Теломеразный компонент РНК
| ТЕРК | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | TERC , DKCA1, PFBMFT2, SCARNA19, TR, TRC3, hTR, компонент теломеразной РНК, TER | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 602322 ; GeneCards : ТРИ ; ОМА : ТРЕТЬЕ – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||


| Теломеразная РНК позвоночных | |
|---|---|
 | |
| Идентификаторы | |
| Символ | Теломераза-хозяин |
| Рфам | RF00024 |
| Другие данные | |
| РНК Тип | Ген |
| Домен(ы) | Эукариоты ; Вирус |
| PDB Структуры | ПДБе |
| Теломеразная РНК инфузорий | |
|---|---|
 | |
| Идентификаторы | |
| Символ | Теломераза-цил |
| Рфам | RF00025 |
| Другие данные | |
| РНК Тип | Ген |
| Домен(ы) | Эукариот |
| PDB Структуры | ПДБе |
| Теломеразная РНК Saccharomyces cerevisiae | |
|---|---|
 | |
| Идентификаторы | |
| Символ | Sacc_теломераза |
| Рфам | RF01050 |
| Другие данные | |
| РНК Тип | Ген |
| Домен(ы) | Эукариот |
| PDB Структуры | ПДБе |
Компонент теломеразной РНК , также известный как TR , TER или TERC , представляет собой нкРНК, обнаруженную у эукариот и являющуюся компонентом теломеразы , фермента, используемого для удлинения теломер . [ 3 ] [ 4 ] TERC служит матрицей для репликации теломер ( обратной транскрипции ) теломеразой. Теломеразные РНК сильно различаются по последовательности и структуре у позвоночных, инфузорий и дрожжей, но они имеют общую структуру 5'- псевдоузла , близкую к матричной последовательности. Теломеразные РНК позвоночных имеют 3' H/ACA snoRNA -подобный домен. [ 5 ] [ 6 ] [ 7 ]
Структура
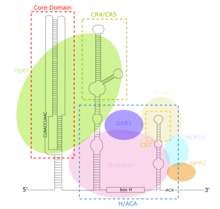
[ редактировать ]TERC представляет собой длинную некодирующую РНК (днРНК) длиной от ~150 нт у инфузорий до 400-600 нт у позвоночных и 1300 нт у дрожжей (Алнафах). TERC зрелого человека (hTR) имеет длину 451 нуклеотид. [ 8 ] TERC имеет обширные вторичные структурные особенности в четырех основных консервативных доменах. [ 9 ] Коровый домен, самый большой домен на 5'-конце TERC, содержит матричную последовательность теломер CUAAC . Его вторичная структура состоит из большой петли, содержащей матричную последовательность, спирали, замыкающей петлю P1, и псевдоузла P2/P3 . [ 10 ] Коровый домен и консервативный домен CR4/CR5 связаны с TERT и являются единственными доменами TERC, необходимыми для каталитической активности теломеразы in vitro. [ 11 ] 3'-конец TERC состоит из консервативного домена H/ACA. [ 10 ] структура из двух шпилек, соединенных одноцепочечным шарниром и граничащая на 3'-конце с одноцепочечной последовательностью ACA. [ 8 ] Домен H/ACA связывает дискерин , GAR1 , NOP10, NHP2 , образуя комплекс H/ACA RNP . [ 10 ] Консервативный домен CR7 также локализован на 3'-конце TERC и содержит 3nt-бокс CAB ( локализация тела Кахаля ), который связывает TCAB1. [ 10 ]
Функция
[ редактировать ]Теломераза представляет собой рибонуклеопротеин-полимеразу, которая поддерживает концы теломер путем добавления теломерного повтора TTAGGG. Этот повтор различается у разных эукариот см. в таблице в статье о теломерах ( полный список ). Фермент состоит из белкового компонента ( TERT ) с активностью обратной транскриптазы и компонента РНК, кодируемого этим геном, который служит матрицей для теломерного повтора. CCCUAA, обнаруженный вблизи положения 50 последовательности TERC позвоночных, действует как матрица. Экспрессия теломеразы играет роль в клеточном старении , поскольку она обычно подавляется в постнатальных соматических клетках, что приводит к прогрессирующему укорочению теломер. Дерегуляция экспрессии теломеразы в соматических клетках может быть вовлечена в онкогенез . Исследования на мышах показывают, что теломераза также участвует в восстановлении хромосом , поскольку de novo синтез теломерных повторов может происходить при двухцепочечных разрывах . [ 12 ] Гомологи TERC также можно обнаружить в вирусах герпеса Gallid . [ 13 ]
Основной домен TERC содержит матрицу РНК , из которой TERT синтезирует теломерные повторы TTAGGG. [ 10 ] В отличие от других РНП, в теломеразе белок TERT является каталитическим, тогда как днРНК TERC является структурным, а не действует как рибозим . [ 14 ] Основной области TERC и TERT достаточно для восстановления каталитической теломеразной активности in vitro. [ 10 ] [ 11 ] Домен H/ACA TERC рекрутирует дискериновый комплекс ( DKC1 , GAR1 , NOP10, NHP2 ), который стабилизирует TERC, увеличивая образование теломеразного комплекса и общую каталитическую активность. [ 10 ] Домен CR7 связывает TCAB1, который локализует теломеразу в тельцах Кахаля , еще больше увеличивая каталитическую активность теломеразы. [ 10 ] TERC экспрессируется повсеместно, даже в клетках, лишенных теломеразной активности и экспрессии TERT. [ 15 ] В результате были предложены различные независимые от TERT функциональные роли TERC. 14 генов, содержащих мотив связывания TERC, непосредственно транскрипционно регулируются TERC посредством увеличения экспрессии, опосредованного образованием триплекса РНК-ДНК. TERC-опосредованная активация Lin37, Trpg1l, tyrobp , Usp16 стимулирует путь NF-κB , что приводит к усилению экспрессии и секреции воспалительных цитокинов . [ 16 ]
Биосинтез
[ редактировать ]В отличие от большинства днРНК , которые собираются из интронов с помощью сплайсосомы напрямую , hTR транскрибируется со специального сайта промотора. [ 8 ] расположен в геномном локусе 3q26.2 [ 17 ] РНК - полимеразой II . [ 8 ] Зрелый hTR имеет длину 451 нуклеотид, но примерно 1/3 клеточных транскриптов hTR в устойчивом состоянии имеют ~10 нуклеотидов, геномно кодируемых 3'-хвосты. Большинство этих расширенных видов hTR имеют дополнительное 3'-удлинение олиго-А. [ 8 ] Процессинг незрелого 3'-хвостого hTR в зрелый 451-нуклеотидный hTR может осуществляться путем прямой 3'-5'- экзорибонуклеолитической деградации или непрямым путем олигоаденилирования с помощью PAPD5, удаления 3'-олиго-А-хвоста с помощью 3'-5' РНК. экзонуклеаза PARN и последующая 3'-5' экзорибонуклеолитическая деградация. [ 8 ] Расширенные транскрипты hTR также разрушаются экзосомой РНК . [ 8 ]
5'-концы транскриптов hTR также подвергаются дополнительному процессингу. TGS-1 гиперметилирует кэп 5'-метилгуанозина до кэпа N2,2,7 триметилгуанозина (TMG), что ингибирует созревание hTR. [ 18 ] Связывание комплекса дискерина с транскрибируемыми доменами H/ACA hTR во время транскрипции способствует терминации транскрипции. [ 8 ] Контроль относительной скорости этих различных конкурирующих путей, которые активируют или ингибируют созревание hTR, является важнейшим элементом регуляции общей активности теломеразы.
Клиническое значение
[ редактировать ]потери функции Мутации в геномном локусе TERC связаны с множеством дегенеративных заболеваний . Мутации в TERC связаны с врожденным дискератозом . [ 19 ] идиопатический легочный фиброз , [ 20 ] апластическая анемия и миелодисплазия . [ 10 ] Сверхэкспрессия и неправильная регуляция TERC связаны с различными видами рака . Повышение регуляции hTR широко наблюдается у пациентов с предраковым фенотипом шейки матки в результате инфекции ВПЧ . [ 21 ] Сверхэкспрессия TERC усиливает MDV -опосредованный онкогенез . [ 22 ] и наблюдается при раке желудка . [ 23 ] Сверхэкспрессия TERC также наблюдается при воспалительных состояниях, таких как диабет II типа и рассеянный склероз , из-за TERC-опосредованной активации воспалительного пути NF-κB . [ 16 ]
TERC играет защитную роль при остеопорозе , а повышенная его экспрессия останавливает скорость остеогенеза . [ 24 ] Из-за его сверхэкспрессии в ряде фенотипов рака TERC был исследован как потенциальный биомаркер рака . Было обнаружено, что он является эффективным биомаркером плоскоклеточного рака легких (LUSC). [ 25 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000277925 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Фэн Дж., Фанк В.Д., Ван С.С., Вайнрих С.Л., Авилион А.А., Чиу К.П. и др. (сентябрь 1995 г.). «РНК-компонент теломеразы человека». Наука . 269 (5228): 1236–41. Бибкод : 1995Sci...269.1236F . дои : 10.1126/science.7544491 . ПМИД 7544491 . S2CID 9440710 .
- ^ Жади Б.Е., Ричард П., Бертран Э., Кисс Т. (февраль 2006 г.). «Зависимое от клеточного цикла рекрутирование теломеразной РНК и телец Кахаля в теломеры человека» . Молекулярная биология клетки . 17 (2): 944–54. doi : 10.1091/mbc.E05-09-0904 . ПМЦ 1356602 . ПМИД 16319170 .
- ^ Маккормик-Грэм М., Ромеро Д.П. (апрель 1995 г.). «Особенности структуры теломеразной РНК ресничек» . Исследования нуклеиновых кислот . 23 (7): 1091–7. дои : 10.1093/нар/23.7.1091 . ПМК 306816 . ПМИД 7739888 .
- ^ Лингнер Дж., Хендрик Л.Л., Чех Т.Р. (август 1994 г.). «Теломеразные РНК разных инфузорий имеют общую вторичную структуру и перестановочную матрицу» . Гены и развитие . 8 (16): 1984–98. дои : 10.1101/гад.16.08.1984 . ПМИД 7958872 .
- ^ Теймер Калифорния, Фейгон Дж (июнь 2006 г.). «Структура и функции теломеразной РНК». Современное мнение в области структурной биологии . 16 (3): 307–18. дои : 10.1016/j.sbi.2006.05.005 . ПМИД 16713250 .
- ^ Jump up to: а б с д и ж г час Роук С.М., Чен Л., Чакраварти А.Л., Феррелл Дж.Э., Раффа Г.Д., Артанди С.Е. (май 2019 г.). «Нарушение кинетики созревания теломеразной РНК провоцирует заболевание» . Молекулярная клетка . 74 (4): 688–700.е3. doi : 10.1016/j.molcel.2019.02.033 . ПМК 6525023 . ПМИД 30930056 .
- ^ Альнафах Р.А., Адишеш М., Баттон Л., Сарецки Г., Хапангама Д.К. (2019). «Теломераза и теломеры при раке эндометрия» . Границы онкологии . 9 : 344. doi : 10.3389/fonc.2019.00344 . ПМК 6533802 . ПМИД 31157162 .
- ^ Jump up to: а б с д и ж г час я Чжан Ц, Ким Н.К., Фейгон Дж. (декабрь 2011 г.). «Архитектура теломеразной РНК человека» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (51): 20325–32. Бибкод : 2011PNAS..10820325Z . дои : 10.1073/pnas.1100279108 . ПМЦ 3251123 . ПМИД 21844345 .
- ^ Jump up to: а б Уэбб CJ, Закян В.А. (август 2016 г.). «Теломеразная РНК — это больше, чем просто матрица ДНК» . Биология РНК . 13 (8): 683–9. дои : 10.1080/15476286.2016.1191725 . ПМЦ 4993324 . ПМИД 27245259 .
- ^ «Ген Энтрез: компонент теломеразной РНК TERC» .
- ^ Фрагне Л., Кут Э., Расшерт Д. (июнь 2005 г.). «Сравнительное функциональное исследование вирусной теломеразной РНК на основе природных мутаций» . Журнал биологической химии . 280 (25): 23502–15. дои : 10.1074/jbc.M501163200 . ПМИД 15811851 . S2CID 24301693 .
- ^ Ван Ю, Сушак Л, Фейгон Дж (декабрь 2019 г.). «Структурная биология теломеразы» . Перспективы Колд-Спринг-Харбор в биологии . 11 (12): а032383. doi : 10.1101/cshperspect.a032383 . ПМК 6886448 . ПМИД 31451513 .
- ^ Шэй Дж.В., Райт МЫ (май 2019 г.). «Теломеры и теломераза: три десятилетия прогресса». Обзоры природы Генетика . 20 (5): 299–309. дои : 10.1038/s41576-019-0099-1 . ПМИД 30760854 . S2CID 61156603 .
- ^ Jump up to: а б Лю Х, Ян Ю, Гэ Ю, Лю Дж, Чжао Ю (сентябрь 2019 г.). «TERC способствует клеточному воспалительному ответу независимо от теломеразы» . Исследования нуклеиновых кислот . 47 (15): 8084–8095. дои : 10.1093/nar/gkz584 . ПМЦ 6735767 . ПМИД 31294790 .
- ^ «Запись OMIM - * 602322 - КОМПОНЕНТ ТЕЛОМЕРАЗНОЙ РНК; TERC» . www.omim.org . Проверено 02 марта 2020 г.
- ^ Чен Л., Роук С.М., Галати А., Бавассо Ф., Микели Э., Саджио И. и др. (февраль 2020 г.). «Потеря человеческой гиперметилазы TGS1 способствует увеличению теломеразной РНК и удлинению теломер» . Отчеты по ячейкам . 30 (5): 1358–1372.e5. дои : 10.1016/j.celrep.2020.01.004 . ПМК 7156301 . ПМИД 32023455 .
- ^ Рич Р.Р. (13 января 2018 г.). Клиническая иммунология: принципы и практика (Пятое изд.). [Св. Луис, Миссури] ISBN 978-0-7020-7039-6 . OCLC 1023865227 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Свигрис Дж. Дж., Браун К. К. (25 июля 2018 г.). Идиопатический легочный фиброз . Сент-Луис. ISBN 978-0-323-54432-0 . OCLC 1053744041 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Лю Ю, Фань П., Ян Ю, Сюй С, Хуан Ю, Ли Д и др. (ноябрь 2019 г.). «Вирус папилломы человека и ген компонента теломеразной РНК человека в прогрессировании рака шейки матки» . Научные отчеты . 9 (1): 15926. Бибкод : 2019NatSR...915926L . дои : 10.1038/s41598-019-52195-5 . ПМЦ 6828729 . ПМИД 31685833 .
- ^ Хеймар А., Тримперт Дж., Гроенке Н., Кауфер Б.Б. (март 2019 г.). «Сверхэкспрессия клеточной теломеразной РНК усиливает индуцированное вирусом образование рака». Онкоген . 38 (10): 1778–1786. дои : 10.1038/s41388-018-0544-1 . PMID 30846849 . S2CID 53085869 .
- ^ Хейне Б., Хаммел М., Демель Г., Штейн Х. (июнь 1998 г.). «Демонстрация постоянной активации компонента теломеразной РНК при карциноме желудка человека с использованием гибридизации in situ». Журнал патологии . 185 (2): 139–44. doi : 10.1002/(SICI)1096-9896(199806)185:2<139::AID-PATH79>3.0.CO;2-L . ПМИД 9713339 . S2CID 21966828 .
- ^ Гао Г.К., Ян Д.В., Лю В. (январь 2020 г.). «LncRNA TERC облегчает прогрессирование остеопороза путем поглощения миРНК-217 для активации RUNX2». Европейский обзор медицинских и фармакологических наук . 24 (2): 526–534. дои : 10.26355/eurrev_202001_20029 . ПМИД 32016954 . S2CID 211024218 .
- ^ Сторти CB, де Оливейра Р.А., де Карвалью М., Хасимото Э.Н., Катанео Д.С., Катанео А.Дж. и др. (февраль 2020 г.). «Связанные с теломерами гены и теломерные днРНК являются кандидатами на биомаркеры при плоскоклеточном раке легких (LUSC)». Экспериментальная и молекулярная патология . 112 : 104354. doi : 10.1016/j.yexmp.2019.104354 . ПМИД 31837325 . S2CID 209385638 .
Дальнейшее чтение
[ редактировать ]- де Ланге Т., Джекс Т. (август 1999 г.). «К лучшему или к худшему? Ингибирование теломеразы и рак» . Клетка . 98 (3): 273–5. дои : 10.1016/S0092-8674(00)81955-8 . ПМИД 10458601 . S2CID 14642341 .
- Марроне А., Докал I (декабрь 2004 г.). «Врожденный дискератоз: молекулярное понимание функции теломеразы, старения и рака». Обзоры экспертов в области молекулярной медицины . 6 (26): 1–23. дои : 10.1017/S1462399404008671 . ПМИД 15613268 . S2CID 38163343 .
- Ямагути Х (июнь 2007 г.). «Мутации генов теломеразного комплекса, связанные с недостаточностью костного мозга» . Журнал медицинской школы Ниппон . 74 (3): 202–9. дои : 10.1272/jnms.74.202 . ПМИД 17625368 .
- Зауг А.Дж., Лингер Дж., Чех Т.Р. (февраль 1996 г.). «Метод определения 3'-концов РНК и применение к теломеразной РНК человека» . Исследования нуклеиновых кислот . 24 (3): 532–3. дои : 10.1093/нар/24.3.532 . ПМК 145649 . ПМИД 8602368 .
- Содер А.И., Хоар С.Ф., Мьюир С., Балмейн А., Паркинсон Э.К., Кейт В.Н. (апрель 1997 г.). «Картирование гена компонента теломеразной РНК мыши, Terc, на хромосоме 3 путем флуоресцентной гибридизации in situ и окраски хромосом мыши». Геномика . 41 (2): 293–4. дои : 10.1006/geno.1997.4621 . ПМИД 9143511 .
- Чжао Дж.К., Хоар С.Ф., Макфарлейн Р., Мьюир С., Паркинсон Е.К., Блэк Д.М., Кейт В.Н. (март 1998 г.). «Клонирование и характеристика последовательностей промотора гена теломеразной РНК человека и мыши». Онкоген . 16 (10): 1345–50. дои : 10.1038/sj.onc.1201892 . ПМИД 9546436 . S2CID 2699389 .
- Митчелл-младший, Вуд Э., Коллинз К. (декабрь 1999 г.). «Компонент теломеразы дефектен при врожденном дискератозе человека». Природа . 402 (6761): 551–5. Бибкод : 1999Natur.402..551M . дои : 10.1038/990141 . ПМИД 10591218 . S2CID 4430482 .
- Чен Дж.Л., Бласко М.А., Грейдер К.В. (март 2000 г.). «Вторичная структура теломеразной РНК позвоночных» . Клетка . 100 (5): 503–14. doi : 10.1016/S0092-8674(00)80687-X . ПМИД 10721988 . S2CID 15642776 .
- Вонг К.К., Чанг С., Вейлер С.Р., Ганесан С., Чаудхури Дж., Чжу С. и др. (сентябрь 2000 г.). «Дисфункция теломер ухудшает восстановление ДНК и повышает чувствительность к ионизирующей радиации». Природная генетика . 26 (1): 85–8. дои : 10.1038/79232 . ПМИД 10973255 . S2CID 1873111 .
- Митчелл-младший, Коллинз К. (август 2000 г.). «Активация теломеразы человека требует двух независимых взаимодействий между теломеразной РНК и обратной транскриптазой теломеразы» . Молекулярная клетка . 6 (2): 361–71. дои : 10.1016/S1097-2765(00)00036-8 . ПМИД 10983983 .
- Имото И., Пимхаокхам А., Фукуда Й., Ян ЗК, Шимада Й., Номура Н. и др. (август 2001 г.). «SNO является вероятной мишенью для амплификации гена 3q26 при плоскоклеточном раке пищевода». Связь с биохимическими и биофизическими исследованиями . 286 (3): 559–65. дои : 10.1006/bbrc.2001.5428 . ПМИД 11511096 .
- Вуллиами Т., Маррон А., Голдман Ф., Дирлав А., Бесслер М., Мейсон П.Дж., Докал I (сентябрь 2001 г.). «РНК-компонент теломеразы мутирует при аутосомно-доминантном врожденном дискератозе». Природа . 413 (6854): 432–5. Бибкод : 2001Natur.413..432V . дои : 10.1038/35096585 . ПМИД 11574891 . S2CID 4348062 .
- Прузан Р., Понграч К., Гитцен К., Валлвебер Г., Грязнов С. (январь 2002 г.). «Аллостерические ингибиторы теломеразы: олигонуклеотиды N3'-->P5' фосфорамидаты» . Исследования нуклеиновых кислот . 30 (2): 559–68. дои : 10.1093/нар/30.2.559 . ПМК 99832 . ПМИД 11788719 .
- Чжан Р.Г., Чжан Р.П., Ван XW, Се Х (март 2002 г.). «Влияние цисплатина на активность теломеразы и длину теломер в клетках гепатомы человека BEL-7404» . Клеточные исследования . 12 (1): 55–62. дои : 10.1038/sj.cr.7290110 . ПМИД 11942411 . S2CID 36839452 .
- Ян Ю, Чен Ю, Чжан С, Хуан Х, Вайсман С.М. (июль 2002 г.). «Ядрышковая локализация белка hTERT связана с функцией теломеразы». Экспериментальные исследования клеток . 277 (2): 201–9. дои : 10.1006/excr.2002.5541 . ПМИД 12083802 .
- Чанг Дж.Т., Чен Ю.Л., Ян Х.Т., Чен С.И., Ченг А.Дж. (июль 2002 г.). «Дифференциальная регуляция теломеразной активности шестью субъединицами теломеразы» . Европейский журнал биохимии . 269 (14): 3442–50. дои : 10.1046/j.1432-1033.2002.03025.x . ПМИД 12135483 .
- Гавори Г., Фэрроу М., Баласубраманиан С. (октябрь 2002 г.). «Требования к минимальной длине домена выравнивания теломеразной РНК человека для поддержания каталитической активности in vitro» . Исследования нуклеиновых кислот . 30 (20): 4470–80. дои : 10.1093/nar/gkf575 . ПМЦ 137139 . ПМИД 12384594 .
- Суд А.К., Коффин Дж., Джаббари С., Буллер Р.Э., Хендрикс М.Дж., Клингельхуц А. (2003). «Нулевые мутации р53 связаны с теломераза-отрицательным фенотипом при карциноме яичника» . Биология и терапия рака . 1 (5): 511–7. дои : 10.4161/cbt.1.5.167 . ПМИД 12496479 . S2CID 45381814 .
- Антал М., Борос Э., Солимози Ф., Кисс Т. (февраль 2002 г.). «Анализ структуры теломеразной РНК человека in vivo» . Исследования нуклеиновых кислот . 30 (4): 912–20. дои : 10.1093/нар/30.4.912 . ПМЦ 100349 . ПМИД 11842102 .