Полиднавириформиды
| Полиднавирус | |
|---|---|

| |
| Электронная микрофотография браковируса | |
| Классификация вирусов | |
| (без рейтинга): | Вириформный |
| Семья: | Полиднавириформиды |
| Роды | |
| Синонимы | |
| |
Polydnaviriformidae ( / p ɒ ˈ l ɪ d n ə v ɪ ə r ˌ f ɔː m ɪ d ɛ / PDV ) [ 1 ] — семейство насекомых вириформных ; члены известны как полиднавирусы . В семействе два рода: Bracoform и Ichnoviriform . Полиднавирусы образуют симбиотические отношения с паразитоидными осами . Ихновириформы (IV) встречаются у ихневмонид , а браковироформы (BV) — у ос Браконид . Личинки ос обеих этих групп сами паразитируют на чешуекрылых (мотыльках и бабочках), а полиднавирусы играют важную роль в обходе иммунного ответа своих паразитированных хозяев. [ 2 ] [ 3 ] Между BV и IV существует небольшая гомология последовательностей или она отсутствует вообще , что позволяет предположить, что эти два рода развивались независимо в течение длительного времени.
Таксономия
[ редактировать ]Браковириформный
- Браковириформная высота
- Браковириформный серебристый фронтис
- Браковировидный черный бурни
- Канадский браковирусный вид
- Браковириформные группы
- Браковириформный крассикорнис
- Браковириформные кроцепопеды
- Браковириформные кривопятнистые пятна
- Браковириформный разрушитель
- Браковириформные экдитолофы
- Браковириформные фасетки
- Браковириформный флавикоксис
- Браковириформный флавипедис
- Браковириформные флавитестовые
- Браковировидные фумифераны
- Браковириформные клубочки
- Браковириформные гифантрии
- Браковириформная пустота
- Браковириформный индийский
- Браковириформ островной
- Браковировидный карияй
- Браковириформный липаридис
- Браковириформный маргинивентрис
- Браковировидные меланосцелы
- Браковириформный нигриципит
- Браковириформный орнигис
- Браковировидные палеакриты
- Браковировидная четырехзубая
- Браковириформные краснухи
- Браковириформное шефери
- Браковириформный техасский
Ichnoviriform
- Ихновириформные акроникты
- Ichnoviriform annulipedis
- Ихновириформный апрель
- Ихновириформные арджуны
- Ихновириформ бенефакторис
- Ихновириформный эрибори
- Небольшая ихновирусная форма
- Ихновириформные желтые
- Ихновирусные щипцы
- Ихновирусные беглецы
- Ихновириформные фумифераны
- Ихновириформные коленчатые
- Ихновирусные инфекции
- Ихновириформное прерывание
- Ихновириформные лимантрии
- Ихновириформная гора
- Ихновириформные волоски
- Ихновириформный соперник
- Ихновириформный ростралис
- Сонорский ихновириформный
- Ихновириформный тенифеморис
- Ихновирусные сверла
Структура
[ редактировать ]Вирусы Polydnaviridae имеют оболочку , вытянутую эллипсоидную и цилиндрическую геометрию. Геномы кольцевые и сегментированные, состоят из множества сегментов двухцепочечной суперспиральной ДНК , упакованных в капсидные белки . Их длина составляет около 2,0–31 КБ. [ 2 ]
| Род | Структура | Симметрия | Капсид | Геномное расположение | Геномная сегментация |
|---|---|---|---|---|---|
| Ichnoviriform | Вытянутый эллипсоид | обернутый | Круговой | Сегментированный | |
| Браковириформный | Вытянутый эллипсоид | обернутый | Круговой | Сегментированный |
Жизненный цикл
[ редактировать ]Репликация вируса ядерная. с помощью ДНК-матрицы Транскрипция - это метод транскрипции. Вирус покидает клетку-хозяина путем экспорта ядерных пор .
Хозяевами вируса служат паразитоидные осы, а хозяевами этих ос - чешуекрылые. Самка осы впрыскивает своему хозяину одно или несколько яиц вместе с некоторым количеством вируса. Вирус и оса находятся в мутуалистических симбиотических отношениях: экспрессия вирусных генов не позволяет иммунной системе хозяина осы убить введенное яйцо осы и вызывает другие физиологические изменения, которые в конечном итоге приводят к смерти зараженного хозяина. Пути передачи – родительские. [ 2 ]
| Род | Сведения о хосте | Тканевой тропизм | Детали входа | Подробности выпуска | Сайт репликации | Монтажный участок | Передача инфекции |
|---|---|---|---|---|---|---|---|
| Ichnoviriform | Паразитоидные осы (Ichneumonidae) | гемоциты; толстые тела | Неизвестный | Почкование через клеточную мембрану [ 2 ] [ 4 ] | Ядро | Ядро | Неизвестный |
| Браковириформный | Осы-паразитоиды (Braconidae) | гемоциты; толстые тела | Неизвестный | Лизис [ 2 ] | Ядро | Ядро | Неизвестный |
Биология
[ редактировать ]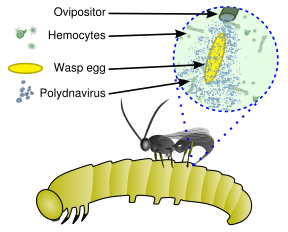
Эти вирусы являются частью уникальной биологической системы, состоящей из эндопаразитической осы ( паразитоида ), личинки-хозяина (обычно чешуекрылых ) и вируса. Полный геном вируса является эндогенным , рассеянным среди генома осы. Вирус размножается только в определенной части яичника, называемой чашечкой , куколок и взрослых самок ос. Вирус впрыскивается вместе с яйцом осы в полость тела гусеницы-хозяина чешуекрылых и заражает клетки гусеницы. Заражение не приводит к репликации новых вирусов; гусеницы скорее, он влияет на иммунную систему , поскольку вирион несет гены вирулентности вместо генов репликации вируса. [ 5 ] Его можно считать разновидностью вирусного вектора . [ 6 ]
Без вирусной инфекции фагоцитарные гемоциты (клетки крови) будут инкапсулировать и убивать осиное яйцо и личинки, но иммуносупрессия, вызванная вирусом, позволяет выжить осиному яйцу и личинкам, что приводит к вылуплению и полному развитию неполовозрелой осы в гусеница. Кроме того, гены, экспрессируемые полиднавирусом у пораженного паразитом хозяина, изменяют развитие и метаболизм хозяина, что благоприятствует росту и выживанию личинки паразитоида. [ 4 ] [ 7 ]
Потенциальные подсемейства носителей
[ редактировать ]Характеристики
[ редактировать ]Оба рода PDV имеют определенные характеристики:
- вирусные частицы каждого из них содержат несколько сегментов дцДНК (двухцепочечная или «нормальная» ДНК, в отличие от одноцепочечной ДНК или РНК с положительным или отрицательным смыслом, обнаруженной в некоторых других вирусах), причем каждый сегмент содержит только часть полного генома (так же, как хромосомы в эукариотических организмах )
- геном вируса имеет эукариотические характеристики, такие как наличие интронов (обычно для генов насекомых, но редко для вирусов) и низкая плотность кодирования. [ 9 ]
- геном каждого вируса интегрирован в геном осы-хозяина [ 10 ]
- геном организован в несколько семейств генов, состоящих из нескольких членов (которые различаются у браковирусов и ихновирусов)
- вирусные частицы производятся только в определенных типах клеток репродуктивных органов самки осы.
Морфология этих двух родов различна при наблюдении с помощью электронной микроскопии. Ихновирусы, как правило, имеют яйцевидную форму, тогда как браковирусы представляют собой короткие палочки. Вирионы браковирусов высвобождаются в результате лизиса клеток ; вирионы ихновирусов выделяются путем почкования.
Эволюция
[ редактировать ]Анализ нуклеиновых кислот предполагает очень длительную связь вирусов с осами (по оценкам, 73,7 миллиона лет ± 10 миллионов). [ 11 ]
Старая теория, основанная на осах
[ редактировать ]Было выдвинуто два предположения о том, как развивалась ассоциация осы и вируса. Первая предполагает, что вирус произошел от генов осы. Многие паразитоиды, не использующие PDV, вводят белки, обеспечивающие многие из тех же функций, то есть подавление иммунного ответа на яйцо паразита. В этой модели осы-бракониды и ихневмониды упаковывают гены для этих функций в вирусы, по сути создавая систему переноса генов, которая приводит к тому, что гусеница вырабатывает факторы, подавляющие иммунитет. В этом сценарии структурные белки (капсиды) PDV, вероятно, были «заимствованы» у существующих вирусов. [ 12 ]
Современная теория эндогенных вирусов
[ редактировать ]Альтернативное предположение предполагает, что предки ос развили полезную связь с существующим вирусом, что в конечном итоге привело к интеграции вируса в геном осы. После интеграции гены, ответственные за репликацию вируса, и капсиды (в конечном итоге) больше не включались в геном PDV. Эта гипотеза подтверждается явными морфологическими различиями между IV и BV, что позволяет предположить разные предковые вирусы для этих двух родов. БВ, вероятно, произошел от нудивируса , в частности бетанудивируса. [ 13 ] ~ 100 миллионов лет назад . [ 14 ] IV имеет менее ясное происхождение: хотя в более ранних сообщениях был обнаружен белок p44/p53, имеющий структурное сходство с асковирусом , связь не была подтверждена в более поздних исследованиях. [ 15 ] В результате в настоящее время существует мнение, что IV произошел от еще не идентифицированного нового вирусного семейства. [ 13 ] со слабой связью с NCLDV . [ 16 ] В любом случае оба рода образовались в результате единого события интеграции в соответствующих линиях ос. [ 5 ]
Две группы вирусов этого семейства на самом деле не связаны филогенетически, что позволяет предположить, что этот таксон, возможно, нуждается в пересмотре. [ 17 ]
Влияние на иммунитет хозяина
[ редактировать ]У хозяина могут сработать несколько механизмов иммунной системы насекомых, когда оса откладывает яйца и когда развивается оса-паразит. Когда в тело насекомого попадает крупное тело (осиное яйцо или мелкая частица, использованная в эксперименте), классической иммунной реакцией является инкапсуляция гематоцитами. Инкапсулированное тело также может быть меланизировано с целью удушья благодаря другому типу гемоцитов, который использует фенолоксидазный путь для производства меланина. Маленькие частицы могут подвергаться фагоцитозу, а затем макрофагальные клетки также меланизироваться в узелке. Наконец, насекомые также могут реагировать выработкой противовирусных пептидов . [ 18 ]
ПолиДНКвирус защищает личинок перепончатокрылых от иммунной системы хозяина, действуя на разных уровнях.
- Сначала они могут вывести из строя или уничтожить гематоциты. ПолиДНКвирус, связанный с Cotesia rubecula , кодирует белок CrV1, который денатурирует актиновые нити в гематоцитах, поэтому эти клетки становятся менее способными двигаться и прилипать к личинкам. [ 18 ] Разрушитель микроплита Браковирус (MdBV) индуцирует апоптоз гематоцитов благодаря своему гену PTP-H2. [ 18 ] Он также снижает адгезионную способность гематоцитов благодаря гену Glc1.8. Ген также ингибирует фагоцитоз. [ 19 ]
- ПолиДНКвирус также может действовать на меланизацию, MdBV препятствует выработке фенолоксидазы . [ 20 ]
- Наконец, полиДНКвирус также может продуцировать вирусные анкирины , которые мешают выработке противовирусных пептидов. [ 21 ] У некоторых ихновирусов Ванкирин также может предотвращать апоптоз — крайнюю реакцию клетки, направленную на блокирование распространения вируса. [ 22 ] [ 23 ]
- Ихновирусы продуцируют некоторые белки, называемые виннексинами, которые признаны гомологичными иннексинам насекомых . Они отвечают за кодировку структурных единиц щелевых переходов. Эти белки могут изменять межклеточную коммуникацию, что может объяснить нарушение процесса инкапсидации . [ 24 ]
Вирусоподобные частицы
[ редактировать ]Другая стратегия, используемая паразитоидными перепончатокрылыми для защиты своего потомства, — это производство вирусоподобных частиц . VLP по своей структуре похожи на вирусы, но не несут нуклеиновой кислоты. Например, Venturia canescens ( Ichneumonidea ) и Leptopilina sp. ( Figitidaea ) производят VLP.
VLP можно сравнить с полиДНКвирусом, поскольку они секретируются одинаковым образом и оба защищают личинок от иммунной системы хозяина. V. canescens -VLP (VcVLP1, VcVLP2, VcNEP ...) продуцируются в клетках чашечки до того, как они попадут в яйцеводы. Работы 2006 года не обнаружили их связи с какими-либо вирусами и предполагали клеточное происхождение. [ 12 ] Более недавнее сравнение связывает их с сильно перетасованными последовательностями одомашненного нудивируса . Эта ссылка дает название эндогенного нудивируса Venturia canescens (VcENV), альфанудивируса, тесно связанного с NlENV, обнаруженного в Nilaparvata lugens . [ 25 ]
VLP защищают личинок перепончатокрылых локально, тогда как полиДНКвирус может иметь более глобальный эффект. VLP позволяют личинкам избегать иммунной системы: личинка не распознается хозяином как вредная, или иммунные клетки не могут взаимодействовать с ней благодаря VLP. [ 12 ] Venturia canescens использует их вместо полиднавирусов, поскольку ихновирус деактивирован. [ 25 ]
Оса Leptopilina гетеротома секретирует VLP, которые способны проникать в ламеллоциты благодаря специфическим рецепторам, а затем изменять форму и свойства поверхности ламеллоцитов, так что они становятся неэффективными, а личинки защищены от инкапсуляции. [ 26 ] VLP Leptopilina или внеклеточные везикулы смешанной стратегии (MSEV) содержат некоторые системы секреции. Их эволюционная картина менее ясна, [ 27 ] но недавно обнаруженный вирус, нитевидный вирус L. boulardi (LbFV), демонстрирует значительное сходство. [ 28 ]
Микро-РНК
[ редактировать ]МикроРНК — это небольшие фрагменты РНК, образующиеся в клетках хозяина благодаря специфическому ферментативному механизму. Они способствуют разрушению вирусной РНК. МикроРНК прикрепляются к вирусной РНК, поскольку они дополняют друг друга. Затем комплекс распознается ферментом, который его разрушает. Это явление известно как PTGS (посттранскрипционное молчание генов). [ 29 ] или РНКи ( РНК-интерференция ).
Интересно рассмотреть феномен микроРНК в контексте полиДНКвирусов. Можно сформулировать множество гипотез:
- Braconidae несут в своем геноме гены, связанные с нудивирусом . [ 30 ] поэтому они могут быть способны производить микроРНК против нудивируса в качестве врожденного иммунитета.
- Осы, возможно, используют микроРНК для контроля вирусных генов, которые они несут.
- ПолиДНКвирус также может использовать PTGS для вмешательства в экспрессию генов хозяина.
- PTGS также используется для развития организмов, используя те же ферменты, что и для подавления антивирусных генов, поэтому мы можем предположить, что если хозяин использует PTGS против полиДНКвируса, возможно, это также влияет на его развитие.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Подробности о таксонах: Polydnaviriformidae» . ictv.global .
Предложение: 2021.006DRPolydnaviriformidae_1renfam_3rensp
- ^ Перейти обратно: а б с д и «Вирусная зона» . ЭксПАСи . Проверено 15 июня 2015 г.
- ^ «Таксономия вирусов: выпуск 2020 г.» . Международный комитет по таксономии вирусов (ICTV). Март 2021 года . Проверено 24 мая 2021 г.
- ^ Перейти обратно: а б с д и ж г Уэбб, бакалавр (1998). Биология полиднавируса, структура генома и эволюция . В Миллер, Л.К., Болл, Лос-Анджелес, ред. Вирусы насекомых . Издательская корпорация «Пленум». стр. 105–139.
- ^ Перейти обратно: а б Стрэнд, MR; Берк, Греция (май 2015 г.). «Полиднавирусы: от открытия к современным открытиям» . Вирусология . 479–480: 393–402. дои : 10.1016/j.virol.2015.01.018 . ПМК 4424053 . ПМИД 25670535 .
- ^ Перейти обратно: а б Берк, Гаэлен Р.; Стрэнд, Майкл Р. (31 января 2012 г.). «Полиднавирусы паразитических ос: одомашнивание вирусов, которые будут действовать как векторы доставки генов» . Насекомые . 3 (1): 91–119. дои : 10.3390/insects3010091 . ПМЦ 4553618 . ПМИД 26467950 .
- ^ Руссинк, MJ (2011). «Изменения в динамике популяций мутуалистических и патогенных вирусов» . Вирусы . 3 (1): 12–19. дои : 10.3390/v3010012 . ПМЦ 3187592 . ПМИД 21994724 .
- ^ Мерфи, Николас; Бэнкс, Джонатан К.; Уитфилд, Джеймс Б.; Остин, Эндрю Д. (1 апреля 2008 г.). «Филогения паразитических подсемейств микрогастроидов (Hymenoptera: Braconidae) на основе данных о последовательностях семи генов с улучшенной оценкой времени происхождения линии». Молекулярная филогенетика и эволюция . 47 (1): 378–395. дои : 10.1016/j.ympev.2008.01.022 . ПМИД 18325792 .
- ^ Уэбб, бакалавр; Стрэнд, MR; Дики, SE; Бек, Миннесота; Хилгарт, РС; Барни, МЫ; Кадаш, К.; Кремер, Дж. А.; Линдстрем, КГ; Раттанадечакул, В.; Шелби, Канзас; Тоеткиаттикул, Х.; Тернбулл, Миссури; Уизерелл, РА; и др. (2006). «Геномы полиднавирусов отражают их двойную роль как мутуалиста и патогена» . Вирусология . 347 (1): 160–174. дои : 10.1016/j.virol.2005.11.010 . ПМИД 16380146 .
- ^ Флеминг, Дж.-АГВ и Саммерс, доктор медицины (1991). «ДНК полиднавируса интегрирована в ДНК его паразитоидной осы-хозяина» . Труды Национальной академии наук . 88 (21): 9770–9774. Бибкод : 1991PNAS...88.9770F . дои : 10.1073/pnas.88.21.9770 . ПМК 52802 . ПМИД 1946402 .
- ^ Уитфилд, Дж. Б. (2002). «Оценка возраста симбиоза полиднавирус-браконид осы» . Труды Национальной академии наук . 99 (11): 7508–7513. Бибкод : 2002PNAS...99.7508W . дои : 10.1073/pnas.112067199 . ПМК 124262 . ПМИД 12032313 .
- ^ Перейти обратно: а б с Аннетт Рейнеке; Сассан Асгари и Отто Шмидт (2006). «Эволюционное происхождение вирусоподобных частиц Venturia canescens». Архив биохимии и физиологии насекомых . 61 (3): 123–133. дои : 10.1002/arch.20113 . ПМИД 16482583 .
- ^ Перейти обратно: а б Дрезен, Дж. М.; Леобольд, М; Безье, А; Юге, Э; Волков А.Н.; Эрниу, Э.А. (август 2017 г.). «Эндогенные вирусы ос-паразитов: вариации на общую тему». Современное мнение в вирусологии . 25 : 41–48. дои : 10.1016/j.coviro.2017.07.002 . ПМИД 28728099 .
- ^ Эрниу Э.А., Уге Э., Тезе Ж., Безье А., Перике Ж., Дрезен Ж.М. (2013). «Когда осы-паразиты захватили вирусы: геномная и функциональная эволюция полиднавирусов» . Philos Trans R Soc Lond B Biol Sci . 368 (1626): 20130051. doi : 10.1098/rstb.2013.0051 . ПМЦ 3758193 . ПМИД 23938758 .
- ^ Волков А.Н.; Жуан, В; Урбах, С; Самаин, С; Бергоэн, М; Винкер, П; Деметрт, Э; Куссеранс, Ф; Провост, Б; Кулибали, Ф; Легеай, Ф; Беливо, К; Кассон, М; Гьяпай, Г; Дрезень, Дж. М. (27 мая 2010 г.). «Анализ структурных компонентов вириона выявил остатки предкового генома ихновируса» . ПЛОС Патогены . 6 (5): e1000923. дои : 10.1371/journal.ppat.1000923 . ПМЦ 2877734 . ПМИД 20523890 .
- ^ Беливо, К; Коэн, А; Стюарт, Д; Перике, Ж; Джумад, А; Кун, Л; Штольц, Д; Бойл, Б; Волков А.Н.; Эрниу, Э.А.; Дрезен, Дж. М.; Кассон, М. (сентябрь 2015 г.). «Геномный и протеомный анализ показывает, что полиднавирусы банчина и кампоплегина имеют схожих, если не идентичных, вирусных предков» . Журнал вирусологии . 89 (17): 8909–21. дои : 10.1128/JVI.01001-15 . ПМК 4524098 . ПМИД 26085165 .
- ^ Дюпюи С., Уге Э., Дрезен Ж.М. (2006). «Развертывание истории эволюции полиднавирусов». Вирус Рес . 117 (1): 81–89. doi : 10.1016/j.virusres.2006.01.001 . ПМИД 16460826 .
- ^ Перейти обратно: а б с ДжМ. Дрезень; С. Савари; М. Пуарье; Г. Перике (1999). «Полиднавирусы, вирусные сущности, одомашненные паразитоидными осами». Вирусология . 3 (1): 11–21.
- ^ Маркус Бек и Майкл Р. Стрэнд (февраль 2005 г.). «Glc1.8 из браковируса, разрушающего микроплитис, вызывает потерю адгезии и фагоцитоза в клетках насекомых High Five и S2» . Журнал вирусологии . 79 (3): 1861–1870. дои : 10.1128/jvi.79.3.1861-1870.2005 . ПМК 544146 . ПМИД 15650210 .
- ^ Лу Чжицян; Бек Маркус Х.; Ван Ян; Цзян Хаобо; Стрэнд Майкл Р. (август 2008 г.). «Вирусный белок Egf1.0 представляет собой ингибитор двойной активности активирующих профенолоксидазу протеиназ 1 и 3 из Manduca sexta » . Журнал биологической химии . 283 (31): 21325–21333. дои : 10.1074/jbc.M801593200 . ПМК 2490783 . ПМИД 18519564 .
- ^ Пэ Сону; Ким Ёнгюн (1 сентября 2009 г.). «Гены IkB, кодируемые в Bracovirus Cotesia plutellae, подавляют противовирусный ответ и усиливают патогенность бакуловируса против бабочки-ножницы Plutella xylostella». Журнал патологии беспозвоночных . 102 (1): 79–87. дои : 10.1016/j.jip.2009.06.007 . ISSN 0022-2011 . ПМИД 19559708 .
- ^ Фатх-Гудин А.; Кремер Дж.А.; Уэбб Б.А. (август 2009 г.). «Белок ванкирина P-ванк-1 ихновируса Campoletis sonorensis ингибирует апоптоз в клетках Sf9 насекомых». Молекулярная биология насекомых . 18 (4): 497–506. дои : 10.1111/j.1365-2583.2009.00892.x . ПМИД 19453763 . S2CID 31146050 .
- ^ Клавихо Г, Доремус Т, Раваллек М, Мануччи М.А., Жуан В., Волков А.Н., Дарбу I (2011). «Мультигенные семейства ихновирусов: исследование специфичности ткани и хозяина посредством анализа экспрессии ванкиринов из Hyposoter Didymator ихновируса » . ПЛОС ОДИН . 6 (11): e27522. Бибкод : 2011PLoSO...627522C . дои : 10.1371/journal.pone.0027522 . ПМК 3210807 . ПМИД 22087334 .
- ^ Марциано НК; Хасэгава Д.К.; Фелан П.; Тернбулл М.В. (октябрь 2011 г.). «Функциональные взаимодействия между полиднавирусами и клеточными иннексинами хозяина» . Журнал вирусологии . 85 (19): 10222–9. дои : 10.1128/jvi.00691-11 . ПМК 3196458 . ПМИД 21813607 .
- ^ Перейти обратно: а б Леобольд, Матье; Безье, Энни; Пишон, Аполлина; Эрниу, Элизабет А; Волков, Анн-Натали; Дрезен, Жан-Мишель; Абергель, Шанталь (июль 2018 г.). «Одомашнивание большого ДНК-вируса осой Venturia canescens предполагает целенаправленную редукцию генома посредством псевдогенизации» . Геномная биология и эволюция . 10 (7): 1745–1764. дои : 10.1093/gbe/evy127 . ПМК 6054256 . ПМИД 29931159 .
- ^ Р.М. Ризки; ТМ Ризки (ноябрь 1990 г.). «Паразитоподобные вирусоподобные частицы разрушают клеточный иммунитет дрозофилы» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (21): 8388–8392. Бибкод : 1990PNAS...87.8388R . дои : 10.1073/pnas.87.21.8388 . ПМК 54961 . ПМИД 2122461 .
- ^ Хивнер, Мэн; Рамруп, Дж; Геген, Г; Рамраттан, Г; Долиос, Г; Скарпати, М; Квиат, Дж; Бхаттачарья, С; Ван, Р; Сингх, С; Говинд, С (25 сентября 2017 г.). «Новые органеллы с элементами бактериальных и эукариотических систем секреции служат оружием для паразитов дрозофилы» . Современная биология . 27 (18): 2869–2877.e6. дои : 10.1016/j.cub.2017.08.019 . ПМЦ 5659752 . ПМИД 28889977 .
- ^ ДиДжованни, Д; Лепети, Д; Гине, Б; Беннетот, Б; Булестей, М; Куте, Ю; Буше, О; Раваллек, М; Варальди, Дж. (1 октября 2020 г.). «Родственник вируса, манипулирующего поведением, как источник адаптивных генов для паразитоидов дрозофилы» . Молекулярная биология и эволюция . 37 (10): 2791–2807. bioRxiv 10.1101/342758 . дои : 10.1093/molbev/msaa030 . ПМИД 32080746 .
- ^ Ли, HW и Дин, SW (2005). «Противовирусное молчание у животных» . ФЭБС Летт . 579 (26): 5965–5973. дои : 10.1016/j.febslet.2005.08.034 . ПМК 1350842 . ПМИД 16154568 .
- ^ Безье А, Аннахайм М, Эрбиньер Ж, Веттервальд С, Гьяпай Г, Бернар-Самен С, Винкер П, Родити И, Хеллер М, Бельгази М, Пфистер-Вильхем Р, Перике Г, Дюпюи С, Юге Е, Волков А.Н., Ланцрейн Б, Дрезень Дж.М. (13 февраля 2009 г.). «Полиднавирусы ос-браконид происходят от предкового нудивируса» . Наука . 13. 323 (5916): 926–30. Бибкод : 2009Sci...323..926B . дои : 10.1126/science.1166788 . ПМИД 19213916 . S2CID 6538583 .
- Управление ICTVdB (2006). 00.055. Полиднавирусиды. В: ICTVdB — Универсальная база данных вирусов, версия 4. Бюхен-Осмонд, К. (ред.), Колумбийский университет, Нью-Йорк, США.
- Эспань, Э.; и др. (2004). «Последовательность генома полиднавируса: взгляд на эволюцию симбиотического вируса» . Наука . 306 (5694): 286–289. Бибкод : 2004Sci...306..286E . дои : 10.1126/science.1103066 . ПМИД 15472078 . S2CID 12260572 .