипция . 7 (1): 26–31. дои : 10.1080/21541264.2015.1128517 . ПМЦ 4802784 . ПМИД 26934309 .
- ^ Ли Э, Дэвидсон Э.Х. (2009). «Создание сетей регулирования генов развития» . Врожденные дефекты Рез . 87 (2): 123–130. дои : 10.1002/bdrc.20152 . ПМЦ 2747644 . ПМИД 19530131 .
- ^ Аэртс, С.; и др. (2003). «Вычислительное обнаружение цис-регуляторных модулей». Биоинформатика . 19 (Приложение 2): ii5–14. doi : 10.1093/биоинформатика/btg1052 . ПМИД 14534164 .
- ^ Врзодек, Клеменс; Шредер, Адриан; Дрегер, Андреас; Ванке, Дирк; Берендзен, Кеннет В.; Кронфельд, Марсель; Хартер, Клаус; Зелл, Андреас (2010). «ModuleMaster: новый инструмент для расшифровки сетей регуляции транскрипции». Биосистемы . 99 (1). Ирландия: Эльзевир: 79–81. doi : 10.1016/j.biosystems.2009.09.005 . ISSN 0303-2647 . ПМИД 19819296 .
- ^ Парра Р.Г., Рор К.О., Койле Д., Перес-Кастро С., Янкилевич П. (2015). «INSECT 2.0: веб-сервер для полногеномного прогнозирования цис-регуляторных модулей» . Биоинформатика . 32 (8): 1229–31. doi : 10.1093/биоинформатика/btv726 . hdl : 11336/37980 . ПМИД 26656931 .
- ^ Рор СО, Парра Р.Г., Янкилевич П., Перес-Кастро С. (2013). «НАСЕКОМОЕ: IN-silico ПОИСК сопутствующих факторов транскрипции» . Биоинформатика . 29 (22): 2852–8. doi : 10.1093/биоинформатика/btt506 . hdl : 11336/12301 . ПМИД 24008418 .
- ^ Jump up to: а б Синха С., Лян Ю., Сиггиа Э. (2006). «Стубб: программа для открытия и анализа цис-регуляторных модулей» . Нуклеиновые кислоты Рез . 34 (проблема с веб-сервером): W555–W559. дои : 10.1093/нар/gkl224 . ПМЦ 1538799 . ПМИД 16845069 .
- ^ Jump up to: а б Чен X, Бланшетт М (2007). «Сравнение последовательностей без использования выравниваний: применение к подтипированию ВИЧ / SIV» . БМК Биоинформатика . 8 : 1–17. дои : 10.1186/1471-2105-8-1 . ПМЦ 1766362 . ПМИД 17199892 .
- ^ Шаран Р., Бен-Гур А., Лутс Г.Г., Овчаренко И. (2004). «CREME: Исследователь цис-регуляторных модулей для генома человека» . Нуклеиновые кислоты Рез . 32 (проблема с веб-сервером): W253–W256. дои : 10.1093/nar/gkh385 . ПМЦ 441523 . ПМИД 15215390 .
- ^ Бекарт М., Ферт А.Е., Чжан Ю., Гладышев В.Н., Аткинс Дж.Ф., Баранов П.В. (январь 2010 г.). «Recode-2: новый дизайн, новые инструменты поиска и еще много генов» . Исследования нуклеиновых кислот . 38 (Проблема с базой данных): D69–74. дои : 10.1093/nar/gkp788 . ПМЦ 2808893 . ПМИД 19783826 .
- ^ Чанг БАЙ, Ферт А.Е., Аткинс Дж.Ф. (март 2010 г.). «Сдвиг рамки у альфавирусов: разнообразие 3'-стимулирующих структур». Журнал молекулярной биологии . 397 (2): 448–456. дои : 10.1016/j.jmb.2010.01.044 . ПМИД 20114053 .
- ^ Гедрок Д.П., Корнуолл П.В. (февраль 2009 г.). «Псевдоузлы РНК со сдвигом рамки: структура и механизм» . Вирусные исследования . 139 (2): 193–208. doi : 10.1016/j.virusres.2008.06.008 . ПМК 2670756 . ПМИД 18621088 .
- ^ Мокрейс М, Вопаленский В, Коленаты О, Масек Т, Фекетова З, Секирова П, Скалудова Б, Криз В, Посписек М (январь 2006 г.). «IRESite: база данных экспериментально проверенных структур IRES (www.iresite.org)» . Исследования нуклеиновых кислот . 34 (Проблема с базой данных): D125–130. дои : 10.1093/nar/gkj081 . ПМЦ 1347444 . ПМИД 16381829 .
- ^ Хентце М.В., Кюн Л.К. (август 1996 г.). «Молекулярный контроль метаболизма железа у позвоночных: регуляторные цепи на основе мРНК, управляемые железом, оксидом азота и окислительным стрессом» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (16): 8175–8182. Бибкод : 1996PNAS...93.8175H . дои : 10.1073/pnas.93.16.8175 . ПМК 38642 . ПМИД 8710843 .
- ^ Платт Т (1986). «Терминация транскрипции и регуляция экспрессии генов». Ежегодный обзор биохимии . 55 : 339–372. дои : 10.1146/annurev.bi.55.070186.002011 . ПМИД 3527045 .
- ^ Брейкер RR (март 2008 г.). «Сложные рибопереключатели». Наука . 319 (5871): 1795–1797. Бибкод : 2008Sci...319.1795B . дои : 10.1126/science.1152621 . ПМИД 18369140 . S2CID 45588146 .
- ^ Кортманн Дж., Нарберхаус Ф (март 2012 г.). «Бактериальные РНК-термометры: молекулярные молнии и переключатели». Обзоры природы. Микробиология . 10 (4): 255–265. дои : 10.1038/nrmicro2730 . ПМИД 22421878 . S2CID 29414695 .
- ^ Вальчак Р., Вестхоф Э., Карбон П., Крол А. (апрель 1996 г.). «Новый структурный мотив РНК в элементе вставки селеноцистеина мРНК эукариотических селенопротеинов» . РНК . 2 (4): 367–379. ПМЦ 1369379 . ПМИД 8634917 .
Дальнейшее чтение
[ редактировать ]- Рэй Г.А. (март 2007 г.). «Эволюционное значение цис-регуляторных мутаций». Обзоры природы Генетика . 8 (3): 206–216. дои : 10.1038/nrg2063 . ПМИД 17304246 . S2CID 560067 .
- Гомпель Н., Прюдом Б., Виткопп П.Дж., Касснер В.А., Кэрролл С.Б. (февраль 2005 г.). «Шанс, попавший на крыло: цис-регуляторная эволюция и происхождение пигментных узоров у дрозофилы». Природа . 433 (7025): 481–487. Бибкод : 2005Natur.433..481G . дои : 10.1038/nature03235 . ПМИД 15690032 . S2CID 16422483 .
- Прюдом Б., Гомпель Н., Рокас А., Касснер В.А., Уильямс Т.М., Йе С.Д., Тру Дж.Р., Кэрролл С.Б. (апрель 2006 г.). «Повторяющаяся морфологическая эволюция посредством цис-регуляторных изменений в плейотропном гене». Природа . 440 (7087): 1050–1053. Бибкод : 2006Natur.440.1050P . дои : 10.1038/nature04597 . ПМИД 16625197 . S2CID 9581516 .
- Стерн Д.Л. (август 2000 г.). «Эволюционная биология развития и проблема изменчивости» . Эволюция; Международный журнал органической эволюции . 54 (4): 1079–1091. дои : 10.1111/j.0014-3820.2000.tb00544.x . ПМИД 11005278 .
- Уэзерби С.Д., Кэрролл С.Б., Гренье Дж.К. (2004). От ДНК к разнообразию: молекулярная генетика и эволюция дизайна животных . Кембридж, Массачусетс: Издательство Blackwell. ISBN 978-1-4051-1950-4 .
Внешние ссылки
[ редактировать ]- Информация о регулировании генов — вручную составленные списки ресурсов, обзоров, обсуждений в сообществе.
- Клеточный дарвинизм
- Регуляция экспрессии генов Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
| Обзор |
| ||||||
|---|---|---|---|---|---|---|---|
| Инженерное дело |
| ||||||