Сокращение мышц

Сокращение мышц – это активация участков, генерирующих напряжение внутри мышечных клеток . [1] [2] В физиологии сокращение мышц не обязательно означает их укорочение, поскольку мышечное напряжение может возникать без изменения длины мышцы, например, при удерживании чего-то тяжелого в одном и том же положении. [1] За прекращением мышечного сокращения следует мышечная релаксация , то есть возвращение мышечных волокон в состояние низкого напряжения. [1]
Чтобы сокращения произошли, мышечные клетки должны полагаться на изменение действия двух типов нитей: тонких и толстых нитей.
Основной составляющей тонких нитей является цепь, образованная спиральным скручиванием двух нитей актина , а толстые нити преимущественно состоят из цепей моторного белка миозина . Вместе эти две нити образуют миофибриллы — основные функциональные органеллы системы скелетных мышц.
У позвоночных , сокращений скелетных мышц являются нейрогенными поскольку они требуют синаптической активности от мотонейронов . Один мотонейрон способен иннервировать несколько мышечных волокон, тем самым заставляя волокна сокращаться одновременно. После иннервации белковые нити внутри каждого волокна скелетной мышцы скользят друг мимо друга, вызывая сокращение, что объясняется теорией скользящих нитей . Произведенное сокращение можно охарактеризовать как подергивание , суммирование или столбняк, в зависимости от частоты потенциалов действия . В скелетных мышцах мышечное напряжение достигает максимального значения, когда мышца растягивается до промежуточной длины, что описывается соотношением длины и напряжения.
В отличие от скелетных мышц, сокращения гладких и сердечных мышц являются миогенными (это означает, что они инициируются самими клетками гладких мышц или сердечных мышц, а не стимулируются внешними событиями, такими как стимуляция нервов), хотя они могут модулироваться стимулами со стороны вегетативная нервная система . Механизмы сокращения этих мышечных тканей аналогичны механизмам сокращения скелетных мышечных тканей.
Сокращение мышц также можно описать с помощью двух переменных: длины и напряжения. [1] В естественных движениях, лежащих в основе двигательной активности , мышечные сокращения многогранны, поскольку они способны вызывать изменения длины и напряжения в зависимости от времени. [3] Следовательно, ни длина, ни напряжение вряд ли останутся неизменными в скелетных мышцах , которые сокращаются во время передвижения. Сокращения можно назвать изометрическими , если напряжение мышц меняется, но длина мышц остается прежней. [1] [4] [5] [6] Напротив, мышечное сокращение описывается как изотоническое , если мышечное напряжение остается одинаковым на протяжении всего сокращения. [1] [4] [5] [6] Если длина мышцы сокращается, сокращение носит концентрический характер; [1] [7] если длина мышцы увеличивается, сокращение носит эксцентрический характер.
Типы
[ редактировать ]
Мышечные сокращения можно описать на основе двух переменных: силы и длины. Саму силу можно дифференцировать как напряжение или нагрузку. Мышечное напряжение — это сила, действующая мышцей на объект, тогда как нагрузка — это сила, действующая объектом на мышцу. [1] Когда мышечное напряжение изменяется без соответствующего изменения длины мышцы, мышечное сокращение описывается как изометрическое. [1] [4] [5] [6] Если длина мышцы изменяется, а напряжение мышц остается прежним, то сокращение мышцы изотоническое. [1] [4] [5] [6] При изотоническом сокращении длина мышцы может либо сокращаться, вызывая концентрическое сокращение, либо удлиняться, вызывая эксцентрическое сокращение. [1] [7] В естественных движениях, лежащих в основе двигательной активности, мышечные сокращения многогранны, поскольку они способны вызывать изменения длины и напряжения в зависимости от времени. [3] Следовательно, ни длина, ни напряжение вряд ли останутся постоянными, когда мышца активна во время двигательной активности.
Изометрическое сокращение
[ редактировать ]Изометрическое сокращение мышцы создает напряжение без изменения длины. [1] [4] [5] [6] Пример можно найти, когда мышцы кисти и предплечья сжимают предмет; суставы руки не двигаются, но мышцы создают достаточную силу , чтобы предотвратить падение предмета.
Изотоническое сокращение
[ редактировать ]При изотоническом сокращении напряжение в мышце остается постоянным, несмотря на изменение длины мышцы. [1] [4] [5] [6] Это происходит, когда сила сокращения мышцы соответствует общей нагрузке на мышцу.
Концентрическое сокращение
[ редактировать ]При концентрическом сокращении мышечного напряжения достаточно для преодоления нагрузки, и по мере сокращения мышца укорачивается. [8] Это происходит, когда сила, создаваемая мышцей, превышает нагрузку, противодействующую ее сокращению.
Во время концентрического сокращения мышца стимулируется к сокращению в соответствии с теорией скользящих нитей . Это происходит по всей длине мышцы, создавая силу в начале и месте прикрепления, вызывая укорочение мышцы и изменение угла сустава. Что касается локтя , концентрическое сокращение бицепса приведет к сгибанию руки в локте, когда рука переместится от ноги к плечу ( сгибание бицепса ). Концентрическое сокращение трицепса меняет угол сустава в противоположную сторону, выпрямляя руку и перемещая кисть к ноге.
Эксцентрическое сокращение
[ редактировать ]При эксцентрическом сокращении напряжения, возникающего во время изометрии, недостаточно для преодоления внешней нагрузки на мышцу, и мышечные волокна удлиняются по мере сокращения. [9] Вместо того, чтобы тянуть сустав в направлении мышечного сокращения, мышца замедляет движение сустава в конце движения или иным образом контролирует изменение положения нагрузки. Это может произойти непроизвольно (например, при попытке поднять вес, слишком тяжелый для мышц) или произвольно (например, когда мышца «сглаживает» движение или сопротивляется силе тяжести, например, при ходьбе под гору). В краткосрочной перспективе силовые тренировки, включающие как эксцентрические, так и концентрические сокращения, по-видимому, увеличивают мышечную силу больше, чем тренировки только с концентрическими сокращениями. [10] Однако повреждение мышц, вызванное физической нагрузкой, также увеличивается во время удлинения сокращений. [11]
Во время эксцентрического сокращения двуглавой мышцы локоть начинает движение в согнутом состоянии, а затем выпрямляется по мере удаления руки от плеча . Во время эксцентрического сокращения трехглавой мышцы локоть начинает движение прямо, а затем сгибается по мере движения руки к плечу. Десмин , тайтин и другие белки z-линии участвуют в эксцентрических сокращениях, но их механизм плохо изучен по сравнению с циклическим переключением мостиков при концентрических сокращениях. [9]
Хотя мышца совершает отрицательную механическую работу (работа совершается над мышцей), химическая энергия (жира или глюкозы или временно запасенная в АТФ ) тем не менее расходуется, хотя и меньше, чем было бы потрачено во время концентрического сокращения. той же силы. Например, человек тратит больше энергии, поднимаясь по лестнице, чем спускаясь по тому же лестнице.
Мышцы, подвергающиеся тяжелой эксцентрической нагрузке, повреждаются при перегрузке (например, во время наращивания мышечной массы или силовых тренировках ) по сравнению с концентрической нагрузкой. Когда эксцентрические сокращения используются в силовых тренировках, их обычно называют негативными . Во время концентрического сокращения сократительные мышечные скользят мимо друга друг миофиламенты миозина и актина , стягивая Z-линии вместе. Во время эксцентрического сокращения миофиламенты скользят друг мимо друга в противоположном направлении, хотя фактическое движение головок миозина во время эксцентрического сокращения неизвестно. Упражнения с тяжелой эксцентрической нагрузкой на самом деле могут выдерживать больший вес (мышцы примерно на 40% сильнее во время эксцентрических сокращений, чем во время концентрических сокращений), а также приводят к большему повреждению мышц и отложенному началу мышечных болей через один-два дня после тренировки. Упражнения, включающие как эксцентрические, так и концентрические сокращения мышц (т. е. включающие сильное сокращение и контролируемое опускание веса), могут привести к большему приросту силы, чем только концентрические сокращения. [10] [12] Хотя непривычные тяжелые эксцентрические сокращения могут легко привести к перетренированности , умеренные тренировки могут обеспечить защиту от травм. [10]
Эксцентрические сокращения при движении.
[ редактировать ]Эксцентрические сокращения обычно возникают как тормозная сила в противовес концентрическому сокращению, защищая суставы от повреждений. Практически во время любого обычного движения эксцентрические сокращения помогают сохранять плавность движений, но также могут замедлять быстрые движения, такие как удары руками или броски. Часть тренировки быстрых движений, таких как подача во время бейсбола, включает в себя уменьшение эксцентрического торможения, что позволяет развивать большую мощность во время движения.
Эксцентрические сокращения исследуются на предмет их способности ускорять восстановление слабых или поврежденных сухожилий. Тендинит ахиллова сухожилия [13] [14] и тендинит надколенника [15] Было доказано, что (также известное как «колено прыгуна» или тендоноз надколенника) эксцентрические сокращения с высокой нагрузкой приносят пользу.
Позвоночные
[ редактировать ]
У позвоночных животных различают три типа мышечной ткани : скелетную, гладкую и сердечную. Скелетные мышцы составляют большую часть мышечной массы тела и отвечают за двигательную активность. Гладкая мускулатура образует кровеносные сосуды , желудочно-кишечный тракт и другие области тела, которые производят устойчивые сокращения. Сердечная мышца образует сердце, которое перекачивает кровь. Скелетные и сердечные мышцы называются поперечно-полосатыми мышцами из-за их полосатого вида под микроскопом, что обусловлено высокоорганизованным чередованием полос А и I полос.
Скелетные мышцы
[ редактировать ]
За исключением рефлексов, все сокращения скелетных мышц происходят в результате сигналов, исходящих из головного мозга. Мозг посылает электрохимические сигналы через нервную систему к двигательному нейрону , который иннервирует несколько мышечных волокон. [16] В случае некоторых рефлексов сигнал о сокращении может исходить из спинного мозга через петлю обратной связи с серым веществом. Другие действия, такие как передвижение, дыхание и жевание, имеют рефлекторный аспект: сокращения могут быть инициированы сознательно или бессознательно.
Нервно-мышечный переход
[ редактировать ]
Нервно -мышечное соединение представляет собой химический синапс, образующийся в результате контакта между мотонейроном и мышечным волокном . [17] Это место, в котором мотонейрон передает сигнал мышечным волокнам, чтобы инициировать мышечное сокращение. Последовательность событий, приводящая к деполяризации мышечного волокна в нервно-мышечном соединении, начинается, когда в теле клетки мотонейрона инициируется потенциал действия, который затем распространяется путем скачкообразной проводимости вдоль его аксона к нервно-мышечному соединению. Как только он достигает терминального бутона , потенциал действия вызывает Ca 2+
приток ионов в терминаль через потенциалзависимые кальциевые каналы . Калифорния 2+
приток вызывает слияние синаптических пузырьков, содержащих нейротрансмиттер ацетилхолин , с плазматической мембраной, высвобождая ацетилхолин в синаптическую щель между окончанием двигательного нейрона и нервно-мышечным соединением волокна скелетных мышц. Ацетилхолин диффундирует через синапс, связывается с никотиновыми ацетилхолиновыми рецепторами нервно-мышечного соединения и активирует их. Активация никотинового рецептора открывает его внутренний натрий / калиевый канал, в результате чего натрий поступает внутрь, а калий вытекает наружу. В результате сарколемма меняет полярность, и ее напряжение быстро подскакивает от потенциала покоя мембраны -90 мВ до +75 мВ при поступлении натрия. Затем мембранный потенциал становится гиперполяризованным, когда калий выходит, а затем снова регулируется до мембранного потенциала покоя. Это быстрое колебание называется потенциалом концевой пластинки. [18] Потенциал-управляемые ионные каналы сарколеммы рядом с концевой пластинкой открываются в ответ на потенциал концевой пластинки. Они специфичны для натрия и калия и пропускают только один из них. Эта волна движения ионов создает потенциал действия, который распространяется от концевой пластины двигателя во всех направлениях. [18] Если потенциалы действия перестают поступать, то ацетилхолин перестает выделяться из терминального бутона. Оставшийся ацетилхолин в синаптической щели либо расщепляется активной ацетилхолинэстеразой , либо реабсорбируется синаптической ручкой, и ничего не остается для замены деградировавшего ацетилхолина.
Связь возбуждения-сокращения
[ редактировать ]Соединение возбуждения-сокращения (ECC) — это процесс, посредством которого мышечный потенциал действия в мышечных волокнах заставляет миофибриллы сокращаться. В скелетных мышцах взаимодействие возбуждения и сокращения основано на прямом взаимодействии между двумя ключевыми белками: каналом высвобождения кальция саркоплазматического ретикулума (SR), идентифицируемым как рианодиновый рецептор 1 (RYR1), и потенциалзависимым кальциевым каналом L-типа, идентифицируемым как дигидропиридиновые рецепторы. , (ДХПР). DHPR расположены на сарколемме (которая включает поверхностную сарколемму и поперечные канальцы ), тогда как RyR находятся через мембрану SR. Тесное расположение поперечных канальцев и двух областей SR, содержащих RyRs, описывается как триада и преимущественно происходит там, где имеет место связь возбуждения-сокращения.

Связь возбуждения-сокращения (ECC) возникает, когда деполяризация скелетных мышц (обычно за счет нервной иннервации) приводит к возникновению потенциала мышечного действия. Этот потенциал действия распространяется по поверхности мышцы и в сеть Т-трубочек мышечного волокна , деполяризуя внутреннюю часть мышечного волокна. Это активирует дигидропиридиновые рецепторы в терминальных цистернах , которые находятся в непосредственной близости от рианодиновых рецепторов в прилежащем саркоплазматическом ретикулуме . Активированные дигидропиридиновые рецепторы физически взаимодействуют с рианодиновыми рецепторами, активируя их посредством отростков стопы (включая конформационные изменения, которые аллостерически активируют рианодиновые рецепторы). Когда рианодиновые рецепторы открываются, Ca 2+ высвобождается из саркоплазматического ретикулума в местное соединительное пространство и диффундирует в основную цитоплазму, вызывая кальциевую искру . [19] Потенциал действия создает почти синхронную активацию тысяч кальциевых искр и вызывает увеличение уровня кальция в масштабах всей клетки, вызывая подъем кальциевого переходного процесса . Калифорния 2+ Высвобождаемый в цитозоль, он связывается с тропонином С актиновыми нитями . Эта связь позволяет актиновым нитям совершать циклическое движение поперек мостиков , создавая силу и, в некоторых ситуациях, движение.
Когда желаемое движение выполнено, можно быстро достичь релаксации многочисленными способами. Релаксация быстро достигается с помощью Ca 2+ буфер с различными цитоплазматическими белками, связывающимися с Ca 2+ с очень высокой близостью. [20] Эти цитоплазматические белки позволяют быстро расслабиться в быстро сокращающихся мышцах. Хотя и медленнее, кальций-АТФаза сарко/эндоплазматического ретикулума (SERCA) активно перекачивает Ca. 2+ обратно в саркоплазматический ретикулум, что приводит к постоянному расслаблению до появления следующего потенциала действия. [19]
Митохондрии также участвуют в Ca 2+ обратный захват, в конечном итоге доставляющий собранный Ca 2+ в SERCA для хранения в саркоплазматическом ретикулуме. Некоторые механизмы релаксации (NCX, Ca2+-насосы и каналы утечки Ca2+) также полностью выводят Ca2+ из клеток. [21] Как Ка 2+ концентрация снижается до уровня покоя, Ca2+ высвобождается из тропонина C, препятствуя перекрестному циклическому циклу, вызывая снижение силы и возникновение релаксации. После полного расслабления мышца снова может сокращаться, тем самым полностью перезапуская цикл.
Теория скользящей нити
[ редактировать ]
Теория скользящих нитей описывает процесс мышц сокращения . Это цикл повторяющихся событий, которые заставляют тонкую нить скользить по толстой нити и создавать напряжение в мышце. [22] Он был независимо разработан Эндрю Хаксли и Рольфом Нидергерке, а также Хью Хаксли и Джин Хэнсон в 1954 году. [23] [24] Физиологически это сокращение неравномерно по всему саркомеру; Центральное положение толстых нитей становится нестабильным и может смещаться во время сокращения, но этому противодействует действие эластичных миофиламентов тайтина . Эта тонкая миофиламент поддерживает равномерное натяжение саркомера, притягивая толстую нить в центральное положение. [25]
Межмостовой цикл
[ редактировать ]
Цикл поперечных мостиков — это последовательность молекулярных событий, лежащая в основе теории скользящих нитей. Поперечный мостик представляет собой выступ миозина, состоящий из двух головок миозина, отходящих от толстых нитей. [1] Каждая головка миозина имеет два сайта связывания: один для аденозинтрифосфата (АТФ), а другой для актина. Связывание АТФ с головкой миозина отделяет миозин от актина , тем самым позволяя миозину связываться с другой молекулой актина. После присоединения АТФ гидролизуется миозином, который использует высвободившуюся энергию для перехода в «взведенное положение», в результате чего он слабо связывается с частью сайта связывания актина. Остальная часть сайта связывания актина блокируется тропомиозином . [26] После гидролизованного АТФ взведенная головка миозина теперь содержит аденозиндифосфат (АДФ) + P i . Два Ка 2+
ионы связываются с тропонином С на актиновых нитях. Тропонин- Ca 2+
Комплекс заставляет тропомиозин скользить и разблокировать оставшуюся часть сайта связывания актина. Разблокирование остальных сайтов связывания актина позволяет двум головкам миозина закрыться и миозин прочно связывается с актином. [26] Затем головка миозина высвобождает неорганический фосфат и инициирует рабочий ход, который создает силу 2 пН. Силовой удар перемещает актиновую нить внутрь, тем самым укорачивая саркомер . Затем миозин высвобождает АДФ, но все еще остается прочно связанным с актином. В конце рабочего хода АДФ высвобождается из головки миозина, оставляя миозин прикрепленным к актину в строгом состоянии до тех пор, пока другой АТФ не свяжется с миозином. Недостаток АТФ может привести к состоянию ригидности, характерному для трупного окоченения . Как только другой АТФ связывается с миозином, головка миозина снова отделяется от актина, и происходит еще один цикл перекрестных мостиков.
Велосипед с перекрестным мостом может продолжаться до тех пор, пока имеется достаточное количество АТФ и кальция. 2+
в цитоплазме. [26] Прекращение перекрестного велосипедного цикла может произойти, когда Ca 2+
обратно активно закачивается в саркоплазматический ретикулум. Когда Ка 2+
на тонкой нити больше нет, тропомиозин меняет конформацию обратно в свое предыдущее состояние, чтобы снова заблокировать сайты связывания. Миозин перестает связываться с тонкой нитью, и мышца расслабляется. Калифорния 2+
ионы покидают молекулу тропонина, чтобы поддерживать Ca 2+
Концентрация ионов в саркоплазме. Активная перекачка Ca 2+
ионы в саркоплазматический ретикулум создают дефицит жидкости вокруг миофибрилл. Это приводит к удалению Ca 2+
ионы тропонина. Таким образом, комплекс тропомиозин-тропонин снова покрывает места связывания на актиновых нитях и сокращение прекращается.
Градация сокращений скелетных мышц
[ редактировать ]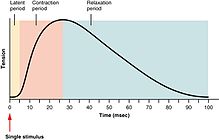

По силе сокращений скелетных мышц можно условно разделить на подергивания , суммации и столбняки . Подергивание — это одиночный цикл сокращения и расслабления, вызываемый потенциалом действия внутри самого мышечного волокна. [27] Время между стимулом двигательного нерва и последующим сокращением иннервируемой мышцы называется латентным периодом , который обычно занимает около 10 мс и обусловлен временем, необходимым для распространения потенциала нервного действия, временем химической передачи в нервно-мышечного соединения, а затем последующие этапы сопряжения возбуждения и сокращения. [28]
Если до полного расслабления мышечного сокращения должен быть создан еще один потенциал действия мышцы, то следующее сокращение просто суммируется с предыдущим сокращением, тем самым производя суммирование . [29] Суммирование может осуществляться двумя способами: [30] суммирование частот и суммирование нескольких волокон . При суммировании частот сила, действующая на скелетные мышцы, контролируется путем изменения частоты, с которой потенциалы действия посылаются к мышечным волокнам. Потенциалы действия не поступают в мышцы синхронно, и во время сокращения некоторая часть волокон мышцы активируется в любой момент времени. В типичных обстоятельствах, когда люди напрягают свои мышцы так сильно, как только могут сознательно, примерно одна треть волокон в каждой из этих мышц активируется одновременно. [ нужна ссылка ] , хотя на это соотношение могут влиять различные физиологические и психологические факторы (включая сухожильные органы Гольджи и клетки Реншоу ). Этот «низкий» уровень сокращения является защитным механизмом, предотвращающим отрыв сухожилия: силы, создаваемой 95%-ным сокращением всех волокон, достаточно, чтобы повредить тело. При суммировании нескольких волокон , если центральная нервная система посылает слабый сигнал о сокращении мышцы, меньшие двигательные единицы в первую очередь стимулируются , поскольку они более возбудимы, чем более крупные. По мере увеличения силы сигнала помимо более крупных двигательных единиц возбуждаются и другие двигательные единицы, причем сократительная сила самых крупных двигательных единиц в 50 раз превышает силу меньших. По мере того, как активируются все более крупные двигательные единицы, сила мышечного сокращения становится все сильнее. Концепция, известная как принцип размера, позволяет постепенно изменять мышечную силу во время слабого сокращения небольшими шагами, которые затем постепенно увеличиваются, когда требуется большее усилие.
Наконец, если частота мышечных потенциалов действия увеличивается настолько, что мышечное сокращение достигает максимальной силы и выходит на плато на этом уровне, то это сокращение является столбнячным .
Соотношение длины и напряжения
[ редактировать ]
Соотношение длины и напряжения связывает силу изометрического сокращения с длиной мышцы, в которой происходит сокращение. Мышцы работают с наибольшим активным напряжением, когда их длина близка к идеальной (часто к длине покоя). При растяжении или сокращении сверх этого значения (будь то из-за действия самой мышцы или внешней силы) максимальное создаваемое активное напряжение уменьшается. [31] Это уменьшение минимально при небольших отклонениях, но напряжение быстро падает по мере дальнейшего отклонения длины от идеальной. Благодаря наличию эластичных белков внутри мышечной клетки (таких как тайтин ) и внеклеточного матрикса, когда мышца растягивается за пределы заданной длины, возникает полностью пассивное напряжение, препятствующее удлинению. В совокупности существует сильное сопротивление удлинению активной мышцы далеко за пределами пика активного напряжения.
Отношения сила-скорость
[ редактировать ]
Соотношение сила-скорость связывает скорость, с которой мышца меняет свою длину (обычно регулируемую внешними силами, такими как нагрузка или другие мышцы), с величиной силы, которую она генерирует. Сила уменьшается гиперболическим образом относительно изометрической силы по мере увеличения скорости сокращения, в конечном итоге достигая нуля при некоторой максимальной скорости. Обратное справедливо для случаев, когда мышца растянута: сила увеличивается выше изометрического максимума, пока, наконец, не достигнет абсолютного максимума. Это внутреннее свойство активной мышечной ткани играет роль в активном демпфировании суставов, которые приводятся в действие одновременно активными противоположными мышцами. В таких случаях профиль сила-скорость увеличивает силу, создаваемую удлиняющейся мышцей за счет укорачивающейся мышцы. Такое предпочтение той мышцы, которая возвращает сустав в равновесие, эффективно увеличивает демпфирование сустава. Причем сила демпфирования увеличивается с ростом мышечной силы. Таким образом, двигательная система может активно контролировать демпфирование суставов посредством одновременного сокращения (совместного сокращения) противоположных групп мышц. [32]
Гладкая мускулатура
[ редактировать ]
Гладкие мышцы можно разделить на две подгруппы: одноединичные и многоединичные . Одноединичные гладкомышечные клетки можно обнаружить в кишечнике и кровеносных сосудах. Поскольку эти клетки связаны между собой щелевыми соединениями, они способны сокращаться как функциональный синцитий . Одноединичные гладкомышечные клетки сокращаются миогенно, что может модулироваться вегетативной нервной системой.
В отличие от одноединичных гладкомышечных клеток, многоединичные гладкомышечные клетки обнаруживаются в мышцах глаз и в основании волосяных фолликулов. Многоединичные гладкомышечные клетки сокращаются при отдельной стимуляции нервами вегетативной нервной системы. По существу, они обеспечивают точный контроль и постепенные реакции, подобно рекрутированию двигательных единиц в скелетных мышцах.
Механизмы сокращения гладких мышц.
[ редактировать ]
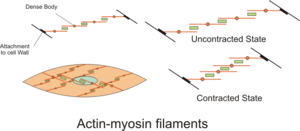
Сократительная активность гладкомышечных клеток может быть тонической (устойчивой) или фазической (преходящей). [33] и на него влияют многочисленные факторы, такие как спонтанная электрическая активность, нервные и гормональные воздействия, локальные изменения химического состава и растяжение. [1] Это контрастирует с сократительной активностью клеток скелетных мышц, которая зависит от единственного нервного импульса. Некоторые типы гладкомышечных клеток способны спонтанно генерировать собственные потенциалы действия, которые обычно возникают вслед за потенциалом водителя ритма или потенциалом медленной волны . Эти потенциалы действия генерируются притоком внеклеточного кальция. 2+
, а не На +
. Как и в скелетных мышцах, цитозольный Ca 2+
ионы также необходимы для перекрестного цикла в гладкомышечных клетках.
Два источника цитозольного Ca 2+
в гладкомышечных клетках находится внеклеточный Са 2+
поступление через кальциевые каналы и Ca 2+
ионы, высвобождаемые из саркоплазматической сети. Повышение цитозольного Ca 2+
приводит к увеличению количества Ca 2+
связывание с кальмодулином , который затем связывается и активирует киназу легкой цепи миозина . Комплекс кальций-кальмодулин-киназа легкой цепи миозина фосфорилирует миозин на легких цепях миозина массой 20 килодальтон (кДа) по аминокислотному остатку серин 19, обеспечивая молекулярное взаимодействие миозина и актина, инициируя сокращение и активируя миозиновую АТФазу . В отличие от клеток скелетных мышц, в клетках гладких мышц отсутствует тропонин, хотя они содержат белок тонких нитей тропомиозин и другие известные белки — кальдесмон и кальпонин. Таким образом, сокращения гладких мышц инициируются Са 2+
-активированное фосфорилирование миозина, а не Са 2+
связывание с тропониновым комплексом, который регулирует места связывания миозина на актине, как в скелетных и сердечных мышцах.
Прекращение цикла перекрестных мостиков (и оставление мышцы в состоянии фиксации) происходит, когда фосфатаза легкой цепи миозина удаляет фосфатные группы из головок миозина. Фосфорилирование легких цепей миозина массой 20 кДа хорошо коррелирует со скоростью сокращения гладких мышц. В этот период наблюдается быстрый всплеск потребления энергии, измеряемый потреблением кислорода. В течение нескольких минут после начала заметно снижается уровень кальция, уменьшается фосфорилирование легких цепей миозина массой 20 кДа и снижается потребление энергии; однако сила тонических гладких мышц сохраняется. Во время сокращения мышц между активированным актином и фосфорилированным миозином образуются быстро циклические перекрестные мостики, генерирующие силу. Предполагается, что поддержание силы происходит за счет дефосфорилированных «мостиков-защелок», которые медленно циклически поддерживают силу. Считается, что ряд киназ, таких как ро-киназа , DAPK3 и протеинкиназа C, участвуют в устойчивой фазе сокращения, а Ca 2+
поток может быть значительным.
Нейромодуляция
[ редактировать ]Хотя сокращения гладких мышц являются миогенными, скорость и сила их сокращений могут модулироваться вегетативной нервной системой . Постганглионарные нервные волокна парасимпатической нервной системы выделяют нейромедиатор ацетилхолин, который связывается с мускариновыми рецепторами ацетилхолина (мАХР) на гладкомышечных клетках. Эти рецепторы являются метаботропными или рецепторами, связанными с G-белком, которые инициируют каскад вторичных мессенджеров. И наоборот, постганглионарные нервные волокна симпатической нервной системы выделяют нейротрансмиттеры адреналин и норадреналин, которые связываются с адренергическими рецепторами, которые также являются метаботропными. Точные эффекты на гладкие мышцы зависят от конкретных характеристик активированного рецептора: как парасимпатическая, так и симпатическая активность может быть либо возбуждающей (сократительной), либо тормозящей (расслабляющей).
Сердечная мышца
[ редактировать ]
Существует два типа клеток сердечной мышцы : авторитмические и сократительные. Ауторитмические клетки не сокращаются, а задают темп сокращения других клеток сердечной мышцы, который может модулироваться вегетативной нервной системой. Напротив, сократительные мышечные клетки (кардиомиоциты) составляют большую часть сердечной мышцы и способны сокращаться.
Связь возбуждения-сокращения
[ редактировать ]При соединении возбуждения-сокращения (ЭК) скелетных и сердечных мышц деполяризационная проводимость и Ca 2+ происходят процессы освобождения. Однако, хотя задействованные белки схожи, они различаются по структуре и регуляции. Дигидропиридиновые рецепторы (DHPR) кодируются разными генами, а рианодиновые рецепторы (RyR) представляют собой отдельные изоформы. Кроме того, DHPR связывается с RyR1 (основная изоформа RyR в скелетных мышцах) для регуляции кальция. 2+ высвобождается в скелетных мышцах, в то время как кальциевые каналы L-типа (DHPR на сердечных миоцитах) и RyR2 (основная изоформа RyR в сердечной мышце) физически не связаны в сердечной мышце, а сталкиваются друг с другом посредством соединительной связи. [34]
В отличие от скелетных мышц, взаимодействие ЭК в сердечной мышце, как полагают, зависит в первую очередь от механизма, называемого кальций-индуцированным высвобождением кальция . [35] который основан на структуре соединения между Т-канальцами и саркоплазматической сетью. Юнктофилин-2 (JPH2) необходим для поддержания этой структуры, а также целостности Т-трубочек . [36] [37] [38] Другой белок, рецепторный дополнительный белок 5 (REEP5), поддерживает нормальную морфологию соединительного SR. [39] Дефекты соединительного сцепления могут быть результатом дефицита любого из двух белков. В процессе высвобождения кальция, индуцированного кальцием, RyR2 активируются кальциевым триггером, который вызывается потоком кальция. 2+ через кальциевые каналы L-типа. После этого сердечная мышца имеет тенденцию проявлять диадную структуру, а не триаду .
Соединение возбуждения-сокращения в клетках сердечной мышцы происходит, когда потенциал действия инициируется пейсмекерными клетками в синоатриальном или атриовентрикулярном узле и передается ко всем клеткам сердца через щелевые контакты . Потенциал действия перемещается вдоль поверхностной мембраны в Т-канальцы (последние наблюдаются не во всех типах сердечных клеток), и деполяризация вызывает внеклеточный Ca. 2+
проникать в клетку через кальциевые каналы L-типа и, возможно, натрий-кальциевый обменник (NCX) на ранней стадии фазы плато . Хотя это Ка 2+ приток учитывается только около 10% Ca 2+ необходимый для активации, он относительно больше, чем у скелетных мышц. Это Калифорния 2+
приток вызывает небольшое локальное увеличение внутриклеточного кальция 2+
. Увеличение внутриклеточного Са 2+
обнаруживается RyR2 в мембране саркоплазматического ретикулума, который высвобождает Ca 2+
в виде физиологической реакции с положительной обратной связью . Эта положительная обратная связь известна как индуцированное кальцием высвобождение кальция. [35] и вызывает образование кальциевых искр ( Ca 2+
искры [40] ). Пространственное и временное суммирование ~30 000 Ca. 2+
искры дают общеклеточное увеличение концентрации цитоплазматического кальция. [41] Увеличение цитозольного кальция после прохождения кальция через клеточную мембрану и саркоплазматический ретикулум сдерживается кальциевыми буферами , которые связывают большую часть внутриклеточного кальция. В результате большое увеличение общего кальция приводит к относительно небольшому увеличению свободного кальция. 2+
. [42]
Цитоплазматический кальций связывается с тропонином С, перемещая тропомиозиновый комплекс с сайта связывания актина, позволяя головке миозина связываться с актиновой нитью. С этого момента сократительный механизм по существу такой же, как и для скелетных мышц (см. выше). Коротко говоря, с помощью гидролиза АТФ головка миозина тянет актиновую нить к центру саркомера.

После систолы внутриклеточный кальций поглощается насосом АТФазы сарко/эндоплазматического ретикулума (SERCA) обратно в саркоплазматический ретикулум, готовый к началу следующего цикла. Кальций также выводится из клетки главным образом посредством натрий-кальциевого обменника (NCX) и, в меньшей степени, кальциевой АТФазы плазматической мембраны . Некоторое количество кальция также поглощается митохондриями. [43] Фермент фосфоламбан служит тормозом для SERCA. При низкой частоте сердечных сокращений фосфоламбан активен и замедляет активность АТФазы, так что Ca 2+
не обязательно покидать клетку полностью. При высокой частоте сердечных сокращений фосфоламбан фосфорилируется и деактивируется, забирая большую часть кальция. 2+
из цитоплазмы обратно в саркоплазматический ретикулум. И снова кальциевые буферы смягчают это падение содержания кальция. 2+
концентрации, позволяющей относительно небольшое уменьшение свободного Ca 2+
концентрации в ответ на большое изменение общего содержания кальция. Падающая Са 2+
концентрация позволяет тропониновому комплексу отделиться от актиновой нити, тем самым прекращая сокращение. Сердце расслабляется, позволяя желудочкам наполниться кровью и снова начать сердечный цикл.
Беспозвоночное
[ редактировать ]Круговые и продольные мышцы
[ редактировать ]
У кольчатых червей, таких как дождевые черви и пиявки , круговые и продольные мышечные клетки образуют стенку тела этих животных и отвечают за их движение. [44] Например, у дождевого червя, движущегося по почве, сокращения круговых и продольных мышц происходят взаимно, в то время как целомическая жидкость служит гидроскелетом , поддерживая набухание дождевого червя. [45] Когда круговые мышцы передних сегментов сокращаются, передняя часть тела животного начинает сжиматься в радиальном направлении, что выталкивает несжимаемую целомическую жидкость вперед и увеличивает длину животного. В результате передняя часть животного выдвигается вперед. Когда передний конец дождевого червя закрепляется, а круговые мышцы в передних сегментах расслабляются, волна продольных мышечных сокращений проходит назад, что тянет остальную часть тела животного вперед. [44] [45] Эти чередующиеся волны круговых и продольных сокращений называются перистальтикой и лежат в основе ползущего движения дождевых червей.
Косополосатые мышцы
[ редактировать ]Беспозвоночные, такие как кольчатые черви, моллюски и нематоды , обладают косо-полосатыми мышцами, которые содержат полосы толстых и тонких нитей, расположенных спирально, а не поперечно, как в скелетных или сердечных мышцах позвоночных. [46] У двустворчатых моллюсков косополосатые мышцы могут сохранять напряжение в течение длительного времени, не затрачивая слишком много энергии. Двустворчатые моллюски используют эти мышцы, чтобы держать раковину закрытой.
Асинхронные мышцы
[ редактировать ]
Развитые насекомые, такие как осы , мухи , пчелы и жуки , обладают асинхронными мышцами , которые составляют мышцы полета у этих животных. [46] Эти летательные мышцы часто называют фибриллярными мышцами, поскольку они содержат толстые и заметные миофибриллы. [47] Замечательной особенностью этих мышц является то, что они не требуют стимуляции для каждого мышечного сокращения. Поэтому их называют асинхронными мышцами , поскольку количество сокращений в этих мышцах не соответствует (или не синхронизируется) количеству потенциалов действия. Например, мышца крыла привязанной мухи может получать потенциалы действия с частотой 3 Гц, но способна сокращаться с частотой 120 Гц. [46] Высокочастотное биение стало возможным благодаря тому, что мышцы подключены к резонансной системе, которая приводит в движение собственную частоту вибрации.
История
[ редактировать ]
В 1780 году Луиджи Гальвани обнаружил, что мышцы ног мертвых лягушек подергиваются при ударе электрической искрой. [49] Это был один из первых набегов на изучение биоэлектричества — области, которая до сих пор изучает электрические закономерности и сигналы в тканях, таких как нервы и мышцы.
В 1952 году был придуман термин «связь возбуждения и сокращения» для описания физиологического процесса преобразования электрического стимула в механическую реакцию. [50] Этот процесс является фундаментальным для физиологии мышц: электрический стимул обычно представляет собой потенциал действия, а механическая реакция — сокращение. При многих заболеваниях связь возбуждения и сокращения может быть нарушена. Хотя взаимодействие возбуждения и сокращения известно уже более полувека, оно до сих пор является активной областью биомедицинских исследований. Общая схема такова, что поступает потенциал действия, деполяризующий клеточную мембрану. По механизмам, специфичным для типа мышц, эта деполяризация приводит к увеличению цитозольного кальция , что называется транзиторным кальцием. Это увеличение содержания кальция активирует чувствительные к кальцию сократительные белки, которые затем используют АТФ, чтобы вызвать укорочение клеток.
Механизм мышечного сокращения годами ускользал от учёных и требует продолжения исследований и обновлений. [51] Теория скользящей нити была независимо разработана Эндрю Ф. Хаксли и Рольфом Нидергерке, а также Хью Хаксли и Джин Хэнсон . Их результаты были опубликованы в виде двух последовательных статей, опубликованных в номере журнала Nature от 22 мая 1954 года под общей темой «Структурные изменения в мышцах во время сокращения». [23] [24]
См. также
[ редактировать ]- Анатомические условия движения
- индуцированное кальцием высвобождение кальция
- Сердечный потенциал действия
- Судороги
- Дистония
- Физиология упражнений
- Фасцикуляция
- Мышечная модель Хилла
- Гипнический придурок
- Тестирование мышц in vitro
- Парадокс Ломбардса
- Миоклонус
- Ригор мертв
- Спазм
- Сокращение матки
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот Видмайер, Эрик П.; Рафф, Херсель; Стрэнг, Кевин Т. (2010). «Мускул». Физиология человека Вандера: механизмы функционирования тела (12-е изд.). Нью-Йорк, штат Нью-Йорк: МакГроу-Хилл. стр. 250–291. ISBN 978-0-321-98122-6 .
- ^ Сильверторн, Ди Унглауб (2016). «Мышцы». Физиология человека: комплексный подход (7-е изд.). Сан-Франциско, Калифорния: Пирсон. стр. 377–416. ISBN 978-0-321-98122-6 .
- ^ Jump up to: а б Бивенер, Эндрю А. (2003). «Мышцы и скелеты: строительные блоки движения животных». Передвижение животных . Оксфордская серия по биологии животных. Нью-Йорк, штат Нью-Йорк: Издательство Оксфордского университета. стр. 15–45. ISBN 978-0-198-50022-3 .
- ^ Jump up to: а б с д и ж Эйдли, Дэвид Дж. (1998). «Механика и энергетика мышечного сокращения». Физиология возбудимых клеток (4-е изд.). Нью-Йорк, штат Нью-Йорк: Издательство Кембриджского университета. стр. 323–335 . ISBN 978-0-521-57421-1 .
- ^ Jump up to: а б с д и ж Сиркар, Сабьясачи (2008). «Упругость мышц». Принципы медицинской физиологии (1-е изд.). Нью-Йорк, штат Нью-Йорк: Тиме. п. 113. ИСБН 978-1-588-90572-7 .
- ^ Jump up to: а б с д и ж Буллок, Джон; Бойл, Джозеф; Ван, Майкл Б. (2001). «Сокращение мышц». Физиология НМС . Том. 578 (4-е изд.). Балтимор, Мэриленд: Липпинкотт Уильямс и Уилкинс. стр. 37–56.
- ^ Jump up to: а б Кумар, Шраван (2008). «Введение и терминология». В Кумаре, Шраван (ред.). Мышечная сила (1-е изд.). Бока-Ратон, Флорида: CRC Press. п. 113. ИСБН 978-0-415-36953-4 .
- ^ Фолкнер, Дж. А. (2003). «Терминология сокращений мышц при укорочении, при изометрии и при удлинении». Журнал прикладной физиологии . 95 (2): 455–459. doi : 10.1152/japplphysicalol.00280.2003 . ПМИД 12851415 . S2CID 28649208 .
- ^ Jump up to: а б «Виды схваток» . 31 мая 2006 года . Проверено 2 октября 2007 г.
- ^ Jump up to: а б с Коллиандер Э.Б., Теш П.А. (1990). «Эффекты эксцентрических и концентрических действий мышц при тренировках с отягощениями». Акта Физиол. Скан . 140 (1): 31–9. дои : 10.1111/j.1748-1716.1990.tb08973.x . ПМИД 2275403 .
- ^ Николаидис М.Г., Кипарос А., Спану С., Пасхалис В., Теодору А.А., Врабас И.С. (2012). «Окислительно-восстановительная биология упражнений: интегративное и сравнительное рассмотрение некоторых упущенных из виду проблем» . Дж. Эксп. Биол . 215 (Часть 10): 1615–25. дои : 10.1242/jeb.067470 . ПМИД 22539728 .
- ^ Брукс, Джорджия; Фэйи, ТД; Уайт, Т.П. (1996). Физиология упражнений: биоэнергетика человека и ее приложения . Мэйфилд Паблишинг Ко.
- ^ Альфредсон, Х; Пиетиля, Т; Йонссон, П; Лоренцон, Р. (1998). «Эксцентрическая тренировка икроножных мышц с тяжелой нагрузкой для лечения хронического тендиноза ахиллова сухожилия» (PDF) . Американский журнал спортивной медицины . 26 (3): 360–6. дои : 10.1177/03635465980260030301 . ПМИД 9617396 . S2CID 30259362 .
- ^ Сатьендра Л., Бил Н. (2006). «Эффективность физиотерапии при тендинопатии ахиллова сухожилия: научно обоснованный обзор эксцентрических упражнений» . Изокинетика и физкультура . 14 (1): 71–80. дои : 10.3233/IES-2006-0223 . Архивировано из оригинала 9 июля 2012 года.
- ^ Каннелл Л.Дж., Тонтон Дж.Э., Клемент Д.Б., Смит С., Хан К.М. (2001). «Рандомизированное клиническое исследование эффективности приседаний или упражнений на разгибание/сгибание ног для лечения клинически диагностированного колена прыгуна у спортсменов: пилотное исследование» . Br J Sports Med . 35 (1): 60–4. дои : 10.1136/bjsm.35.1.60 . ПМЦ 1724276 . ПМИД 11157465 .
- ^ Тассинарий; Качиоппо (2000). «Скелетомоторная система: поверхностная электромиография». В Качиоппо, Джон Т.; Тассинари, Луис Г.; Бернтсон, Гэри Г. (ред.). Справочник по психофизиологии (второе изд.). Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-62634-7 .
- ^ Левитан, Ирвин; Качмарек, Леонард (19 августа 2015 г.). «Межклеточная связь». Нейрон: клеточная и молекулярная биология (4-е изд.). Нью-Йорк, штат Нью-Йорк: Издательство Оксфордского университета. стр. 153–328. ISBN 978-0199773893 .
- ^ Jump up to: а б Саладин, Кеннет С., Стивен Дж. Салливан и Кристина А. Ган. Анатомия и физиология: единство формы и функции. 7-е изд. Нью-Йорк: McGraw-Hill Education, 2015. Печать.
- ^ Jump up to: а б Ланнер, Джоанна Т.; Георгиу, Димитра К.; Джоши, Адитья Д.; Гамильтон, Сьюзен Л. (ноябрь 2010 г.). «Рианодиновые рецепторы: структура, экспрессия, молекулярные детали и функция высвобождения кальция» . Перспективы Колд-Спринг-Харбор в биологии . 2 (11): а003996. doi : 10.1101/cshperspect.a003996 . ISSN 1943-0264 . ПМЦ 2964179 . ПМИД 20961976 .
- ^ Кальдерон, Хуан К.; Боланьос, Пура; Капуто, Карло (март 2014 г.). «Механизм связи возбуждения-сокращения в скелетных мышцах» . Биофизические обзоры . 6 (1): 133–160. дои : 10.1007/s12551-013-0135-x . ISSN 1867-2450 . ПМЦ 5425715 . ПМИД 28509964 .
- ^ Уэбб, Р. Клинтон (декабрь 2003 г.). «СОКРАЩЕНИЕ И РАССЛАБЛЕНИЕ ГЛАДКИХ МЫШЦ» . Достижения в области физиологического образования . 27 (4): 201–206. дои : 10.1152/advan.00025.2003 . ISSN 1043-4046 .
- ^ Саладин, Кеннет (2012). Анатомия и физиология: единство формы и функции . Нью-Йорк: МакГроу Хилл. ISBN 978-0-07-337825-1 .
- ^ Jump up to: а б Хаксли А.Ф., Нидергерке Р. (1954). «Структурные изменения в мышцах во время сокращения: интерференционная микроскопия живых мышечных волокон». Природа . 173 (4412): 971–973. Бибкод : 1954Natur.173..971H . дои : 10.1038/173971a0 . ПМИД 13165697 . S2CID 4275495 .
- ^ Jump up to: а б Хаксли Х, Хэнсон Дж (1954). «Изменения поперечной исчерченности мышц при сокращении и растяжении и их структурная интерпретация». Природа . 173 (4412): 973–976. Бибкод : 1954Natur.173..973H . дои : 10.1038/173973a0 . ПМИД 13165698 . S2CID 4180166 .
- ^ Горовиц Р., Подольский Р.Ю. (ноябрь 1987 г.). «Позиционная стабильность толстых нитей в активированных скелетных мышцах зависит от длины саркомера: доказательства роли тайтиновых нитей» . Дж. Клеточная Биол . 105 (5): 2217–23. дои : 10.1083/jcb.105.5.2217 . ПМК 2114850 . ПМИД 3680378 .
- ^ Jump up to: а б с Энока, Роджер М.; Пирсон, Кейр Г. (2013). «Двигатель и действие мышц». В Канделе, Эрик Р.; Шварц, Джеймс Х.; Джесселл, Томас М.; Сигельбаум, Стивен А.; Хадспет, Эй.Дж. (ред.). Принципы нейронауки (5-е изд.). Нью-Йорк, штат Нью-Йорк: McGraw-Hill Medical. стр. 768–789. ISBN 978-0-071-39011-8 .
- ^ Фехер, Джозеф (2012). «Глава 3.4: Механика скелетных мышц». Количественная физиология человека: Введение . Серия Academic Press по биомедицинской инженерии (1-е изд.). Нью-Йорк, штат Нью-Йорк: Academic Press. стр. 239–248. ISBN 978-0-123-82163-8 .
- ^ Хурана, Инду (2006). «Особенности возбудимости и сократимости мышц». Учебник медицинской физиологии (1-е изд.). Эльзевир. стр. 101–2.
- ^ Смит, Ян К.; Адам, Хелен; Херцог, Вальтер (апрель 2020 г.). «Краткое сокращение оказывает сложное влияние на суммирование пар сокращений в приводящей большой палец мышцы человека» . Экспериментальная физиология . 105 (4): 676–689. дои : 10.1113/ep088401 . ПМИД 32052487 . S2CID 211100581 . Проверено 5 апреля 2022 г.
- ^ Шведик, Э.; Баласубраманян, Р.; Скотт, Р.Н. (1977). «Нестационарная модель электромиограммы». Транзакции IEEE по биомедицинской инженерии . 24 (5): 417–424. дои : 10.1109/TBME.1977.326175 . ПМИД 892834 . S2CID 1770255 .
- ^ Гордон А.М., Хаксли А.Ф., Джулиан Ф.Дж. (1966). «Изменение изометрического напряжения в зависимости от длины саркомера в мышечных волокнах позвоночных» . Дж. Физиол . 184 (1): 170–92. дои : 10.1113/jphysicalol.1966.sp007909 . ПМЦ 1357553 . ПМИД 5921536 .
- ^ Хайтманн, Стюарт; Папоротники, Норм; Брейпсир, Майкл (2011). «Совместное сокращение мышц модулирует демпфирование и стабильность суставов в трехзвенной биомеханической конечности» . Границы нейроробототехники . 5 : 5. дои : 10.3389/fnbot.2011.00005 . ISSN 1662-5218 . ПМЦ 3257849 . ПМИД 22275897 .
- ^ Чжан, Ю; Хермансон, Мэн; Эдингер, Ти Джей (2013). «Тоническое и фазовое сокращение гладких мышц не регулируется путем PKCα-CPI-17 в антральном отделе и на дне желудка свиньи» . ПЛОС ОДИН . 8 (9): е74608. Бибкод : 2013PLoSO...874608Z . дои : 10.1371/journal.pone.0074608 . ПМЦ 3776813 . ПМИД 24058600 .
- ^ Мартоноси, Энтони Н.; Пикула, Славомир (2003). «Сеть регуляции кальция в мышцах» . Акта Биохимика Полоника . 50 (1): 1–30. дои : 10.18388/abp.2003_3711 . ISSN 0001-527X . ПМИД 12673344 .
- ^ Jump up to: а б Фабиато, А. (1983). «Высвобождение кальция из сердечного саркоплазматического ретикулума, индуцированное кальцием». Американский журнал физиологии . 245 (1): С1–14. дои : 10.1152/ajpcell.1983.245.1.C1 . ПМИД 6346892 .
- ^ Го, Анг; Чжан, Сяоин; Айер, Венкат Рамеш; Чен, Бийи; Чжан, Каймэй; Кучке, Уильям Дж.; Вайс, Роберт М.; Франзини-Армстронг, Клара; Сун, Лун-Шэн (19 августа 2014 г.). «Сверхэкспрессия юнктофилина-2 не улучшает исходную функцию, но ослабляет развитие сердечной недостаточности после сердечного стресса» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (33): 12240–12245. Бибкод : 2014PNAS..11112240G . дои : 10.1073/pnas.1412729111 . ISSN 1091-6490 . ПМК 4143026 . ПМИД 25092313 .
- ^ Вэй, Шэн; Го, Анг; Чен, Бийи; Кучке, Уильям; Се, Ю-Пин; Циммерман, Кэти; Вайс, Роберт М.; Андерсон, Марк Э.; Ченг, Хэпин; Сун, Лун-Шэн (20 августа 2010 г.). «Ремоделирование Т-трубочек при переходе от гипертрофии к сердечной недостаточности» . Исследование кровообращения . 107 (4): 520–531. дои : 10.1161/CIRCRESAHA.109.212324 . ISSN 1524-4571 . ПМЦ 2927862 . ПМИД 20576937 .
- ^ Такэсима, Х.; Комазаки, С.; Ниши, М.; Иино, М.; Кангава, К. (июль 2000 г.). «Юнктофилины: новое семейство белков соединительного мембранного комплекса» . Молекулярная клетка . 6 (1): 11–22. дои : 10.1016/s1097-2765(00)00003-4 . ISSN 1097-2765 . ПМИД 10949023 .
- ^ Яо, Лей; Се, Дуаньян; Гэн, Ли; Ши, Дэн; Хуан, Цзянь; Ву, Юфэй; Льв, Фей; Лян, Дандан; Ли, Ли; Лю, Йи; Ли, Цзюнь (3 февраля 2018 г.). «REEP5 (рецепторный вспомогательный белок 5) действует как формирователь мембран саркоплазматического ретикулума, модулируя сердечную функцию» . Журнал Американской кардиологической ассоциации . 7 (3). дои : 10.1161/JAHA.117.007205 . ISSN 2047-9980 . ПМЦ 5850239 . ПМИД 29431104 .
- ^ Ченг Х., Ледерер В.Дж., Каннелл М.Б. (октябрь 1993 г.). «Кальциевые искры: элементарные события, лежащие в основе связи возбуждения-сокращения в сердечной мышце». Наука . 262 (5134): 740–4. Бибкод : 1993Sci...262..740C . дои : 10.1126/science.8235594 . ПМИД 8235594 .
- ^ Каннелл М.Б., Ченг Х., Ледерер В.Дж. (ноябрь 1994 г.). «Пространственные неоднородности в Ca 2+
i во время сопряжения возбуждения-сокращения в кардиомиоцитах» . Biophys. J. 67 ( 5): 1942–56. Bibcode : 1994BpJ....67.1942C . doi : 10.1016/S0006-3495(94)80677-0 . PMC 1225569 ПМИД 7858131 . - ^ Берс, доктор медицинских наук (2001). Связь возбуждения-сокращения и сократительная сила сердца (2-е изд.). Дордрехт: Kluwer Academic Publishers. ISBN 9780792371571 . OCLC 47659382 .
- ^ Креспо Л.М., Грэнтэм С.Дж., Каннелл М.Б. (июнь 1990 г.). «Кинетика, стехиометрия и роль механизма обмена Na-Ca в изолированных кардиомиоцитах». Природа . 345 (6276): 618–21. Бибкод : 1990Natur.345..618C . дои : 10.1038/345618a0 . ПМИД 2348872 . S2CID 4348240 .
- ^ Jump up to: а б Хиллис, Дэвид М.; Садава, Дэвид Э.; Прайс, Мэри В. (2014). «Мышцы и движение». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 681–698. ISBN 978-1-464-10947-8 .
- ^ Jump up to: а б Гарднер, ЧР (1976). «Нейронный контроль передвижения дождевого червя». Биологические обзоры Кембриджского философского общества . 51 (1): 25–52. дои : 10.1111/j.1469-185X.1976.tb01119.x . ПМИД 766843 . S2CID 9983649 .
- ^ Jump up to: а б с Александр, Р. Макнил (2003). «Мышцы, мотор». Принципы передвижения животных (2-е изд.). Принстон, Нью-Джерси: Издательство Принстонского университета. стр. 15–37. ISBN 978-0-691-12634-0 .
- ^ Джозефсон, РК; Маламуд, Дж.Г.; Стоукс, ДР (15 сентября 2000 г.). «Асинхронная мышца: букварь» . Журнал экспериментальной биологии . 203 (18): 2713–2722. дои : 10.1242/jeb.203.18.2713 . ISSN 0022-0949 . ПМИД 10952872 .
- ^ Уэллс, Дэвид Эймс (1859). «Как было открыто гальваническое электричество» . Наука об обычных вещах: знакомое объяснение основных принципов физической науки . Нью-Йорк: Айвисон и Финни. п. 290.
- ^ Уиттакер, ET (1951), История теорий эфира и электричества. Том 1 , Нельсон, Лондон
- ^ Сандов, А (1952). «Связь возбуждения-сокращения в мышечной реакции» . Йельский университет биологической медицины . 25 (3): 176–201. ПМК 2599245 . ПМИД 13015950 .
- ^ Хаксли, HE (апрель 2000 г.). «Прошлые, настоящие и будущие эксперименты на мышцах» . Философские труды: Биологические науки . 355 (1396): 539–543. дои : 10.1098/rstb.2000.0595 . JSTOR 3066716 . ПМЦ 1692762 . ПМИД 10836507 .
Дальнейшее чтение
[ редактировать ]- Кранс, Дж. Л. (2010) Теория мышечного сокращения скользящих нитей. Природное образование 3(9):66
- Саладин, Кеннет С., Стивен Дж. Салливан и Кристина А. Ган. (2015). Анатомия и физиология: единство формы и функции. 7-е изд. Нью-Йорк: Образование Макгроу-Хилл.