Перо

Перья -это эпидермальные росты, которые образуют отличительное внешнее покрытие или оперение , как на птичьей (птице), так и на некоторых неавийских динозаврах и других археозаврах . Они являются самыми сложными покровными структурами, обнаруженными у позвоночных [ 1 ] [ 2 ] и пример сложной эволюционной новизны. [ 3 ] Они являются одними из характеристик, которые отличают существующих птиц от других живых групп. [ 4 ]
Хотя перья покрывают большую часть тела птицы, они возникают только из некоторых четко определенных участков на коже. Они помогают в полете, теплоизоляции и гидроизоляции. Кроме того, окраска помогает в связи и защите . [ 5 ] Изучение перьев называется плумология (или наука о оперении ). [ 6 ] [ 7 ]
Люди используют перья во многих отношениях, которые являются практическими, культурными и религиозными. Перья являются мягкими и отличными для захвата тепла ; Таким образом, они иногда используются в постельных принадлежностях высокого класса , особенно на подушках , одеялах и матрасах . Они также используются в качестве начинки для зимней одежды и открытых постельных принадлежностей, таких как стеганые пальто и спальные мешки . Goose и Eider Down имеют отличный чердак , способность расширяться из сжатого, хранящегося состояния, чтобы захватить большое количество компартментализированного, изолирующего воздуха. [ 8 ] Перья крупных птиц (чаще всего гусей ) были и используются для приготовления перо . Исторически, охота на птиц на декоративные и декоративные перья подвергала угрозу некоторым видам и помогла внести свой вклад в вымирание других. [ 9 ] Сегодня перья, используемые в моде и в военных головных узерах и одежде, получены как отходы птицеводства, включая цыплят , гусей , индеек , фазанов и страуса . Эти перья окрашиваются и манипулируют, чтобы усилить их внешний вид, поскольку птичья перья, естественно, часто скучны по внешнему виду по сравнению с перьями диких птиц.
Этимология

- Иметь
- Вал, Рахис
- Обновой
- Фафл, после фарма
- Quill, Каламус
Перо происходит от старого английского «feþer», который имеет германское происхождение; Связано с голландским «Veer» и немецким «федеральным» из индоевропейского корня, разделяемого «патрой» Санскрита, означающим «крыло», латинское «Пенна», означающее «перьев» и «Птерон» Грека, «Птерокс». '
Из -за того, что перья, являющиеся неотъемлемой частью переноек , которые были ранними ручками, использованными для письма, сама ручка получена из латинской пенны , что означает перьев. [ 10 ] Французский шлейф может означать перо , перо или ручку .
Структуры и характеристики

Перья являются одними из наиболее сложных покровных придаток, обнаруженных у позвоночных и образуются в крошечных фолликулах в эпидермисе , или наружном слое кожи, которые продуцируют кератиновые белки . Β -кератины в перьях, клювах и когтях -и когти, масштабах и оболочки рептилий , -состоит из водородно-водородных цепей связанных с β-считанными листами , которые затем скручены и сшиваются мостами дисульфидными мостами в структуры еще более жестче, которые затем скручены и сшиваются дисульфидными в структуры. α -кератины млекопитающих волос , рога и копыта . [ 11 ] [ 12 ] Точные сигналы , которые вызывают рост перьев на коже, неизвестны, но было обнаружено, что фактор транскрипции Cdermo-1 вызывает рост перьев на коже и масштабах на ноге. [ 13 ]
Классификация


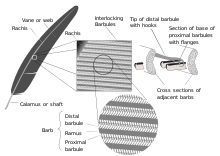
Существует два основных типа перьев: фунополированные перья, которые покрывают внешнюю часть тела, и вниз по перьям , которые находятся под перьями. Пенжевые перья - перья. Также называемые контурными перьями, ножными перьями возникают из трактов и покрывают все тело. Третий режарный тип пера, филоплимы , волосы рода и тесно связаны с пенсисными перьями и часто полностью скрыты, с одним или двумя филопумами, прикрепленными и прорастающими из той точки кожи, что и каждое пеннасовое перьев, по крайней мере, перо, по крайней мере, перо, по крайней мере На головой птицы, шеи и туловище. [ 14 ] [ 15 ] Филопумы полностью отсутствуют в Ratites . [ 16 ] В некоторых пробинах филоплимы возникают, выставленные за ножными перьями на шее. [ 1 ] Ремиги, или перья полета крыла, а также первые перья или перья летательных перевозок являются наиболее важными перьями для полета. Типичное перьев Vaned имеет основной вал, называемый Rachis . Обеспеченные на рахисе, представляют собой серию ветвей или колючек ; Сами колючки также разветвляются и образуют уарлы . Эти борчаты имеют мельчайшие крючки, называемые Барбицелями для перекрестного привязки. Вниз по перьям пушистые, потому что им не хватает барбицелей, поэтому у барбов плавают свободно друг от друга, что позволяет ловушке воздуха и обеспечивает отличную теплоизоляцию. У основания пера рахис расширяется, образуя полого канальца (или перо ), который вставляется в фолликул в коже . Базальная часть каламуса без лопастей. Эта часть встроена в фолликул кожи и имеет отверстие в основании (проксимальный пупок) и небольшое отверстие на стороне (дистальный пупок). [ 17 ]
Птицы из каких -либо видов имеют особый вид натала вниз по перьям (неосоптилям), которые выталкиваются, когда появляются нормальные перья (телеоптилы). [ 1 ]
Печатные перья усилены, чтобы работать против воздуха вниз, но уступил в других направлениях. Было отмечено, что характер ориентации β-кератиновых волокон в перьях летающих птиц отличается от таблицы у нелетающих птиц: волокна лучше выровнены вдоль направления оси вала в направлении наконечника, [ 18 ] [ 19 ] и боковые стены в районе Рахис демонстрируют структуру скрещенных волокон. [ 20 ] [ 21 ]
Функции
Перья изолируют птиц от воды и холодных температур. Они также могут быть сорваны, чтобы выровнять гнездо и обеспечить изоляцию яиц и молодых. Отдельные перья в крыльях и хвосте играют важную роль в управлении полетом. [ 20 ] У некоторых видов есть гребень перьев на головах. Хотя перья легкие, оперение птиц весит в два или три раза больше, чем в его скелете, поскольку многие кости являются пустыми и содержат воздушные мешки. Цветовые узоры служат камуфляжем против хищников для птиц в их среде обитания и служат камуфляжем для хищников, ищущих еду. Как и в случае с рыбой, верхние и нижние цвета могут быть разными, чтобы обеспечить камуфляж во время полета. Поразительные различия в узорах и цветах перьев являются частью сексуального диморфизма многих видов птиц и особенно важны при выборе спаривающихся пар. В некоторых случаях существуют различия в ультрафиолетовой отражательной способности перьев в полах, даже если в видимом диапазоне не отмечается различия в цвете. [ 22 ] Крыловые перья мужского клубного класса Manakins Machaeropterus deliciosus имеют специальные структуры, которые используются для изготовления звуков путем стридуляции . [ 23 ]

У некоторых птиц есть запас порошка вниз по перьям, которые растут непрерывно, с небольшими частицами, регулярно отрывающимися от концов шишки. Эти частицы производят порошок , который просеивает перья на теле птицы и действует как гидроизоляционный агент и кондиционер с перьями . Порошок вниз развивался независимо в нескольких таксонах и может быть найден вниз, а также у нористы. Они могут быть разбросаны в оперении, как у голубей и попугаев, или на локализованных участках на груди, животе или по бокам, как у цапель и лягушек. Цериноны используют свой счет, чтобы сломать порошок вниз по перьям и распределить их, в то время как какаду могут использовать свою голову в качестве порошкового слойки, чтобы нанести порошок. [ 24 ] Гидроизоляция может быть потеряна за счет воздействия эмульгирующих агентов из -за загрязнения человека. Затем перья могут стать заболоченными, заставляя птицу тонуть. Также очень трудно чистить и спасать птиц, перья которых были загрязнены разливами нефти . Перья бакланов впитывают воду и помогают уменьшить плавучесть, что позволяет птицам плавать погружение. [ 25 ]

Компания жесткие, сужающиеся перья с большими рахисами, но немного зазубрин. Риктальные щетинки встречаются вокруг глаз и Билла. Они могут служить аналогичной целью с ресницами и вибрациями у млекопитающих . Несмотря на то, что пока нет четких доказательств, было высказано предположение, что риктальные щетинки имеют сенсорные функции и могут помочь насекомым птицам захватить добычу. [ 26 ] В одном исследовании было обнаружено, что мухоловка Willow ( Empidonax Traillii ) ловит насекомых до столь же до и после удаления риктовых щетинок. [ 27 ]
Гребс своеобразны в их привычке приглашать свои перья и кормить их молодым. Наблюдения за их рационом по рыбе и частоте питания перьев предполагают, что проглатывание перьев, особенно вниз по бокам, помогает формировать легко выброшенные гранулы. [ 28 ]
Распределение

Контурные перья не равномерно распределены на коже птицы, за исключением некоторых групп, таких как пингвины , катисты и крикулы. [ 29 ] У большинства птиц перья растут из определенных участков кожи, называемых птеролами ; Между Pterylae есть области, которые свободны от перьев, называемых Apterylae (или Apteria ). Филопумы и вниз могут возникнуть из Apterylae. Расположение этих перьев, птерилоза или птерилографии различается по семействам птиц и использовалось в прошлом в качестве средства для определения эволюционных отношений семейств птиц. [ 30 ] [ 31 ] Виды, которые инкубируют свои собственные яйца, часто теряют свои перья в области живота, образуя задумчивый участок . [ 32 ]
Окраска

Слева: Турацин (красный) и Тураковердин (зеленый, с некоторой структурной голубым переливанием на нижнем конце) на крыле Таурако Баннермани
Справа: каротиноиды (красный) и меланины (темные) на животе/крыльям Ramphocelus bresilius
Цвета перьев производятся пигментами, микроскопическими конструкциями, которые могут преломлять , отражать или разбросать выбранные длина волн света или комбинацией обоих.
Большинство пера -пигментов - меланины (коричневые и бежевые феомеланины , черные и серые эймеланины ) и каротиноиды (красный, желтый, оранжевый); Другие пигменты встречаются только в определенных таксонах - от желтого до красного псаттакофулвина [ 33 ] (найдено в некоторых попугаях ) и красный турацин и зеленый тураковердин ( пигменты порфирина, обнаруженные только в Туракос ).
Структурная окраска [ 5 ] [ 34 ] [ 35 ] участвует в производстве синих цветов, переливаемости , наиболее ультрафиолетовой отражательной способности и улучшения пигментальных цветов. Сообщалось о структурной переливании [ 36 ] в ископаемых перьях, начиная с 40 миллионов лет. У белых перьев не хватает пигмента и рассеивания света диффузно; Альбинизм у птиц вызван дефектной производством пигментов, хотя структурная окраска не будет затронута (как можно увидеть, например, в сине-белых буджеригарах ).

Блюз и яркая зелень многих попугаев производятся путем конструктивного интерференции света, отражающегося от разных слоев структур в перьях. В случае зеленого оперения, в дополнение к желтому, конкретная структура перья вызывается некоторыми текстурой DYCK. [ 37 ] [ 38 ] Меланин часто участвует в поглощении света; В сочетании с желтым пигментом он производит скучный оливково-зеленый.

У некоторых птиц цветы перьев могут быть созданы или изменены выделениями из уропигиальной железы , также называемой Preen Rand. Желтые цвета счетов многих рогатых зажиганий производятся такими выделениями. Было высказано предположение, что существуют другие цветовые различия, которые могут быть видны только в ультрафиолетовой области, [ 24 ] Но исследования не смогли найти доказательства. [ 39 ] Секреция нефти из уропигиальной железы также может оказывать ингибирующее влияние на перьевые бактерии. [ 40 ]
Красные, оранжевые и желтые цвета многих перьев вызваны различными каротиноидами. Пигменты на основе каротиноидов могут быть честными сигналами пригодности, потому что они получены из специальных диет и, следовательно, могут быть трудно получить, [ 41 ] [ 42 ] и/или потому, что каротиноиды необходимы для иммунной функции и, следовательно, сексуальные показы происходят за счет здоровья. [ 43 ]
Птичьи перья подвергаются износу и периодически заменяются в течение жизни птицы через лишение . Новые перья, известные при развитии как кровь, или перья для штифтов , в зависимости от стадии роста, образуются через те же фолликулы, из которых старые лежали. Присутствие меланина в перьях повышает их сопротивление истиранию. [ 44 ] В одном исследовании отмечается, что перья на основе меланина быстрее разлагались при бактериальном действии, даже по сравнению с неправленными перьями из одного и того же вида, чем те, которые не раскрыты или с каротиноидными пигментами. [ 45 ] Однако в другом исследовании в том же году сравнивалось действие бактерий на пигментации двух видов Song Sparrow и наблюдалось, что более темные пигментированные перья были более устойчивыми; Авторы процитировали другие исследования, также опубликованные в 2004 году, в которых говорилось, что увеличение меланина обеспечило большую устойчивость. Они заметили, что большее сопротивление темных птиц подтвердило правило Глогера . [ 46 ]
Хотя половой отбор играет важную роль в развитии перьев, в частности, цвет перьев это не единственный доступный вывод. Новые исследования предполагают, что уникальные перья птиц также оказывают большое влияние на многие важные аспекты поведения птиц, такие как высота, на которой разные виды строят свои гнезда. Поскольку женщины являются основными лицами, осуществляющими уход, Evolution помогла отбирать женщин, чтобы показать более тупые цвета, чтобы они могли смешаться в среде гнездования. Положение гнезда и есть ли у него больше шансов на пребывание в хищнике, наложило ограничения на оперение птиц -птиц. [ 47 ] Вид птицы, который гнездятся на земле, а не навес деревьев, должен иметь много тупых цветов, чтобы не привлекать внимание к гнезду. Исследование высоты показало, что птицы, которые гнездятся в навесах деревьев, часто имеют гораздо больше хищных атак из -за более яркого цвета перьев, которые демонстрирует самка. [ 47 ] Другое влияние эволюции, которое может сыграть роль в том, почему перья птиц настолько красочны и демонстрируют так много узоров, что птицы развили свои яркие цвета от растительности и цветов, которые процветают вокруг них. Птицы разрабатывают свои яркие цвета от жизни вокруг определенных цветов. Большинство видов птиц часто смешиваются в их окружающую среду, из -за некоторой степени камуфляжа, поэтому, если среда обитания видов полна цветов и узоров, это виды в конечном итоге развивались, чтобы смешать, чтобы избежать употребления. Птицы перья показывают большой ассортимент цветов, даже превышающие разнообразие многих растений, листьев и цветов. [ 48 ]
Паразиты
Поверхность перья является домом для некоторых эктопаразитов, особенно перья вшами ( phthiraptera ) и перьев. Вши с перьями обычно живут на одном хозяине и могут перемещаться только от родителей к цыплятам, между спаривающимися птицами и, иногда, Форези . Эта история жизни привела к тому, что большинство видов паразитов специфичны для хозяина и совместно с хозяином, что делает их интересными в филогенетических исследованиях. [ 49 ]
Пеховые отверстия - это следов вшей (скорее всего, Brueelia spp. Вшей) на крыло и хвостовых перьях. Они были описаны на ласточках сараев , и из -за легкой счетности многие эволюционные, экологические и поведенческие публикации используют их для количественной оценки интенсивности заражения.
Паразитические кукурузы, которые растут в гнездах других видов, также имеют специфические для хозяина вшей, и они, кажется, передаются только после того, как молодые кукушки покинут гнездо хозяина. [ 50 ]
Птицы поддерживают состояние перья, притягая и купая в воде или пыли . Было высказано предположение, что своеобразное поведение птиц, антинг , в котором муравьи вводятся в оперение, помогает уменьшить паразиты, но никаких подтверждающих доказательств не было обнаружено. [ 51 ]
Использование человека
Утилитарный


перья давно использовались для стрел . Птичьи Красочные перья, такие как те, которые принадлежат к фазанам, использовались для украшения приманки для рыбалки .
Перья также ценны в том, чтобы помочь идентификации видов в судебно -медицинских исследованиях, особенно при ударах птиц в самолет. Соотношения изотопов водорода в перьях помогают определить географическое происхождение птиц. [ 52 ] Перья также могут быть полезны в неразрушающей выборке загрязняющих веществ. [ 53 ]
Птичья промышленность производит большое количество перьев в виде отходов, которые, как и другие формы кератина, медленно разлагаются. Перовые отходы использовались в ряде промышленных применений в качестве среды для культивирования микробов, [ 54 ] биоразлагаемые полимеры, [ 55 ] и производство ферментов. [ 56 ] Белки перья были опробованы как клей для деревянной доски. [ 57 ]
Некоторые группы коренных жителей на Аляске использовали перья Ptarmigan в качестве характера (непластичные добавки) в производстве керамики со времен первого тысячелетия до н.э., чтобы способствовать сопротивлению и прочности теплового шока. [ 58 ]
В религии и культуре

Орлиные перья имеют большую культурную и духовную ценность для коренных американцев в Соединенных Штатах и первых наций народах в Канаде как религиозные объекты. В Соединенных Штатах религиозное использование перьев орла и ястребов регулируется законом о орлиных перьях , федеральным законом, ограничивающим владение орлиными перьями сертифицированными и зачисленными членами федерально признанных коренных американских племен.
В Южной Америке варева, изготовленные из перьев кондоров, используются в традиционных лекарствах. [ 59 ] В Индии перья индийского павлина использовались в традиционной медицине для укуса змеи, бесплодия и кашля. [ 60 ] [ 61 ]
Известно, что члены Шотландии Кэмпбелл носят перья на своих капотах, чтобы обозначить власть в клане. Клановые вожди носят три, вождь носят два, а Армигер носит один. Любой член клана, который не соответствует критериям, не уполномочен носить перья как часть традиционной одежды, и это считается самонадеянным. [ 62 ]
В течение 18 -го, 19 -го и начала 20 -го веков произошла быстро растущая международная торговля в перья для экстравагантных женских шляп и других головных уборов (в том числе викторианской модой ). В 1886 году Фрэнк Чепмен отметил, что перья до 40 видов птиц использовались примерно в трех четвертях из 700 женских шляп, которые он наблюдал в Нью-Йорке. [ 63 ] Например, в прошлом были использованы перья колибри в Южной Америке , чтобы одеть некоторых миниатюрных птиц, представленных в поющих птичьих коробках . Эта сделка вызвала серьезные потери для популяций птиц (например, цаплы и копейки ). Защитники возглавляли крупную кампанию против использования перьев в шляпах. Это способствовало принятию Закона о Лейси в 1900 году и изменениям в моде. Рынок декоративных перьев затем в значительной степени рухнул. [ 64 ] [ 65 ]
Совсем недавно оперение Rooster стало популярной тенденцией в качестве аксессуара для прически , причем перья, ранее используемые в качестве рыболовных приманков, теперь используются для обеспечения цвета и стиля для волос. [ 66 ]
Производство перьев в Европе сократилось за последние 60 лет, в основном из -за конкуренции со стороны Азии. Перья украшали шляпы на многих престижных мероприятиях, таких как свадьбы и дамы в День в Racecourses (Royal Ascot).
Эволюция


Функциональные соображения
Функциональный взгляд на эволюцию перьев традиционно фокусировался на изоляции, полете и дисплее. Открытия не летящих поздних меловых пернатых динозавров в Китае, [ 67 ] Тем не менее, предполагайте, что полете не мог быть исходной основной функцией, поскольку перья просто не были бы способны обеспечить какую -либо форму подъема. [ 68 ] [ 69 ] Были предположения о том, что перья, возможно, имели свою первоначальную функцию в терморегуляции, гидроизоляции или даже в качестве раковин для метаболических отходов, таких как серная. [ 70 ] Утверждается, что недавние открытия поддерживают терморегуляторную функцию, по крайней мере, в небольших динозаврах. [ 71 ] [ 72 ] Некоторые исследователи даже утверждают, что терморегуляция возникла из -за щетины на лице, которые использовались в качестве тактильных датчиков. [ 73 ] В то время как перья были предложены как развивающиеся из рептилийских масштабов , существуют многочисленные возражения против этой идеи, и более поздние объяснения возникли в результате парадигмы эволюционной биологии развития . [ 2 ] Теории основанных на масштабе происхождения перьев предполагают, что плоская структура масштаба была изменена для развития на перья путем расщепления, чтобы сформировать лямки; Однако этот процесс развития включает в себя трубчатую структуру, возникающую из фолликула и продольного расщепления трубки для формирования лямки. [ 1 ] [ 2 ] Количество перьев на единицу площади кожи выше у мелких птиц, чем у более крупных птиц, и эта тенденция указывает на их важную роль в теплоизоляции, поскольку мелкие птицы теряют больше тепла из -за относительно большей площади поверхности пропорционально их массу тела Полем [ 5 ] Миниатюризация птиц также сыграла свою роль в эволюции полета с питанием. [ 74 ] Считается, что окраска перьев развивалась главным образом в ответ на половой отбор . В ископаемых образцах паравии Anciornis Huxleyi и Pterosaur Tupandactylus Imperator , признаки настолько хорошо сохраняются, что меланосом можно наблюдать структуру цвет и рисунок перьев на анщорнисе и tupandactylus . (пигментные клетки). Сравнивая форму ископаемых меланосомов с меланосомами от существующих птиц, можно определить [ 75 ] [ 76 ] Было обнаружено, что у Анчиорниса были перья с черно-белыми пальцами на передних конечностях и задних конечностях, с красновато-коричневым гребнем. Этот рисунок похож на окраску многих существующих видов птиц, которые используют окраску оперения для отображения и общения, включая половой отбор и камуфляж. Вполне вероятно, что неавийские виды динозавров использовали паттерны оперения для оперения для аналогичных функций, что и современные птицы до происхождения полета. Во многих случаях физиологическое состояние птиц (особенно мужчин) обозначается качеством их перьев, и это используется (женщинами) в выборе партнера . [ 77 ] [ 78 ] Кроме того, при сравнении различных образцов Ornithomimus edmontonicus , пожилых людей, обнаружили, что имеют пеннибрихиум (похожая на крыло структура, состоящая из удлиненных перьев), а более молодые-нет. Это говорит о том, что Pennibrachium был вторичным половым характеристиком и, вероятно, имел сексуальную функцию. [ 79 ]
Молекулярная эволюция
несколько генов Было обнаружено, что определяют развитие перьев. Они будут ключом к пониманию эволюции перьев. Например, некоторые гены преобразуют масштабы в перья или похожие на перья, когда выражаются или индуцируются в ногах птиц, такие как преобразователи для шкалы SOX2 , ZIC1 , GREM1 , SPRY2 и SOX18 . [ 80 ]
Перья и весы состоят из двух отдельных форм кератина , и долго думали, что каждый тип кератина был эксклюзивным для каждой структуры кожи (перья и весы). Тем не менее, перо кератин также присутствует на ранних стадиях развития американских масштабов аллигатора . Этот тип кератина, ранее считался специфическим для перьев, подавляется во время эмбриологического развития аллигатора и поэтому не присутствует в масштабах зрелых аллигаторов. Присутствие этого гомологичного кератина как у птиц, так и у крокодилов указывает на то, что он был унаследован от общего предка. [ 81 ]
птерозавров Это может свидетельствовать о том, что крокодильские чешуйки, птицы и фотозащиты и пикнофибры - все это выражения в области развития одних и тех же примитивных кожных структур архозавра; предполагая, что перья и пикноволокны могут быть гомологичными. [ 82 ] Методы молекулярного датирования в 2011 году показывают, что подсемейство β-кератинов перьев, обнаруженных у существующих птиц, начало расходиться с 143 миллионами лет назад, предполагая, что переночные перья Анчиорниса не были сделаны из перо β-кератинов, присутствующих у существующих птиц. [ 83 ] Тем не менее, исследование ископаемых перьев из динозавра sinosauropteryx и других окаменелостей выявило следы бета-листовых белков, используя инфракрасную спектроскопию и спектроскопию серы-x-ray. Было показано, что наличие обильных альфа-белков в некоторых ископаемых перьях является артефактом процесса ископаемости, поскольку структуры бета-белка легко изменяются на альфа-спирали во время теплового деградации. [ 84 ] В 2019 году ученые обнаружили, что гены для производства перьев эволюционировали у основания Архосаврии, подтверждая, что перья присутствовали на ранних орнитодиранах и согласуются с ископаемым записи. [ 85 ]
Пернатые динозавры

У нескольких неавийских динозавров были перья на своих конечностях, которые не работали бы для полета. [ 67 ] [ 2 ] Одна теория предполагает, что перья изначально развивались на динозаврах из -за их изоляционных свойств; Затем небольшие виды динозавров, которые росли более длинными перьями, могли найти их полезными в скольжении, что приводит к эволюции прото-птиц, таких как Archeopteryx и Microraptor Zhaoianus . Другая теория утверждает, что первоначальным адаптивным преимуществом ранних перьев была их пигментация или переливаемость, способствующая сексуальным предпочтениям при выборе партнеров. [ 86 ] Динозавры, у которых были перья или протофичи, включают Pedopenna daohugouensis [ 87 ] и Delong Paradoxus , тиранозавра , который на 60-70 миллионов лет старше, чем Tyrannosaurus rex . [ 88 ]
У большинства динозавров, как известно, имели перья или протофичи, являются теропод , однако, похожие на перьев «нитчатые покровные структуры», также известны из орнитишских динозавров Тяньюлонга и Пситтакозавра . [ 89 ] Точная природа этих структур все еще изучается. Тем не менее, считается, что перья стадии 1 (см. Раздел эволюционных этапов ниже), такие как те, которые наблюдаются в этих двух орнитишах, вероятно, функционировали на дисплее. [ 90 ] Ornithischian Kulindadromeus , которые имели структуры, напоминающие перья стадии 3. В 2014 году, как сообщалось, [ 91 ] Вероятность того, что весы, развивающиеся на ранних предках динозавров, высока. Однако это было предполагающим, что примитивные птерозавры были чешуйчаты. [ 92 ] [ 93 ] В исследовании 2016 года анализируется морфология мякоти хвостовой щетины Psittacosaurus и обнаруживает, что они похожи на перья, но отмечает, что они также похожи на щетину на голове павпочки Конго , бороду индейки и позвоночнику на голове рогатого крикера . [ 94 ] Повторная ретимеция максимальной вероятности палеонтолога Томаса Хольца обнаруживает, что филаменты с большей вероятностью станут наследственным состоянием динозавров. [ 95 ]
В 2010 году у Carcharodontosaurid под названием Congcovatus было обнаружено, что была установлена установка на локтевой кости, предполагая, что у нее могут быть капитальные структуры на AMS. [ 96 ] Однако Foth et al. В 2014 году раздается публикации, где они указывают на то, что удары на локте -конъекторе находятся на переднелатеральной части , которая не похожа на REMIGES, которые находятся в постерататеральной на локте некоторых птиц, они считают, что это более вероятно, что это привязанности к межсборным связям. [ 97 ] Это было опровергнуто Куэста Фидальго и ее коллегами, они указали, что эти удары на локтевой кости являются постератолатеральными, которые не похожи на межзнательные связки. [ 98 ]
были обнаружены десятки пернатых динозавров С 1990 -х годов в кладе Манираптора , в которую входят клада Avialae и недавних общих предков птиц, овираптозавейров и deinonychosauria . В 1998 году открытие пернатого овираптозавриана Caudipteryx Zoui бросило вызов понятию перьев как структуры, исключительной для Avialae. [ 99 ] Похоронен в формировании Йикса в Ляонинге, Китай, К. Зуи жил в начале мелового периода. Присутствует на передних конечностях и хвостах, их покровная структура была принята [ кем? ] как пенсиозные перья на основе рахиса и рисунка херрингбоун колючи. В Clade Deinonychosauria продолжающееся расхождение перьев также очевидно в семьях Troodontidae и Dromaeosauridae . Распространенные перья с рахисом, колючками и барбами были обнаружены во многих членах, включая Sinornithosaurus millenii , Dromaeosaurid, обнаруженный в формации Yixian (124,6 млн лет). [ 100 ]
Ранее в эволюции перьев существовал временный парадокс-лечебцы с высокоэтальными птицами, похожими на птицами, произошли в более позднее время, чем археоптерикс, -поступив, что потомки птиц возникли перед предком. Тем не менее, открытие Анчиорниса Хакслии в формировании поздней юры Тиаоджишан (160 млн лет) в Западном Ляонинге в 2009 году [ 101 ] [ 102 ] решил этот парадокс. Предыдущая археоптерикс , Анчиорнис доказывает существование современного пернатым предком Теропода, предоставляя представление о переходе динозавр-птицы. Образец показывает распределение больших пеншетных перьев на передних конечностях и хвосте, что подразумевает, что пенистые перья распространяются на остальную часть тела на более ранней стадии в эволюции теропода. [ 103 ] Развитие новичных перьев не заменило более ранних нитевидных перьев. Неснатных перьев сохраняются вместе с современными перьями полета, в том числе с некоторыми с модификациями, обнаруженными в перьях существующих птиц, в 80 миллионов лет Амбер из Альберты. [ 104 ]
Два маленьких крыла, попавших в янтарную датировку до 100 млн лет, существовали в некоторых предшественниках птиц. Крылья, скорее всего, принадлежали Enantiornithes , разнообразной группе птичьих динозавров. [ 105 ] [ 106 ]
Большой филогенетический анализ ранних динозавров Мэтью Барона, Дэвида Б. Нормана и Пола Барретта (2017) обнаружил, что Theropoda на самом деле более тесно связана с Ornithischia , с которой она сформировала сестринскую группу в пределах Clade Ornithoscelida . Исследование также показало, что если перьев, похожие на структуры теропод и орнитишхов, имеют общее эволюционное происхождение, то было бы возможно, что перья были ограничены Ornithoscelida. Если это так, то происхождение перьев, вероятно, произошло бы еще в среднем триасе , [ 107 ] Хотя это было не согласено. [ 108 ] [ 109 ] Отсутствие перьев, присутствующих в крупных сауроподах и анкилозаврах, может заключаться в том, что перья были подавлены геномными регуляторами. [ 110 ]
Эволюционные этапы

Несколько исследований развития перьев у эмбрионов современных птиц, в сочетании с распределением типов перьев среди различных доисторических предшественников птиц, позволили ученым попытаться реконструкции последовательности, в которой перья сначала развивались и превратились в типы, обнаруженные на современных птицах.
Эволюция перьев была разбита на следующие этапы Сюй и Го в 2009 году: [ 90 ]
- Одиночная нить
- Многочисленные нити соединены на их базе
- Многочисленные нити соединены у их базы в центральную нить
- Многочисленные нити по длине центральной нити
- Многочисленные филаменты, возникающие из края мембранной структуры
- Пеннсенозное перо с лопатом колючек и барбов и центральным рахисом
- Пеннашесное перо с асимметричным рахисом
- Недифференцированный лодок с центральным рахисом
Тем не менее, Foth (2011) показал, что некоторые из этих предполагаемых этапов (в частности, этапах 2 и 5), вероятно, являются просто артефактами сохранения, вызванными тем, как ископаемые перья раздавлены, а перья остаются или отпечатки. Фот переосмысленные перья стадии 2 как раздавленные или неверно идентифицированные перья, по крайней мере, стадии 3, и перья 5 стадии 5 в виде перьев измельченных стадий 6. [ 111 ]
Следующая упрощенная диаграмма взаимосвязи динозавров следует по этим результатам и показывает вероятное распределение плюмасовых (пухлых) и пенсиссовых (Vaned) перьев среди динозавров и доисторических птиц. Диаграмма следует за тем, что представлен Сюй и Го (2009) [ 90 ] Модифицировано с результатами Foth (2011) [ 111 ] Числа, сопровождающие каждое имя, относятся к наличию конкретных этапов перья. Обратите внимание, что «S» указывает на известное присутствие масштабов на теле.
В птерозаврах
Давно известно, что птерозавры имеют нитевидные структуры, покрывающие их тело, известные как пикнофибристы , которые, как правило, считались отличными от «истинных красителей» птиц и их родственников динозавров. Тем не менее, исследование двух небольших, хорошо сохранившихся окаменелостей птерозавров из юрского периода внутренней Монголии , Китай указал, что птерозавры были покрыты в массивах различных структурированных пикнофибриев (а не просто нитевидные), с несколькими из этих структур, демонстрирующих диагностику. Особенности перьев, таких как неэверные сгруппированные нити и двусторонние филаменты, оба из которых первоначально считались исключительными для птиц и других манирапторанских динозавров. Учитывая эти выводы, возможно, что перья имеют глубокое эволюционное происхождение у наследственных архозавров , хотя существует также вероятность того, что эти структуры независимо развивались, чтобы напоминать птичьи перья посредством сходящейся эволюции . [ 112 ] Майк Бентон, старший автор исследования, одолжил бывшую теорию, заявив, что «мы не могли найти никаких анатомических доказательств того, что четыре типа пикноволока каким -либо образом отличаются от перьев птиц и динозавров. Поэтому, потому что они одинаковы Они должны разделить эволюционное происхождение, и это было около 250 миллионов лет назад, задолго до происхождения птиц ». [ 113 ] [ 114 ] [ 115 ] [ 116 ] Но покровные структуры образцов анурогнатидов по -прежнему основаны на грубой морфологии, как указала Лилиана д'Альба. Пикнофибры двух образцов анурогнатидов могут не быть гомологичными с нитевидными придатками к динозаврам. [ 117 ] Пол М. Барретт подозревает, что во время интенсивной эволюции птерозавров птерозавры примитивно потеряли масштабы, а пикноволокны начали появляться. [ 93 ]
Cascocauda была почти полностью покрыта обширным слоем пикнофибриков, которые, по -видимому, происходили в двух типах. Первые являются простыми, изогнутыми нитями, которые в длину варьируются от 3,5 до 12 мм. Эти нити покрывают большую часть животного, включая голову, шею, тело, конечности и хвост. Второй тип состоит из пучков филаментов, соединенных вблизи основания, аналогично разветвленным перьям птиц и других колурозаврных динозавров, длиной около 2,5–8,0 мм и покрывают только мембраны крыла. Исследования отобранных пикнофибридов выявили наличие микрободов в нитях, напоминающих пигменты меланосом, идентифицированные в других ископаемых покровных, в частности, феомеланосомы. Кроме того, инфракрасный спектральный анализ этих пикнофибриров показывает сходные спектры поглощения с красными человеческими волосами . Эти пикнофибры, вероятно, обеспечивали как изоляцию и, возможно, помогли оптимизировать тело и крылья во время полета. [ 118 ] Идентичность этих ветвящихся структур в качестве пикнофибриров или перьев была оспорена Unwin & Martill (2020), который интерпретировал их как сгруппированные и деградированные актинофибрилл-наживающие волокна, обнаруженные в мембране крыла с скорее кожи птерозавров и приписывали меланосомы и кератин с кожей в кожу и кожу, а также приписывали меланосомы и кератин в кожу и кератин. чем нити. [ 119 ] Эти претензии были опровергнуты Ян и коллегами, которые утверждают, что интерпретации Unwin и Martill не соответствуют сохранению образца. А именно, они утверждают, что последовательная структура, регулярное расстояние и расширение филаментов за мембраной крыла подтверждают их идентификацию как пикнофибры. Кроме того, они утверждают, что ограничение меланосом и кератина к волокнам, как это происходит в перьях ископаемых динозавров, подтверждает случай, когда они являются филаментами и не согласуются с загрязнением со консервированной кожи. [ 120 ] Протоферы, вероятно, развивались в ранних архизаврах, вскоре после события вымирания PT в течение времени метаболизма ранних архозавров и синапсидов увеличивались, позы становятся прямыми и устойчивыми активностью. [ 85 ]
Смотрите также
Ссылки
- ^ Подпрыгнуть до: а беременный в дюймовый Prum, Richard O .; Ah Brush (2002). «Эволюционное происхождение и диверсификация перьев» (PDF) . Ежеквартальный обзор биологии . 77 (3): 261–295. doi : 10.1086/341993 . PMID 12365352 . S2CID 6344830 . Архивировано (PDF) из оригинала 29 июня 2011 года . Получено 7 июля 2010 года .
- ^ Подпрыгнуть до: а беременный в дюймовый Prum, Ro & Brush, Ah (март 2003 г.). "Что произошло первым, перо или птица?" (PDF) . Scientific American . 288 (3): 84–93. Bibcode : 2003sciam.288c..84p . doi : 10.1038/Scientificamerican0303-84 (неактивная 30 июня 2024 г.). PMID 12616863 . Архивировано (PDF) из оригинала 29 июня 2011 года . Получено 7 июля 2010 года .
{{cite journal}}: CS1 Maint: doi неактивен по состоянию на июнь 2024 года ( ссылка ) - ^ Prum, Richard O (1999). «Развитие и эволюционное происхождение перьев» (PDF) . Журнал экспериментальной зоологии Часть B: молекулярная эволюция и развитие . 285 (4): 291–306. Bibcode : 1999jez ... 285..291p . doi : 10.1002/(sici) 1097-010x (19991215) 285: 4 <291 :: Aid-Jez1> 3.0.co; 2-9 . PMID 10578107 . Архивировано из оригинала (PDF) 9 апреля 2011 года . Получено 7 июля 2010 года .
- ^ Ли, Quanguo (9 марта 2012 г.). «Реконструкция микрораптора и эволюция переливающегося оперения». Наука . 335 (6073): 1215–1219. BIBCODE : 2012SCI ... 335.1215L . doi : 10.1126/science.1213780 . PMID 22403389 . S2CID 206537426 .
- ^ Подпрыгнуть до: а беременный в Pettingill, OS Jr. (1970). Орнитология в лаборатории и на местах. Четвертое издание . Burgess Publishing Company. С. 29–58 . ISBN 0-8087-1609-3 .
- ^ «Галапагос плумология» (PDF) . DarwinFoundation.org . База данных Charles Darwin Collections Фонда Чарльза Дарвина. Архивировано с оригинала 17 марта 2016 года . Получено 24 апреля 2015 года .
- ^ Эйххорн, изд. Манфред (2005). Langenscheidt Specialist Dictionary Biology английский: английский - немецкий, немецкий, английский (1 -е изд.). Берлин [UA]: Langenscheidt. п. 537. ISBN 3-86117-228-3 Полем Архивировано из оригинала 17 декабря 2017 года . Получено 24 апреля 2015 года .
- ^ Bonser, RHC; Доусон, С. (1999). «Структурные механические свойства перьев и биомимикирующих натуральных изоляционных материалов». Журнал писем о материалостике . 18 (21): 1769–1770. doi : 10.1023/a: 1006631328233 . S2CID 135061282 .
- ^ Джонстон, Николь и Парсонс, Джин (20 сентября 2018 г.). «Перья: находятся под угрозой исчезновения - фауна и мода» . Университет Миссури исторический костюм и текстиль.
- ^ "Печка (3)" . Онлайн-словарь Merriam-Webster . Merriam-Webster, Inc. Архивирована из оригинала 19 сентября 2011 года . Получено 16 октября 2010 года .
- ^ Schor, R.; Кримм С. (1961). «Исследования структуры перьев кератина: II. Модель β-спирали для структуры перо кератина» . Биофиз. Дж . 1 (6): 489–515. Bibcode : 1961bpj ..... 1..489s . doi : 10.1016/s0006-3495 (61) 86904-x . PMC 1366335 . PMID 19431311 .
- ^ Полинг, Линус; Кори, Роберт Б. (1951). «Структура перьев рахис кератин» . Труды Национальной академии наук Соединенных Штатов Америки . 37 (5): 256–261. Bibcode : 1951pnas ... 37..256p . doi : 10.1073/pnas.37.5.256 . PMC 1063351 . PMID 14834148 .
- ^ HORNIK, C.; Кришан, К.; Yusuf, F.; Scaal, M.; Brand-Saberi, B. (2005). «Misexpression Cdermo-1 вызывает плотную дерму, перья и масштаб» . Биология развития . 277 (1): 42–50. doi : 10.1016/j.ydbio.2004.08.050 . PMID 15572138 .
- ^ Ницш, Кристиан Людвиг (1867). Птерилография Ницша . Рэй Общество. п. 14
- ^ Чандлер 1916 , с. 261
- ^ Чендлер, Аса С. (1916). «Изучение структуры перьев, со ссылкой на их таксономическое значение» . Университет Калифорнийского университета в области зоологии . 13 (11): 243–446 [284].
- ^ McLelland, J. (1991). Цветный атлас птичьего анатомии . WB Saunders Co. ISBN 0-7216-3536-9 .
- ^ Cameron, G.; Wess, T.; Bonser, R. (2003). «Модуль Янга варьируется в зависимости от дифференциальной ориентации кератина в перьях». Журнал структурной биологии . 143 (2): 118–23. doi : 10.1016/s1047-8477 (03) 00142-4 . PMID 12972348 .
- ^ Bonser, R.; Saker, L.; Jeronimidis, G. (2004). «Прочность анизотропия в перьев кератин». Журнал материаловедения . 39 (8): 2895–2896. Bibcode : 2004jmats..39.2895b . doi : 10.1023/b: JMSC.0000021474.75864.ff . S2CID 135873731 .
- ^ Подпрыгнуть до: а беременный Ван, Бин (2016). «Свет как перо: волокнистый натуральный композит с формой, изменяющейся от круглого к квадрату» . Продвинутая наука . 4 (3): 1600360. DOI : 10.1002/Advs.201600360 . PMC 5357985 . PMID 28331789 .
- ^ Lingham-Soliar, Theagarten (2013). «Новая спиральная структура скрещенного волокна B-кератина в перьевых перьях и ее биомеханические последствия» . Plos один . 8 (6): E65849. BIBCODE : 2013PLOSO ... 865849L . doi : 10.1371/journal.pone.0065849 . PMC 3677936 . PMID 23762440 .
- ^ Eaton, Muir D.; Ланьон, Скотт М. (2003). «Вездесущность отражательной способности ультрафиолетового оперения птиц» . Труды: биологические науки . 270 (1525): 1721–1726. doi : 10.1098/rspb.2003.2431 . PMC 1691429 . PMID 12965000 .
- ^ Bostwick, Kimberly S.; Ричард О., Прум (2005). «Ухаживающая за птица поет со стридулирующими перьями крыла» (PDF) . Наука . 309 (5735): 736. doi : 10.1126/science.1111701 . PMID 16051789 . S2CID 22278735 . Архивировано (PDF) из оригинала 7 июля 2010 года . Получено 19 июля 2010 года .
- ^ Подпрыгнуть до: а беременный Делхи, К; Петерс, А.; Kempenaers, B. (2007). «Косметическая окраска у птиц: возникновение, функция и эволюция» (PDF) . Являюсь. НАТ 169 : S145–158. doi : 10.1086/510095 . PMID 19426089 . S2CID 29592388 . Архивировано из оригинала (PDF) 3 декабря 2007 года.
- ^ Рибак, Г.; Weihs, D.; Arad, Z. (2005). «Задержка воды в оперении в дайвингах великих бакланов Phalacrocorax Carbo sinensis ». J. Avian Biol . 36 (2): 89–95. doi : 10.1111/j.0908-8857.2005.03499.x .
- ^ Ледерер, Роджер Дж. (1972). «Роль птичьей риктальной щетины» (PDF) . Бюллетень Уилсона . 84 : 193–97. Архивировано (PDF) из оригинала 4 февраля 2014 года.
- ^ Коновер, мистер; Миллер, Де (1980). «Риктальная функция щетины в Willow Flycatcher» . Кондор . 82 (4): 469–471. doi : 10.2307/1367580 . JSTOR 1367580 . Архивировано с оригинала 22 февраля 2014 года.
- ^ Piersma, t; Van Eerden, MR (1989). «Поедание перья в великолепных гребняках Grebes Podiceps Cristatus : уникальное решение проблем мусора и желудочных паразитов у птиц, питающихся рыбой» (PDF) . Ибис . 131 (4): 477–486. doi : 10.1111/j.1474-919x.1989.tb04784.x .
- ^ Демей, Ида С. (1940). «Изучение птерилоза и пневматичности крикера» . Кондор . 42 (2): 112–118. doi : 10.2307/13644475 . JSTOR 1364475 . Архивировано из оригинала 21 февраля 2014 года.
- ^ Холл, К.; Сюзанна С. (2005). «Имеют ли девять привилегированных проходов девять или десять первичных перьев? Эволюция концепции». Журнал орнитологии . 146 (2): 121–126. doi : 10.1007/s10336-004-0070-5 . S2CID 36055848 .
- ^ Pycraft, WP (1895). «О птерографии Хоацина ( Opistocomus cristatus )» . Ибис . 37 (3): 345–373. doi : 10.1111/j.1474-919x.1895.tb06744.x .
- ^ Тернер, Дж. Скотт (1997). «О тепловой способности птичьего яйца, нагретого пятном вывода» (PDF) . Физиологическая зоология . 70 (4): 470–80. doi : 10.1086/515854 . PMID 9237308 . S2CID 26584982 . Архивировано из оригинала (PDF) 20 октября 2022 года . Получено 29 июля 2020 года - через EBSCO.
- ^ МакГроу, КХ; Nogare, MC (2005). «Распределение уникальных пигментов красных перьев у попугаев» . Биологические письма . 1 (1): 38–43. doi : 10.1098/rsbl.2004.0269 . PMC 1629064 . PMID 17148123 .
- ^ Hausmann, F.; Арнольд, Ке; Маршалл, Нью -Джерси; Оуэнс, IPF (2003). «Ультрафиолетовые сигналы у птиц являются особенными» . Труды Королевского общества б . 270 (1510): 61–67. doi : 10.1098/rspb.2002.2200 . PMC 1691211 . PMID 12590772 .
- ^ Шоуки, Мэтью Д; Хилл, Джеффри Е. (2005). «Каротиноидам нужны структурные цвета, чтобы сиять» (PDF) . Биол. Летал 1 (2): 121–124. doi : 10.1098/rsbl.2004.0289 . PMC 1626226 . PMID 17148144 . Архивировано из оригинала (PDF) 26 марта 2009 года.
- ^ Винтер, Якоб; Бриггс, Дерек, Эг; Кларк, Джулия; Майр, Джеральд; Prum, Richard O. (2009). «Структурная окраска в ископаемом перьев» (PDF) . Биологические письма . 6 (1): 128–31. doi : 10.1098/rsbl.2009.0524 . PMC 2817243 . PMID 19710052 . Архивировано из оригинала (PDF) 21 июня 2010 года . Получено 19 июля 2010 года .
- ^ Dyck, J. (1971). «Структура и спектральная отражательная способность зеленых и голубых перьев любимой птицы ( агапорнис roseicollis )». Биол. Скрипт 18 : 1–67.
- ^ Shawkey, MD; Хилл, GE (2005). «Перья в прекрасном масштабе» . Аук . 121 (3): 652–655. doi : 10.1642/0004-8038 (2004) 121 [0652: FAAFS] 2.0.co; 2 .
- ^ Delhey, K.; Петерс, А.; Бидерманн, PHW; Kempenaers, B. (2008). «Оптические свойства секреции уропигиальной железы: нет доказательств ультрафиолетовой косметики у птиц» . Naturwissenschaften . 95 (10): 939–46. Bibcode : 2008nw ..... 95..939d . doi : 10.1007/s00114-008-0406-8 . HDL : 11858/00-001M-0000-0010-509C-A . PMID 18560743 .
- ^ Shawkey, MD; Pillai, Sr; Хилл, GE (2003). «Химическая война? Влияние уропигиального масла на бактерии, разрушающие перья» (PDF) . Журнал птичьей биологии . 34 (4): 345–349. doi : 10.1111/j.0908-8857.2003.03193.x . Архивировано из оригинала (PDF) 10 сентября 2008 года.
- ^ Эндлер, JA (1980). «Естественный отбор на цветовых узорах в Poeci-Lia reticulata». Эволюция 34 (1): 76–91. doi : 10.2307/2408316 . JSTOR 2408316 . PMID 28563214 .
- ^ Badyaev, Av; Хилл, GE (2000). «Эволюция сексуального дихроматизма: вклад окраски каротиноида и меланина» . Биологический журнал Линневого общества . 69 (2): 153–172. doi : 10.1111/j.1095-8312.2000.tb01196.x . S2CID 201965078 .
- ^ Lozano, GA (1994). «Каротиноиды, паразиты и половой отбор». Оикос . 70 (2): 309–311. Bibcode : 1994oikos..70..309L . doi : 10.2307/3545643 . JSTOR 3545643 . S2CID 86971117 .
- ^ Bonser, RHC (1995). «Меланин и сопротивление истирания перьев» . Кондор . 97 (2): 590–591. doi : 10.2307/1369048 . JSTOR 1369048 . Архивировано из оригинала 23 февраля 2014 года.
- ^ Гранде, JM; Негр, JJ; Торрес, MJ (2004). «Эволюция окрашивания оперения птиц: роль бактерий, разрушающих перьев?» (PDF) . Ардеола . 51 (2): 375–383. Архивировано (PDF) из оригинала 10 сентября 2008 года.
- ^ Бертт, Эдвард Х. младший; Ичида, Янн М. (2004). «Правило Глогера, разрушающие перья бактерии и цветовые вариации среди песенных воробьев» (PDF) . Кондор . 106 (3): 681–686. doi : 10.1650/7383 . S2CID 5857742 . Архивировано (PDF) из оригинала 20 ноября 2012 года.
- ^ Подпрыгнуть до: а беременный Мартин, те; Badyaev, AV (1996). «Сексуальные дихроматические у птиц; важность хищничества гнезда и местоположения гнезда для женщин против мужчин». Эволюция 50 (6): 2454–2460. doi : 10.2307/2410712 . JSTOR 2410712 . PMID 28565684 .
- ^ Касвелл Стоддард, Мэри; Prum, Richard O. (2011). «Насколько красочными птицы? Эволюция цветовой гаммы птичьего оперения» . Поведенческая экология . 22 (5): 1042–1052. doi : 10.1093/beheco/arr088 . HDL : 10.1093/beheco/arr088 .
- ^ Toon, A. & Hughes, J. (2008). «Хорошими доверенными лицами вшами для истории хозяина? Сравнительный анализ австралийской сороки, гимноорхины Тибицен и двух видов в вож в перьев» . Наследственность . 101 (2): 127–135. doi : 10.1038/hdy.2008.37 . PMID 18461081 .
- ^ Брук, М. де Л.; Хироши Накамура (1998). «Приобретение специфических для хозяина вшей с помощью обычных кукушек ( Cuculus canorus )». Журнал зоологии . 244 (2): 167–173. doi : 10.1017/s0952836998002027 .
- ^ Ревис, Ханна С.; Дебора А. Уоллер (2004). «Бактерицидная и фунгицидная активность химических веществ муравьев на паразитах перьев: оценка поведения ансинга как метода самолечения у певчих птиц» . Аук . 121 (4): 1262–1268. doi : 10.1642/0004-8038 (2004) 121 [1262: bafaoa] 2.0.co; 2 . S2CID 85677766 .
- ^ Боуэн, Габриэль Дж; Вассенаар, Леонард I; Хобсон, Кит А (2005). «Глобальное применение стабильных изотопов водорода и кислорода к криминалистике дикой природы». Oecologia . 143 (3): 337–348. Bibcode : 2005oecol.143..337b . doi : 10.1007/s00442-004-1813-y . PMID 15726429 . S2CID 1762342 .
- ^ Jaspers, v.; Voorspoels, S.; Covaci, A.; Lepoint, G. & Eens, M. (2007). «Оценка полезности птичьих перьев в качестве неразрушающего инструмента биомонитора для органических загрязнителей: сравнительный и метааналитический подход» . Environment International . 33 (3): 328–337. Bibcode : 2007enint..33..328j . doi : 10.1016/j.envint.2006.11.011 . HDL : 2268/1067 . PMID 17198730 .
- ^ Poopathi, S.; Абидха С. (2007). «Использование культурных средств на основе перьев для производства комаров». Биологический контроль . 43 (1): 49–55. Bibcode : 2007biolc..43 ... 49p . doi : 10.1016/j.bioControl.2007.04.019 .
- ^ Шмидт, WF; Barone, Jr (2004). «Новое использование для куриных перьев кератинового волокна». Симпозиум по управлению птицеводством . С. 99–101.
- ^ Касарин, Франциани; Бранделли, Флоренсия Кладера-Оливера Адриано; Brandelli, Adriano (2008). «Использование побочного продукта птицы для производства кератинолитических ферментов». Технология питания и биопроцесса . 1 (3): 301–305. doi : 10.1007/s11947-008-0091-9 . S2CID 96154497 .
- ^ Jiang, Z.; Цинь, Д.; HSE, C.; Kuo, M.; Luo, Z.; Ван, Г.; и др. (2008). «Предварительное исследование на клейке на основе белка на основе куриных перьев» . Журнал Wood Chemistry & Technology . 28 (3): 240–246. doi : 10.1080/02773810802347073 . S2CID 3656808 . Архивировано из оригинала 19 февраля 2014 года.
- ^ Нойсюс, Сара В. и Г. Тимоти Гросс 2007 В поисках нашего прошлого: введение в археологию Северной Америки. Издательство Оксфордского университета, Нью -Йорк.
- ^ Froemming, Steve (2006). «Традиционное использование андского мерцания (Colaptes Rupicola) в качестве галактагога в перуанских Андах» . Журнал этнобиологии и этномедицины . 2 : 23. DOI : 10.1186/1746-4269-2-23 . PMC 1484469 . PMID 16677398 .
- ^ Мурари, С.К.; Фрей, FJ; Фрей, Б.М.; Говда, телевизор; Вишванат, Б.С. (2005). «Использование экстракта пера Pavo Cristatus для лучшего лечения змеиных кубиков: нейтрализация воспалительных реакций». Журнал этнофармакологии . 99 (2): 229–237. doi : 10.1016/j.jep.2005.02.027 . PMID 15894132 .
- ^ Матч, мм; Jaroli, DP (2007). Племя Сахаррии в Раджастане, Индия " Журнал этнобиологии и этномедицины . 3 : 25. doi : 10.1186/ 1746-4269-3-2 1892771PMC PMID 1754781 .
- ^ «Шотландское горное платье: клан Кэмпбелл» . Клан Кэмпбелл Общество . Получено 10 ноября 2020 года .
- ^ Doughty, Robin W. Feather Fashions и сохранение птиц, исследование в области защиты природы . Калифорнийский университет. Страница 197.
- ^ Эрлих, Пол Р.; Добкин, Дэвид С.; Сэй, Дэррил (1988). «Торговля в шлейфе» . Стэнфордский университет. Архивировано из оригинала 30 сентября 2008 года.
- ^ Торговля перьями архивировала 23 июня 2008 г. На машине Wayback , Смитсоновском институте
- ^ Боннер, Джесси Л. (6 июня 2011 г.). «Высокая мода или приманка? Укладки мух теперь наращивания волос» . Сиэтл времена . Архивировано из оригинала 10 июня 2011 года.
- ^ Подпрыгнуть до: а беременный Сент -Флер, Николас (8 декабря 2016 года). «Эта вещь с перьями, пойманными в янтарь? Это был хвост динозавра» . New York Times . Архивировано с оригинала 8 декабря 2016 года . Получено 8 декабря 2016 года .
- ^ Sumida, SS; CA Brochu (2000). «Филогенетический контекст для происхождения перьев» . Американский зоолог . 40 (4): 486–503. doi : 10.1093/icb/40.4.486 .
- ^ Dimond, CC; RJ Cabin; JS Brooks (2011). «Перья, динозавры и поведенческие сигналы: определение гипотезы визуального дисплея для адаптивной функции перьев в неавийских тероподах». Биос . 82 (3): 58–63. doi : 10.1893/011.082.0302 . S2CID 98221211 .
- ^ Bock, WJ (2000). «Пояснительная история происхождения перьев» . Являюсь. Zool . 40 (4): 478–485. doi : 10.1093/icb/40.4.478 .
- ^ Уитфилд, Джон (4 апреля 2012 года). «Крупнейший пернатый динозавр, но обнаружился в Китае» . Блог Nature News . Архивировано из оригинала 6 апреля 2012 года . Получено 4 апреля 2012 года .
- ^ Xu x.; Ван К.; Чжан К.; MA Q.; Син Л.; Салливан С.; Hu D.; Cheng S.; Wang S.; и др. (2012). «Гигантский пернатый динозавр из нижнего мела Китая» (PDF) . Природа . 484 (7392): 92–95. Bibcode : 2012natr.484 ... 92x . doi : 10.1038/nature10906 . PMID 22481363 . S2CID 29689629 . Архивировано из оригинала (PDF) 17 апреля 2012 года.
- ^ Люди, Уолтер С.; Керри, Филипп Дж. (2015). «Литлс перед падением: новая перспектива функционального происхождения перьев» . Эволюция 69 (4): 857–862. doi : 10.1111/evo.12634 . ISSN 1558-5646 . PMID 25756292 . S2CID 24319963 .
- ^ De ricqes; AJ; К. Падиан; JR Horner; Et lamm; N. Myhrvolld (2003). "Работа Sanctus. А 23 (2): 373–3 doi : 0272-4634 (2003) 023: OK] 10.1671 / S2CID 84936431 .
- ^ Ли, Quanguo; Gao, Ke-Qin; Винтер, Якоб; Шоуки, Мэтью; Кларк, Джулия; Д'Альба, Лилиана; Мэн, Циндзин; Бриггс, Дерек; Прум, Ричард (12 марта 2010 г.). «Цветовые узоры оперения вымершего динозавра» (PDF) . Наука . 327 (5971): 1369–1372. Bibcode : 2010sci ... 327.1369L . doi : 10.1126/science.1186290 . PMID 20133521 . S2CID 206525132 .
- ^ Cincotta, A., Nicolaï, M., Campos, Hbn et al. Меланосомы птерозавров поддерживают сигнальные функции для ранних перьев. Nature 604, 684–688 (2022). Два : 10.1038/S41586-022-04622-3 .
- ^ Сайно, Никола; Риккардо Стради (1999). «Концентрация каротиноидной в плазме, иммунный профиль и орнамент оперения мужских сарая ласточки». Американский натуралист . 154 (4): 441–448. doi : 10.1086/303246 . PMID 10523490 . S2CID 4400888 .
- ^ Эндлер, Джон А.; Дэвид А. Уэсткотт; Джоа Р. Мэдден; Тим Робсон и Патрик Филлипс (2005). «Визуальные системы животных и эволюция цветовых узоров: сенсорная обработка освещает эволюцию сигнала» . Эволюция 59 (8): 1795–1818. doi : 10.1111/j.0014-3820.2005.tb01827.x . PMID 16329248 . S2CID 25683790 .
- ^ Зеленицкий, DK; Терриен, Ф.; Эриксон, GM; Debuhr, cl; Kobayashi, Y.; Эберт, да; Хэдфилд, Ф. (26 октября 2012 г.). «Пернатые неавийские динозавры из Северной Америки дают представление о происхождении крыла». Наука . 338 (6106): 510–514. Bibcode : 2012sci ... 338..510Z . doi : 10.1126/science.1225376 . ISSN 0036-8075 . PMID 23112330 . S2CID 2057698 .
- ^ Ву, Пинг; Ян, Цзе; Лай, Юнг-Чих; Нг, Чен Сян; Ли, Анг; Цзян, Сюэюан; Else, Ruth M.; Widelitz, Randall; Баджпай, Ручи; Ли, Вэнь-Хсинг; Чуонг, Ченг-Мин (1 февраля 2018 года). «Для преобразования масштабирования в обращение требуются несколько регуляторных модулей» . Молекулярная биология и эволюция . 35 (2): 417–430. doi : 10.1093/molbev/msx295 . ISSN 1537-1719 . PMC 5850302 . PMID 29177513 .
- ^ Alibardi, L.; Knapp, LW; Сойер, RH (июнь 2006 г.). «Локализация бета-кератина в развивающихся масштабах аллигаторов и перьев в отношении развития и эволюции перьев» . Журнал субмикроскопической цитологии и патологии . 38 (2–3): 175–192. ISSN 1122-9497 . PMID 17784647 .
- ^ Алибарди, L; Knapp, LW; Sawyer, RH (2006). «Локализация бета-кератина в развивающихся масштабах аллигаторов и перьев в отношении развития и эволюции перьев» . Журнал субмикроскопической цитологии и патологии . 38 (2–3): 175–92. PMID 17784647 .
- ^ Гринволд, Мэтью Дж.; Сойер, Роджер Х. (15 декабря 2011 г.). «Связывание молекулярной эволюции кератинов птичьей бета (β) с эволюцией перьев» (PDF) . Журнал экспериментальной зоологии Часть B: молекулярная эволюция и развитие . 316b (8): 609–616. Bibcode : 2011jezb..316..609g . doi : 10.1002/jez.b.21436 . PMID 21898788 . Архивировано из оригинала (PDF) 15 октября 2022 года . Получено 15 октября 2022 года .
- ^ Слейтер, Тиффани с; Эдвардс, Николас П; Уэбб, Сэмюэль М; Чжан, Фученг; Макнамара, Мария Э (2023). «Сохранение роговых β-белков в мезозойских перьях» . Природа экология и эволюция . 7 (10): 1706–1713. Bibcode : 2023natee ... 7.1706s . doi : 10.1038/s41559-023-02177-8 . PMID 37735563 . S2CID 262125827 .
- ^ Подпрыгнуть до: а беременный Бентон, Майкл Дж.; Dhouailly, Danielle; Цзян, Баою; Макнамара, Мария (1 сентября 2019 г.). «Раннее происхождение перьев» (PDF) . Тенденции в экологии и эволюции . 34 (9): 856–869. Bibcode : 2019tecoe..34..856b . doi : 10.1016/j.tree.2019.04.018 . HDL : 10468/8068 . ISSN 0169-5347 . PMID 31164250 . S2CID 174811556 .
- ^ Dimond, CC; RJ Cabin; JS Brooks (2011). «Перья, динозавры и поведенческие сигналы: определение гипотезы визуального дисплея для адаптивной функции перьев в неавийских тероподах». Биос . 82 (3): 58–63. doi : 10.1893/011.082.0302 . S2CID 98221211 .
- ^ Сюй, Син; Фученг Чжан (2005). «Новый динозавр манираптора из Китая с длинными перьями на плюсневой кости». Naturwissenschaften . 92 (4): 173–177. Bibcode : 2005nw ..... 92..173X . doi : 10.1007/s00114-004-0604-y . PMID 15685441 . S2CID 789908 .
- ^ Сюй, Син (2006). «Пернатые динозавры из Китая и эволюция главных птичьих персонажей» . Интегративная зоология . 1 (1): 4–11. doi : 10.1111/j.1749-4877.2006.00004.x . PMID 21395983 . S2CID 1516713 .
- ^ Чжэн, XT; HL вам; X. Xu & Zm Dong (2009). «Ранний меловой гетеродонтозавридной динозавр с нитевидными покровными структурами». Природа . 458 (7236): 333–336. Bibcode : 2009natur.458..333Z . doi : 10.1038/nature07856 . PMID 19295609 . S2CID 4423110 .
- ^ Подпрыгнуть до: а беременный в Сюй, х.; Го, Ю. (2009). «Происхождение и ранняя эволюция перьев: понимание недавних палеонтологических и неонтологических данных». Повертеблат Паласиатика . 47 (4): 311–329.
- ^ Godefroit, Pascal; Sinitsa, Sofia M.; Dhouailly, Danielle; Болотский, Юрий Л.; Сизов, Александр V.; Макнамара, Мария Э.; Бентон, Майкл Дж.; Spagna, Paul (2014). «Юрский орнитишский динозавр из Сибири с как перья, так и весы» . Наука . 345 (6195): 451–455. Bibcode : 2014sci ... 345..451G . doi : 10.1126/science.1253351 . HDL : 1983/A7AE6DFB-55BF-4CA4-BD8B-A5EA5F323103 . PMID 25061209 . S2CID 206556907 .
- ^ Барретт, Пол М.; Эванс, Дэвид С.; Кампионе, Николас Э. (30 июня 2015 г.). «Эволюция эпидермальных структур динозавров» . Биологические письма . 11 (6): 20150229. DOI : 10.1098/rsbl.2015.0229 . PMC 4528472 . PMID 26041865 .
- ^ Подпрыгнуть до: а беременный Эти перья не будут придерживаться: максимальное количество моделирования правдоподобия подтверждает масштабы как примитивные для динозаврий (66 -й симпозиум по палеонтологии позвоночных и сравнительной анатомии) , 9 ноября 2018 года, архивировав с оригинала 11 декабря 2021 года , извлеченные 3 октября 2021 года 2021 г.
- ^ Майр, Джеральд; Питман, Майкл; Сатта, Эван; Кэй, Томас Г.; Винтер, Якоб (30 августа 2016 г.). Бенсон, Роджер (ред.). «Структура и гомология хвоста Пситтакозавра» . Палеонтология . 59 (6): 793–802. Bibcode : 2016Palgy..59..793m . doi : 10.1111/pala.12257 . HDL : 1983/029C668F-08B9-45F6-A0C5-30CE9256E593 . S2CID 89156313 .
- ^ Хольц, Томас (19 октября 2018 г.). « Посятый статус: это сложно»: филогенетические, осадочные и биологические препятствия для разрешения наследственного поклевания мезозозой динозаврий » (PDF) . Общество палеонтологии позвоночных . Получено 16 июля 2022 года .
- ^ Ортега, Франциско; Эскасо, Фернандо; Санз, Хосе Л. (9 сентября 2010 г.). «Странная, горбатая карачадонтозаврия (Theropoda) из нижнего мела Испании» . Природа . 467 (7312): 203–206. Bibcode : 2010natur.467..203o . doi : 10.1038/nature09181 . ISSN 1476-4687 . PMID 20829793 . S2CID 4395795 .
- ^ Фот, христианин; Тишлингер, Гельмут; Раухут, Оливер Вм (2 июня 2014 г.). «Новый образец Archeopteryx дает представление о эволюции ножных перьев» . Природа . 511 (7507): 79–82. Bibcode : 2014natur.511 ... 79f . doi : 10.1038/nature13467 . PMID 24990749 . S2CID 4464659 .
- ^ Куэста, Елена; Ортега, Франциско; Санз, Хосе Луис (4 июля 2018 г.). «Аппендикулярная остеология Concavenator Corcovatus (Theropoda: Carcharodontosauridae) из нижнего мела Испании » . Журнал палеонтологии позвоночных . 38 (4): (1) - (24). Doi : 10.1080/02724634.2018.1485153 . ISSN 0272-4634 . S2CID 91976402 .
- ^ Ji, Q.; PJ Currie; Ма Норелл; SA JI (1998). «Два пернатых динозавра из северо -восточного Китая» (PDF) . Природа . 393 (6687): 753–761. Bibcode : 1998natur.393..753q . doi : 10.1038/31635 . S2CID 205001388 .
- ^ Сюй, х.; HH Zhou & Ro Prum (2001). «Разветвленные покровные структуры в Sinornithosaurus и происхождение перьев». Природа . 410 (6825): 200–204. Bibcode : 2001natur.410..200x . doi : 10.1038/35065589 . PMID 11242078 . S2CID 4426803 .
- ^ Ху, Да; LH HOU; LJ Чжан; X. Сюй (2009). «Догареоптерикс Troodontid Theropod из Китая с длинными перьями на плюсанке». Природа . 461 (7264): 640–643. Bibcode : 2009natur.461..640h . doi : 10.1038/nature08322 . PMID 19794491 . S2CID 205218015 .
- ^ Сюй, х.; Q. Чжао; М. Норелл; C. Салливан; Д. Хон; Г. Эриксон; XL Wang; и др. (2009). «Новая пернатая манирапторанская ископаемость динозавров, которая заполняет морфологический разрыв в птичьем происхождении» . Китайский научный бюллетень . 54 (3): 430–435. Bibcode : 2009scibu..54..430x . doi : 10.1007/s11434-009-0009-6 .
- ^ Witmer, LM (2009). "Перьяны динозавры в клутке" . Природа . 461 (7264): 601–602. Bibcode : 2009natur.461..601W . doi : 10.1038/461601a . PMID 197944481 . S2CID 205049989 .
- ^ «Перья динозавра, найденные в Альберте Эмбер» . CBC News . 15 сентября 2011 года. Архивировано с оригинала 15 сентября 2011 года.
- ^ «Редкие птичьи крылья эпохи динозавра, найденные в янтаре» . 28 июня 2016 года. Архивировано с оригинала 28 июня 2016 года . Получено 28 июня 2016 года .
- ^ Син, Лида; МакКеллар, Райан С.; Ван, мин; Бай, Мин; O'Connor, Jingmai K.; Бентон, Майкл Дж.; Чжан, Цзянпинг; Ван, Ян; Ценг, Куоуэй (28 июня 2016 г.). «Мумифицированные предварительные крылья птиц в середине мозговой бирманской янтарь» . Природная связь . 7 : 12089. Bibcode : 2016natco ... 712089x . doi : 10.1038/ncomms12089 . PMC 4931330 . PMID 27352215 .
- ^ Барон, мг; Норман, дБ; Барретт, PM (2017). «Новая гипотеза о отношениях динозавров и ранней эволюции динозавров». Природа . 543 (7646): 501–506. Bibcode : 2017natur.543..501b . doi : 10.1038/nature21700 . PMID 28332513 . S2CID 205254710 .
- ^ Ян, Zixiao; Цзян, Баою; Макнамара, Мария Э.; Кернс, Стюарт Л.; Питман, Майкл; Кэй, Томас Г.; Орр, Патрик Дж.; Сюй, Син; Бентон, Майкл Дж. (17 декабря 2018 г.). «Птерозавр покровные структуры со сложным перьев ветвление» (PDF) . Природа экология и эволюция . 3 (1): 24–30. Bibcode : 2018natee ... 3 ... 24y . doi : 10.1038/s41559-018-0728-7 . HDL : 1983/1F7893A1-924D-4CB3-A4BF-C4B1592356E9 . ISSN 2397-334X . PMID 30568282 . S2CID 56480710 .
- ^ Лангер, Макс С.; Эзкурра, Мартин Д.; Раухут, Оливер Вм; Бентон, Майкл Дж.; Кнолл, Фабен; Макфи, Блэр У.; Novas, Fernando E.; Пол, Диего; Брусатт, Стивен Л. (2 ноября 2017 г.). «Распутывание семейного древа динозавров» (PDF) . Природа . 551 (7678): E1 - E3. Bibcode : 2017natur.551e ... 1L . doi : 10.1038/nature24011 . HDL : 1983/D088DAE2-C7FA-4D41-9FA2-AEEBBFCD2FA3 . ISSN 1476-4687 . PMID 29094688 . S2CID 205260354 .
- ^ Бентон, Майкл Дж.; Dhouailly, Danielle; Цзян, Баою; Макнамара, Мария (1 сентября 2019 г.). «Раннее происхождение перьев» (PDF) . Тенденции в экологии и эволюции . 34 (9): 856–869. Bibcode : 2019tecoe..34..856b . doi : 10.1016/j.tree.2019.04.018 . HDL : 10468/8068 . ISSN 0169-5347 . PMID 31164250 . S2CID 174811556 .
- ^ Подпрыгнуть до: а беременный Foth, C (2011). «О идентификации структур перьев у линии ствола представителей птиц: доказательства окаменелостей и аккупальэонтологии». Paläontologische Zeitschrift . 86 : 91–102. doi : 10.1007/s12542-011-0111-3 . S2CID 86362907 .
- ^ Unwin, David M.; Мартилл, Дэвид М. (декабрь 2020 г.). «Нет протофиков на птерозаврах» . Природа экология и эволюция . 4 (12): 1590–1591. Bibcode : 2020natee ... 4.1590U . doi : 10.1038/s41559-020-01308-9 . ISSN 2397-334X . PMID 32989266 . S2CID 222168569 .
- ^ Ян, Zixiao; Цзян, Баою; Макнамара, Мария Э.; Кернс, Стюарт Л.; Питман, Майкл; Кэй, Томас Г.; Орр, Патрик Дж.; Сюй, Син; Бентон, Майкл Дж. (Январь 2019 г.). «Птерозавр покровные структуры со сложным перьев ветвление» (PDF) . Природа экология и эволюция . 3 (1): 24–30. Bibcode : 2018natee ... 3 ... 24y . doi : 10.1038/s41559-018-0728-7 . HDL : 1983/1F7893A1-924D-4CB3-A4BF-C4B1592356E9 . ISSN 2397-334X . PMID 30568282 . S2CID 56480710 .
- ^ «У птерозавров было четыре типа перьев, новое исследование показывает | палеонтология | Sci-news.com» . Последние научные новости | Sci-news.com . 18 декабря 2018 года . Получено 19 декабря 2018 года .
- ^ Сент -Флер, Николас (17 декабря 2018 года). «Перья и мех летают над ископаемым обнаружением птерозавра - анализ двух окаменелостей оттолкнет происхождение перьев примерно на 70 миллионов лет, но для подтверждения может потребоваться больше образцов» . New York Times . Получено 19 декабря 2018 года .
- ^ Бриггс, Хелен (17 декабря 2018 г.). «Мех летит по новым окаменелостям птерозавра» . BBC News . Получено 19 декабря 2018 года .
- ^ Д'Альба, Лилиана (2019). «Оперение Птерозавра» . Природа экология и эволюция . 3 (1): 12–13. doi : 10.1038/s41559-018-0767-0 . ISSN 2397-334X . PMID 30568284 . S2CID 56480834 .
- ^ Zixiao Yang; Баою Цзян; Мария Э. Макнамара; Стюарт Л. Кернс; Майкл Питман; Томас Г. Кэй; Патрик Дж. Орр; Син Сюй; Майкл Дж. Бентон (2019). «Птерозавр покровные структуры со сложным перьев ветвление» . Природа экология и эволюция . 3 (1): 24–30. Bibcode : 2018natee ... 3 ... 24y . doi : 10.1038/s41559-018-0728-7 . HDL : 1983/1F7893A1-924D-4CB3-A4BF-C4B1592356E9 . PMID 30568282 . S2CID 56480710 .
- ^ Unwin, DM; Martill, DM (2020). «Нет протофиков на птерозаврах». Природа экология и эволюция . 4 (12): 1590–1591. Bibcode : 2020natee ... 4.1590U . doi : 10.1038/s41559-020-01308-9 . PMID 32989266 . S2CID 222168569 .
- ^ Ян, Z.; Цзян, Б.; Макнамара, я; Кернс, SL; Питтман, М.; Кэй, Тг; Orr, PJ; Сюй, х.; Бентон, MJ (2020). «Ответьте: нет протофиков на птерозаврах». Природа экология и эволюция . 4 (12): 1592–1593. Bibcode : 2020natee ... 4.1592y . doi : 10.1038/s41559-020-01309-8 . HDL : 10468/11874 . PMID 32989267 . S2CID 222163211 .
Дальнейшее чтение
- Хансон, Тор (2011). Перья: эволюция естественного чуда . Нью -Йорк: Основные книги. ISBN 978-0-465-02013-3 .
- Лукас, Альфред М.; Стеттенхайм, Питер Р. (1972). «Структура перьев» . Анатомия птиц . Тол. Часть 1. Вашингтон, округ Колумбия: Министерство сельского хозяйства США. С. 235–276.
Внешние ссылки
- McGraw, KJ 2005. Полли хотите пигмент? Тресну химический код в красную окраску у попугаев. Австралийский журнал Birdkeeper 18: 608–611.
- Демео, Антония М. Доступ к орлам и запчастям орла: защита окружающей среды v. Свободное упражнение по религии коренных американцев (1995)
- Электронный кодекс федеральных правил (E-CFR), название 50:
- США против Тридцати восемь Золотых Орлов (1986)
- Механическая структура перьев
- Документальный фильм об эволюции перьев
- Заметки на лекции по общению птиц
- Атлас национальной криминалистики национальной рыбной и дикой природы
- Federn.org